
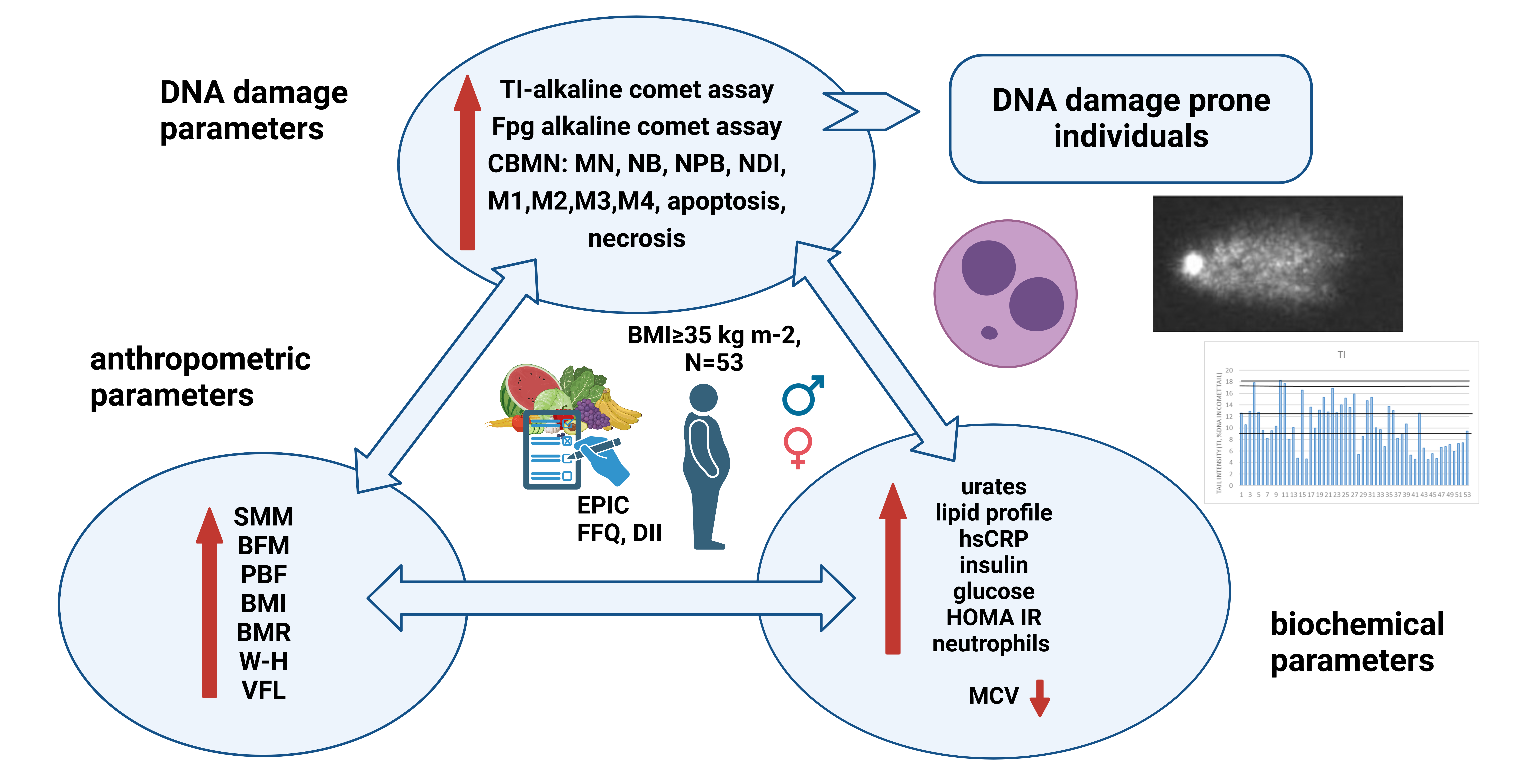
Combined Approach: FFQ, DII, Anthropometric, Biochemical and DNA Damage Parameters in Obese with BMI ≥ 35 kg m−2

 ,
,  ,
,
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Subject Recruitment, Food and Lifestyle Questionnaires
2.3. DII Calculation and Analysis
2.4. Anthropometric Analysis
2.5. Sampling
2.6. Biochemical Parameters Analysis
2.7. Alkaline and Fpg Comet Assay
2.8. Micronucleus Cytome Assay
2.9. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO-World Health Organization. World Obesity Day 2022—Accelerating Action to Stop Obesity. Available online: https://www.who.int/news/item/04-03-2022-world-obesity-day-2022-accelerating-action-to-stop-obesity (accessed on 28 December 2022).
- Weir, C.B.; Jan, A. BMI Classification Percentile and Cut Off Points. [Updated 2022 Jun 27]. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK541070/] (accessed on 3 January 2022).
- Eurostat—An Official Site of European Union. Available online: https://ec.europa.eu/eurostat/web/productseurostat-news/-/ddn-20210721-2 (accessed on 28 December 2022).
- Poirier, P.; Giles, T.D.; Bray, G.A.; Hong, Y.; Stern, J.S.; Pi-Sunyer, F.X.; Eckel, R.H.; American Heart Association; Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Obesity and Cardiovascular Disease: Pathophysiology, Evaluation, and Effect of Weight Loss: An update of the 1997 American Heart Association Scientific Statement on Obesity and Heart Disease from the Obesity Committee of the Council on Nutrition, Physical Activity, and Metabolism. Circulation 2006, 113, 898–918. [Google Scholar] [CrossRef]
- Guh, D.P.; Zhang, W.; Bansback, N.; Amarsi, Z.; Birmingham, C.L.; Anis, A.H. The incidence of co-morbidities related to obesity and overweight: A systematic review and meta-analysis. BMC Public Health 2009, 9, 88. [Google Scholar] [CrossRef]
- Lauby-Secretan, B.; Scoccianti, C.; Loomis, D.; Grosse, Y.; Bianchini, F.; Straif, K.; International Agency for Research on Cancer Handbook Working Group. Body Fatness and Cancer—Viewpoint of the IARC Working Group. N. Engl. J. Med. 2016, 375, 794–798. [Google Scholar] [CrossRef]
- Tsatsoulis, A.; Paschou, S.A. Metabolically Healthy Obesity: Criteria, Epidemiology, Controversies, and Consequences. Curr. Obes. Rep. 2020, 9, 109–120. [Google Scholar] [CrossRef]
- Lassale, C.; Tzoulaki, I.; Moons, K.G.M.; Sweeting, M.; Boer, J.; Johnson, L.; Huerta, J.M.; Agnoli, C.; Freisling, H.; Weiderpass, E.; et al. Separate and combined associations of obesity and metabolic health with coronary heart disease: A pan-European case-cohort analysis. Eur. Hearth J. 2018, 39, 397–406. [Google Scholar] [CrossRef]
- Bell, J.A.; Hamer, M.; Batty, G.D.; Singh-Manoux, A.; Sabia, S.; Kivimäki, M. Incidence of Metabolic Risk Factors Among Healthy Obese Adults: 20-year follow-up. J. Am. Coll. Cardiol. 2015, 66, 871–873. [Google Scholar] [CrossRef]
- Nyberg, S.T.; Batty, G.D.; Pentti, J.; Virtanen, M.; Alfredsson, L.; Fransson, E.I.; Goldberg, M.; Heikkilä, K.; Jokela, M.; Knutsson, A.; et al. Obesity and loss of disease-free years owing to major non-communicable diseases: A multicohort study. Lancet Public Health 2018, 3, e490–e497. [Google Scholar] [CrossRef]
- Lee, D.H.; Giovannucci, E.L. The Obesity Paradox in Cancer: Epidemiologic Insights and Perspectives. Curr. Nutr. Rep. 2019, 8, 175–181. [Google Scholar] [CrossRef]
- Milic, M.; Frustaci, A.; Del Bufalo, A.; Sánchez-Alarcón, J.; Valencia-Quintana, R.; Russo, P.; Bonassi, S. DNA damage in non-communicable diseases: A clinical and epidemiological perspective. Mutat. Res. Mol. Mech. Mutagen. 2015, 776, 118–127. [Google Scholar] [CrossRef]
- Neri, M.; Milazzo, D.; Ugolini, D.; Milic, M.; Campolongo, A.; Pasqualetti, P.; Bonassi, S. Worldwide interest in the comet assay: A bibliometric study. Mutagenesis 2015, 30, 155–163. [Google Scholar] [CrossRef]
- Bonassi, S.; Znaor, A.; Ceppi, M.; Lando, C.; Chang, W.P.; Holland, N.; Kirsch-Volders, M.; Zeiger, E.; Ban, S.; Barale, R.; et al. An increased micronucleus frequency in peripheral blood lymphocytes predicts the risk of cancer in humans. Carcinogenesis 2007, 28, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Fenech, M. Cytokinesis-block micronucleus cytome assay. Nat. Protoc. 2007, 2, 1084–1104. [Google Scholar] [CrossRef] [PubMed]
- Kopjar, N.; Kašuba, V.; Milić, M.; Rozgaj, R.; Želježić, D.; Gajski, G.; Mladinic, M.; Garaj-Vrhovac, V. Normalne i granicne vrijednosti mikronukleus-testa na limfocitima periferne krvi u ispitanika opće populacije Republike Hrvatske [Normal and Cut-Off Values of the Cytokinesis-Block Micronucleus Assay on Peripheral Blood Lymphocytes in the Croatian General Population]. Arch. Ind. Hyg. Toxicol. 2010, 61, 219–234. [Google Scholar] [CrossRef]
- Milić, M.; Ceppi, M.; Bruzzone, M.; Azqueta, A.; Brunborg, G.; Godschalk, R.; Koppen, G.; Langie, S.; Møller, P.; Teixeira, J.P.; et al. The hCOMET project: International database comparison of results with the comet assay in human biomonitoring. Baseline frequency of DNA damage and effect of main confounders. Mutat. Res. Mol. Mech. Mutagen. 2021, 787, 108371. [Google Scholar] [CrossRef]
- Bonassi, S.; Ceppi, M.; Møller, P.; Azqueta, A.; Milić, M.; Neri, M.; Brunborg, G.; Godschalk, R.; Koppen, G.; Langie, S.A.S.; et al. DNA damage in circulating leukocytes measured with the comet assay may predict the risk of death. Sci. Rep. 2021, 11, 16793. [Google Scholar] [CrossRef]
- Vodicka, P.; Vodenkova, S.; Opattova, A.; Vodickova, L. DNA damage and repair measured by comet assay in cancer patients. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 843, 95–110. [Google Scholar] [CrossRef]
- Franzke, B.; Schwingshackl, L.; Wagner, K.-H. Chromosomal damage measured by the cytokinesis block micronucleus cytome assay in diabetes and obesity—A systematic review and meta-analysis. Mutat. Res. Mol. Mech. Mutagen. 2020, 786, 108343. [Google Scholar] [CrossRef]
- Bankoglu, E.E.; Mukama, T.; Katzke, V.; Stipp, F.; Johnson, T.; Kühn, T.; Seyfried, F.; Godschalk, R.; Collins, A.; Kaaks, R.; et al. Short- and long-term reproducibility of the COMET assay for measuring DNA damage biomarkers in frozen blood samples of the EPIC-Heidelberg cohort. Mutat. Res. Toxicol. Environ. Mutagen. 2022, 874–875, 503442. [Google Scholar] [CrossRef]
- Azqueta, A.; Muruzabal, D.; Boutet-Robinet, E.; Milic, M.; Dusinska, M.; Brunborg, G.; Møller, P.; Collins, A.R. Technical recommendations to perform the alkaline standard and enzyme-modified comet assay in human biomonitoring studies. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 843, 24–32. [Google Scholar] [CrossRef]
- Collins, A.; Milic, M.; Bonassi, S.; Dusinska, M. The comet assay in human biomonitoring: Technical and epidemiological perspectives. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 843, 1–2. [Google Scholar] [CrossRef]
- Setayesh, T.; Nersesyan, A.; Mišík, M.; Ferk, F.; Langie, S.; Andrade, V.M.; Haslberger, A.; Knasmüller, S. Impact of obesity and overweight on DNA stability: Few facts and many hypotheses. Mutat. Res. Rev. Mutat. Res. 2018, 777, 64–91. [Google Scholar] [CrossRef]
- Włodarczyk, M.; Nowicka, G. Obesity, DNA Damage, and Development of Obesity-Related Diseases. Int. J. Mol. Sci. 2019, 20, 1146. [Google Scholar] [CrossRef]
- Setayesh, T.; Mišík, M.; Langie, S.A.S.; Godschalk, R.; Waldherr, M.; Bauer, T.; Leitner, S.; Bichler, C.; Prager, G.; Krupitza, G.; et al. Impact of Weight Loss Strategies on Obesity-Induced DNA Damage. Mol. Nutr. Food Res. 2019, 63, e1900045. [Google Scholar] [CrossRef] [PubMed]
- Ubago-Guisado, E.; Rodríguez-Barranco, M.; Ching-López, A.; Petrova, D.; Molina-Montes, E.; Amiano, P.; Barricarte-Gurrea, A.; Chirlaque, M.-D.; Agudo, A.; Sánchez, M.-J. Evidence Update on the Relationship between Diet and the Most Common Cancers from the European Prospective Investigation into Cancer and Nutrition (EPIC) Study: A Systematic Review. Nutrients 2021, 13, 3582. [Google Scholar] [CrossRef]
- Mulligan, A.A.; Luben, R.; Bhaniani, A.; Parry-Smith, D.J.; O’Connor, L.; Khawaja, A.; Forouhi, N.; Khaw, K.-T.; EPIC-Norfolk FFQ Study. A new tool for converting food frequency questionnaire data into nutrient and food group values: FETA research methods and availability. BMJ Open 2014, 4, e004503. [Google Scholar] [CrossRef] [PubMed]
- Riboli, E.; Hunt, K.; Slimani, N.; Ferrari, P.; Norat, T.; Fahey, M.; Charrondière, U.; Hémon, B.; Casagrande, C.; Vignat, J.; et al. European Prospective Investigation into Cancer and Nutrition (EPIC): Study populations and data collection. Public Health Nutr. 2002, 5, 1113–1124. [Google Scholar] [CrossRef] [PubMed]
- Hébert, J.R.; Shivappa, N.; Wirth, M.D.; Hussey, J.R.; Hurley, T.G. Perspective: The Dietary Inflammatory Index (DII)—Lessons Learned, Improvements Made, and Future Directions. Adv. Nutr. Int. Rev. J. 2019, 10, 185–195. [Google Scholar] [CrossRef]
- Shivappa, N.; Steck, S.E.; Hurley, T.G.; Hussey, J.R.; Hébert, J.R. Designing and developing a literature-derived, population-based dietary inflammatory index. Public Health Nutr. 2014, 17, 1689–1696. [Google Scholar] [CrossRef]
- Park, Y.-M.M.; Choi, M.K.; Lee, S.-S.; Shivappa, N.; Han, K.; Steck, S.E.; Hébert, J.R.; Merchant, A.T.; Sandler, D.P. Dietary inflammatory potential and risk of mortality in metabolically healthy and unhealthy phenotypes among overweight and obese adults. Clin. Nutr. 2019, 38, 682–688. [Google Scholar] [CrossRef]
- Rothwell, J.A.; Medina-Remón, A.; Pérez-Jiménez, J.; Neveu, V.; Knaze, V.; Slimani, N.; Scalbert, A. Effects of food processing on polyphenol contents: A systematic analysis using Phenol-Explorer data. Mol. Nutr. Food Res. 2015, 59, 160–170. [Google Scholar] [CrossRef]
- Milić, M.; Ožvald, I.; Vrček, I.V.; Lovrenčić, M.V.; Oreščanin, V.; Bonassi, S.; Del Castillo, E.R. Alkaline comet assay results on fresh and one-year frozen whole blood in small volume without cryo-protection in a group of people with different health status. Mutat. Res. Toxicol. Environ. Mutagen. 2019, 843, 3–10. [Google Scholar] [CrossRef]
- Collins, A.; Møller, P.; Gajski, G.; Vodenková, S.; Abdulwahed, A.; Anderson, D.; Bankoglu, E.E.; Bonassi, S.; Boutet-Robinet, E.; Brunborg, G.; et al. Measuring DNA modifications with the comet assay: A compendium of protocols. Nat. Protoc. 2023, 1–61. [Google Scholar] [CrossRef]
- Collins, A.R.; El Yamani, N.; Lorenzo, Y.; Shaposhnikov, S.; Brunborg, G.; Azqueta, A. Controlling variation in the comet assay. Front. Genet. 2014, 5, 359. [Google Scholar] [CrossRef] [PubMed]
- Ožvald, I.; Božičević, D.; Duh, L.; Vrček, I.V.; Pavičić, I.; Domijan, A.-M.; Milić, M. Effects of a 3-Week Hospital-Controlled Very-Low-Calorie Diet in Severely Obese Patients. Nutrients 2021, 13, 4468. [Google Scholar] [CrossRef] [PubMed]
- Eastmond, D.A.; Tucker, J.D. Identification of aneuploidy-inducing agents using cytokinesis-blocked human lymphocytes and an antikinetochore antibody. Environ. Mol. Mutagen. 1989, 13, 34–43. [Google Scholar] [CrossRef]
- Bonassi, S.; Fenech, M.; Lando, C.; Lin, Y.P.; Ceppi, M.; Chang, W.P.; Holland, N.; Kirsch-Volders, M.; Zeiger, E.; Ban, S.; et al. HUman MicroNucleus project: International database comparison for results with the cytokinesis-block micronucleus assay in human lymphocytes: I. Effect of laboratory protocol, scoring criteria, and host factors on the frequency of micronuclei. Environ. Mol. Mutagen. 2001, 37, 31. [Google Scholar] [CrossRef]
- Karelis, A.D.; Brochu, M.; Rabasa-Lhoret, R. Can we identify metabolically health but obese individuals (MHO)? Diabetes Metab. 2004, 30, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Liu, M.; Su, E.; Yu, P.; Jiang, H.; Zhao, J.; Ge, J. Elevated circulating high-sensitivity cardiac troponin t and cardiac remodeling in obesity. BMC Cardiovasc. Disord. 2021, 21, 620. [Google Scholar] [CrossRef]
- Jeong, H.R.; Lee, H.S.; Shim, Y.S.; Hwang, J.S. Positive Associations between Body Mass Index and Hematological Parameters, Including RBCs, WBCs, and Platelet Counts, in Korean Children and Adolescents. Children 2022, 9, 109. [Google Scholar] [CrossRef] [PubMed]
- Uribe-Querol, E.; Rosales, C. Neutrophils Actively Contribute to Obesity-Associated Inflammation and Pathological Complications. Cells 2022, 11, 1883. [Google Scholar] [CrossRef]
- Sapio, L.; Naviglio, S. Inorganic phosphate in the development and treatment of cancer: A Janus Bifrons? World J. Clin. Oncol. 2015, 6, 198–201. Available online: http://www.wjgnet.com/2218-4333/full/v6/i6/198.htm (accessed on 9 December 2022). [CrossRef]
- Nuttall, F.Q. Body Mass Index: Obesity, BMI, and health: A critical review. Nutr. Today 2015, 50, 117–128. [Google Scholar] [CrossRef]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic. Report of a WHO Consultation; World Health Organ Technical Report Series 2000; World Health Organization: Geneva, Switzerland, 2000; Volume 894. [Google Scholar]
- NHLBI. Obesity Education Initiative Expert Panel on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults (US). In Clinical Guidelines on the Identification, Evaluation, and Treatment of Overweight and Obesity in Adults: The Evidence Report; Report No.: 98; National Institutes of Health, Ed.; National Heart, Lung, and Blood Institute: Bethesda, MD, USA, 1998. Available online: https://www.ncbi.nlm.nih.gov/books/NBK2003/ (accessed on 3 January 2023).
- Nimptsch, K.; Konigorski, S.; Pischon, T. Diagnosis of obesity and use of obesity biomarkers in science and clinical medicine. Metabolism 2019, 92, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Ahima, R.S.; Lazar, M.A. Physiology. The Health Risk of Obesity—Better Metrics Imperative. Science 2013, 341, 856–858. [Google Scholar] [CrossRef] [PubMed]
- Drabsch, T.; Holzapfel, C.; Stecher, L.; Petzold, J.; Skurk, T.; Hauner, H. Associations between C-reactive protein, insu-lin sensitivity, and resting metabolic rate in adults: A mediator analysis. Front. Endocrin. 2018, 9, 556. [Google Scholar] [CrossRef]
- Ruggiero, C.; Metter, E.J.; Melenovsky, V.; Cherubini, A.; Najjar, S.S.; Ble, A.; Senin, U.; Longo, D.L.; Ferrucci, L. High basal metabolic rate is a risk factor for mortality: The Baltimore longitudinal study of aging. J. Gerontol. A Biol. 2008, 63, 698–706. [Google Scholar] [CrossRef]
- Kliemann, N.; Murphy, N.; Viallon, V.; Freisling, H.; Tsilidis, K.K.; Rinaldi, S.; Mancini, F.R.; Fagherazzi, G.; Boutron-Ruault, M.; Boeing, H.; et al. Predicted basal metabolic rate and cancer risk in the European Prospective Investigation into Cancer and Nutrition. Int. J. Cancer 2020, 147, 648–661. [Google Scholar] [CrossRef]
- Chaput, J.-P.; Doucet, E.; Tremblay, A. Obesity: A disease or a biological adaptation? An update. Obes. Rev. 2012, 13, 681–691. [Google Scholar] [CrossRef]
- Mozaffarian, D. Dietary and Policy Priorities for Cardiovascular Disease, Diabetes, and Obesity: A Comprehensive Review. Circulation 2016, 133, 187–225. [Google Scholar] [CrossRef]
- World Health Organization. Diet, Nutrition and the Prevention of Chronic Diseases; World Health Organ Technical Report Series 2000; World Health Organization: Geneva, Switzerland, 2003; Volume 916. [Google Scholar]
- Schlesinger, S.; Neuenschwander, M.; Schwedhelm, C.; Hoffmann, G.; Bechthold, A.; Boeing, H.; Schwingshackl, L. Food Groups and Risk of Overweight, Obesity, and Weight Gain: A Systematic Review and Dose-Response Meta-Analysis of Prospective Studies. Adv. Nutr. Int. Rev. J. 2019, 10, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Milić, M.; Kišan, M.; Rogulj, D.; Radman, M.; Lovrenčić, M.V.; Konjevoda, P.; Domijan, A.-M. Level of primary DNA damage in the early stage of metabolic syndrome. Mutat. Res. Toxicol. Environ. Mutagen. 2013, 758, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rogulj, D.; Konjevoda, P.; Milić, M.; Mladinić, M.; Domijan, A.-M. Fatty liver index as an indicator of metabolic syndrome. Clin. Biochem. 2012, 45, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Ladeira, C.; Giovannelli, L.; Boutet-Robinet, E.; Bonassi, S.; Neri, M.; Gajski, G.; Duthie, S.; Del Bo’, C.; Riso, P.; et al. Application of the comet assay in human biomonitoring: An hCOMET perspective. Mutat. Res. Mol. Mech. Mutagen. 2020, 783, 108288. [Google Scholar] [CrossRef] [PubMed]
- Bankoglu, E.E.; Stopper, H. Obesity related genomic instability and altered xenobiotic metabolism: Possible consequences for cancer risk and chemotherapy. Expert Rev. Mol. Med. 2022, 24, e28. [Google Scholar] [CrossRef]
- Wilson, P.W.; D’Agostino, R.B.; Parise, H.; Sullivan, L.; Meigs, J.B. Metabolic Syndrome as a Precursor of Cardiovascular Disease and Type 2 Diabetes Mellitus. Circulation 2005, 112, 3066–3072. [Google Scholar] [CrossRef]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Kim, J.; Lee, J. Role of obesity-induced inflammation in the development of insulin resistance and type 2 diabetes: History of the research and remaining questions. Ann. Pediatr. Endocrinol. Metab. 2021, 26, 1–13. [Google Scholar] [CrossRef]
- Stępień, M.; Stępień, A.; Wlazeł, R.N.; Paradowski, M.; Banach, M.; Rysz, J. Obesity indices and inflammatory markers in obese non-diabetic normo- and hypertensive patients: A comparative pilot study. Lipids Health Dis. 2014, 13, 29. [Google Scholar] [CrossRef]
- Ellulu, M.S.; Patimah, I.; KhazáAi, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Gilbert-Diamond, D.; Baylin, A.; Mora-Plazas, M.; Villamor, E. Chronic inflammation is associated with overweight in Colombian school children. Nutr. Metab. Cardiovasc. Dis. 2012, 22, 244–251. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Shin, Y.-H.; Kim, J.-K.; Shim, J.; Kang, D.-R.; Lee, H.-R. Metabolic syndrome and its association with white blood cell count in children and adolescents in Korea: The 2005 Korean National Health and Nutrition Examination Survey. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 165–172. [Google Scholar] [CrossRef]
- Nieto, F.J.; Szklo, M.; Folsom, A.R.; Rock, R.; Mercuri, M. Leukocyte Count Correlates in Middle-aged Adults: The Atherosclerosis Risk in Communities (ARIC) Study. Am. J. Epidemiol. 1992, 136, 525–537. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.; Weiss, S.T. Host and Environmental Factors Influencing the Peripheral Blood Leukocyte Count. Am. J. Epidemiology 1991, 134, 1402–1409. [Google Scholar] [CrossRef]
- Samocha-Bonet, D.; Justo, D.; Rogowski, O.; Saar, N.; Abu-Abeid, S.; Shenkerman, G.; Shapira, I.; Berliner, S.; Tomer, A. Platelet Counts and Platelet Activation Markers in Obese Subjects. Mediat. Inflamm. 2008, 2008, 834153. [Google Scholar] [CrossRef]
- Santilli, F.; Vazzana, N.; Liani, R.; Guagnano, M.T.; Davì, G. Platelet activation in obesity and metabolic syndrome. Obes. Rev. 2012, 13, 27–42. [Google Scholar] [CrossRef]
- Timerga, A.; Haile, K. Patterns of Calcium- and Chloride-Ion Disorders and Predictors among Obese Outpatient Adults in Southern Ethiopia. Diabetes, Metab. Syndr. Obesity: Targets Ther. 2021, 14, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.B. Obesity and Cancer: Potential Mediation by Dysregulated Dietary Phosphate. Obesities 2022, 2, 64–75. [Google Scholar] [CrossRef]
- Mazzarella, L.; Falvo, P.; Adinolfi, M.; Tini, G.; Gatti, E.; Piccioni, R.; Bonetti, E.; Gavilan Dorronzoro, E.; Bodini, M.; Gallo, B.; et al. High-fat diet promotes Acute Promyelocytic Leukemia through PPARδ-enhanced self-renewal of preleukemic progenitors. bioRxiv 2022. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable/Unit, Reference Values | Mean | Median | Min | Max | SD | SE | Spearman Correlation (R) |
|---|---|---|---|---|---|---|---|
| Weight/kg, 85–115 | 126.62 | 126.00 | 95.30 | 183.60 | 19.29 | 2.65 | SMM(0.84), BFM(0.68), BMI(0.72), BMR(0.83) |
| SMM/kg, 19.8–24.2 | 37.27 | 34.90 | 23.80 | 56.20 | 8.13 | 1.12 | weight(0.84), BMI (0.34), BMR(1), PBF%(−0.47), VFL(−0.42) |
| BFM/kg, 10.8–17.2 | 60.78 | 62.00 | 40.40 | 90.30 | 11.52 | 1.58 | weight(0.68), PBF%(0.56), BMI(0.88), VFL(0.28) |
| PBF/%, 10–20 M, 18–28 F | 47.94 | 50.00 | 30.30 | 64.50 | 6.59 | 0.91 | BFM(0.56), BMI(0.37), VFL(0.37), SMM(−0.47), BMR(−0.48) |
| BMI/kgm−2, 18.5–25 | 44.58 | 43.60 | 35.40 | 57.50 | 5.77 | 0.79 | weight(0.72), SMM(0.34), BFM(0.88), PBF%(0.37), BMR(0.33) |
| BMR/kcal day−1, 1255–1451 | 1792.28 | 1703.00 | 1299.00 | 2462.00 | 290.91 | 39.96 | weight(0.83), SMM(1), BMI(0.33), PBF%(−0.48), VFL(−0.42) |
| W-H ratio, 0.75–0.85 | 1.05 | 1.06 | 0.70 | 1.27 | 0.11 | 0.01 | - |
| VFL, 10 | 19.77 | 20.00 | 16.00 | 20.00 | 0.80 | 0.11 | BFM(0.28), PBF%(0.37), SMM(−0.42), BMR(−0.42) |
| Variables, Reference Values | Mean | Median | Min | Max | SD | SE | Spearman Correlations |
|---|---|---|---|---|---|---|---|
| Erythrocytes, 3.86–5.08 F, 4.34–5.72 M ∗ 1012/L | 4.85 | 4.82 | 3.86 | 5.81 | 0.41 | 0.06 | |
| Hemoglobin, 119–157 F, 138–175 M g/L | 139.04 | 138.00 | 107.00 | 165.00 | 13.65 | 1.88 | |
| Hematocrit, 0.356–0.47 F, 0.414–0.53 M L/L | 0.41 | 0.41 | 0.33 | 0.48 | 0.03 | 0.00 | |
| MCV, 83–97.2 FM fl | 84.63 | 85.30 | 62.80 | 94.30 | 6.14 | 0.84 | insulin(−0.56), HOMA-IR(−0.51), erythrocytes(−0.46) |
| RDW %, 9–15 FM | 13.91 | 13.50 | 12.40 | 19.70 | 1.28 | 0.18 | chlorides(0.42) |
| Platelets, 158–424 FM ∗ 109/L | 272.30 | 249.00 | 167.00 | 563.00 | 86.76 | 11.92 | leukocytes(0.61), neutrophils(0.58), monocytes(0.41) |
| Leukocytes, 3.4–9.7 FM ∗ 109/L | 8.20 | 7.70 | 5.04 | 14.26 | 2.12 | 0.29 | neutrophils(0.91), monocytes(0.71), platelets(0.61), lymphocytes(0.52), %neutro(0.44), mono%(−0.43) |
| Basophils, 0–0.06 FM, ∗ 109/L | 0.03 | 0.03 | 0.01 | 0.09 | 0.02 | 0.00 | |
| Basophils %, 0–1 FM | 0.44 | 0.40 | 0.10 | 1.30 | 0.24 | 0.03 | |
| Neutrophils, 2.06–6.49 FM ∗ 109/L | 4.85 | 4.63 | 2.00 | 9.65 | 1.66 | 0.23 | leukocytes(0.91), lymphocytes%(−0.65), monocytes(0.63), platelets(0.58), hsCRP(0.45), monocytes%(−0.42) |
| Neutrophils %, 44–72 FM | 58.28 | 58.30 | 38.60 | 75.00 | 8.22 | 1.13 | lymphocytes%(−0.95), lymphocytes(−0.45), leukocytes(0.44), hsCRP(0.44) |
| Lymphocytes, 1.19–3.35 FM *109/L | 2.39 | 2.26 | 1.18 | 4.10 | 0.71 | 0.10 | leukocytes(0.52), LDL-C(0.47), neutrophils%(−0.45), cholesterol(0.43) |
| Lymphocytes %, 20–46 FM | 29.77 | 29.00 | 14.60 | 49.20 | 7.88 | 1.08 | neutrophils(−0.65), hsCRP(−0.43) |
| Monocytes, 0.12–0.84 FM ∗ 109/L | 0.68 | 0.64 | 0.36 | 1.24 | 0.18 | 0.02 | leukocytes(0.71), neutrophils(0.63), platelets(0.41), W-H-ratio(0.40) |
| Monocytes %, 2–12 FM | 8.48 | 8.20 | 4.80 | 13.10 | 1.81 | 0.25 | leukocytes(−0.43), neutrophils(−0.42), urates(0.41 |
| Glucose, 4.4–6.4 FM mmol/L | 7.18 | 6.40 | 4.30 | 19.10 | 2.30 | 0.32 | HOMA-IR(0.57), hs troponin(0.50), fT4(0.49) |
| Urate, 134–337 F, 182–403 M µmol/L | 412.19 | 395.00 | 247.00 | 838.00 | 122.28 | 16.80 | monocytes%(0.41), platelets(−0.39), SMM(0.37), BMR(0.37), hs troponin(0.35) |
| Urea, 2.8–8.3 mmol/L | 6.64 | 6.00 | 3.90 | 18.70 | 2.58 | 0.35 | erythrocytes(0.50), hemoglobin(0.45), hematocrit(0.41) |
| Total calcium, 2.14–2.53 FM mmol/L | 2.43 | 2.43 | 2.23 | 2.71 | 0.11 | 0.02 | inorganic phosphates(0.51), chlorides(0.46), albumin(0.41) |
| Inorganic phosphates, 0.79–1.42 FM mmol/L | 1.03 | 0.93 | 0.74 | 1.53 | 0.23 | 0.03 | chlorides(0.71), Ca total(0.51), hstroponin(0.51), weight(0.36), BMR(0.36), platelets(−0.36), fT4(0.35), SMM(0.35), BMI(0.35) |
| Chlorides, 97–108 mmol/L | 50.55 | 1.35 | 0.84 | 108.00 | 50.91 | 6.99 | inorganic phosphates(0.71), Ca total(0.46), BMI(0.46), hs troponin(0.44), RDW(0.42), weight(0.41), platelets(−0.39), BMR(0.38), SMM(0.37) |
| Albumin, 40.6–51.4 FM g/L | 44.07 | 44.00 | 38.20 | 49.30 | 2.48 | 0.34 | BFM(−0.47),Ca total(0.41), fT3(0.41) |
| hsCRP, ≤5 FM mg/L | 9.94 | 6.78 | 0.99 | 39.64 | 9.20 | 1.26 | lymphocytes%(−0.43) |
| hs troponin I, ≤15.6 F, ≤34.2 M ng/L | 3.12 | 2.00 | 0.10 | 18.70 | 3.47 | 0.48 | inorganic phosphates(0.51), glucose(0.50), BMR(0.49), SMM(0.46), chlorides(0.44), VFL(−0.39), basophils(0.37), weight(0.37), basophils%(0.36), TC(−0.36), fT4(0.36), LDL-C(−0.36), urates(0.35) |
| TC, ≤5 FM, mmol/L | 5.65 | 5.60 | 3.00 | 10.10 | 1.46 | 0.20 | LDL-C(0.96), lymphocytes(0.43), TG(0.41), HDL-C(0.40), hs troponin(−0.36) |
| HDL-C, ≤1.2 F, ≤1 M mmol/L | 1.24 | 1.20 | 0.80 | 2.10 | 0.30 | 0.04 | BMR(−0.45), SMM(−0.44), weight(−0.41), TC(0.40) |
| LDL-C, ≤3 FM mmol/L | 3.56 | 3.20 | 1.00 | 7.70 | 1.25 | 0.17 | TC(0.96), lymphocytes(0.47), hs troponin(−0.36), TG(0.35) |
| TG, ≤1.7 FM mmol/L | 2.02 | 2.00 | 0.80 | 4.60 | 0.82 | 0.11 | TC(0.41), LDL-C(0.35) |
| TSH, 0.35–4.94 FM mIU/L | 2.38 | 2.05 | 0.08 | 7.69 | 1.39 | 0.19 | |
| fT4, 9–19.5 FM pmol/L | 12.80 | 12.60 | 9.32 | 18.57 | 1.91 | 0.26 | glucose(0.49) |
| fT3, 2.42–6 FM, pmol/L | 4.22 | 4.25 | 2.64 | 5.59 | 0.72 | 0.10 | albumin(0.41), BMI(−0.40) |
| Insulin, 2–22 FM mU/L | 17.40 | 15.10 | 4.30 | 56.30 | 8.87 | 1.22 | HOMA-IR(0.90), MCV(−0.56), weight(0.40), erythrocytes(0.39) |
| HOMA-IR, <2.5 FM | 5.85 | 4.68 | 1.40 | 19.77 | 4.06 | 0.56 | insulin(0.90), glucose(0.57), MCV(−0.51), BMI(0.43), weight(0.41) |
| Micronucleus Assay, Reference Range | Mean | Median | Min | Max | SD | SE |
|---|---|---|---|---|---|---|
| M1 | 538.96 | 545.00 | 186.00 | 912.00 | 264.62 | 36.35 |
| SC: NDI(−0.98), M4(0.91), M3(−0.91), M2(−0.89), NB(−0.43), inorganic phosphates(0.52), chlorides(0.50), BMI(0.48), fT3(−0.45), weight(0.38), Ca total(0.38), BFM(0.36), MN(0.35) | ||||||
| M2 | 316.94 | 330.00 | 84.00 | 575.00 | 156.10 | 21.44 |
| SC: M1(0.89), NDI(0.81), M4(0.72), M3(0.71), fT3(0.49), chlorides(−0.48), NB(0.45), BMI(−0.46), BFM(−0.38), inorganic phosphates(−0.42), weight(−0.35), MN(−0.35) | ||||||
| M3 | 51.83 | 51.00 | 1.00 | 140.00 | 42.55 | 5.84 |
| SC: NDI(0.93), M4(0.92), M1(−0.91), M2(0.71), inorganic phosphates(−0.55), chlorides(−0.53), BMI(−0.46), hs troponin(−0.43), fT3(0.42), weight(−0.40), Ca total(−0.40), MN(−0.39), fT4(−0.38), NB(0.35) | ||||||
| M4 | 91.87 | 61.00 | 0.00 | 302.00 | 94.59 | 12.99 |
| SC: NDI(0.96), M3(0.92), M1(−0.91), M2(0.72), chlorides(−0.58), inorganic phosphates(−0.55), BMI(−0.54), NB(0.47), weight(−0.46), hs troponin(−0.43), Ca total(−0.41), fT3(0.40), SMM(−0.35), BFM(−0.35) | ||||||
| NDI, ~2 | 1.70 | 1.74 | 1.09 | 2.55 | 0.48 | 0.07 |
| SC: M1(−0.98), M4(0.96), M3(0.93), M2(0.81), inorganic phosphates(−0.53), chlorides(−0.52), BMI(−0.48), weight(−0.42), fT3(0.41), NB(0.40), hs troponin(−0.39), Ca total(−0.37) | ||||||
| FREQ MN, 0–12.5 | 9.00 | 7.50 | 1.50 | 24.00 | 5.10 | 0.70 |
| SC: inorganic phosphates(0.42), M3(−0.39), fT3(−0.39) | ||||||
| FREQ NB, 0–5 | 5.60 | 3.50 | 0.00 | 21.00 | 5.12 | 0.70 |
| SC: M4(0.47), chlorides(−0.46), M2(0.45), Ca total(−0.43), M1(−0.43), NDI(0.40), BMI(−0.39), fT3(0.39), W-H-ratio(0.38), M3(0.35) | ||||||
| FREQ NPB, 0–10 | 5.74 | 4.50 | 0.50 | 17.00 | 4.31 | 0.59 |
| SC: urates(0.36) | ||||||
| APOPTOSIS, 0–7% | 7.91 | 7.00 | 0.00 | 42.00 | 7.02 | 0.96 |
| SC: mean comet(0.36) | ||||||
| NECROSIS, 0–9% | 2.79 | 0.00 | 0.00 | 33.00 | 7.45 | 1.02 |
| SC: TSH(0.53), inorganic phosphates(0.37), glucose(0.35) | ||||||
| Alkaline comet assay | ||||||
| Mean comet (TI), 0–9% | 10.51 | 10.04 | 4.50 | 18.27 | 4.00 | 0.55 |
| SC: TG(0.43), chlorides(0.41), apoptosis(0.36), glucose(0.35) | ||||||
| Fpg alkaline comet assay | ||||||
| Net Fpg (TI), 0% | 6.28 | 4.45 | 0.00 | 31.59 | 6.20 | 0.85 |
| Food Groups (g) | Mean | Median | Min | Max | SD | SE | Spearman Correlation(R) |
|---|---|---|---|---|---|---|---|
| Alcoholic beverages | 27.26 | 7.00 | 0.00 | 316.00 | 52.57 | 7.22 | Positive: Alcohol(0.95), Negative: Age(−0.31) |
| Cereals and cereal products | 264.80 | 188.91 | 0.56 | 2585.28 | 364.44 | 50.06 | Negative: DII(−0.44), TG(−0.27), Net Fpg(−0.36), TI (−0.28) |
| Eggs and egg dishes | 16.28 | 17.50 | 0.00 | 50.00 | 13.07 | 1.80 | Positive: weight(0.28), MN(0.33) |
| Fats and oils | 12.32 | 6.51 | 0.00 | 177.12 | 26.51 | 3.64 | Negative: TG(−0.29) |
| Fish and fish products | 27.45 | 24.15 | 0.00 | 101.00 | 19.14 | 2.63 | Negative: DII(−0.32), Net Fpg(−0.30) |
| Fruit | 184.25 | 129.75 | 17.50 | 1477.80 | 232.51 | 31.94 | Negative: DII(−0.47) |
| Meat and meat products | 158.88 | 134.89 | 20.65 | 613.13 | 108.22 | 14.87 | Negative: DII(−0.47), M4(−0.28) |
| Milk and milk products | 101.37 | 78.30 | 0.00 | 423.20 | 97.54 | 13.40 | Positive: Insulin(0.29), Negative: DII(−0.38) |
| Non-alcoholic beverages | 634.95 | 515.00 | 9.48 | 1791.10 | 427.55 | 58.73 | Negative: DII(−0.35) |
| Nuts and seeds | 13.40 | 2.10 | 0.00 | 137.66 | 26.01 | 3.57 | - |
| Potatoes | 91.49 | 74.54 | 0.00 | 392.69 | 75.24 | 10.34 | Positive: Weight(0.33), BFM(0.28), Negative: DII(−0.46) |
| Soups and sauces | 97.30 | 78.00 | 0.00 | 377.90 | 76.06 | 10.45 | Positive: NPB(0.28), Negative: DII(−0.34) |
| Sugars | 50.50 | 22.49 | 0.00 | 530.50 | 83.34 | 11.45 | Negative: hs troponin(−0.35), TI(−0.34), age(−0.51) |
| Preserves and snacks | 225.85 | 213.05 | 20.64 | 747.81 | 140.21 | 19.26 | Positive: HDL-C(0.28), Negative: DII(−0.64), Net Fpg(−0.27) |
| Vegetables | 4.34 | 5.00 | 0.00 | 5.00 | 1.71 | 0.23 | Positive: fT3(0.31), M2(0.37), M3(0.40), M4(0.35), NDI(0.39), NB(0.44), Negative: hs troponin(−0.32), weight(−0.34), M1(−0.38), TI(−0.29) |
| Age 51–59, n = 9 | Age 41–50, n = 17 | Age < 40, n = 9 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Variable | Mean | Median | Min | Max | SD | SE | Mean | Median | Min | Max | SD | SE | Mean | Median | Min | Max | SD | SE |
| Alpha carotene Alcohol (mcg) | 105.04 | 101.09 | 8.58 | 203.51 | 76.84 | 25.61 | 204.76 | 106.30 | 4.62 | 994.71 | 261.21 | 63.35 | 123.85 | 100.32 | 9.43 | 201.53 | 72.72 | 24.24 |
| Alcohol (g) | 5.70 | 0.51 | 0.00 | 34.79 | 11.33 | 3.78 | 5.93 | 2.50 | 0.00 | 34.03 | 9.82 | 2.38 | 5.60 | 0.51 | 0.00 | 43.75 | 14.36 | 4.79 |
| Beta carotene (mcg) | 1573.50 | 1634.15 | 608.99 | 2479.17 | 690.09 | 230.03 | 2218.75 | 1764.11 | 632.18 | 5381.19 | 1501.02 | 364.05 | 1697.99 | 1125.44 | 301.25 | 4972.47 | 1492.09 | 497.36 |
| Calcium (mg) | 573.15 | 487.27 | 219.54 | 1238.94 | 294.13 | 98.04 | 832.65 | 571.35 | 415.44 | 3734.16 | 795.68 | 192.98 | 645.69 | 568.80 | 263.16 | 1184.92 | 329.03 | 109.68 |
| Carotene—total (carotene equivalents) (mcg) | 1845.98 | 2056.93 | 716.80 | 2729.51 | 802.05 | 267.35 | 2598.26 | 2023.78 | 638.77 | 5945.25 | 1705.91 | 413.74 | 1947.99 | 1277.80 | 359.58 | 5597.43 | 1697.27 | 565.76 |
| Carbohydrate—total (g) | 185.08 | 179.68 | 105.96 | 298.98 | 68.85 | 22.95 | 300.65 | 214.45 | 54.23 | 1771.66 | 390.31 | 94.66 | 250.55 | 175.76 | 104.99 | 633.13 | 176.59 | 58.86 |
| Cholesterol (mg) | 286.21 | 305.22 | 118.47 | 460.33 | 114.99 | 38.33 | 411.87 | 265.58 | 187.77 | 2284.20 | 491.91 | 119.31 | 281.40 | 210.48 | 124.05 | 617.10 | 171.84 | 57.28 |
| Chloride (mg) | 3626.55 | 3867.86 | 1727.77 | 5274.23 | 1236.18 | 412.06 | 4781.56 | 3537.11 | 2382.51 | 18,279.1 | 3747.57 | 908.92 | 3585.57 | 2563.62 | 1528.34 | 7917.96 | 2386.75 | 795.584 |
| Copper (mg) | 1.53 | 1.36 | 0.76 | 3.43 | 0.80 | 0.27 | 2.35 | 1.40 | 0.70 | 11.89 | 2.72 | 0.66 | 1.24 | 0.97 | 0.39 | 3.10 | 0.85 | 0.28 |
| Englyst Fibre—non-starch polysaccharides(NSP)(g) | 13.71 | 14.06 | 7.91 | 18.94 | 3.94 | 1.31 | 20.05 | 15.74 | 7.85 | 62.67 | 13.89 | 3.37 | 4677.40 | 10.94 | 3.81 | 42,003.0 | 13,997.1 | 4665.70 |
| Iron (mg) | 10.14 | 11.46 | 5.01 | 14.39 | 2.87 | 0.96 | 14.76 | 11.65 | 6.93 | 54.90 | 11.25 | 2.729 | 9.61 | 7.84 | 3.71 | 20.95 | 5.78 | 1.93 |
| Total folate (mcg) | 241.21 | 253.96 | 121.49 | 342.18 | 82.86 | 27.62 | 300.43 | 277.25 | 152.30 | 726.66 | 153.91 | 37.33 | 203.91 | 175.71 | 73.20 | 432.60 | 125.17 | 41.72 |
| Carbohydrate—fructose (g) | 23.16 | 24.00 | 10.60 | 46.92 | 11.47 | 3.82 | 25.66 | 16.24 | 5.48 | 110.14 | 25.85 | 6.27 | 21.02 | 11.22 | 4.82 | 88.20 | 26.56 | 8.85 |
| Carbohydrate—galactose (g) | 0.54 | 0.27 | 0.00 | 1.44 | 0.55 | 0.18 | 0.91 | 0.48 | 0.00 | 4.41 | 1.22 | 0.30 | 0.32 | 0.12 | 0.00 | 1.01 | 0.34 | 0.12 |
| Carbohydrate—glucose (g) | 22.15 | 20.41 | 11.37 | 40.93 | 10.23 | 3.41 | 26.48 | 15.62 | 4.90 | 125.52 | 28.60 | 6.94 | 22.37 | 15.68 | 5.36 | 77.32 | 22.38 | 7.46 |
| Carbohydrate—glucose (mcg) | 68.18 | 69.99 | 31.57 | 104.29 | 24.76 | 8.25 | 127.15 | 76.06 | 45.18 | 662.59 | 146.71 | 35.58 | 85.85 | 62.65 | 38.10 | 194.69 | 53.28 | 17.76 |
| Potassium (mg) | 2750.84 | 2580.86 | 1869.45 | 3835.49 | 684.62 | 228.21 | 3985.89 | 3824.90 | 1821.53 | 12,996.9 | 2649.54 | 642.61 | 3002.97 | 2198.26 | 1155.01 | 7816.25 | 2120.70 | 706.90 |
| Energy_kcal | 1470.51 | 1511.38 | 844.57 | 2055.98 | 478.51 | 159.50 | 2443.35 | 1709.93 | 1156.04 | 12,727.6 | 2728.69 | 661.81 | 2132.80 | 2016.03 | 870.56 | 4563.85 | 1161.46 | 387.16 |
| Energy_kj | 6191.10 | 6358.97 | 3563.39 | 8645.21 | 2013.56 | 671.19 | 10,273.7 | 7195.29 | 4805.64 | 53,529.7 | 11,478.7 | 2783.99 | 8098.61 | 5961.07 | 3664.38 | 19,180.3 | 5060.99 | 1687.00 |
| Carbohydrate—lactose (g) | 3.66 | 3.14 | 1.44 | 8.61 | 2.25 | 0.75 | 6.61 | 3.54 | 0.96 | 45.08 | 10.62 | 2.58 | 5.51 | 5.28 | 1.18 | 10.34 | 2.98 | 0.99 |
| Carbohydrate—maltose (g) | 1.57 | 1.23 | 0.14 | 3.38 | 1.07 | 0.36 | 3.35 | 1.617 | 0.166 | 24.30 | 5.66 | 1.37 | 3.85 | 2.54 | 0.84 | 11.90 | 3.53 | 1.18 |
| Magnesium (mg) | 239.46 | 248.76 | 132.02 | 340.09 | 58.93 | 19.64 | 353.51 | 283.81 | 146.11 | 1139.36 | 238.08 | 57.74 | 254.28 | 186.78 | 100.45 | 687.19 | 183.24 | 61.08 |
| Manganese (mg) | 3.01 | 2.82 | 1.43 | 4.37 | 0.95 | 0.32 | 4.00 | 2.785 | 1.39 | 14.70 | 3.35 | 0.81 | 3.21 | 2.50 | 0.71 | 11.01 | 3.07 | 1.02 |
| Sodium (mg) | 2488.07 | 2642.56 | 1178.08 | 3557.61 | 863.31 | 287.77 | 3376.43 | 2492.29 | 1631.38 | 13,993.4 | 2897.73 | 702.80 | 2388.65 | 1677.65 | 1106.84 | 5050.05 | 1536.96 | 512.32 |
| Niacin (mg) | 21.88 | 24.37 | 13.55 | 27.37 | 5.14 | 1.71 | 31.39 | 25.08 | 12.45 | 81.92 | 17.30 | 4.20 | 18.68 | 14.39 | 9.13 | 39.30 | 10.06 | 3.35 |
| Phosphorus (mg) | 1097.58 | 1109.68 | 560.59 | 1730.58 | 347.45 | 115.82 | 1667.92 | 1158.57 | 800.31 | 6624.44 | 1375.70 | 333.66 | 1051.78 | 859.97 | 528.78 | 1872.52 | 504.47 | 168.16 |
| Protein (g) | 75.09 | 75.34 | 44.90 | 117.93 | 23.66 | 7.89 | 111.66 | 85.30 | 53.46 | 363.30 | 74.73 | 18.12 | 68.26 | 55.70 | 38.75 | 115.37 | 29.67 | 9.89 |
| Vitamin A—retinol (mcg) | 1427.28 | 926.11 | 168.39 | 5250.17 | 1522.96 | 507.65 | 2065.08 | 1076.95 | 54.90 | 14,070.9 | 3473.56 | 842.46 | 698.10 | 292.58 | 176.59 | 1738.48 | 694.60 | 231.53 |
| Vitamin A—retinol equivalents (mcg) | 1736.80 | 1381.10 | 511.82 | 5370.13 | 1450.29 | 483.43 | 2501.95 | 1674.59 | 161.84 | 14,829.7 | 3546.31 | 860.11 | 1022.37 | 553.03 | 253.20 | 2672.78 | 891.64 | 297.21 |
| Vitamin B2—riboflavin (mg) | 1.44 | 1.44 | 0.71 | 2.23 | 0.50 | 0.17 | 1.99 | 1.22 | 0.86 | 8.31 | 1.80 | 0.44 | 1.10 | 0.82 | 0.61 | 2.09 | 0.56 | 0.19 |
| Selenium (mcg) | 70.61 | 78.64 | 31.36 | 91.94 | 20.04 | 6.68 | 85.26 | 73.02 | 37.65 | 263.07 | 54.23 | 13.15 | 56.48 | 49.12 | 29.95 | 104.80 | 28.18 | 9.39 |
| Carbohydrate—starch (g) | 89.89 | 87.31 | 40.61 | 143.99 | 33.11 | 11.04 | 150.15 | 123.03 | 13.84 | 800.47 | 174.38 | 42.29 | 113.25 | 87.67 | 42.84 | 221.95 | 60.84 | 20.28 |
| Carbohydrate—sucrose (g) | 32.94 | 29.84 | 12.11 | 62.12 | 15.25 | 5.08 | 70.33 | 31.15 | 12.13 | 605.67 | 139.84 | 33.92 | 76.47 | 39.35 | 19.77 | 219.10 | 74.32 | 24.78 |
| Vitamin B1—thiamin (mg) | 1.27 | 1.16 | 0.74 | 1.95 | 0.41 | 0.14 | 142.55 | 1.69 | 0.76 | 2395.00 | 580.44 | 140.78 | 1.12 | 0.87 | 0.47 | 2.13 | 0.59 | 0.20 |
| Nitrogen (g) | 12.14 | 12.17 | 7.23 | 19.00 | 3.82 | 1.28 | 18.18 | 13.76 | 8.62 | 59.56 | 12.26 | 2.97 | 11.11 | 9.09 | 6.36 | 19.16 | 4.88 | 1.63 |
| Carbohydrate—sugars (total) (g) | 88.30 | 75.76 | 46.22 | 143.32 | 37.07 | 12.36 | 141.13 | 78.11 | 33.95 | 939.79 | 212.75 | 51.60 | 132.48 | 81.78 | 36.00 | 404.63 | 122.91 | 40.97 |
| Vitamin B12—cobalamin (mcg) | 7.38 | 7.07 | 1.45 | 19.61 | 5.39 | 1.80 | 10.60 | 6.318 | 1.71 | 52.37 | 12.09 | 2.93 | 4.38 | 2.98 | 1.65 | 8.41 | 2.64 | 0.88 |
| Vitamin B6—pyridoxine (mg) | 1.60 | 1.49 | 1.02 | 2.31 | 0.44 | 0.15 | 2.40 | 1.80 | 1.08 | 7.14 | 1.49 | 0.36 | 1.63 | 1.39 | 0.66 | 3.89 | 1.02 | 0.34 |
| Vitamin C—ascorbic acid (mg) | 86.22 | 76.94 | 45.63 | 147.31 | 36.58 | 12.19 | 164.62 | 77.91 | 53.70 | 1111.13 | 254.00 | 61.60 | 83.55 | 49.14 | 18.41 | 289.33 | 88.00 | 29.33 |
| Vitamin D—ergocalciferol (mcg) | 2.33 | 2.60 | 0.81 | 3.81 | 1.17 | 0.39 | 3.69 | 2.11 | 1.35 | 22.85 | 5.07 | 1.23 | 3.17 | 2.28 | 1.40 | 9.05 | 2.40 | 0.80 |
| Vitamin E—alpha tocopherol equivalents (mg) | 7.73 | 7.84 | 3.95 | 10.83 | 2.43 | 0.81 | 14.10 | 9.99 | 4.21 | 75.19 | 16.19 | 3.93 | 10.90 | 7.42 | 3.34 | 33.66 | 9.61 | 3.20 |
| Zinc (mg) | 8.37 | 8.28 | 4.51 | 14.26 | 2.77 | 0.92 | 12.52 | 9.25 | 6.01 | 40.46 | 8.28 | 2.00 | 7.35 | 5.94 | 3.75 | 12.70 | 3.38 | 1.13 |
| Fat—total (g) | 48.55 | 45.72 | 22.78 | 86.31 | 20.41 | 6.80 | 91.93 | 63.09 | 29.39 | 513.83 | 112.25 | 27.23 | 74.90 | 56.78 | 32.22 | 191.07 | 50.06 | 16.69 |
| Monounsaturated fatty acids (MUFA—total) (g) | 17.49 | 15.87 | 8.11 | 28.68 | 7.29 | 2.43 | 36.54 | 23.69 | 8.86 | 210.71 | 46.88 | 11.37 | 30.15 | 22.04 | 13.06 | 87.17 | 23.44 | 7.81 |
| Polyunsaturated fatty acids (PUFA—total) (g) | 8.32 | 7.89 | 4.29 | 13.60 | 3.16 | 1.05 | 14.63 | 9.65 | 6.08 | 68.10 | 14.63 | 3.55 | 10.29 | 6.90 | 3.60 | 32.18 | 8.78 | 2.93 |
| Saturated fatty acids (SFA—total) (g) | 17.75 | 16.40 | 7.57 | 39.98 | 9.58 | 3.19 | 32.17 | 19.88 | 8.57 | 186.60 | 40.75 | 9.88 | 27.59 | 24.29 | 10.79 | 54.82 | 14.42 | 4.81 |
| Alcoholic beverages (g) | 20.40 | 3.50 | 0.00 | 125.50 | 40.51 | 13.50 | 52.01 | 35.63 | 0.00 | 316.00 | 76.18 | 18.48 | 27.37 | 3.50 | 0.00 | 138.00 | 46.22 | 15.41 |
| Cereals and cereal products (g) | 190.89 | 178.11 | 66.67 | 307.88 | 77.09 | 25.70 | 360.09 | 177.23 | 0.56 | 2585.28 | 591.43 | 143.44 | 266.67 | 219.76 | 93.24 | 643.45 | 176.93 | 58.98 |
| Eggs and egg dishes (g) | 14.67 | 7.00 | 3.50 | 39.50 | 12.16 | 4.05 | 16.35 | 17.50 | 0.00 | 50.00 | 13.60 | 3.30 | 16.22 | 17.50 | 3.50 | 50.00 | 15.30 | 5.10 |
| Fats and oils (g) | 8.43 | 6.79 | 1.82 | 17.42 | 5.67 | 1.89 | 16.08 | 5.60 | 0.21 | 177.12 | 41.83 | 10.15 | 20.99 | 6.51 | 4.69 | 91.82 | 27.91 | 9.30 |
| Fish and fish products (g) | 18.40 | 16.24 | 0.00 | 46.48 | 16.11 | 5.37 | 33.92 | 27.23 | 4.20 | 101.00 | 24.78 | 6.01 | 23.69 | 19.32 | 8.12 | 47.46 | 14.26 | 4.75 |
| Fruit (g) | 179.28 | 191.55 | 77.25 | 290.05 | 78.72 | 26.24 | 186.10 | 135.20 | 47.95 | 1047.35 | 228.21 | 55.35 | 243.75 | 67.90 | 17.50 | 1477.80 | 468.90 | 156.30 |
| Meat and meat products (g) | 136.46 | 142.58 | 83.52 | 183.89 | 41.33 | 13.78 | 215.23 | 191.01 | 69.05 | 533.97 | 121.41 | 29.45 | 108.71 | 100.55 | 50.61 | 156.00 | 32.42 | 10.81 |
| Milk and milk products (g) | 96.20 | 90.26 | 22.82 | 182.64 | 46.81 | 15.60 | 126.84 | 89.28 | 4.06 | 423.20 | 118.47 | 28.73 | 67.40 | 57.52 | 14.98 | 165.60 | 54.06 | 18.02 |
| Non-alcoholic beverages (g) | 758.66 | 656.30 | 196.30 | 1494.14 | 445.84 | 148.61 | 588.49 | 503.00 | 9.48 | 1752.20 | 460.08 | 111.59 | 614.78 | 477.80 | 166.20 | 1748.64 | 488.15 | 162.72 |
| Nuts and seeds (g) | 3.60 | 2.10 | 0.00 | 12.90 | 3.89 | 1.30 | 23.55 | 2.10 | 0.00 | 137.66 | 39.98 | 9.70 | 11.23 | 2.10 | 0.00 | 75.00 | 24.22 | 8.07 |
| Potatoes (g) | 59.48 | 59.08 | 17.50 | 128.36 | 33.49 | 11.16 | 125.10 | 83.36 | 0.00 | 392.69 | 113.43 | 27.51 | 93.20 | 83.65 | 26.39 | 207.83 | 53.72 | 17.91 |
| Soups and sauces (g) | 131.71 | 116.10 | 44.80 | 230.10 | 65.55 | 21.85 | 85.88 | 72.50 | 0.00 | 280.40 | 62.23 | 15.09 | 102.51 | 32.20 | 23.80 | 377.90 | 128.17 | 42.72 |
| Sugars (g) | 34.21 | 21.44 | 3.92 | 133.96 | 40.57 | 13.53 | 68.07 | 39.22 | 0.00 | 530.50 | 124.68 | 30.24 | 84.49 | 76.37 | 22.90 | 254.77 | 72.12 | 24.04 |
| preserves and snacks (g) | 233.87 | 224.45 | 106.01 | 383.01 | 105.59 | 35.20 | 270.98 | 218.28 | 20.64 | 747.81 | 182.05 | 44.15 | 187.66 | 174.48 | 34.86 | 430.16 | 143.55 | 47.85 |
| Vegetables | 2.78 | 5.00 | 0.00 | 5.00 | 2.64 | 0.88 | 4.71 | 5.00 | 0.00 | 5.00 | 1.21 | 0.29 | 5.00 | 5.00 | 5.00 | 5.00 | 0.00 | 0.00 |
| DII | 1.88 | 2.07 | −1.24 | 4.71 | 1.86 | 0.62 | 1.44 | 2.01 | −3.077 | 6.52 | 2.25 | 0.55 | 2.62 | 3.18 | −2.80 | 6.00 | 2.90 | 0.97 |
| hs troponin I | 4.79 | 2.70 | 1.30 | 14.10 | 4.43 | 1.48 | 1.94 | 1.80 | 0.10 | 7.80 | 1.86 | 0.45 | 1.02 | 0.80 | 0.30 | 2.20 | 0.59 | 0.20 |
| TC | 5.49 | 4.60 | 3.50 | 10.10 | 2.19 | 0.73 | 6.01 | 5.90 | 3.90 | 8.40 | 1.24 | 0.30 | 5.64 | 6.00 | 3.20 | 7.80 | 1.55 | 0.52 |
| HDL-C | 1.13 | 1.10 | 0.80 | 1.50 | 0.26 | 0.09 | 1.19 | 1.20 | 0.80 | 1.80 | 0.29 | 0.07 | 1.21 | 1.30 | 0.90 | 1.50 | 0.20 | 0.07 |
| LDL-C | 3.54 | 2.60 | 2.20 | 7.70 | 1.82 | 0.61 | 3.84 | 3.80 | 2.20 | 6.00 | 1.01 | 0.24 | 3.67 | 4.10 | 2.00 | 5.40 | 1.29 | 0.43 |
| TG | 1.93 | 1.80 | 0.80 | 3.70 | 1.01 | 0.34 | 2.31 | 2.40 | 0.80 | 3.60 | 0.64 | 0.16 | 1.87 | 1.50 | 0.80 | 4.60 | 1.16 | 0.39 |
| TSH | 1.76 | 1.74 | 0.88 | 2.77 | 0.61 | 0.19 | 2.27 | 2.06 | 0.75 | 5.74 | 1.19 | 0.31 | 3.31 | 3.00 | 2.05 | 4.79 | 1.02 | 0.32 |
| fT4 | 13.20 | 13.90 | 9.48 | 15.84 | 2.01 | 0.67 | 12.25 | 12.04 | 10.19 | 14.68 | 1.51 | 0.37 | 11.79 | 12.07 | 9.32 | 13.98 | 1.40 | 0.47 |
| fT3 | 3.94 | 4.21 | 2.71 | 4.97 | 0.69 | 0.23 | 4.31 | 4.27 | 3.43 | 5.59 | 0.55 | 0.13 | 4.68 | 4.64 | 3.75 | 5.57 | 0.65 | 0.22 |
| Insulin | 18.91 | 16.30 | 4.30 | 31.20 | 9.83 | 3.28 | 17.26 | 15.30 | 4.50 | 56.30 | 11.14 | 2.70 | 18.66 | 18.80 | 6.80 | 31.40 | 8.58 | 2.86 |
| HOMA-IR | 17.90 | 6.50 | 1.70 | 113.40 | 35.91 | 11.9710 | 34.89 | 33.40 | 1.40 | 127.21 | 32.22 | 7.81 | 43.62 | 35.76 | 11.61 | 87.12 | 26.29 | 8.77 |
| Weight | 135.48 | 130.70 | 119.40 | 154.70 | 12.23 | 4.08 | 125.80 | 125.60 | 95.90 | 183.60 | 24.20 | 5.87 | 123.91 | 119.80 | 104.70 | 145.10 | 12.66 | 4.22 |
| SMM | 41.23 | 40.30 | 33.50 | 52.20 | 7.08 | 2.36 | 38.17 | 36.50 | 26.90 | 54.20 | 9.05 | 2.20 | 36.13 | 32.90 | 30.30 | 56.20 | 8.40 | 2.80 |
| BFM | 62.87 | 64.30 | 43.80 | 78.40 | 11.89 | 3.96 | 58.61 | 55.90 | 40.40 | 90.30 | 13.22 | 3.21 | 60.26 | 62.00 | 42.20 | 71.40 | 10.12 | 3.37 |
| PBF% | 45.30 | 43.90 | 34.30 | 55.90 | 7.19 | 2.40 | 47.64 | 49.40 | 33.50 | 64.50 | 7.14 | 1.73 | 48.83 | 51.00 | 30.30 | 55.10 | 7.75 | 2.58 |
| BMI | 47.64 | 49.80 | 36.40 | 57.50 | 6.97 | 2.32 | 42.21 | 41.70 | 35.40 | 54.80 | 5.69 | 1.38 | 42.90 | 42.40 | 37.70 | 49.20 | 3.53 | 1.18 |
| BMR | 1938.22 | 1903.00 | 1648.00 | 2341.00 | 255.32 | 85.11 | 1821.35 | 1768.00 | 1418.00 | 2385.00 | 321.52 | 77.99 | 1745.11 | 1635.00 | 1532.00 | 2462.00 | 300.39 | 100.13 |
| W-H ratio | 1.05 | 1.04 | 0.84 | 1.17 | 0.12 | 0.04 | 1.09 | 1.07 | 0.92 | 1.27 | 0.08 | 0.02 | 1.06 | 1.06 | 0.93 | 1.22 | 0.09 | 0.03 |
| VFL | 19.44 | 20.00 | 16.00 | 20.00 | 1.33 | 0.44 | 19.94 | 20.00 | 19.00 | 20.00 | 0.24 | 0.06 | 19.89 | 20.00 | 19.00 | 20.00 | 0.33 | 0.11 |
| M1 | 680.22 | 788.00 | 197.00 | 890.00 | 238.42 | 79.47 | 503.94 | 423.00 | 186.00 | 883.00 | 257.74 | 62.51 | 331.89 | 255.00 | 223.00 | 633.00 | 156.68 | 52.23 |
| M2 | 248.56 | 177.00 | 102.00 | 504.00 | 143.89 | 47.96 | 330.18 | 357.00 | 113.00 | 569.00 | 152.32 | 36.94 | 412.44 | 410.00 | 249.00 | 575.00 | 111.31 | 37.10 |
| M3 | 22.78 | 10.00 | 1.00 | 85.00 | 28.87 | 9.62 | 60.82 | 54.00 | 2.00 | 128.00 | 40.05 | 9.71 | 91.22 | 87.00 | 40.00 | 140.00 | 32.34 | 10.78 |
| M4 | 46.11 | 2.00 | 0.00 | 214.00 | 82.86 | 27.62 | 105.06 | 80.00 | 0.00 | 302.00 | 97.15 | 23.56 | 164.44 | 184.00 | 24.00 | 285.00 | 92.40 | 30.80 |
| NDI | 1.43 | 1.21 | 1.11 | 2.32 | 0.43 | 0.14 | 1.77 | 1.76 | 1.12 | 2.55 | 0.47 | 0.12 | 2.09 | 2.22 | 1.54 | 2.46 | 0.34 | 0.11 |
| FREQ MN TOTAL | 9.94 | 10.00 | 4.00 | 19.00 | 4.72 | 1.57 | 8.21 | 9.00 | 1.50 | 14.50 | 3.87 | 0.94 | 4.78 | 5.00 | 2.00 | 7.00 | 1.73 | 0.58 |
| FREQ NB TOTAL | 3.83 | 2.00 | 0.00 | 18.00 | 5.42 | 1.81 | 5.56 | 5.00 | 1.00 | 13.00 | 3.80 | 0.920 | 5.39 | 4.00 | 1.50 | 14.00 | 4.10 | 1.37 |
| FREQ NPB TOTAL | 5.61 | 4.50 | 1.50 | 14.50 | 4.05 | 1.35 | 6.41 | 5.50 | 1.00 | 17.00 | 5.02 | 1.22 | 3.78 | 3.00 | 2.00 | 10.00 | 2.46 | 0.82 |
| APOPTOSIS | 11.44 | 10.00 | 1.00 | 42.00 | 12.13 | 4.04 | 6.82 | 5.00 | 2.00 | 18.00 | 4.38 | 1.06 | 5.00 | 4.00 | 3.00 | 9.00 | 2.29 | 0.76 |
| NECROSIS | 2.56 | 0.00 | 0.00 | 23.00 | 7.67 | 2.56 | 2.65 | 0.00 | 0.00 | 29.00 | 7.33 | 1.78 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Net Fpg, TI | 5.55 | 3.03 | 0.00 | 15.06 | 5.55 | 1.85 | 5.22 | 3.14 | 0.23 | 13.01 | 3.97 | 0.96 | 6.63 | 5.30 | 1.41 | 13.50 | 4.37 | 1.46 |
| TI | 12.02 | 10.53 | 6.65 | 17.72 | 3.84 | 1.28 | 10.28 | 10.04 | 4.61 | 17.88 | 4.12 | 0.998 | 8.97 | 8.52 | 4.50 | 15.93 | 3.58 | 1.193 |
| Age | 55.69 | 56.00 | 51.00 | 59.00 | 2.76 | 0.92 | 45.43 | 45.80 | 40.26 | 51.41 | 3.58 | 0.869 | 32.95 | 35.31 | 26.35 | 38.73 | 4.95 | 1.65 |
| DNA Damage Parameters | T/NT | Mean | Median | Min | Max | SD | SE |
|---|---|---|---|---|---|---|---|
| MN | NonT | 8.76 | 7.00 | 1.50 | 24.00 | 5.09 | 0.76 |
| T | 10.38 | 9.75 | 3.00 | 17.50 | 5.30 | 1.88 | |
| NB | NonT | 6.03 | 4.00 | 0.00 | 21.00 | 5.29 | 0.79 |
| T | 3.19 | 2.00 | 0.50 | 11.00 | 3.40 | 1.20 | |
| NPB | NonT | 6.02 | 4.50 | 0.50 | 17.00 | 4.44 | 0.66 |
| T | 4.13 | 3.00 | 1.00 | 10.50 | 3.23 | 1.14 | |
| APOPTOSIS | NonT | 7.84 | 7.00 | 1.00 | 42.00 | 6.88 | 1.02 |
| T | 8.25 | 7.00 | 0.00 | 26.00 | 8.29 | 2.93 | |
| NECROSIS | NonT | 2.44 | 0.00 | 0.00 | 33.00 | 7.39 | 1.10 |
| T | 4.75 | 1.00 | 0.00 | 23.00 | 8.01 | 2.83 | |
| Net Fpg, TI, % | NonT | 6.45 | 4.73 | 0.00 | 31.59 | 5.94 | 0.89 |
| T | 5.29 | 2.57 | 0.00 | 24.06 | 7.93 | 2.80 | |
| TI, % | NonT | 10.32 | 9.98 | 4.50 | 17.88 | 3.83 | 0.57 |
| T | 11.56 | 12.74 | 4.68 | 18.27 | 5.02 | 1.77 | |
| AGE, years | NonT | 49.68 | 50.00 | 26.35 | 68.00 | 11.39 | 1.70 |
| T | 59.42 | 64.00 | 41.00 | 67.00 | 9.47 | 3.35 | |
| MN | NT | 8.3 | 7.0 | 3.0 | 17.5 | 4.2 | 0.80 |
| FMT | 9.7 | 9.3 | 1.5 | 24.0 | 5.9 | 1.16 | |
| NB | NT | 4.7 | 3.0 | 0.5 | 20.5 | 4.4 | 0.85 |
| FMT | 6.5 | 4.5 | 0.0 | 21.0 | 5.7 | 1.12 | |
| NPB | NT | 5.5 | 3.5 | 0.5 | 16.0 | 4.3 | 0.83 |
| FMT | 6.0 | 4.8 | 1.0 | 17.0 | 4.4 | 0.85 | |
| APOPTOSIS | NT | 7.4 | 6.0 | 2.0 | 26.0 | 5.4 | 1.05 |
| FMT | 8.5 | 7.0 | 0.0 | 42.0 | 8.4 | 1.65 | |
| NECROSIS | NT | 2.7 | 0.0 | 0.0 | 33.0 | 7.8 | 1.49 |
| FMT | 2.9 | 0.0 | 0.0 | 29.0 | 7.3 | 1.43 | |
| Net Fpg, TI, % | NT | 6.0 | 3.0 | 0.0 | 31.6 | 6.6 | 1.28 |
| FMT | 6.5 | 5.0 | 0.0 | 24.1 | 5.8 | 1.15 | |
| TI, % | NT | 10.0 | 9.6 | 4.5 | 17.9 | 3.6 | 0.70 |
| FMT | 11.0 | 11.6 | 4.6 | 18.3 | 4.4 | 0.86 | |
| AGE | NT | 48.9 | 49.0 | 26.4 | 68.0 | 12.5 | 2.41 |
| FMT | 53.5 | 56.5 | 28.9 | 67.0 | 10.3 | 2.01 |
| Individuals with previous tumors | |||||
| Individual No. | >9% TI | >12.4% TI | >13 MN | >10 NPB | >5 NB |
| 1 | + | + | + | ||
| 2 | + | + | |||
| 3 | + | + | |||
| 4 | + | + | |||
| 5 | + | + | |||
| 6 | + | + | |||
| 7 | + | + | |||
| 8 | + | ||||
| Individuals with family history of tumors | |||||
| Individual No. | >9% TI | >12.4% TI | >13 MN | >10 NPB | >5 NB |
| 1 | |||||
| 2 | + | + | + | ||
| 3 | + | + | |||
| 4 | + | + | + | ||
| 5 | + | + | + | ||
| 6 | + | + | |||
| 7 | + | + | |||
| 8 | + | + | |||
| 9 | + | + | |||
| 10 | + | + | + | + | |
| 11 | + | + | |||
| 12 | + | + | + | ||
| 13 | + | + | + | ||
| 14 | + | + | + | + | |
| 15 | + | + | |||
| 16 | + | + | + | + | |
| 17 | + | ||||
| 18 | + | + | + | + | |
| 19 | + | + | |||
| 20 | |||||
| 21 | + | + | |||
| 22 | + | ||||
| 23 | + | + | |||
| 24 | + | ||||
| 25 | + | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Milić, M.; Ožvald, I.; Matković, K.; Radašević, H.; Nikolić, M.; Božičević, D.; Duh, L.; Matovinović, M.; Bituh, M. Combined Approach: FFQ, DII, Anthropometric, Biochemical and DNA Damage Parameters in Obese with BMI ≥ 35 kg m−2. Nutrients 2023, 15, 899. https://doi.org/10.3390/nu15040899
Milić M, Ožvald I, Matković K, Radašević H, Nikolić M, Božičević D, Duh L, Matovinović M, Bituh M. Combined Approach: FFQ, DII, Anthropometric, Biochemical and DNA Damage Parameters in Obese with BMI ≥ 35 kg m−2. Nutrients. 2023; 15(4):899. https://doi.org/10.3390/nu15040899
Chicago/Turabian StyleMilić, Mirta, Ivan Ožvald, Katarina Matković, Hrvoje Radašević, Maja Nikolić, Dragan Božičević, Lidija Duh, Martina Matovinović, and Martina Bituh. 2023. "Combined Approach: FFQ, DII, Anthropometric, Biochemical and DNA Damage Parameters in Obese with BMI ≥ 35 kg m−2" Nutrients 15, no. 4: 899. https://doi.org/10.3390/nu15040899
APA StyleMilić, M., Ožvald, I., Matković, K., Radašević, H., Nikolić, M., Božičević, D., Duh, L., Matovinović, M., & Bituh, M. (2023). Combined Approach: FFQ, DII, Anthropometric, Biochemical and DNA Damage Parameters in Obese with BMI ≥ 35 kg m−2. Nutrients, 15(4), 899. https://doi.org/10.3390/nu15040899