
Maternal Intake of Polyunsaturated Fatty Acids in Autism Spectrum Etiology and Its Relation to the Gut Microbiota: What Do We Know?


 , , , , , ,
, , , , , ,
Abstract
1. Introduction
2. Methodology
3. Understanding Autism Spectrum Disorder: General Concepts
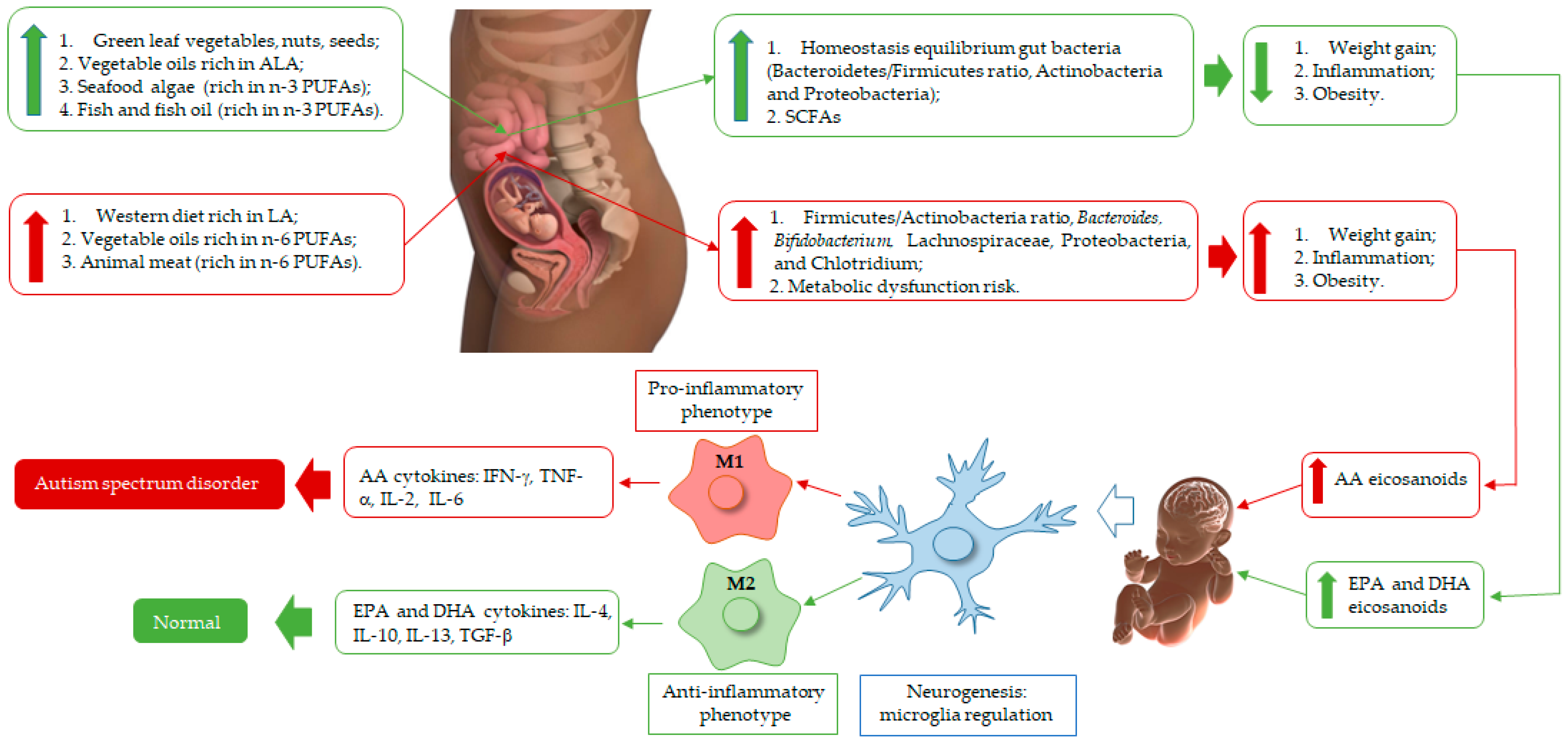
4. The Role of PUFA Intake during Pregnancy and Its Risk for the Development of Autistic Traits

{kind=link}
| Study Design/Kind of Study | Sample | Diet | Child Autistic Traits |
|---|---|---|---|
| 3802 mothers | Low intake or concentrations of ω-3 | No associations of individual n-3 PUFAs with child autistic traits |
| Higher total ω-6 levels (linoleic acid only) | More child autistic traits | ||
| 258 mother-child pairs | Higher consumption of total n-3 in second half of pregnancy | 40% lower risk of having children with Autism Spectrum Disorder |
| 92,011 BAP mothers | Pregnant women with higher BAP level presented lower consumption of vegetables and fish | A potential risk factor for children’s food choices, mental and physical development |
| 1892 and 1589 mother-child pairs at the ages of 14 months and 5 years, respectively | Consumption of seafood by pregnant women over 454 g/week | A consistent reduction in Autism Spectrum Disorder traits |
| 11,875 pregnant women | Maternal consumption of seafood over 340 g per week | Beneficial for the children’s neurodevelopment (considering verbal IQ and communication skills) |
5. Gut Microbiota of Pregnant Women: A Link with the Child’s Autism Spectrum Disorder?
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Rojas, D.C.; Singel, D.; Steinmetz, S.; Hepburn, S.; Brown, M.S. Decreased Left Perisylvian GABA Concentration in Children with Autism and Unaffected Siblings. NeuroImage 2014, 86, 28–34. [Google Scholar] [CrossRef] [PubMed]
- Kojovic, N.; Ben Hadid, L.; Franchini, M.; Schaer, M. Sensory Processing Issues and Their Association with Social Difficulties in Children with Autism Spectrum Disorders. J. Clin. Med. 2019, 8, 1508. [Google Scholar] [CrossRef] [PubMed]
- Soler, N.; Hardwick, C.; Perkes, I.E.; Mohammad, S.S.; Dossetor, D.; Nunn, K.; Bray, P.; Dale, R.C. Sensory Dysregulation in Tic Disorders Is Associated with Executive Dysfunction and Comorbidities. Mov. Disord 2019, 34, 1901–1909. [Google Scholar] [CrossRef] [PubMed]
- Yonkman, J.; Lawler, B.; Talty, J.; O’Neil, J.; Bull, M. Safely Transporting Children with Autism Spectrum Disorder: Evaluation and Intervention. Am. J. Occup. Ther. 2013, 67, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Afif, I.Y.; Farkhan, M.; Kurdi, O.; Maula, M.I.; Ammarullah, M.I.; Setiyana, B.; Jamari, J.; Winarni, T.I. Effect of short-term deep-pressure portable seat on behavioral and biological stress in children with autism spectrum disorders: A pilot study. Bioengineering 2022, 9, 48. [Google Scholar] [CrossRef]
- Kogan, M.D.; Vladutiu, C.J.; Schieve, L.A.; Ghandour, R.M.; Blumberg, S.J.; Zablotsky, B.; Perrin, J.M.; Shattuck, P.; Kuhlthau, K.A.; Harwood, R.L.; et al. The Prevalence of Parent-Reported Autism Spectrum Disorder Among US Children. American Academy of Pediatrics. Pediatrics 2018, 142, e20174161. Available online: https://pediatrics.aappublications.org/content/142/6/e20174161 (accessed on 11 April 2021). [CrossRef]
- Fombonne, E. Editorial: The Rising Prevalence of Autism. J. Child Psychol. Psychiatry 2018, 59, 717–720. Available online: https://acamh.onlinelibrary.wiley.com/doi/full/10.1111/jcpp.12941 (accessed on 11 April 2021). [CrossRef]
- Agrawal, S.; Rao, S.C.; Bulsara, M.K.; Patole, S.K. Prevalence of Autism Spectrum Disorder in Preterm Infants: A Meta-Analysis. Pediatrics 2018, 142, e20180134. [Google Scholar] [CrossRef]
- Sharp, M.; French, N.; McMichael, J.; Campbell, C. Survival and neurodevelopmental outcome in extremely preterm infants 22-24 weeks of gestation born in western Australia. J. Pediatr. Child. Health 2018, 54, 188–193. [Google Scholar] [CrossRef]
- Cheong, J.L.; Doyle, L.W.; Burnett, A.C.; Lee, K.J.; Walsh, J.M.; Potter, C.R.; Treyvaud, K.; Thompson, D.K.; Olsen, J.E.; Anderson, P.J.; et al. Association between moderate and late preterm birth and neurodevelopment and social-emotional development at age 2 years. JAMA Pediatr. 2017, 171, e164805. [Google Scholar] [CrossRef]
- Maramara, L.A.; He, W.; Ming, X. Pre-and perinatal risk factors for autism spectrum disorder in a New Jersey cohort. J. Child. Neurol. 2014, 29, 1645–1651. [Google Scholar] [CrossRef] [PubMed]
- Limperopoulos, C. Extreme prematurity, cerebellar injury, and autism. Semin. Pediatr. Neurol. 2010, 17, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Han, Y.; Dy, A.B.C.; Hagerman, R.J. The Gut Microbiota and Autism Spectrum Disorders. Front. Cell. Neurosci. 2017, 11, 120. [Google Scholar] [CrossRef] [PubMed]
- Lundgren, S.N.; Madan, J.C.; Emond, J.A.; Morrison, H.G.; Christensen, B.C.; Karagas, M.R.; Hoen, A.G. Maternal diet during pregnancy is related with the infant stool microbiome in a delivery mode-dependent manner. Microbiome 2018, 6, 109. [Google Scholar] [CrossRef] [PubMed]
- Galley, J.D.; Bailey, M.; Dush, C.K.; Schoppe-Sullivan, S.; Christian, L.M. Maternal Obesity Is Associated with Alterations in the Gut Microbiome in Toddlers. PLoS ONE 2014, 9, e113026. [Google Scholar] [CrossRef] [PubMed]
- Connolly, N.; Anixt, J.; Manning, P.; Lin, D.P.-I.; Marsolo, K.; Bowers, K. Maternal Metabolic Risk Factors for Autism Spectrum Disorder-An Analysis of Electronic Medical Records and Linked Birth Data. Autism Res. Off. J. Int. Soc. Autism Res. 2016, 9, 829–837. [Google Scholar] [CrossRef]
- Edlow, A.G. Maternal Obesity and Neurodevelopmental and Psychiatric Disorders in Offspring. Prenat. Diagn. 2017, 37, 95–110. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, C.E.; Barry, C.; Sabhlok, A.; Russell, K.; Majors, A.; Kollins, S.H.; Fuemmeler, B.F. Maternal Pre-Pregnancy Obesity and Child Neurodevelopmental Outcomes: A Meta-Analysis. Obes. Rev. 2018, 19, 464–484. [Google Scholar] [CrossRef]
- Madore, C.; Leyrolle, Q.; Lacabanne, C.; Benmamar-Badel, A.; Joffre, C.; Nadjar, A.; Layé, S. Neuroinflammation in Autism: Plausible Role of Maternal Inflammation, Dietary Omega 3, and Microbiota. Neural Plast. 2016, 2016, 3597209. [Google Scholar] [CrossRef]
- Lyall, K.; Munger, K.L.; O’Reilly, É.J.; Santangelo, S.L.; Ascherio, A. Maternal Dietary Fat Intake in Association with Autism Spectrum Disorders. Am. J. Epidemiol. 2013, 178, 209–220. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Schwartz, C.E. Intellectual Disability and Autism Spectrum Disorders: Causal Genes and Molecular Mechanisms. Neurosci. Biobehav. Rev. 2014, 46 Pt 2, 161–174. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.H.; Smith, T.; Paciorkowski, A.R. Autism Spectrum Disorder and Epilepsy: Disorders with a Shared Biology. Epilepsy Behav. 2015, 47, 191–201. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, M.; Piccolo, M.; Vannini, L.; Siragusa, S.; De Giacomo, A.; Serrazzanetti, D.I.; Cristofori, F.; Guerzoni, M.E.; Gobbetti, M.; Francavilla, R. Fecal Microbiota and Metabolome of Children with Autism and Pervasive Developmental Disorder Not Otherwise Specified. PLoS ONE 2013, 8, e76993. [Google Scholar] [CrossRef]
- Berding, K.; Donovan, S.M. Diet Can Impact Microbiota Composition in Children with Autism Spectrum Disorder. Front. Neurosci. 2018, 12, 515. [Google Scholar] [CrossRef] [PubMed]
- Esteban-Figuerola, P.; Canals, J.; Cao-Fernández, J.C.; Val, V.A. Differences in food consumption and nutritional intake between children with autism spectrum disorders and typically developing children: A meta-analysis. Autism 2019, 23, 1079–1095. [Google Scholar] [CrossRef] [PubMed]
- Chiarotti, F.; Venerosi, A. Epidemiology of autism spectrum disorders: A review of worldwide prevalence estimates since 2014. Brain Sci. 2020, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Fombonne, E. Epidemiological controversies in autism. Swiss Arch. Neurol. Psychiatry Psychother. 2020, 171, w03084. [Google Scholar] [CrossRef]
- Baio, J.; Wiggins, L.; Christensen, D.L.; Maenner, M.J.; Daniels, J.; Warren, Z.; Kurzius-Spencer, M.; Zahorodny, W.; Robinson Rosenberg, C.; White, T.; et al. Prevalence of autism spectrum disorder among children aged 8 years-autism and developmental disabilities monitoring network, 11 sites, United States, 2014. MMWR Surveill. Summ. 2018, 67, 1–23. [Google Scholar] [CrossRef]
- Xu, G.; Strathearn, L.; Liu, B.; Bao, W. Prevalence of autism spectrum disorder among US children and adolescents, 2014–2016. JAMA 2018, 319, 81–82. [Google Scholar] [CrossRef]
- Maenner, M.J. Prevalence of Autism Spectrum Disorder Among Children Aged 8 Years—Autism and Developmental Disabilities Monitoring Network, 11 Sites, United States, 2016. MMWR Surveill. Summ. 2020, 69, 1–12. [Google Scholar] [CrossRef]
- Narzisi, A.; Posada, M.; Barbieri, F.; Chericoni, N.; Ciuffolini, D.; Pinzino, M.; Romano, R.; Scattoni, M.L.; Tancredi, R.; Calderoni, S.; et al. Prevalence of Autism Spectrum Disorder in a large Italian catchment area: A school-based population study within the ASDEU project. Epidemiol. Psychiatr. Sci. 2020, 29, e5. [Google Scholar] [CrossRef] [PubMed]
- Fombonne, E. Epidemiology of Pervasive Developmental Disorders. Pediatr. Res. 2009, 65, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Elsabbagh, M.; Divan, G.; Koh, Y.-J.; Kim, Y.S.; Kauchali, S.; Marcín, C.; Montiel-Nava, C.; Patel, V.; Paula, C.S.; Wang, C.; et al. Global Prevalence of Autism and Other Pervasive Developmental Disorders. Autism Res. Off. J. Int. Soc. Autism Res. 2012, 5, 160–179. [Google Scholar] [CrossRef]
- Salari, N.; Rasoulpoor, S.; Rasoulpoor, S.; Shohaimi, S.; Jafarpour, S.; Abdoli, N.; Khaledi-Paveh, B.; Mohammadi, M. The global prevalence of autism spectrum disorder: A comprehensive systematic review and meta-analysis. Ital. J. Pediatr. 2022, 48, 112. [Google Scholar] [CrossRef] [PubMed]
- Hallmayer, J.; Cleveland, S.; Torres, A.; Phillips, J.; Cohen, B.; Torigoe, T.; Miller, J.; Fedele, A.; Collins, J.; Smith, K.; et al. Genetic Heritability and Shared Environmental Factors among Twin Pairs with Autism. Arch. Gen. Psychiatry 2011, 68, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, T.; Morisaki, N.; Honda, Y.; Sampei, M.; Tani, Y. Chemicals, Nutrition, and Autism Spectrum Disorder: A Mini-Review. Front. Neurosci. 2016, 10, 174. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.-C.; Lombardo, M.V.; Auyeung, B.; Chakrabarti, B.; Baron-Cohen, S. Sex/Gender Differences and Autism: Setting the Scene for Future Research. J. Am. Acad. Child Adolesc. Psychiatry 2015, 54, 11–24. [Google Scholar] [CrossRef]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, Interconversion, and Dose Response of n-3 Fatty Acids in Humans. Am. J. Clin. Nutr. 2006, 83, 1467S–1476S. [Google Scholar] [CrossRef]
- Childs, C.E.; Romeu-Nadal, M.; Burdge, G.C.; Calder, P.C. Gender Differences in the N-3 Fatty Acid Content of Tissues. Proc. Nutr. Soc. 2008, 67, 19–27. [Google Scholar] [CrossRef]
- Field, S.S. Interaction of Genes and Nutritional Factors in the Etiology of Autism and Attention Deficit/Hyperactivity Disorders: A Case Control Study. Med. Hypotheses 2014, 82, 654–661. [Google Scholar] [CrossRef]
- Bennett, C.N.; Horrobin, D.F. Gene Targets Related to Phospholipid and Fatty Acid Metabolism in Schizophrenia and Other Psychiatric Disorders: An Update. Prostaglandins Leukot. Essent. Fatty Acids 2000, 63, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. Genetic Variants in the Metabolism of Omega-6 and Omega-3 Fatty Acids: Their Role in the Determination of Nutritional Requirements and Chronic Disease Risk. Exp. Biol. Med. 2010, 235, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Schneider, A.; Hagerman, R.J.; Hessl, D. Fragile X Syndrome—From Genes to Cognition. Dev. Disabil. Res. Rev. 2009, 15, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Srikantha, P.; Mohajeri, M.H. The Possible Role of the Microbiota-Gut-Brain-Axis in Autism Spectrum Disorder. Int. J. Mol. Sci 2019, 20, 2115. [Google Scholar] [CrossRef]
- Zurita, M.F.; Cárdenas, P.A.; Sandoval, M.E.; Peña, M.C.; Fornasini, M.; Flores, N.; Monaco, M.H.; Berding, K.; Donovan, S.M.; Kuntz, T.; et al. Analysis of Gut Microbiome, Nutrition and Immune Status in Autism Spectrum Disorder: A Case-Control Study in Ecuador. Gut Microbes 2020, 11, 453–464. [Google Scholar] [CrossRef]
- Mathias, P.C.F.; Elmhiri, G.; de Oliveira, J.C.; Delayre-Orthez, C.; Barella, L.F.; Tófolo, L.P.; Fabricio, G.S.; Chango, A.; Abdennebi-Najar, L. Maternal Diet, Bioactive Molecules, and Exercising as Reprogramming Tools of Metabolic Programming. Eur. J. Nutr. 2014, 53, 711–722. [Google Scholar] [CrossRef]
- Rustan, A.; Drevon, C. Fatty Acids: Structures and Properties. In Encyclopedia of Life Sciences; John Wiley & Sons: Hoboken, NJ, USA, 2005; ISBN 978-0-470-01590-2. [Google Scholar]
- Imhoff-Kunsch, B.; Briggs, V.; Goldenberg, T.; Ramakrishnan, U. Effect of N-3 Long-chain Polyunsaturated Fatty Acid Intake during Pregnancy on Maternal, Infant, and Child Health Outcomes: A Systematic Review. Paediatr. Perinat. Epidemiol. 2012, 26, 91–107. [Google Scholar] [CrossRef]
- Lee, J.M.; Lee, H.; Kang, S.B.; Park, W.J. Fatty acid desaturases, polyunsaturated fatty acid regulation, and biotechnological advances. Nutrients 2016, 8, 23. [Google Scholar] [CrossRef]
- Russo, G.L. Dietary n-6 and n-3 polyunsaturated fatty acids: From biochemistry to clinical implications in cardiovascular prevention. Biochem. Pharmacol. 2009, 77, 937–946. [Google Scholar] [CrossRef]
- Wiktorowska-Owczarek, A.; Berezinska, M.; Nowak, J. PUFAs: Structures, Metabolism and Functions. Adv. Clin. Exp. Med. 2015, 24, 931–941. [Google Scholar] [CrossRef]
- Van Vlies, N.; Hogenkamp, A.; Fear, A.L.; van Esch, B.C.; Oosting, A.; van de Heijning, B.; van der Beek, E.; Calder, P.C.; Garssen, J. Perinatal Programming of Murine Immune Responses by Polyunsaturated Fatty Acids. J. Dev. Orig. Health Dis. 2011, 2, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Steer, C.D.; Lattka, E.; Koletzko, B.; Golding, J.; Hibbeln, J.R. Maternal fatty acids in pregnancy, FADS polymorphisms, and child intelligence quotient at 8 y of age. Am. J. Clin. Nutr. 2013, 986, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Steenweg-de Graaff, J.; Tiemeier, H.; Ghassabian, A.; Rijlaarsdam, J.; Jaddoe, V.W.V.; Verhulst, F.C.; Roza, S.J. Maternal Fatty Acid Status During Pregnancy and Child Autistic Traits the Generation R Study. Am. J. Epidemiol. 2016, 183, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Iosif, A.-M.; Hansen, R.L.; Schmidt, R.J. Maternal Polyunsaturated Fatty Acids and Risk for Autism Spectrum Disorder in the MARBLES High-Risk Study. Autism Int. J. Res. Pract. 2020, 24, 1191–1200. [Google Scholar] [CrossRef] [PubMed]
- Hirokawa, K.; Kimura, T.; Ikehara, S.; Honjo, K.; Ueda, K.; Sato, T.; Iso, H.; Kishi, R.; Yaegashi, N.; Hashimoto, K.; et al. Associations Between Broader Autism Phenotype and Dietary Intake: A Cross-Sectional Study (Japan Environment & Children’s Study). J. Autism Dev. Disord. 2020, 50, 2698–2709. [Google Scholar] [CrossRef] [PubMed]
- Julvez, J.; Méndez, M.; Fernandez-Barres, S.; Romaguera, D.; Vioque, J.; Llop, S.; Ibarluzea, J.; Guxens, M.; Avella-Garcia, C.; Tardón, A.; et al. Maternal Consumption of Seafood in Pregnancy and Child Neuropsychological Development: A Longitudinal Study Based on a Population with High Consumption Levels. Am. J. Epidemiol. 2016, 183, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Hibbeln, J.R.; Davis, J.M.; Steer, C.; Emmett, P.; Rogers, I.; Williams, C.; Golding, J. Maternal Seafood Consumption in Pregnancy and Neurodevelopmental Outcomes in Childhood (ALSPAC Study): An Observational Cohort Study. Lancet 2007, 369, 578–585. [Google Scholar] [CrossRef]
- Vuillermot, S.; Luan, W.; Meyer, U.; Eyles, D. Vitamin D Treatment during Pregnancy Prevents Autism-Related Phenotypes in a Mouse Model of Maternal Immune Activation. Mol. Autism 2017, 8, 9. [Google Scholar] [CrossRef]
- Martins, B.P.; Bandarra, N.M.; Figueiredo-Braga, M. The Role of Marine Omega-3 in Human Neurodevelopment, Including Autism Spectrum Disorders and Attention-Deficit/Hyperactivity Disorder—A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 1431–1446. [Google Scholar] [CrossRef]
- Gluckman, P.D.; Hanson, M.A.; Cooper, C.; Thornburg, K.L. Effect of in Utero and Early-Life Conditions on Adult Health and Disease. N. Engl. J. Med. 2008, 359, 61–73. [Google Scholar] [CrossRef]
- US Food and Drug Administration. FDA/EPA 2004 Advice on What You Need to Know about Mercury in Fish and Shellfish. 2020. Available online: https://www.fda.gov/food/environmental-contaminants-food/fdaepa-2004-advice-what-you-need-know-about-mercury-fish-and-shellfish (accessed on 15 November 2022).
- Mennitti, L.V.; Oliveira, J.L.; Morais, C.A.; Estadella, D.; Oyama, L.M.; Oller do Nascimento, C.M.; Pisani, L.P. Type of Fatty Acids in Maternal Diets during Pregnancy and/or Lactation and Metabolic Consequences of the Offspring. J. Nutr. Biochem. 2015, 26, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Heiss, C.N.; Olofsson, L.E. The Role of the Gut Microbiota in Development, Function and Disorders of the Central Nervous System and the Enteric Nervous System. J. Neuroendocrinol. 2019, 31, e12684. [Google Scholar] [CrossRef] [PubMed]
- Kelly, J.R.; Minuto, C.; Cryan, J.F.; Clarke, G.; Dinan, T.G. Cross Talk: The Microbiota and Neurodevelopmental Disorders. Front. Neurosci. 2017, 11, 490. [Google Scholar] [CrossRef] [PubMed]
- Codagnone, M.G.; Spichak, S.; O’Mahony, S.M.; O’Leary, O.F.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Programming Bugs: Microbiota and the Developmental Origins of Brain Health and Disease. Biol. Psychiatry 2019, 85, 150–163. [Google Scholar] [CrossRef]
- Thion, M.S.; Low, D.; Silvin, A.; Chen, J.; Grisel, P.; Schulte-Schrepping, J.; Blecher, R.; Ulas, T.; Squarzoni, P.; Hoeffel, G.; et al. Microbiome Influences Prenatal and Adult Microglia in a Sex-Specific Manner. Cell 2018, 172, 500–516. [Google Scholar] [CrossRef]
- Chait, A.; Kim, F. Saturated fatty acids and inflammation: Who pays the toll? Arterioscler. Thromb. Vasc. Biol. 2010, 30, 692. [Google Scholar] [CrossRef]
- Santos, S.; Oliveira, A.; Lopes, C. Systematic review of saturated fatty acids on inflammation and circulating levels of adipokines. Nutr. Res. 2013, 33, 687–695. [Google Scholar] [CrossRef]
- Valdearcos, M.; Douglass, J.D.; Robblee, M.M.; Dorfman, M.D.; Stifler, D.R.; Bennett, M.L.; Gerritse, I.; Fasnacht, R.; Barres, B.A.; Thaler, J.P.; et al. Microglial Inflammatory Signaling Orchestrates the Hypothalamic Immune Response to Dietary Excess and Mediates Obesity Susceptibility. Cell Metab. 2018, 27, 1356. [Google Scholar] [CrossRef]
- Marrone, M.C.; Coccurello, R. Dietary fatty acids and microbiota-brain communication in neuropsychiatric diseases. Biomolecules 2020, 10, 12. [Google Scholar] [CrossRef]
- Lombardi, V.C.; De Meirleir, K.L.; Subramanian, K.; Nourani, S.M.; Dagda, R.K.; Delaney, S.L.; Palotás, A. Nutritional Modulation of the Intestinal Microbiota; Future Opportunities for the Prevention and Treatment of Neuroimmune and Neuroinflammatory Disease. J. Nutr. Biochem. 2018, 61, 1–16. [Google Scholar] [CrossRef]
- Nitschke, A.; Deonandan, R.; Konkle, A.T. The Link between Autism Spectrum Disorder and Gut Microbiota: A Scoping Review. Autism Int. J. Res. Pract. 2020, 24, 1328–1344. [Google Scholar] [CrossRef] [PubMed]
- Val-Laillet, D.; Besson, M.; Guérin, S.; Coquery, N.; Randuineau, G.; Kanzari, A.; Quesnel, H.; Bonhomme, N.; Bolhuis, J.E.; Kemp, B.; et al. A Maternal Western Diet during Gestation and Lactation Modifies Offspring’s Microbiota Activity, Blood Lipid Levels, Cognitive Responses, and Hippocampal Neurogenesis in Yucatan Pigs. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 2037–2049. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Molcan, E.; DeCoffe, D.; Dai, C.; Gibson, D.L. Diets Rich in N-6 PUFA Induce Intestinal Microbial Dysbiosis in Aged Mice. Br. J. Nutr. 2013, 110, 515–523. [Google Scholar] [CrossRef] [PubMed]
- Vohr, B.R.; Poggi Davis, E.; Wanke, C.A.; Krebs, N.F. Neurodevelopment: The Impact of Nutrition and Inflammation During Preconception and Pregnancy in Low-Resource Settings. Pediatrics 2017, 139, S38–S49. [Google Scholar] [CrossRef] [PubMed]
- Sotgiu, S.; Manca, S.; Gagliano, A.; Minutolo, A.; Melis, M.C.; Pisuttu, G.; Scoppola, C.; Bolognesi, E.; Clerici, M.; Guerini, F.R.; et al. Immune Regulation of Neurodevelopment at the Mother-Foetus Interface: The Case of Autism. Clin. Transl. Immunol. 2020, 9, e1211. [Google Scholar] [CrossRef] [PubMed]
- Irwin, J.L.; Yeates, A.J.; Mulhern, M.S.; McSorley, E.M.; Strain, J.J.; Watson, G.E.; Grzesik, K.; Thurston, S.W.; Love, T.M.; Smith, T.H.; et al. Maternal Gestational Immune Response and Autism Spectrum Disorder Phenotypes at 7 Years of Age in the Seychelles Child Development Study. Mol. Neurobiol. 2019, 56, 5000–5008. [Google Scholar] [CrossRef]
- Gibson, D.; Gill, S.; Brown, K.; Tasnim, N.; Ghosh, S.; Innis, S.; Jacobson, K. Maternal Exposure to Fish Oil Primes Offspring to Harbor Intestinal Pathobionts Associated with Altered Immune Cell Balance. Gut Microbes 2015, 6, 24–32. [Google Scholar] [CrossRef]
- Patterson, E.; O’ Doherty, R.M.; Murphy, E.F.; Wall, R.; O’ Sullivan, O.; Nilaweera, K.; Fitzgerald, G.F.; Cotter, P.D.; Ross, R.P.; Stanton, C. Impact of Dietary Fatty Acids on Metabolic Activity and Host Intestinal Microbiota Composition in C57BL/6J Mice. Br. J. Nutr. 2014, 111, 1905–1917. [Google Scholar] [CrossRef]
- Pusceddu, M.M.; El Aidy, S.; Crispie, F.; O’Sullivan, O.; Cotter, P.; Stanton, C.; Kelly, P.; Cryan, J.F.; Dinan, T.G. N-3 Polyunsaturated Fatty Acids (PUFAs) Reverse the Impact of Early-Life Stress on the Gut Microbiota. PLoS ONE 2015, 10, e0139721. [Google Scholar] [CrossRef]
- Oken, E.; Bellinger, D.C. Fish Consumption, Methylmercury and Child Neurodevelopment. Curr. Opin. Pediatr. 2008, 20, 178–183. [Google Scholar] [CrossRef]
- Johnson, D.; Letchumanan, V.; Thurairajasingam, S.; Lee, L.-H. A Revolutionizing Approach to Autism Spectrum Disorder Using the Microbiome. Nutrients 2020, 12, 1983. [Google Scholar] [CrossRef] [PubMed]
- Buffington, S.A.; Di Prisco, G.V.; Auchtung, T.A.; Ajami, N.J.; Petrosino, J.F.; Costa-Mattioli, M. Microbial Reconstitution Reverses Maternal Diet-Induced Social and Synaptic Deficits in Offspring. Cell 2016, 165, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Lohuis, M.A.M.; Werkman, C.C.N.; Harmsen, H.J.M.; Tietge, U.J.F.; Verkade, H.J. Absence of Intestinal Microbiota during Gestation and Lactation Does Not Alter the Metabolic Response to a Western-Type Diet in Adulthood. Mol. Nutr. Food Res. 2019, 63, 1800809. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.C.; Kaliannan, K.; Strain, C.R.; Ross, R.P.; Stanton, C.; Kang, J.X. Maternal omega-3 fatty acids regulate offspring obesity through persistent modulation of gut microbiota. Microbiome 2018, 6, 95. [Google Scholar] [CrossRef]
- Mesa, M.D.; Loureiro, B.; Iglesia, I.; Fernandez Gonzalez, S.; LlurbaOlivé, E.; García Algar, O.; Solana, M.J.; Cabero Perez, M.J.; Sainz, T.; Martinez, L.; et al. The Evolving Microbiome from Pregnancy to Early Infancy: A Comprehensive Review. Nutrients 2020, 12, 133. [Google Scholar] [CrossRef]
- Eshraghi, R.S.; Davies, C.; Iyengar, R.; Perez, L.; Mittal, R.; Eshraghi, A.A. Gut-Induced Inflammation during Development May Compromise the Blood-Brain Barrier and Predispose to Autism Spectrum Disorder. J. Clin. Med. 2020, 10, 27. [Google Scholar] [CrossRef]
- Rey, C.; Delpech, J.C.; Madore, C.; Nadjar, A.; Greenhalgh, A.D.; Amadieu, C.; Aubert, A.; Pallet, V.; Vaysse, C.; Layé, S.; et al. Dietary n-3 long chain PUFA supplementation promotes a pro-resolving oxylipin profile in the brain. Brain Behav. Immun. 2019, 76, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Calatayud, M.; Koren, O.; Collado, M.C. Maternal Microbiome and Metabolic Health Program Microbiome Development and Health of the Offspring. Trends Endocrinol. Metab. 2019, 30, 735–744. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, E.L.; Figueiredo, P.S.; Marcelino, G.; de Cássia Avellaneda Guimarães, R.; Pott, A.; Santana, L.F.; Hiane, P.A.; do Nascimento, V.A.; Bogo, D.; de Cássia Freitas, K. Maternal Intake of Polyunsaturated Fatty Acids in Autism Spectrum Etiology and Its Relation to the Gut Microbiota: What Do We Know? Nutrients 2023, 15, 1551. https://doi.org/10.3390/nu15071551
Rodrigues EL, Figueiredo PS, Marcelino G, de Cássia Avellaneda Guimarães R, Pott A, Santana LF, Hiane PA, do Nascimento VA, Bogo D, de Cássia Freitas K. Maternal Intake of Polyunsaturated Fatty Acids in Autism Spectrum Etiology and Its Relation to the Gut Microbiota: What Do We Know? Nutrients. 2023; 15(7):1551. https://doi.org/10.3390/nu15071551
Chicago/Turabian StyleRodrigues, Elisana Lima, Priscila Silva Figueiredo, Gabriela Marcelino, Rita de Cássia Avellaneda Guimarães, Arnildo Pott, Lidiani Figueiredo Santana, Priscila Aiko Hiane, Valter Aragão do Nascimento, Danielle Bogo, and Karine de Cássia Freitas. 2023. "Maternal Intake of Polyunsaturated Fatty Acids in Autism Spectrum Etiology and Its Relation to the Gut Microbiota: What Do We Know?" Nutrients 15, no. 7: 1551. https://doi.org/10.3390/nu15071551
APA StyleRodrigues, E. L., Figueiredo, P. S., Marcelino, G., de Cássia Avellaneda Guimarães, R., Pott, A., Santana, L. F., Hiane, P. A., do Nascimento, V. A., Bogo, D., & de Cássia Freitas, K. (2023). Maternal Intake of Polyunsaturated Fatty Acids in Autism Spectrum Etiology and Its Relation to the Gut Microbiota: What Do We Know? Nutrients, 15(7), 1551. https://doi.org/10.3390/nu15071551