
Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Substances
2.2. Animals
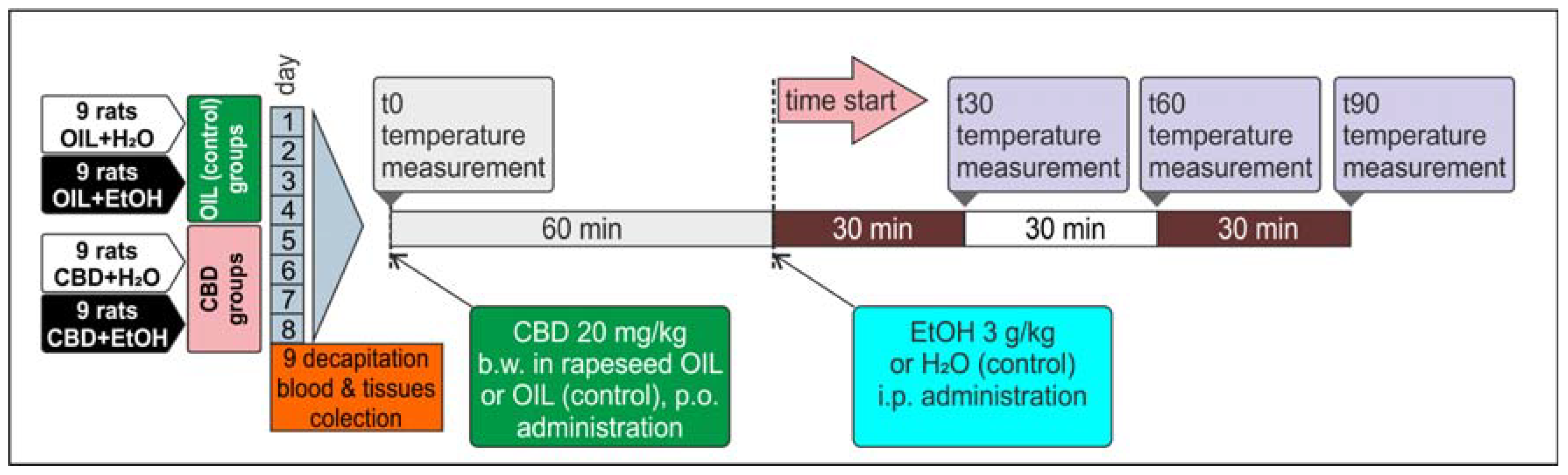
2.3. Experiment Protocol
2.4. Temperature Measurement
2.5. Loss of the Righting Reflex and Sleep Duration
2.6. Measurement of Ethanol Blood Concentration
2.7. Evaluation of RNA Isolation and mRNA Level Changes
2.8. Statistical Analysis
3. Results
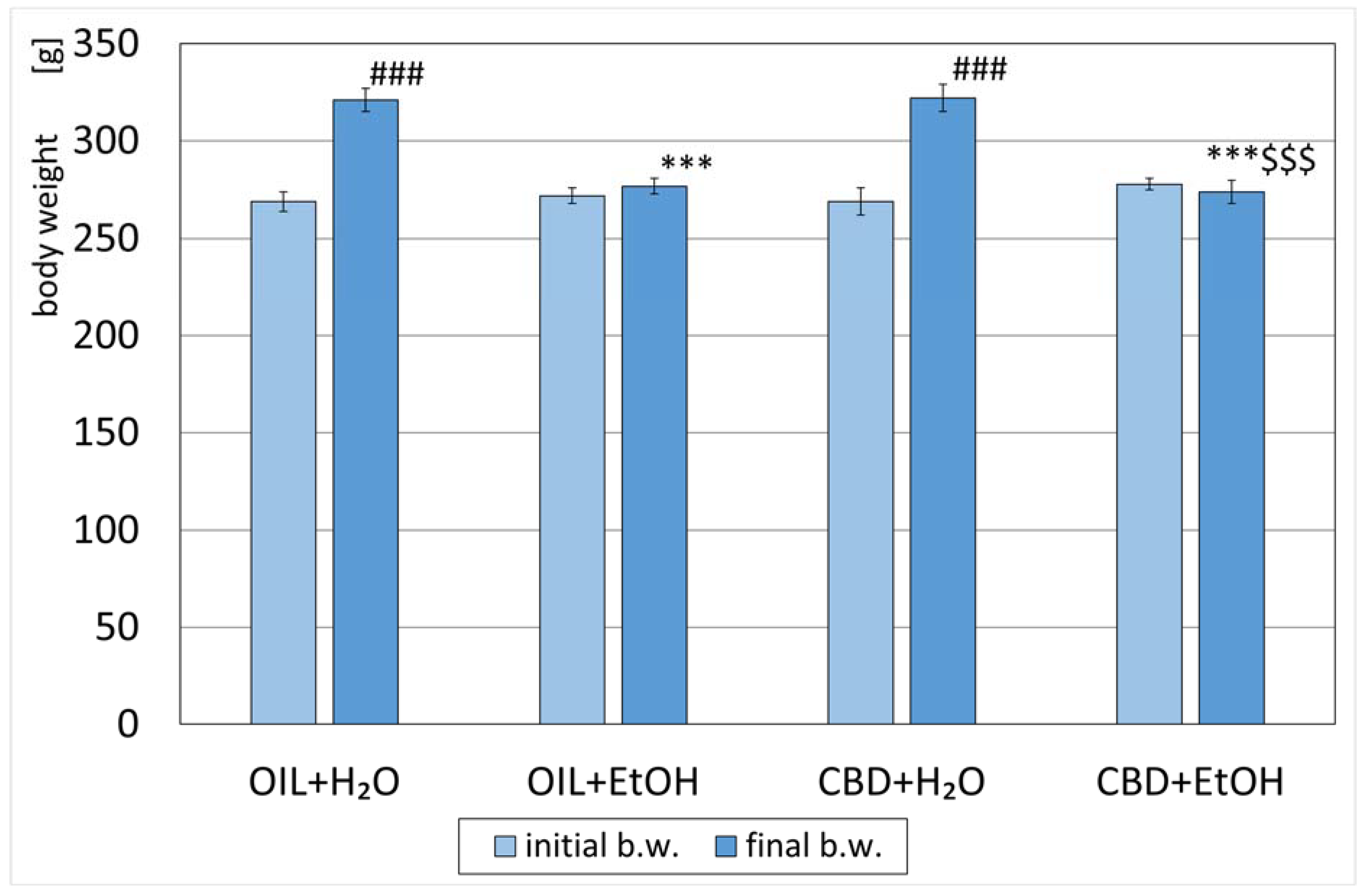
3.1. Animals’ Body Weight
3.2. Animals Body Temperature
3.2.1. T0 min
3.2.2. T30 min
3.2.3. T60 min
3.2.4. T90 min
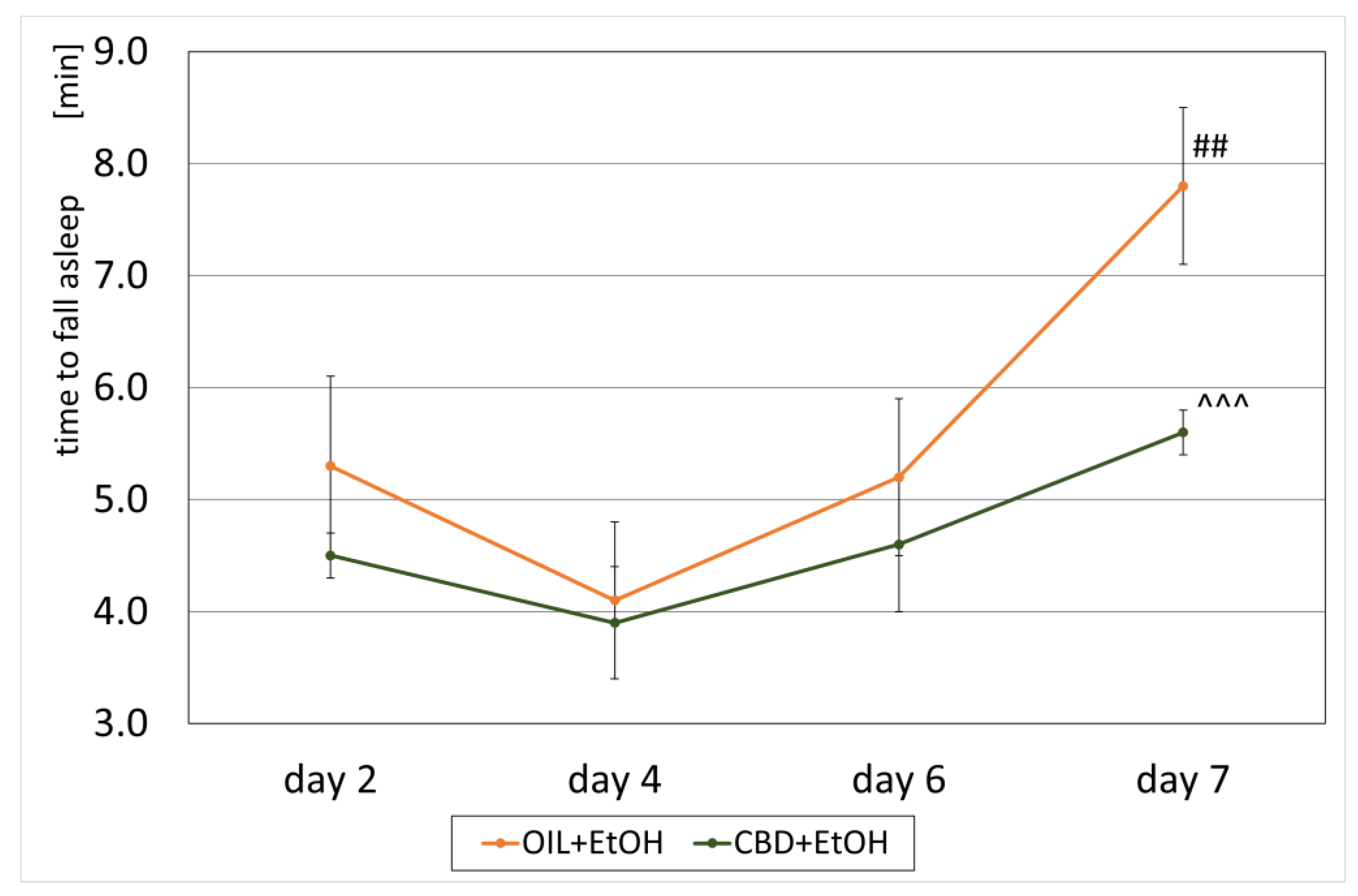
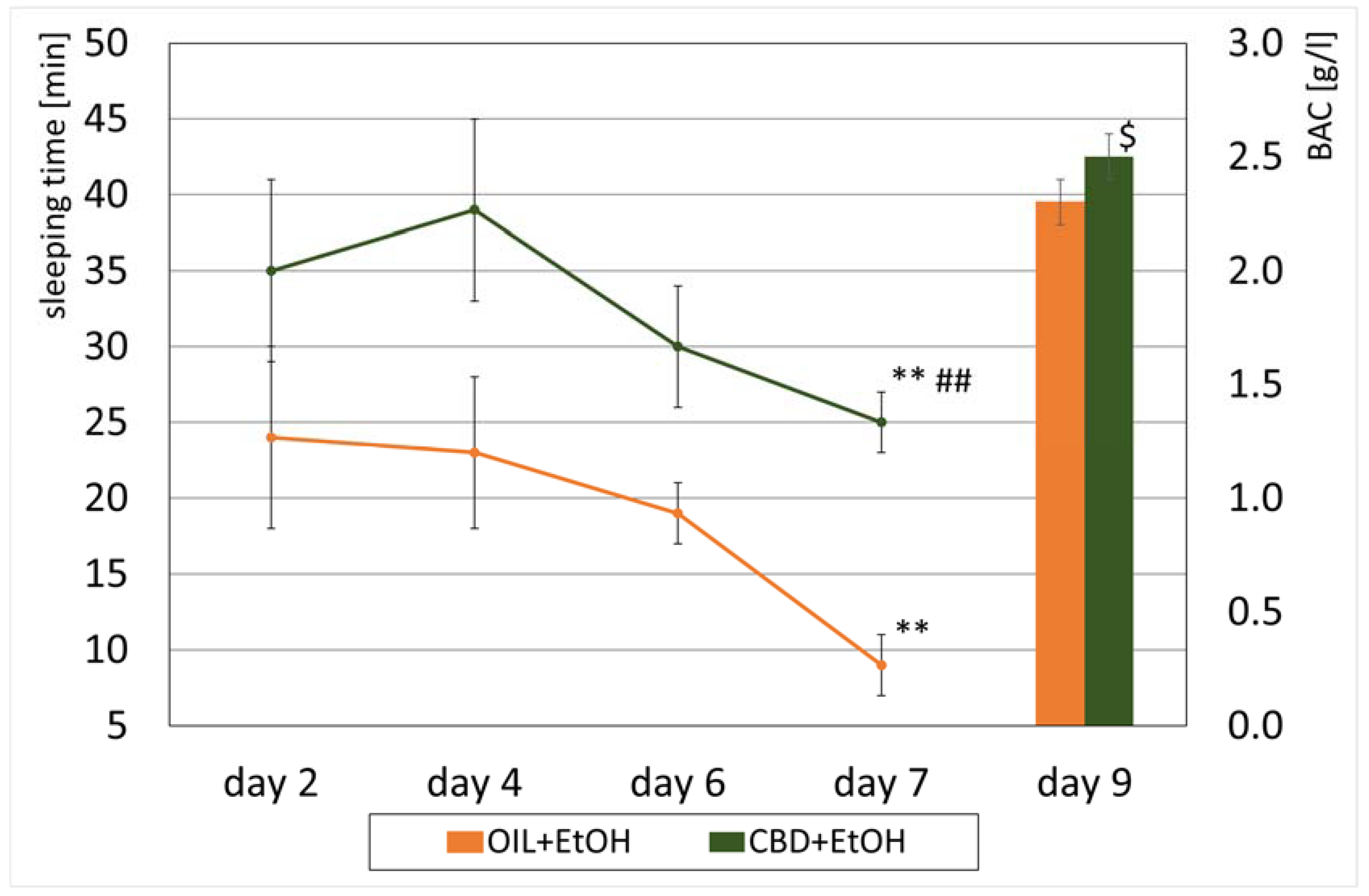
3.3. Loss of the Righting Reflex and Sleep Duration
3.4. Peripheral Blood Alcohol Concentration (BAC)
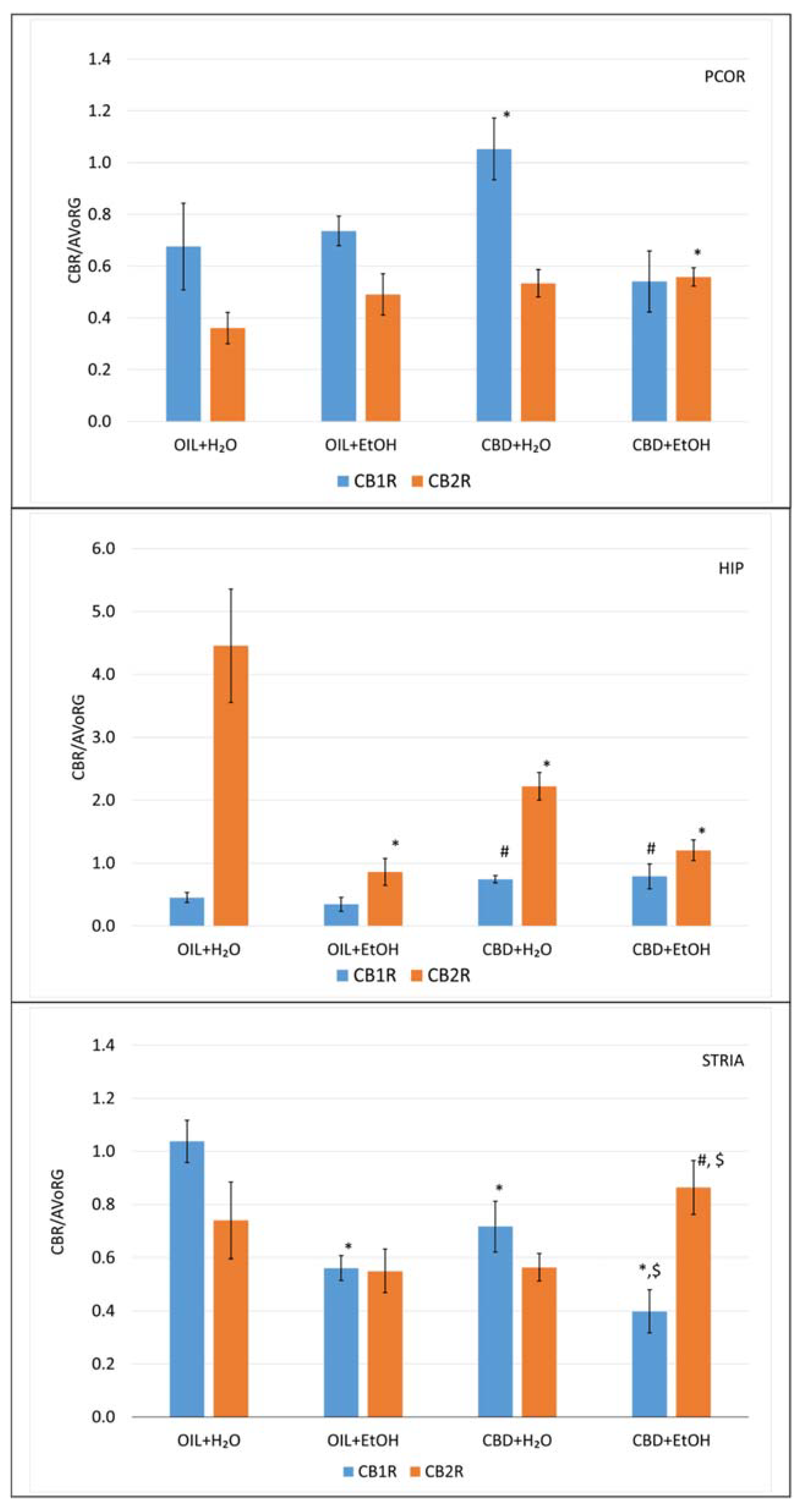
3.5. Evaluation of mRNA Level for Genes Encoding Cannabinoid Receptors in Selected Brain Structures
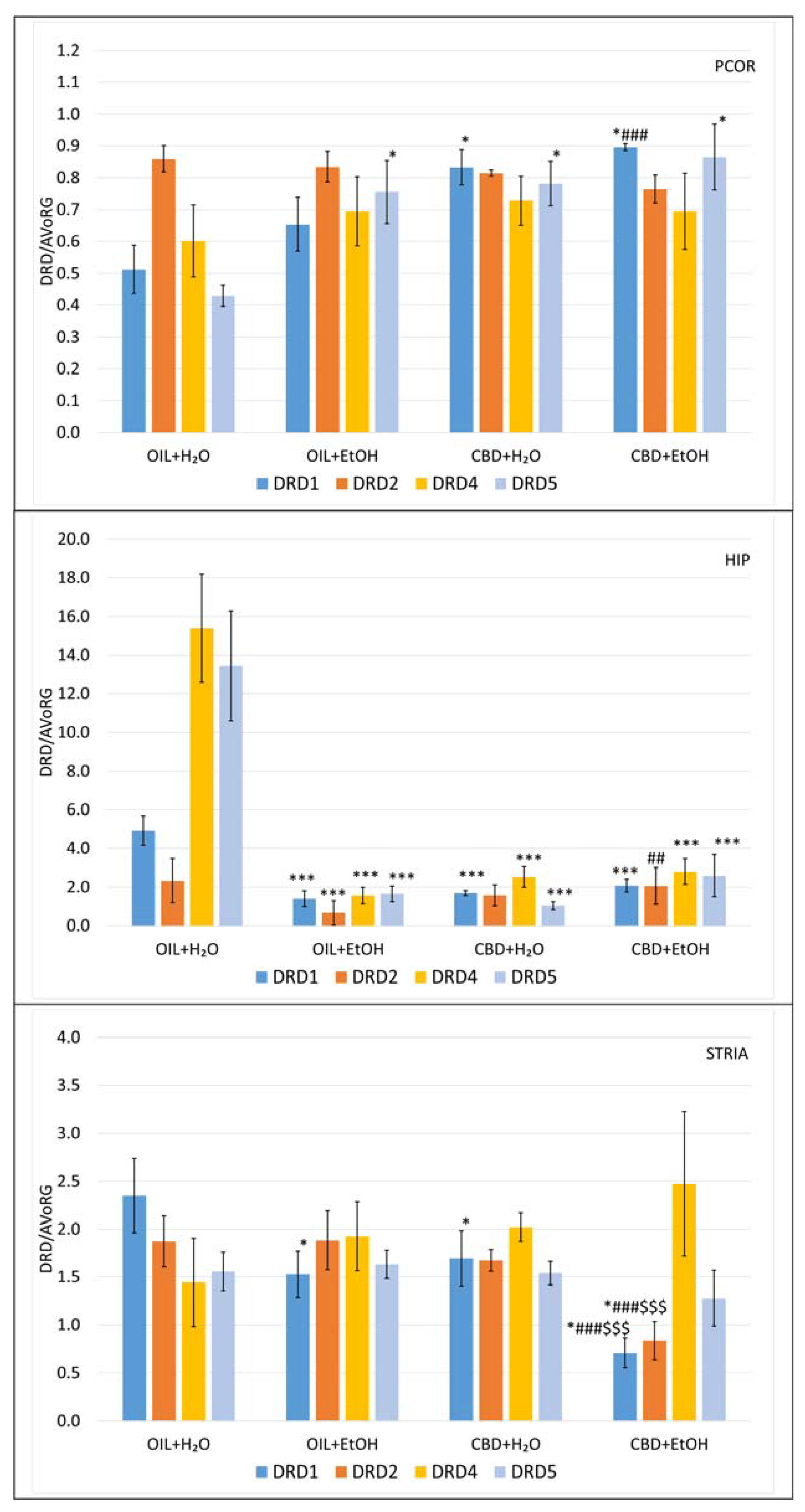
3.6. Evaluation of mRNA Level for Genes Encoding Dopamine Receptors in Selected Brain Structures
4. Discussion
4.1. Behavioral Studies
4.2. Molecular Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ray, L.A.; Bujarski, S.; Grodin, E.; Hartwell, E.; Green, R.; Venegas, A.; Lim, A.C.; Gillis, A.; Miotto, K. State-of-the-art behavioral and pharmacological treatments for alcohol use disorder. Am. J. Drug Alcohol Abus. 2019, 45, 124–140. [Google Scholar] [CrossRef]
- Tabakoff, B.; Kiianmaa, K. Does tolerance develop to the activating, as well as the depressant, effects of ethanol? Pharmacol. Biochem. Behav. 1982, 17, 1073–1076. [Google Scholar] [CrossRef]
- Morris, S.A.; Kelso, M.L.; Liput, D.J.; Marshall, S.A.; Nixon, K. Similar withdrawal severity in adolescents and adults in a rat model of alcohol dependence. Alcohol 2010, 44, 89–98. [Google Scholar] [CrossRef]
- Szulc, M.; Kujawski, R.; Baraniak, J.; Kania-Dobrowolska, M.; Kamińska, E.; Gryszczyńska, A.; Czora-Poczwardowska, K.; Winiarska, H.; Mikołajczak, P.Ł. Differential Influence of Pueraria lobata Root Extract and Its Main Isoflavones on Ghrelin Levels in Alcohol-Treated Rats. Pharmaceuticals 2021, 15, 25. [Google Scholar] [CrossRef] [PubMed]
- Szulc, M.; Mikołajczak, P.; Okulicz-Kozaryn, I.; Kamińska, E.; Dyr, W.; Kostowski, W. Acamprosate effect on short-term memory in chronically EtOH treated rats. Pol. J. Pharmacol. 2002, 54, 539–540. [Google Scholar]
- Haass-Koffler, C.L.; Perciballi, R. Alcohol Tolerance in Human Laboratory Studies for Development of Medications to treat Alcohol Use Disorder. Alcohol Alcohol. 2020, 55, 129–135. [Google Scholar] [CrossRef]
- Rustay, N.R.; Crabbe, J.C. Genetic analysis of rapid tolerance to ethanol’s incoordinating effects in mice: Inbred strains and artificial selection. Behav. Genet. 2004, 34, 441–451. [Google Scholar] [CrossRef] [PubMed]
- Ozburn, A.R.; Harris, R.A.; Blednov, Y.A. Chronic voluntary alcohol consumption results in tolerance to sedative/hypnotic and hypothermic effects of alcohol in hybrid mice. Pharmacol. Biochem. Behav. 2013, 104, 33–39. [Google Scholar] [CrossRef]
- Spiers, D.E. Thermoregulation and Alcohol. In Alcohol and Hormones; Watson, R.R., Ed.; Humana Press: Totowa, NJ, USA, 1995; pp. 193–208. ISBN 978-1-4612-0243-1. [Google Scholar]
- Carreño, C.F.; Ferreira, V.M.; Morato, G.S. Ethanol-induced hypothermia in rats is antagonized by dexamethasone. Braz. J. Med. Biol. Res. 1997, 30, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Morena, M.; Roozendaal, B.; Trezza, V.; Ratano, P.; Peloso, A.; Hauer, D.; Atsak, P.; Trabace, L.; Cuomo, V.; McGaugh, J.L.; et al. Endogenous cannabinoid release within prefrontal-limbic pathways affects memory consolidation of emotional training. Proc. Natl. Acad. Sci. USA 2014, 111, 18333–18338. [Google Scholar] [CrossRef] [PubMed]
- Lovinger, D.M.; Alvarez, V.A. Alcohol and basal ganglia circuitry: Animal models. Neuropharmacology 2017, 122, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Diez-Alarcia, R.; Ibarra-Lecue, I.; Lopez-Cardona, Á.P.; Meana, J.; Gutierrez-Adán, A.; Callado, L.F.; Agirregoitia, E.; Urigüen, L. Biased agonism of three different cannabinoid receptor agonists in mouse brain cortex. Front. Pharmacol. 2016, 7, 415. [Google Scholar] [CrossRef] [PubMed]
- Normandin, M.D.; Zheng, M.-Q.; Lin, K.-S.; Mason, N.S.; Lin, S.-F.; Ropchan, J.; Labaree, D.; Henry, S.; Williams, W.A.; Carson, R.E.; et al. Imaging the cannabinoid CB1 receptor in humans with [11C]OMAR: Assessment of kinetic analysis methods, test-retest reproducibility, and gender differences. J. Cereb. Blood Flow Metab. 2015, 35, 1313–1322. [Google Scholar] [CrossRef]
- Basavarajappa, B.S.; Hungund, B.L. Chronic ethanol increases the cannabinoid receptor agonist anandamide and its precursor N-arachidonoylphosphatidylethanolamine in SK-N-SH cells. J. Neurochem. 1999, 72, 522–528. [Google Scholar] [CrossRef] [PubMed]
- González, S.; Cascio, M.G.; Fernández-Ruiz, J.; Fezza, F.; Di Marzo, V.; Ramos, J.A. Changes in endocannabinoid contents in the brain of rats chronically exposed to nicotine, ethanol or cocaine. Brain Res. 2002, 954, 73–81. [Google Scholar] [CrossRef]
- Zou, S.; Kumar, U. Cannabinoid receptors and the endocannabinoid system: Signaling and function in the central nervous system. Int. J. Mol. Sci. 2018, 19, 833. [Google Scholar] [CrossRef] [PubMed]
- Basavarajappa, B.S.; Nagre, N.N.; Xie, S.; Subbanna, S. Elevation of endogenous anandamide impairs LTP, learning, and memory through CB1 receptor signaling in mice. Hippocampus 2014, 24, 808–818. [Google Scholar] [CrossRef]
- Dyr, W.; Ligieza, J.; Kostowski, W. The effect of cannabinoid CB(1) receptor antagonist rimonabant (SR-141716) on ethanol drinking in high-preferring rats. Alcohol 2008, 42, 509–512. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Jaimes, L.; Polis, I.; Parsons, L.H. Regional Influence of Cannabinoid CB1 Receptors in the Regulation of Ethanol Self-Administration by Wistar Rats. Open Neuropsychopharmacol. J. 2009, 2, 77–85. [Google Scholar] [CrossRef]
- Colombo, G.; Serra, S.; Brunetti, G.; Atzori, G.; Pani, M.; Vacca, G.; Addolorato, G.; Froestl, W.; Carai, M.A.M.; Gessa, G.L. The GABA(B) receptor agonists baclofen and CGP 44532 prevent acquisition of alcohol drinking behaviour in alcohol-preferring rats. Alcohol Alcohol. 2002, 37, 499–503. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Hirvonen, J.; Zanotti-Fregonara, P.; Umhau, J.C.; George, D.T.; Rallis-Frutos, D.; Lyoo, C.H.; Li, C.T.; Hines, C.S.; Sun, H.; Terry, G.E.; et al. Reduced cannabinoid CB1 receptor binding in alcohol dependence measured with positron emission tomography. Mol. Psychiatry 2013, 18, 916–921. [Google Scholar] [CrossRef]
- Pertwee, R.G.; Ross, R.A. Cannabinoid receptors and their ligands. Prostaglandins Leukot. Essent. Fat. Acids 2002, 66, 101–121. [Google Scholar] [CrossRef]
- Buckley, N.E.; McCoy, K.L.; Mezey, E.; Bonner, T.; Zimmer, A.; Felder, C.C.; Glass, M.; Zimmer, A. Immunomodulation by cannabinoids is absent in mice deficient for the cannabinoid CB(2) receptor. Eur. J. Pharmacol. 2000, 396, 141–149. [Google Scholar] [CrossRef]
- Onaivi, E.S.; Ishiguro, H.; Gong, J.-P.; Patel, S.; Perchuk, A.; Meozzi, P.A.; Myers, L.; Mora, Z.; Tagliaferro, P.; Gardner, E.; et al. Discovery of the presence and functional expression of cannabinoid CB2 receptors in brain. Ann. N. Y. Acad. Sci. 2006, 1074, 514–536. [Google Scholar] [CrossRef]
- Gong, J.-P.; Onaivi, E.S.; Ishiguro, H.; Liu, Q.-R.; Tagliaferro, P.A.; Brusco, A.; Uhl, G.R. Cannabinoid CB2 receptors: Immunohistochemical localization in rat brain. Brain Res. 2006, 1071, 10–23. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.K.; Devi, L.A. The highs and lows of cannabinoid receptor expression in disease: Mechanisms and their therapeutic implications. Pharmacol. Rev. 2011, 63, 461–470. [Google Scholar] [CrossRef]
- Ma, Z.; Gao, F.; Larsen, B.; Gao, M.; Luo, Z.; Chen, D.; Ma, X.; Qiu, S.; Zhou, Y.; Xie, J.; et al. Mechanisms of cannabinoid CB2 receptor-mediated reduction of dopamine neuronal excitability in mouse ventral tegmental area. EBioMedicine 2019, 42, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Powers, M.S.; Breit, K.R.; Chester, J.A. Genetic Versus Pharmacological Assessment of the Role of Cannabinoid Type 2 Receptors in Alcohol Reward-Related Behaviors. Alcohol. Clin. Exp. Res. 2015, 39, 2438–2446. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.S.; Navarrete, F.; Gasparyan, A.; Navarro, D.; Morcuende, Á.; Femenía, T.; Manzanares, J. Role of cannabinoid CB2 receptor in alcohol use disorders: From animal to human studies. Int. J. Mol. Sci. 2022, 23, 5908. [Google Scholar] [CrossRef] [PubMed]
- Pellati, F.; Borgonetti, V.; Brighenti, V.; Biagi, M.; Benvenuti, S.; Corsi, L. Cannabis sativa L. and Nonpsychoactive Cannabinoids: Their Chemistry and Role against Oxidative Stress, Inflammation, and Cancer. Biomed. Res. Int. 2018, 2018, 1691428. [Google Scholar] [CrossRef]
- Guerrero-Alba, R.; Barragán-Iglesias, P.; González-Hernández, A.; Valdez-Moráles, E.E.; Granados-Soto, V.; Condés-Lara, M.; Rodríguez, M.G.; Marichal-Cancino, B.A. Some prospective alternatives for treating pain: The endocannabinoid system and its putative receptors GPR18 and GPR. Front. Pharmacol. 2018, 9, 1496. [Google Scholar] [CrossRef]
- Elsaid, S.; Kloiber, S.; Le Foll, B. Effects of cannabidiol (CBD) in neuropsychiatric disorders: A review of pre-clinical and clinical findings. Prog. Mol. Biol. Transl. Sci. 2019, 167, 25–75. [Google Scholar] [CrossRef]
- Maccioni, P.; Bratzu, J.; Carai, M.A.M.; Colombo, G.; Gessa, G.L. Reducing Effect of Cannabidiol on Alcohol Self-Administration in Sardinian Alcohol-Preferring Rats. Cannabis Cannabinoid Res. 2022, 7, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, F.; García-Gutiérrez, M.S.; Gasparyan, A.; Austrich-Olivares, A.; Manzanares, J. Role of cannabidiol in the therapeutic intervention for substance use disorders. Front. Pharmacol. 2021, 12, 626010. [Google Scholar] [CrossRef] [PubMed]
- Kunos, G. Interactions between alcohol and the endocannabinoid system. Alcohol. Clin. Exp. Res. 2020, 44, 790–805. [Google Scholar] [CrossRef]
- Bustamante, D.; Quintanilla, M.E.; Tampier, L.; Gonzalez-Lira, V.; Israel, Y.; Herrera-Marschitz, M. Ethanol induces stronger dopamine release in nucleus accumbens (shell) of alcohol-preferring (bibulous) than in alcohol-avoiding (abstainer) rats. Eur. J. Pharmacol. 2008, 591, 153–158. [Google Scholar] [CrossRef]
- Wise, R.A.; Robble, M.A. Dopamine and Addiction. Annu. Rev. Psychol. 2020, 71, 79–106. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Jaimes, L.; Stouffer, D.G.; Parsons, L.H. Chronic ethanol treatment potentiates ethanol-induced increases in interstitial nucleus accumbens endocannabinoid levels in rats. J. Neurochem. 2009, 111, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Dahchour, A.; Ward, R.J. Changes in Brain Dopamine Extracellular Concentration after Ethanol Administration; Rat Microdialysis Studies. Alcohol Alcohol. 2022, 57, 165–175. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Palomo-Garo, C.; Gómez-Gálvez, Y.; Fernández-Ruiz, J. Cannabinoid-dopamine interactions in the physiology and physiopathology of the basal ganglia. Br. J. Pharmacol. 2016, 173, 2069–2079. [Google Scholar] [CrossRef]
- Hungund, B.L.; Szakall, I.; Adam, A.; Basavarajappa, B.S.; Vadasz, C. Cannabinoid CB1 receptor knockout mice exhibit markedly reduced voluntary alcohol consumption and lack alcohol-induced dopamine release in the nucleus accumbens. J. Neurochem. 2003, 84, 698–704. [Google Scholar] [CrossRef]
- Hill, S.Y.; Hoffman, E.K.; Zezza, N.; Thalamuthu, A.; Weeks, D.E.; Matthews, A.G.; Mukhopadhyay, I. Dopaminergic mutations: Within-family association and linkage in multiplex alcohol dependence families. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2008, 147B, 517–526. [Google Scholar] [CrossRef]
- Kalivas, P.W.; Stewart, J. Dopamine transmission in the initiation and expression of drug- and stress-induced sensitization of motor activity. Brain Res. Brain Res. Rev. 1991, 16, 223–244. [Google Scholar] [CrossRef]
- Ng, G.Y.; O’Dowd, B.F.; George, S.R. Genotypic differences in brain dopamine receptor function in the DBA/2J and C57BL/6J inbred mouse strains. Eur. J. Pharmacol. 1994, 269, 349–364. [Google Scholar] [CrossRef]
- McBride, W.J.; Chernet, E.; Russell, R.N.; Wong, D.T.; Guan, X.M.; Lumeng, L.; Li, T.K. Regional CNS densities of monoamine receptors in alcohol-naive alcohol-preferring P and -nonpreferring NP rats. Alcohol 1997, 14, 141–148. [Google Scholar] [CrossRef] [PubMed]
- McBride, W.J.; Chernet, E.; Russell, R.N.; Chamberlain, J.K.; Lumeng, L.; Li, T.K. Regional CNS densities of serotonin and dopamine receptors in high alcohol-drinking (HAD) and low alcohol-drinking (LAD) rats. Alcohol 1997, 14, 603–609. [Google Scholar] [CrossRef] [PubMed]
- Syvälahti, E.K.; Pohjalainen, T.; Korpi, E.R.; Pälvimäki, E.P.; Ovaska, T.; Kuoppamäki, M.; Hietala, J. Dopamine D2 receptor gene expression in rat lines selected for differences in voluntary alcohol consumption. Alcohol. Clin. Exp. Res. 1994, 18, 1029–1031. [Google Scholar] [CrossRef]
- Tupala, E.; Hall, H.; Mantere, T.; Räsänen, P.; Särkioja, T.; Tiihonen, J. Dopamine receptors and transporters in the brain reward circuits of type 1 and 2 alcoholics measured with human whole hemisphere autoradiography. Neuroimage 2003, 19, 145–155. [Google Scholar] [CrossRef]
- Dyr, W.; McBride, W.J.; Lumeng, L.; Li, T.K.; Murphy, J.M. Effects of D1 and D2 dopamine receptor agents on ethanol consumption in the high-alcohol-drinking (HAD) line of rats. Alcohol 1993, 10, 207–212. [Google Scholar] [CrossRef]
- Silvestre, J.S.; O’Neill, M.F.; Fernandez, A.G.; Palacios, J.M. Effects of a range of dopamine receptor agonists and antagonists on ethanol intake in the rat. Eur. J. Pharmacol. 1996, 318, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Sunahara, R.K.; Guan, H.C.; O’Dowd, B.F.; Seeman, P.; Laurier, L.G.; Ng, G.; George, S.R.; Torchia, J.; Van Tol, H.H.; Niznik, H.B. Cloning of the gene for a human dopamine D5 receptor with higher affinity for dopamine than D. Nature 1991, 350, 614–619. [Google Scholar] [CrossRef]
- Shuto, T.; Nishi, A. Treatment of the psychostimulant-sensitized animal model of schizophrenia. CNS Neurosci. Ther. 2011, 17, 133–139. [Google Scholar] [CrossRef] [PubMed]
- McBride, W.J.; Chernet, E.; Dyr, W.; Lumeng, L.; Li, T.K. Densities of dopamine D2 receptors are reduced in CNS regions of alcohol-preferring P rats. Alcohol 1993, 10, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Hietala, J.; West, C.; Syvälahti, E.; Någren, K.; Lehikoinen, P.; Sonninen, P.; Ruotsalainen, U. Striatal D2 dopamine receptor binding characteristics in vivo in patients with alcohol dependence. Psychopharmacology 1994, 116, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Logan, J.; Hitzemann, R.; Ding, Y.S.; Pappas, N.; Shea, C.; Piscani, K. Decreases in dopamine receptors but not in dopamine transporters in alcoholics. Alcohol. Clin. Exp. Res. 1996, 20, 1594–1598. [Google Scholar] [CrossRef]
- Heidbreder, C.A.; Andreoli, M.; Marcon, C.; Hutcheson, D.M.; Gardner, E.L.; Ashby, C.R. Evidence for the role of dopamine D3 receptors in oral operant alcohol self-administration and reinstatement of alcohol-seeking behavior in mice. Addict. Biol. 2007, 12, 35–50. [Google Scholar] [CrossRef]
- Boyce-Rustay, J.M.; Risinger, F.O. Dopamine D3 receptor knockout mice and the motivational effects of ethanol. Pharmacol. Biochem. Behav. 2003, 75, 373–379. [Google Scholar] [CrossRef]
- McQuade, J.-A.; Xu, M.; Woods, S.C.; Seeley, R.J.; Benoit, S.C. Ethanol consumption in mice with a targeted disruption of the dopamine-3 receptor gene. Addict. Biol. 2003, 8, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Thrasher, M.J.; Freeman, P.A.; Risinger, F.O. Clozapine’s effects on ethanol’s motivational properties. Alcohol. Clin. Exp. Res. 1999, 23, 1377–1385. [Google Scholar] [CrossRef]
- Rubinstein, M.; Phillips, T.J.; Bunzow, J.R.; Falzone, T.L.; Dziewczapolski, G.; Zhang, G.; Fang, Y.; Larson, J.L.; McDougall, J.A.; Chester, J.A.; et al. Mice lacking dopamine D4 receptors are supersensitive to ethanol, cocaine, and methamphetamine. Cell 1997, 90, 991–1001. [Google Scholar] [CrossRef][Green Version]
- Szulc, M.; Mularczyk, P.; Kujawski, R.; Gryszczyńska, A.; Kamińska, E.; Geppert, B.; Baraniak, J.; Kania-Dobrowolska, M.; Ożarowski, M.; Krajewska-Patan, A.; et al. Influence of salidroside, a neuroactive compound of Rhodiola rosea L., on alcohol tolerance development in rats. Herba Pol. 2018, 64, 22–35. [Google Scholar] [CrossRef]
- Szulc, M.; Mularczyk, P.; Grządzielski, P.; Zakowicz, P.; Kujawski, R.; Gryszczyńska, A.; Buchwald, W.; Teżyk, A.; Krajewska-Patan, A.; Kamińska, E. Others Influence of extracts from Rhodiola rosea and Rhodiola kirilowii on the development of alcohol tolerance in rats. Herba Pol. 2018, 64, 34–43. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Okulicz-Kozaryn, I.; Mikolajczak, P.; Kaminska, E. Tolerance to hypothermia and hypnotic action of ethanol in 3 and 14 months old rats. Pharmacol. Res. 1992, 25, 63–64. [Google Scholar] [CrossRef]
- Crabbe, J.C.; Janowsky, J.S.; Young, E.R.; Kosobud, A.; Stack, J.; Rigter, H. Tolerance to ethanol hypothermia in inbred mice: Genotypic correlations with behavioral responses. Alcohol. Clin. Exp. Res. 1982, 6, 446–458. [Google Scholar] [CrossRef] [PubMed]
- Costa, B.; Trovato, A.E.; Comelli, F.; Giagnoni, G.; Colleoni, M. The non-psychoactive cannabis constituent cannabidiol is an orally effective therapeutic agent in rat chronic inflammatory and neuropathic pain. Eur. J. Pharmacol. 2007, 556, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Viudez-Martínez, A.; García-Gutiérrez, M.S.; Navarrón, C.M.; Morales-Calero, M.I.; Navarrete, F.; Torres-Suárez, A.I.; Manzanares, J. Cannabidiol reduces ethanol consumption, motivation and relapse in mice. Addict. Biol. 2018, 23, 154–164. [Google Scholar] [CrossRef] [PubMed]
- Jamontt, J.M.; Molleman, A.; Pertwee, R.G.; Parsons, M.E. The effects of Delta-tetrahydrocannabinol and cannabidiol alone and in combination on damage, inflammation and in vitro motility disturbances in rat colitis. Br. J. Pharmacol. 2010, 160, 712–723. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.B.; Watson, M.R.; James, K.; Kastner, A.; Schneider, A.; Mittleman, G. The impact of low to moderate chronic intermittent ethanol exposure on behavioral endpoints in aged, adult, and adolescent rats. Alcohol 2019, 78, 33–42. [Google Scholar] [CrossRef]
- Marty, V.N.; Farokhnia, M.; Munier, J.J.; Mulpuri, Y.; Leggio, L.; Spigelman, I. Long-Acting Glucagon-Like Peptide-1 Receptor Agonists Suppress Voluntary Alcohol Intake in Male Wistar Rats. Front. Neurosci. 2020, 14, 599646. [Google Scholar] [CrossRef] [PubMed]
- Gautier, H. Body temperature regulation in the rat. J. Therm. Biol. 2000, 25, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Hampson, A.J.; Grimaldi, M.; Lolic, M.; Wink, D.; Rosenthal, R.; Axelrod, J. Neuroprotective antioxidants from marijuana. Ann. N. Y. Acad. Sci. 2000, 899, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Hampson, S.E.; Andrews, J.A.; Barckley, M.; Severson, H.H. Personality predictors of the development of elementary school children’s intentions to drink alcohol: The mediating effects of attitudes and subjective norms. Psychol. Addict. Behav. 2006, 20, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Bischof, L.L. Stress and ethanol-induced hypothermia. Physiol. Behav. 1987, 40, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Stout, S.M.; Cimino, N.M. Exogenous cannabinoids as substrates, inhibitors, and inducers of human drug metabolizing enzymes: A systematic review. Drug Metab. Rev. 2014, 46, 86–95. [Google Scholar] [CrossRef]
- Crabbe, J.C. Use of animal models of alcohol-related behavior. Handb. Clin. Neurol. 2014, 125, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Treistman, S.N.; Martin, G.E. BK Channels: Mediators and models for alcohol tolerance. Trends Neurosci. 2009, 32, 629–637. [Google Scholar] [CrossRef]
- Kyzar, E.J.; Pandey, S.C. Molecular mechanisms of synaptic remodeling in alcoholism. Neurosci. Lett. 2015, 601, 11–19. [Google Scholar] [CrossRef]
- Basavarajappa, B.S.; Joshi, V.; Shivakumar, M.; Subbanna, S. Distinct functions of endogenous cannabinoid system in alcohol abuse disorders. Br. J. Pharmacol. 2019, 176, 3085–3109. [Google Scholar] [CrossRef]
- Sanchez-Marin, L.; Pavon, F.J.; Decara, J.; Suarez, J.; Gavito, A.; Castilla-Ortega, E.; Rodriguez de Fonseca, F.; Serrano, A. Effects of intermittent alcohol exposure on emotion and cognition: A potential role for the endogenous cannabinoid system and neuroinflammation. Front. Behav. Neurosci. 2017, 11, 15. [Google Scholar] [CrossRef]
- Pava, M.J.; Woodward, J.J. A review of the interactions between alcohol and the endocannabinoid system: Implications for alcohol dependence and future directions for research. Alcohol 2012, 46, 185–204. [Google Scholar] [CrossRef]
- Ferrer, B.; Bermúdez-Silva, F.J.; Bilbao, A.; Alvarez-Jaimes, L.; Sanchez-Vera, I.; Giuffrida, A.; Serrano, A.; Baixeras, E.; Khaturia, S.; Navarro, M.; et al. Regulation of brain anandamide by acute administration of ethanol. Biochem. J. 2007, 404, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Van Sickle, M.D.; Duncan, M.; Kingsley, P.J.; Mouihate, A.; Urbani, P.; Mackie, K.; Stella, N.; Makriyannis, A.; Piomelli, D.; Davison, J.S.; et al. Identification and functional characterization of brainstem cannabinoid CB2 receptors. Science 2005, 310, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Al Mansouri, S.; Ojha, S.; Al Maamari, E.; Al Ameri, M.; Nurulain, S.M.; Bahi, A. The cannabinoid receptor 2 agonist, β-caryophyllene, reduced voluntary alcohol intake and attenuated ethanol-induced place preference and sensitivity in mice. Pharmacol. Biochem. Behav. 2014, 124, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Marco, E.M.; Ballesta, J.A.; Irala, C.; Hernández, M.-D.; Serrano, M.E.; Mela, V.; López-Gallardo, M.; Viveros, M.-P. Sex-dependent influence of chronic mild stress (CMS) on voluntary alcohol consumption; study of neurobiological consequences. Pharmacol. Biochem. Behav. 2017, 152, 68–80. [Google Scholar] [CrossRef]
- Beaulieu, J.-M.; Borrelli, E.; Carlsson, A.; Caron, M.G.; Civelli, O.; Espinoza, S.; Fisone, G.; Gainetdinov, R.R.; Grandy, D.K.; Kebabian, J.W.; et al. Dopamine receptors (version 2019.4) in the IUPHAR/BPS Guide to Pharmacology Database. GtoPdb CITE 2019, 2019, 162–167. [Google Scholar] [CrossRef]
- Hill, R.A.; Xu, W.; Yoshimura, M. Role of an adenylyl cyclase isoform in ethanol’s effect on cAMP regulated gene expression in NIH 3T3 cells. Biochem. Biophys. Rep. 2016, 8, 162–167. [Google Scholar] [CrossRef][Green Version]
- Eravci, M.; Grosspietsch, T.; Pinna, G.; Schulz, O.; Kley, S.; Bachmann, M.; Wolffgramm, J.; Götz, E.; Heyne, A.; Meinhold, H.; et al. Dopamine receptor gene expression in an animal model of “behavioral dependence” on ethanol. Brain Res. Mol. Brain Res. 1997, 50, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Hungund, B.L.; Basavarajappa, B.S. Distinct differences in the cannabinoid receptor binding in the brain of C57BL/6 and DBA/2 mice, selected for their differences in voluntary ethanol consumption. J. Neurosci. Res. 2000, 60, 122–128. [Google Scholar] [CrossRef]
- Diana, M.; Melis, M.; Muntoni, A.L.; Gessa, G.L. Mesolimbic dopaminergic decline after cannabinoid withdrawal. Proc. Natl. Acad. Sci. USA 1998, 95, 10269–10273. [Google Scholar] [CrossRef] [PubMed]
- Balla, A.; Dong, B.; Shilpa, B.M.; Vemuri, K.; Makriyannis, A.; Pandey, S.C.; Sershen, H.; Suckow, R.F.; Vinod, K.Y. Cannabinoid-1 receptor neutral antagonist reduces binge-like alcohol consumption and alcohol-induced accumbal dopaminergic signaling. Neuropharmacology 2018, 131, 200–208. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, D.L.; Devi, L.A. Diversity of molecular targets and signaling pathways for CBD. Pharmacol. Res. Perspect. 2020, 8, e00682. [Google Scholar] [CrossRef] [PubMed]
- D’Souza, D.C.; Cortes-Briones, J.A.; Ranganathan, M.; Thurnauer, H.; Creatura, G.; Surti, T.; Planeta, B.; Neumeister, A.; Pittman, B.; Normandin, M.; et al. Rapid Changes in CB1 Receptor Availability in Cannabis Dependent Males after Abstinence from Cannabis. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2016, 1, 60–67. [Google Scholar] [CrossRef] [PubMed]
- ClinicalTrials.gov CBD for the Treatment of Alcohol Use Disorder. 2022. Available online: https://clinicaltrials.gov/ct2/show/NCT04873453 (accessed on 1 March 2023).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward (5′ → 3′) | Reverse (5′ → 3′) | |
|---|---|---|
| HPRT1 | TGGAGATTCAAGTCC | ATGAGGCTGTCTGTGATGTC |
| SDHA | CAGGAGTTGCCTTCCTTTGTG | GTAAATAAATGTCCTGTGAAG |
| CB1 | TAATATGAAGCAAGATACCAG | CCATTTACAGAGACAACAAG |
| CB2 | CAGTTACAGAGACAGAGGC | TGTTTCCATTACCCTAGAGC |
| DRD1 | TCCTTCAAGAGGGAGAGACGAA | CCACACAAACACATCGAAGG |
| DRD2 | TTGCAGACCACCACCAACTA | AATTTCCACTCACCCACCAC |
| DRD4 | GATGTGTTGGACGCCTTTCT | TCGGCATTGAAGATGGTGTA |
| DRD5 | CCACATGATACCGAATGCAG | CACAGTCAAGCTCCCAGACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szulc, M.; Kujawski, R.; Pacholak, A.; Poprawska, M.; Czora-Poczwardowska, K.; Geppert, B.; Mikołajczak, P.Ł. Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats. Nutrients 2023, 15, 1702. https://doi.org/10.3390/nu15071702
Szulc M, Kujawski R, Pacholak A, Poprawska M, Czora-Poczwardowska K, Geppert B, Mikołajczak PŁ. Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats. Nutrients. 2023; 15(7):1702. https://doi.org/10.3390/nu15071702
Chicago/Turabian StyleSzulc, Michał, Radosław Kujawski, Amanda Pacholak, Marta Poprawska, Kamila Czora-Poczwardowska, Bogna Geppert, and Przemysław Ł. Mikołajczak. 2023. "Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats" Nutrients 15, no. 7: 1702. https://doi.org/10.3390/nu15071702
APA StyleSzulc, M., Kujawski, R., Pacholak, A., Poprawska, M., Czora-Poczwardowska, K., Geppert, B., & Mikołajczak, P. Ł. (2023). Cannabidiol as a Modulator of the Development of Alcohol Tolerance in Rats. Nutrients, 15(7), 1702. https://doi.org/10.3390/nu15071702