
New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food


 , ,
, ,  , ,
, ,  ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Metabolic Syndrome
2.1. Past-Current Definition and Classification
- ▪
- High waist circumference (WC), whose thresholds depend on populations and country-specific definitions (≥102 cm and ≥88 cm for European men and women respectively) [9];
- ▪
- Blood TG ≥ 150 mg/dL;
- ▪
- Blood HDL cholesterol < 40 mg/dL in men and <50 mg/dL in women;
- ▪
- Blood pressure (BP) ≥ 130/85 mmHg;
- ▪
- Blood fasting glucose ≥ 100 mg/dL.
2.2. Pathophysiology
2.3. MetS Comorbidities and Complications
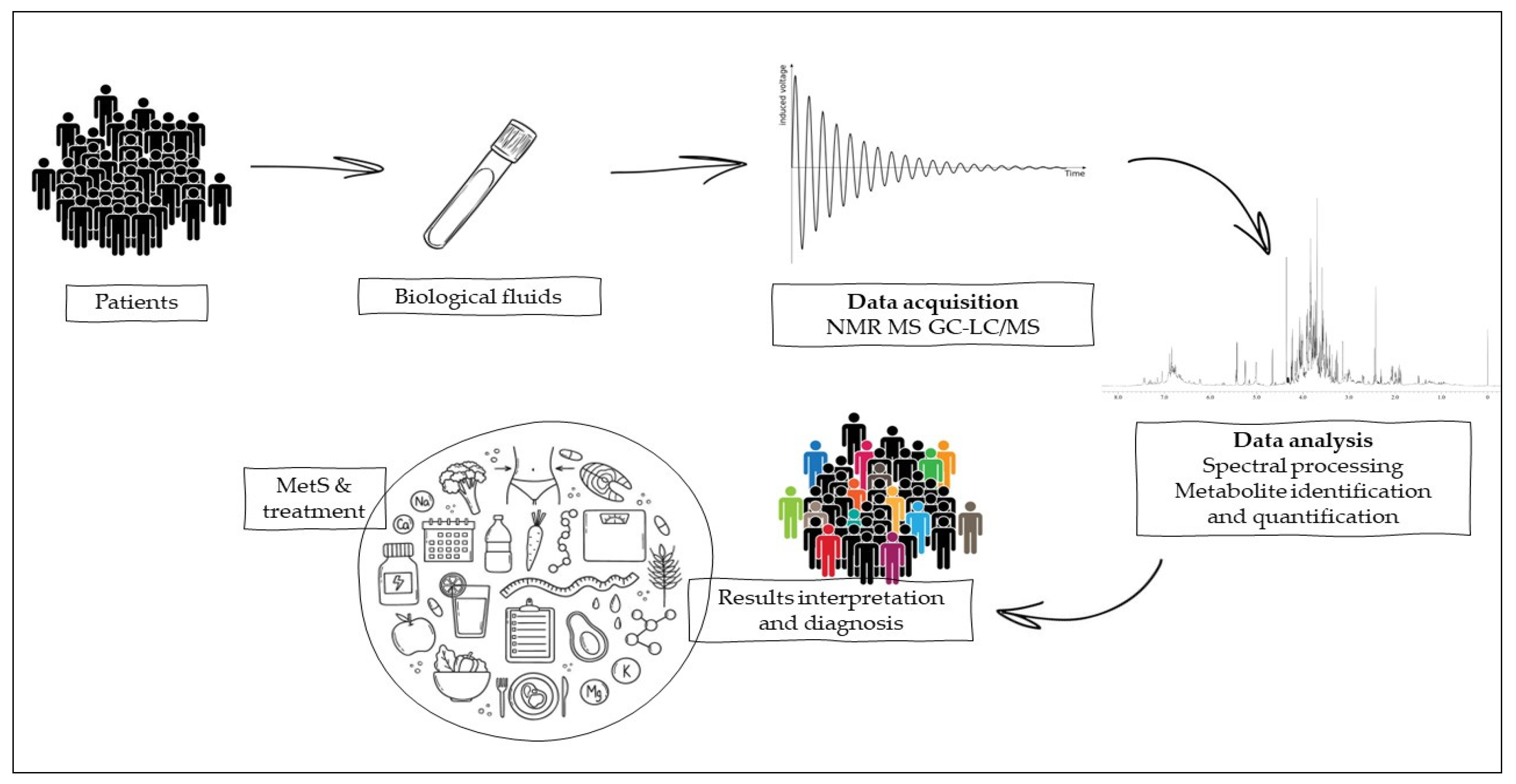
3. Methodologies for Timely Prediction and Diagnosis of MetS
3.1. Nuclear Magnetic Resonance
3.2. MS-Chromatographic Techniques
3.3. Metabolic Syndrome Management, Interventions, and Challenges
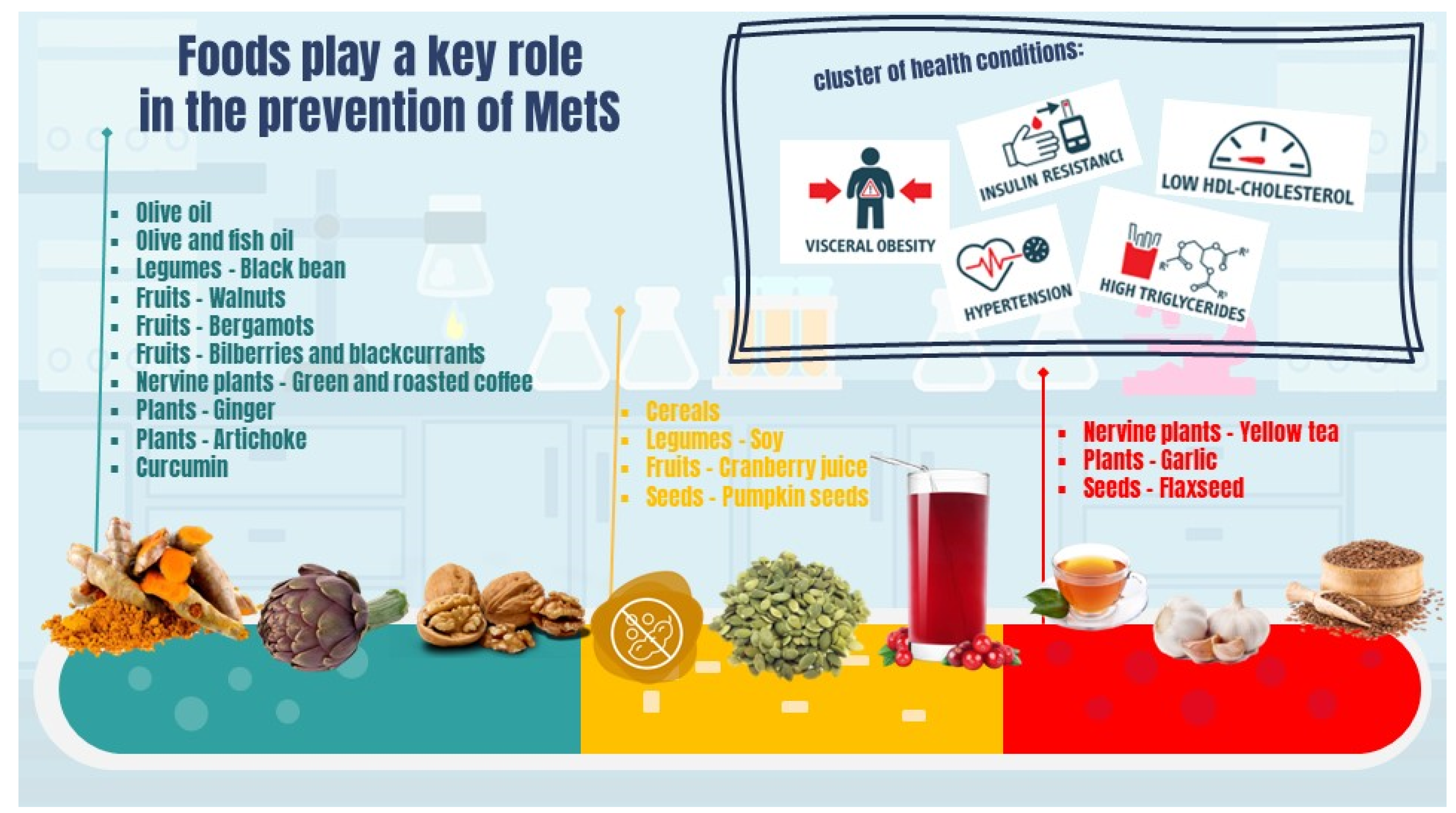
4. New Aspects Implicated in the Prevention and Treatment of MetS
4.1. Gender Medicine and Metabolomics
4.2. Dietary Patterns
5. Food and Foods Components
5.1. Seeds
5.2. Plants
5.3. Nervine Plants
5.4. Fruits
5.5. Legumes
5.6. Cereals
5.7. Olive Oil
5.8. Omega 3 Long-Chain Polyunsaturated Fatty Acids and Fish Products
5.9. Polyphenols
5.9.1. Flavonoids
5.9.2. Chlorogenic Acid
5.10. Curcumin
5.11. Prebiotics and Probiotics
6. Microbiota and Nutrigenetics
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Levesque, J.; Lamarche, B. The Metabolic Syndrome: Definitions, Prevalence and Management. J. Nutr. Nutr. 2008, 1, 100–108. [Google Scholar] [CrossRef]
- Sarafidis, P.A.; Nilsson, P.M. The Metabolic Syndrome: A Glance at Its History. J. Hypertens. 2006, 24, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M. Role of Insulin Resistance in Human Disease. Diabetes 1988, 37, 1595–1607. [Google Scholar] [CrossRef]
- DeFronzo, R.A.; Ferrannini, E. Insulin Resistance: A Multifaceted Syndrome Responsible for NIDDM, Obesity, Hypertension, Dyslipidemia, and Atherosclerotic Cardiovascular Disease. Diabetes Care 1991, 14, 173–194. [Google Scholar] [CrossRef]
- Kaplan, N.M. The Deadly Quartet. Upper-Body Obesity, Glucose Intolerance, Hypertriglyceridemia, and Hypertension. Arch. Intern. Med. 1989, 149, 1514–1520. [Google Scholar] [CrossRef] [PubMed]
- Pasternak, R.C. Report of the Adult Treatment Panel III: The 2001 National Cholesterol Education Program Guidelines on the Detection, Evaluation and Treatment of Elevated Cholesterol in Adults. Cardiol. Clin. 2003, 21, 393–398. [Google Scholar] [CrossRef]
- Alberti, K.G.M.M.; Zimmet, P.Z. Definition, Diagnosis and Classification of Diabetes Mellitus and Its Complications. Part 1: Diagnosis and Classification of Diabetes Mellitus. Provisional Report of a WHO Consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Balkau, B.; Charles, M.A. Comment on the Provisional Report from the WHO Consultation. European Group for the Study of Insulin Resistance (EGIR). Diabet Med. 1999, 16, 442–443. [Google Scholar] [PubMed]
- Alberti, K.G.M.M.; Eckel, R.H.; Grundy, S.M.; Zimmet, P.Z.; Cleeman, J.I.; Donato, K.A.; Fruchart, J.C.; James, W.P.T.; Loria, C.M.; Smith, S.C. Harmonizing the Metabolic Syndrome: A Joint Interim Statement of the International Diabetes Federation Task Force on Epidemiology and Prevention; National Heart, Lung, and Blood Institute; American Heart Association; World Heart Federation; International Atherosclerosis Society; And International Association for the Study of Obesity. Circulation 2009, 120, 1640–1645. [Google Scholar]
- Kassi, E.; Pervanidou, P.; Kaltsas, G.; Chrousos, G. Metabolic Syndrome: Definitions and Controversies. BMC Med. 2011, 9, 48. [Google Scholar] [CrossRef]
- Bovolini, A.; Garcia, J.; Andrade, M.A.; Duarte, J.A. Metabolic Syndrome Pathophysiology and Predisposing Factors. Int. J. Sports Med. 2021, 42, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Hull, R.L.; Utzschneider, K.M. Mechanisms Linking Obesity to Insulin Resistance and Type 2 Diabetes. Nature 2006, 444, 840–846. [Google Scholar] [CrossRef]
- Debnath, M.; Agrawal, S.; Agrawal, A.; Dubey, G.P. Metaflammatory Responses during Obesity: Pathomechanism and Treatment. Obes. Res. Clin. Pract. 2016, 10, 103–113. [Google Scholar] [CrossRef]
- Chait, A.; den Hartigh, L.J. Adipose Tissue Distribution, Inflammation and Its Metabolic Consequences, Including Diabetes and Cardiovascular Disease. Front. Cardiovasc. Med. 2020, 7, 22. [Google Scholar] [CrossRef]
- Rochlani, Y.; Pothineni, N.V.; Kovelamudi, S.; Mehta, J.L. Metabolic Syndrome: Pathophysiology, Management, and Modulation by Natural Compounds. Ther. Adv. Cardiovasc. Dis. 2017, 11, 215–225. [Google Scholar] [CrossRef]
- Boden, G.; Shulman, G.I. Free Fatty Acids in Obesity and Type 2 Diabetes: Defining Their Role in the Development of Insulin Resistance and β-Cell Dysfunction. Eur. J. Clin. Invest. 2002, 32, 14–23. [Google Scholar] [CrossRef]
- McCracken, E.; Monaghan, M.; Sreenivasan, S. Pathophysiology of the Metabolic Syndrome. Clin. Dermatol. 2018, 36, 14–20. [Google Scholar] [CrossRef]
- Li, X.; Zhai, Y.; Zhao, J.; He, H.; Li, Y.; Liu, Y.; Feng, A.; Li, L.; Huang, T.; Xu, A.; et al. Impact of Metabolic Syndrome and It’s Components on Prognosis in Patients With Cardiovascular Diseases: A Meta-Analysis. Front. Cardiovasc. Med. 2021, 8, 704145. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Cleeman, J.I.; Bairey Merz, C.N.; Brewer, H.B.; Clark, L.T.; Hunninghake, D.B.; Pasternak, R.C.; Smith, S.C.; Stone, N.J. Implications of Recent Clinical Trials for the National Cholesterol Education Program Adult Treatment Panel III Guidelines. Circulation 2004, 110, 227–239. [Google Scholar] [CrossRef] [PubMed]
- IDF. IDF Diabetes Atlas, 9th ed.; IDF: Brussels, Belgium, 2019. [Google Scholar]
- American Diabetes Association. 2. Classification and Diagnosis of Diabetes: Standards of Medical Care in Diabetes-2021. Diabetes Care 2021, 44, S15–S33. [Google Scholar] [CrossRef]
- Poggiogalle, E.; Lubrano, C.; Sergi, G.; Coin, A.; Gnessi, L.; Mariani, S.; Lenzi, A.; Donini, L.M. Sarcopenic Obesity and Metabolic Syndrome in Adult Caucasian Subjects. J. Nutr. Health Aging 2016, 20, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Gaines, J.; Vgontzas, A.N.; Fernandez-Mendoza, J.; Bixler, E.O. Obstructive Sleep Apnea and the Metabolic Syndrome: The Road to Clinically-Meaningful Phenotyping, Improved Prognosis, and Personalized Treatment. Sleep Med. Rev. 2018, 42, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Wluka, A.; Cicuttini, F.M.; Spector, T.D. Obesity, Arthritis, and Gout; CRC Press: New York, NY, USA, 1998. [Google Scholar]
- Vainio, H.; Kaaks, R.; Bianchini, F. Weight Control and Physical Activity in Cancer Prevention: International Evaluation of the Evidence. Eur. J. Cancer Prev. 2002, 11, S94–S100. [Google Scholar] [PubMed]
- Uzunlulu, M.; Telci Caklili, O.; Oguz, A. Association between Metabolic Syndrome and Cancer. Ann. Nutr. Metab. 2016, 68, 179–719. [Google Scholar] [CrossRef] [PubMed]
- Pujia, R.; Tarsitano, M.G.; Arturi, F.; de Lorenzo, A.; Lenzi, A.; Pujia, A.; Montalcini, T. Advances in Phenotyping Obesity and in Its Dietary and Pharmacological Treatment: A Narrative Review. Front. Nutr. 2022, 9, 804719. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.P.; de Souza Santos, R.; Palmer, B.F.; Clegg, D.J. Determinants of body fat distribution in humans may provide insight about obesity-related health risks. J. Lipid Res. 2019, 60, 1710–1719. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Magno, C.P.; Lane, K.T.; Hinojosa, M.W.; Lane, J.S. Association of Hypertension, Diabetes, Dyslipidemia, and Metabolic Syndrome with Obesity: Findings from the National Health and Nutrition Examination Survey, 1999 to 2004. J. Am. Coll. Surg. 2008, 207, 928–934. [Google Scholar] [CrossRef]
- Skurk, T.; Hauner, H. Obesity and Impaired Fibrinolysis: Role of Adipose Production of Plasminogen Activator Inhibitor-1. Int. J. Obes. 2004, 28, 1357–1364. [Google Scholar] [CrossRef]
- Williams, B.; Mancia, G.; Spiering, W.; Rosei, E.A.; Azizi, M.; Burnier, M.; Clement, D.; Coca, A.; de Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the Management of Arterial Hypertension. Eur. Heart J. 2018, 39, 3021–3104, Erratum in Eur. Heart J. 2019, 40, 475. [Google Scholar] [CrossRef]
- Catapano, A.L.; Reiner, Ž.; de Backer, G.; Graham, I.; Taskinen, M.R.; Wiklund, O.; Agewall, S.; Alegria, E.; Chapman, M.J.; Durrington, P.; et al. ESC/EAS Guidelines for the Management of Dyslipidaemias. The Task Force for the Management of Dyslipidaemias of the European Society of Cardiology (ESC) and the European Atherosclerosis Society (EAS). Atherosclerosis 2011, 217, 1769–1818. [Google Scholar]
- Atar, D.; Jukema, J.W.; Molemans, B.; Taub, P.R.; Goto, S.; Mach, F.; CerezoOlmos, C.; Underberg, J.; Keech, A.; Tokgözoğlu, L.; et al. New Cardiovascular Prevention Guidelines: How to Optimally Manage Dyslipidaemia and Cardiovascular Risk in 2021 in Patients Needing Secondary Prevention? Atherosclerosis 2021, 319, 51–61. [Google Scholar] [CrossRef]
- Friedman, S.L.; Neuschwander-Tetri, B.A.; Rinella, M.; Sanyal, A.J. Mechanisms of NAFLD Development and Therapeutic Strategies. Nat. Med. 2018, 24, 908–922. [Google Scholar] [CrossRef]
- Chan, K.L.; Cathomas, F.; Russo, S.J. Central and Peripheral Inflammation Link Metabolic Syndrome and Major Depressive Disorder. Physiology 2019, 34, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Salvio, G.; Ciarloni, A.; Cutini, M.; Delli Muti, N.; Finocchi, F.; Perrone, M.; Rossi, S.; Balercia, G. Metabolic Syndrome and Male Fertility: Beyond Heart Consequences of a Complex Cardiometabolic Endocrinopathy. Int. J. Mol. Sci. 2022, 23, 5497. [Google Scholar] [CrossRef]
- Sanchez-Garrido, M.A.; Tena-Sempere, M. Metabolic Dysfunction in Polycystic Ovary Syndrome: Pathogenic Role of Androgen Excess and Potential Therapeutic Strategies. Mol. Metab. 2020, 35, 100937. [Google Scholar] [CrossRef]
- Gatto, N.M.; Henderson, V.W.; St. John, J.A.; McCleary, C.; Hodis, H.N.; Mack, W.J. Metabolic Syndrome and Cognitive Function in Healthy Middle-Aged and Older Adults without Diabetes. Aging Neuropsychol. Cogn. 2008, 15, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Taylor, V.H.; MacQueen, G.M. Cognitive Dysfunction Associated with Metabolic Syndrome. Obes. Rev. 2007, 8, 409–418. [Google Scholar] [CrossRef]
- Gordon Smith, A. Impaired Glucose Tolerance and Metabolic Syndrome in Idiopathic Neuropathy. J. Peripher. Nerv. Syst. 2012, 17, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Pal, K.; Mukadam, N.; Petersen, I.; Cooper, C. Mild Cognitive Impairment and Progression to Dementia in People with Diabetes, Prediabetes and Metabolic Syndrome: A Systematic Review and Meta-Analysis. Soc. Psychiatry Psychiatr. Epidemiol. 2018, 53, 1149–1160. [Google Scholar] [CrossRef]
- Nishikawa, H.; Asai, A.; Fukunishi, S.; Nishiguchi, S.; Higuchi, K. Metabolic Syndrome and Sarcopenia. Nutrients 2021, 13, 3519. [Google Scholar] [CrossRef]
- Zuin, M.; Rigatelli, G.; Bilato, C.; Cervellati, C.; Zuliani, G.; Roncon, L. Prognostic Role of Metabolic Syndrome in COVID-19 Patients: A Systematic Review Meta-Analysis. Viruses 2021, 13, 1938. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, G.; Liccardi, A.; Graziadio, C.; Barrea, L.; Muscogiuri, G.; Colao, A. Obesity and Infectious Diseases: Pathophysiology and Epidemiology of a Double Pandemic Condition. Int. J. Obes. 2022, 46, 449–465. [Google Scholar] [CrossRef]
- Costa, F.F.; Rosário, W.R.; Ribeiro Farias, A.C.; de Souza, R.G.; Duarte Gondim, R.S.; Barroso, W.A. Metabolic Syndrome and COVID-19: An Update on the Associated Comorbidities and Proposed Therapies. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 809–814. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, A.P.; Ciampa, A.; Ingallina, C.; Mannina, L.; Capitani, D.; Ernesti, I.; Maggi, E.; Businaro, R.; del Ben, M.; Engel, P.; et al. Blueberry-Based Meals for Obese Patients with Metabolic Syndrome: A Multidisciplinary Metabolomic Pilot Study. Metabolites 2019, 9, 138. [Google Scholar] [CrossRef] [PubMed]
- Pasanta, D.; Chancharunee, S.; Tungjai, M.; Kim, H.J.; Kothan, S. Effects of Obesity on the Lipid and Metabolite Profiles of Young Adults by Serum 1H-NMR Spectroscopy. PeerJ 2019, 7, e7137. [Google Scholar] [CrossRef]
- Salek, R.M.; Maguire, M.L.; Bentley, E.; Rubtsov, D.V.; Hough, T.; Cheeseman, M.; Nunez, D.; Sweatman, B.C.; Haselden, J.N.; Cox, R.D.; et al. A Metabolomic Comparison of Urinary Changes in Type 2 Diabetes in Mouse, Rat, and Human. Physiol. Genom. 2007, 29, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Yang, S.O.; Kim, H.S.; Kim, Y.; Park, T.; Choi, H.K. 1H-Nuclear Magnetic Resonance Spectroscopy-Based Metabolic Assessment in a Rat Model of Obesity Induced by a High-Fat Diet. Anal. Bioanal Chem 2009, 395, 1117–1124. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, H.; Rong, S.; Bian, C.; Yang, Y.; Pan, H. NMR Spectroscopy Based Metabolomics Confirms the Aggravation of Metabolic Disorder in Metabolic Syndrome Combined with Hyperuricemia. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2449–2457. [Google Scholar] [CrossRef]
- Gonzalez-Franquesa, A.; Burkart, A.M.; Isganaitis, E.; Patti, M.E. What Have Metabolomics Approaches Taught Us About Type 2 Diabetes? Curr. Diab. Rep. 2016, 16, 74. [Google Scholar] [CrossRef]
- Abu Bakar Sajak, A.; Mediani, A.; Maulidiani; Ismail, A.; Abas, F. Metabolite Variation in Lean and Obese Streptozotocin (STZ)-Induced Diabetic Rats via 1H NMR-Based Metabolomics Approach. Appl. Biochem. Biotechnol. 2017, 182, 653–668. [Google Scholar] [CrossRef]
- Wolak-Dinsmore, J.; Gruppen, E.G.; Shalaurova, I.; Matyus, S.P.; Grant, R.P.; Gegen, R.; Bakker, S.J.L.; Otvos, J.D.; Connelly, M.A.; Dullaart, R.P.F. A Novel NMR-Based Assay to Measure Circulating Concentrations of Branched-Chain Amino Acids: Elevation in Subjects with Type 2 Diabetes Mellitus and Association with Carotid Intima Media Thickness. Clin. Biochem. 2018, 54, 92–99. [Google Scholar] [CrossRef]
- Tillin, T.; Hughes, A.D.; Wang, Q.; Würtz, P.; Ala-Korpela, M.; Sattar, N.; Forouhi, N.G.; Godsland, I.F.; Eastwood, S.V.; McKeigue, P.M.; et al. Diabetes Risk and Amino Acid Profiles: Cross-Sectional and Prospective Analyses of Ethnicity, Amino Acids and Diabetes in a South Asian and European Cohort from the SABRE (Southall And Brent REvisited) Study. Diabetologia 2015, 58, 968–979. [Google Scholar] [CrossRef] [PubMed]
- Würtz, P.; Tiainen, M.; Mäkinen, V.P.; Kangas, A.J.; Soininen, P.; Saltevo, J.; Keinänen-Kiukaanniemi, S.; Mäntyselkä, P.; Lehtimäki, T.; Laakso, M.; et al. Circulating Metabolite Predictors of Glycemia in Middle-Aged Men and Women. Diabetes Care 2012, 35, 1749–1756. [Google Scholar] [CrossRef] [PubMed]
- Abdul Ghani, Z.D.F.; Ab Rashid, A.H.; Shaari, K.; Chik, Z. Urine NMR Metabolomic Study on Biochemical Activities to Investigate the Effect of P. Betle Extract on Obese Rats. Appl. Biochem. Biotechnol. 2019, 189, 690–708. [Google Scholar] [CrossRef]
- Won, E.Y.; Yoon, M.K.; Kim, S.W.; Jung, Y.; Bae, H.W.; Lee, D.; Park, S.G.; Lee, C.H.; Hwang, G.S.; Chi, S.W. Gender-Specific Metabolomic Profiling of Obesity in Leptin-Deficient Ob/Ob Mice by 1H NMR Spectroscopy. PLoS ONE 2013, 8, e75998. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.; Wiklund, P.K.; Pekkala, S.; Autio, R.; Munukka, E.; Xu, L.; Saltevo, J.; Cheng, S.; Kujala, U.M.; Alen, M. Serum Metabolic Profiles in Overweight and Obese Women with and without Metabolic Syndrome. Diabetol. Metab. Syndr. 2014, 6, 40. [Google Scholar]
- Surowiec, I.; Noordam, R.; Bennett, K.; Beekman, M.; Slagboom, P.E.; Lundstedt, T.; van Heemst, D. Metabolomic and Lipidomic Assessment of the Metabolic Syndrome in Dutch Middle-Aged Individuals Reveals Novel Biological Signatures Separating Health and Disease. Metabolomics 2019, 15, 23. [Google Scholar] [CrossRef]
- Dai, L.; Gonçalves, C.M.V.; Lin, Z.; Huang, J.; Lu, H.; Yi, L.; Liang, Y.; Wang, D.; An, D. Exploring Metabolic Syndrome Serum Free Fatty Acid Profiles Based on GC-SIM-MS Combined with Random Forests and Canonical Correlation Analysis. Talanta 2015, 135, 108–114. [Google Scholar] [CrossRef]
- Yun, H.; Sun, L.; Wu, Q.; Zong, G.; Qi, Q.; Li, H.; Zheng, H.; Zeng, R.; Liang, L.; Lin, X. Associations among Circulating Sphingolipids, β-Cell Function, and Risk of Developing Type 2 Diabetes: A Population-Based Cohort Study in China. PLoS Med. 2020, 17, e1003451. [Google Scholar] [CrossRef]
- Yun, H.; Qi, Q.B.; Zong, G.; Wu, Q.Q.; Niu, Z.H.; Chen, S.S.; Li, H.X.; Sun, L.; Zeng, R.; Lin, X. Plasma Sphingolipid Profile in Association with Incident Metabolic Syndrome in a Chinese Population-Based Cohort Study. Nutrients 2021, 13, 2263. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Vicente Gonçalves, C.M.; Dai, L.; Lu, H.-M.; Huang, J.-H.; Ji, H.; Wang, D.-S.; Yi, L.-Z.; Liang, Y.-Z. Exploring Metabolic Syndrome Serum Profiling Based on Gas Chromatography Mass Spectrometry and Random Forest Models. Anal. Chim. Acta 2014, 827, 22–27. [Google Scholar] [CrossRef]
- Esperanza, M.G.; Wrobel, K.; Ojeda, A.G.; Garay-Sevilla, M.E.; Escobosa, A.R.C.; Barrientos, E.Y.; Wrobel, K. Liquid Chromatography-Mass Spectrometry Untargeted Metabolomics Reveals Increased Levels of Tryptophan Indole Metabolites in Urine of Metabolic Syndrome Patients. Eur. J. Mass Spectrom. 2020, 26, 379–387. [Google Scholar] [CrossRef]
- Bruzzone, C.; Gil-Redondo, R.; Seco, M.; Barragán, R.; de la Cruz, L.; Cannet, C.; Schäfer, H.; Fang, F.; Diercks, T.; Bizkarguenaga, M.; et al. A Molecular Signature for the Metabolic Syndrome by Urine Metabolomics. Cardiovasc. Diabetol. 2021, 20, 155. [Google Scholar] [CrossRef]
- Kanbay, M.; Jensen, T.; Solak, Y.; Le, M.; Roncal-Jimenez, C.; Rivard, C.; Lanaspa, M.A.; Nakagawa, T.; Johnson, R.J. Uric Acid in Metabolic Syndrome: From an Innocent Bystander to a Central Player. Eur. J. Intern. Med. 2016, 29, 3–8. [Google Scholar] [CrossRef]
- Bombelli, M.; Quarti-Trevano, F.; Tadic, M.; Facchetti, R.; Cuspidi, C.; Mancia, G.; Grassi, G. Uric Acid and Risk of New-Onset Metabolic Syndrome, Impaired Fasting Glucose and Diabetes Mellitus in a General Italian Population: Data from the Pressioni Arteriose Monitorate e Loro Associazioni Study. J. Hypertens. 2018, 36, 1492–1498. [Google Scholar] [CrossRef]
- Saito, N.; Saito, T.; Yamazaki, T.; Fujimine, Y.; Ihara, T. Establishment of an Analytical Method for Accurate Purity Evaluations of Acylcarnitines by Using Quantitative 1H NMR Spectroscopy. Accredit. Qual. Assur. 2017, 22, 171–178. [Google Scholar] [CrossRef]
- Lawler, P.R.; Akinkuolie, A.O.; Chandler, P.D.; Moorthy, M.V.; Vandenburgh, M.J.; Schaumberg, D.A.; Lee, I.M.; Glynn, R.J.; Ridker, P.M.; Buring, J.E.; et al. Circulating N-Linked Glycoprotein Acetyls and Longitudinal Mortality Risk. Circ. Res. 2016, 118, 1106–1115. [Google Scholar] [CrossRef] [PubMed]
- Otvos, J.D.; Shalaurova, I.; Wolak-Dinsmore, J.; Connelly, M.A.; Mackey, R.H.; Stein, J.H.; Tracy, R.P. GlycA: A Composite Nuclear Magnetic Resonance Biomarker of Systemic Inflammation. Clin. Chem. 2015, 61, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, S.C.; Würtz, P.; Nath, A.P.; Abraham, G.; Havulinna, A.S.; Fearnley, L.G.; Sarin, A.P.; Kangas, A.J.; Soininen, P.; Aalto, K.; et al. The Biomarker GlycA Is Associated with Chronic Inflammation and Predicts Long-Term Risk of Severe Infection. Cell Syst. 2015, 1, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Adosraku, R.K.; Choi, G.T.Y.; Constantinou-Kokotos, V.; Anderson, M.M.; Gibbons, W.A. NMR Lipid Profiles of Cells, Tissues, and Body Fluids: Proton NMR Analysis of Human Erythrocyte Lipids. J. Lipid Res. 1994, 35, 1925–1931. [Google Scholar] [CrossRef]
- Guo, W.; Jiang, C.; Yang, L.; Li, T.; Liu, X.; Jin, M.; Qu, K.; Chen, H.; Jin, X.; Liu, H.; et al. Quantitative Metabolomic Profiling of Plasma, Urine, and Liver Extracts by 1H NMR Spectroscopy Characterizes Different Stages of Atherosclerosis in Hamsters. J. Proteome Res. 2016, 15, 3500–3510. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.L.M.; Vogelzangs, N.; Mook-Kanamori, D.O.; Brahimaj, A.; Nano, J.; van der Heijden, A.A.W.A.; van Dijk, K.W.; Slieker, R.C.; Steyerberg, E.W.; Arfan Ikram, M.; et al. Blood Metabolomic Measures Associate with Present and Future Glycemic Control in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2018, 103, 4569–4579. [Google Scholar] [CrossRef] [PubMed]
- Denimal, D.; Nguyen, A.; Pais de Barros, J.P.; Bouillet, B.; Petit, J.M.; Vergès, B.; Duvillard, L. Major Changes in the Sphingophospholipidome of HDL in Non-Diabetic Patients with Metabolic Syndrome. Atherosclerosis 2016, 246, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Martínez, P.; Mikhailidis, D.P.; Athyros, V.G.; Bullo, M.; Couture, P.; Covas, M.I.; de Koning, L.; Delgado-Lista, J.; Díaz-López, A.; Drevon, C.A.; et al. Lifestyle Recommendations for the Prevention and Management of Metabolic Syndrome: An International Panel Recommendation. Nutr. Rev. 2017, 75, 307–326. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, E.; Farjami, Z.; Rezayi, M.; Lngari, H.; Darroudi, M.; Avan, A.; Ghayour-Mobarhan, M. Recent Advances in Nanotechnology for the Treatment of Metabolic Syndrome. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 1561–1568. [Google Scholar] [CrossRef]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef]
- Xie, Y.; Wu, Z.; Sun, L.; Zhou, L.; Wang, G.; Xiao, L.; Wang, H. The Effects and Mechanisms of Exercise on the Treatment of Depression. Front. Psychiatry 2021, 12, 705559. [Google Scholar] [CrossRef]
- Kapoor, G.; Chauhan, P.; Singh, G.; Malhotra, N.; Chahal, A. Physical Activity for Health and Fitness: Past, Present and Future. J. Lifestyle Med. 2022, 12, 9. [Google Scholar] [CrossRef]
- Myers, J.; Kokkinos, P.; Nyelin, E. Physical Activity, Cardiorespiratory Fitness, and the Metabolic Syndrome. Nutrients 2019, 11, 1652. [Google Scholar] [CrossRef]
- Wewege, M.A.; Thom, J.M.; Rye, K.A.; Parmenter, B.J. Aerobic, Resistance or Combined Training: A Systematic Review and Meta-Analysis of Exercise to Reduce Cardiovascular Risk in Adults with Metabolic Syndrome. Atherosclerosis 2018, 274, 162–171. [Google Scholar] [CrossRef]
- Reljic, D.; Frenk, F.; Herrmann, H.J.; Neurath, M.F.; Zopf, Y. Low-Volume High-Intensity Interval Training Improves Cardiometabolic Health, Work Ability and Well-Being in Severely Obese Individuals: A Randomized-Controlled Trial Sub-Study. J. Transl. Med. 2020, 18, 419. [Google Scholar] [CrossRef]
- Chiang, L.C.; Chiang, S.L.; Tzeng, W.C.; Lee, M.S.; Hung, Y.J.; Lin, C.H. Active Physical Activity Patterns Are Associated with Improved Quality of Life and Depression Status in Taiwanese Women with Metabolic Syndrome. J. Cardiovasc. Nurs. 2019, 34, 491–502. [Google Scholar] [CrossRef]
- Haufe, S.; Kahl, K.G.; Kerling, A.; Protte, G.; Bayerle, P.; Stenner, H.T.; Rolff, S.; Sundermeier, T.; Eigendorf, J.; Kück, M.; et al. Employers With Metabolic Syndrome and Increased Depression/Anxiety Severity Profit Most From Structured Exercise Intervention for Work Ability and Quality of Life. Front. Psychiatry 2020, 11, 562. [Google Scholar] [CrossRef]
- Emerenziani, G.P.; Gallotta, M.C.; Migliaccio, S.; Ferrari, D.; Greco, E.A.; Saavedra, F.J.; Iazzoni, S.; Aversa, A.; Donini, L.M.; Lenzi, A.; et al. Effects of an Individualized Home-Based Unsupervised Aerobic Training on Body Composition and Physiological Parameters in Obese Adults Are Independent of Gender. J. Endocrinol. Investig. 2018, 41, 465–473. [Google Scholar] [CrossRef]
- Emerenziani, G.P.; Ferrari, D.; Migliaccio, S.; Lenzi, A.; Greco, E.A.; Marocco, C.; Baldari, C.; Guidetti, L. Effects of Body Weight Loss Program on Parameters of Muscle Performance in Female Obese Adults. J. Sport. Med. Phys. Fit. 2019, 59, 624–631. [Google Scholar] [CrossRef]
- Grundy, S.M.; Vega, G.L.; Yuan, Z.; Battisti, W.P.; Brady, W.E.; Palmisano, J. Effectiveness and Tolerability of Simvastatin plus Fenofibrate for Combined Hyperlipidemia (the SAFARI Trial). Am. J. Cardiol. 2005, 95, 462–468. [Google Scholar] [CrossRef]
- Rubins, H.B.; Robins, S.J.; Collins, D.; Nelson, D.B.; Elam, M.B.; Schaefer, E.J.; Faas, F.H.; Anderson, J.W. Diabetes, Plasma Insulin, and Cardiovascular Disease: Subgroup Analysis from the Department of Veterans Affairs High-Density Lipoprotein Intervention Trial (VA-HIT). Arch. Intern. Med. 2002, 162, 2597–2604. [Google Scholar] [CrossRef]
- Chobanian, A.V.; Bakris, G.L.; Black, H.R.; Cushman, W.C.; Green, L.A.; Izzo, J.L.; Jones, D.W.; Materson, B.J.; Oparil, S.; Wright, J.T.; et al. Seventh Report of the Joint National Committee on Prevention, Detection, Evaluation, and Treatment of High Blood Pressure. Hypertension 2003, 42, 1206–1252. [Google Scholar] [CrossRef]
- Diabetes Prevention Program Research Group. Reduction in the Incidence of Type 2 Diabetes with Lifestyle Intervention or Metformin—NEJMoa012512. New Engl. J. Med. 2002, 346, 393–403. [Google Scholar] [CrossRef]
- Thi Quynh, D.; van Nga, V.; Thi Hoa, L.; Thi Hong Diem, L.; Thi Thom, V. Applying Framingham Risk Score 2008 to Predict the 10-Year Risk of Cardiovascular Disease in a Group of Office Workers in Hanoi, Vietnam. VNU J. Sci. Med. Pharm. Sci. 2019, 35, 128–136. [Google Scholar] [CrossRef]
- Christensen, P.; Meinert Larsen, T.; Westerterp-Plantenga, M.; Macdonald, I.; Martinez, J.A.; Handjiev, S.; Poppitt, S.; Hansen, S.; Ritz, C.; Astrup, A.; et al. Men and Women Respond Differently to Rapid Weight Loss: Metabolic Outcomes of a Multi-Centre Intervention Study after a Low-Energy Diet in 2500 Overweight, Individuals with Pre-Diabetes (PREVIEW). Diabetes Obes. Metab. 2018, 20, 2840–2851. [Google Scholar] [CrossRef]
- Geidenstam, N.; Al-Majdoub, M.; Ekman, M.; Spégel, P.; Ridderstråle, M. Metabolite Profiling of Obese Individuals before and after a One Year Weight Loss Program. Int. J. Obes. 2017, 41, 1369–1378. [Google Scholar] [CrossRef]
- Geidenstam, N.; Hsu, Y.H.H.; Astley, C.M.; Mercader, J.M.; Ridderstråle, M.; Gonzalez, M.E.; Gonzalez, C.; Hirschhorn, J.N.; Salem, R.M. Using Metabolite Profiling to Construct and Validate a Metabolite Risk Score for Predicting Future Weight Gain. PLoS ONE 2019, 14, e0222445. [Google Scholar] [CrossRef]
- Pigsborg, K.; Magkos, F. Metabotyping for Precision Nutrition and Weight Management: Hype or Hope? Curr. Nutr. Rep. 2022, 11, 117–123. [Google Scholar] [CrossRef]
- O’Donovan, C.B.; Walsh, M.C.; Woolhead, C.; Forster, H.; Celis-Morales, C.; Fallaize, R.; Macready, A.L.; Marsaux, C.F.M.; Navas-Carretero, S.; Rodrigo San-Cristobal, S.; et al. Metabotyping for the Development of Tailored Dietary Advice Solutions in a European Population: The Food4Me Study. Br. J. Nutr. 2017, 118, 561–569. [Google Scholar] [CrossRef] [PubMed]
- Berry, S.E.; Valdes, A.M.; Drew, D.A.; Asnicar, F.; Mazidi, M.; Wolf, J.; Capdevila, J.; Hadjigeorgiou, G.; Davies, R.; al Khatib, H.; et al. Human Postprandial Responses to Food and Potential for Precision Nutrition. Nat. Med. 2020, 26, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Gijbels, A.; Trouwborst, I.; Jardon, K.M.; Hul, G.B.; Siebelink, E.; Bowser, S.M.; Yildiz, D.; Wanders, L.; Erdos, B.; Thijssen, D.H.J.; et al. The PERSonalized Glucose Optimization Through Nutritional Intervention (PERSON) Study: Rationale, Design and Preliminary Screening Results. Front. Nutr. 2021, 8, 694568. [Google Scholar] [CrossRef]
- Wu, Q.; Li, J.; Sun, X.; He, D.; Cheng, Z.; Li, J.; Zhang, X.; Xie, Y.; Zhu, Y.; Lai, M. Multi-Stage Metabolomics and Genetic Analyses Identified Metabolite Biomarkers of Metabolic Syndrome and Their Genetic Determinants. EBioMedicine 2021, 74, 103707. [Google Scholar] [CrossRef]
- Riedl, A.; Gieger, C.; Hauner, H.; Daniel, H.; Linseisen, J. Metabotyping and Its Application in Targeted Nutrition: An Overview. Br. J. Nutr. 2017, 117, 1631–1644. [Google Scholar] [CrossRef]
- Agodi, A.; Maugeri, A.; Kunzova, S.; Sochor, O.; Bauerova, H.; Kiacova, N.; Barchitta, M.; Vinciguerra, M. Association of Dietary Patterns with Metabolic Syndrome: Results from the Kardiovize Brno 2030 Study. Nutrients 2018, 10, 898. [Google Scholar] [CrossRef]
- Salas-Salvadó, J.; Guasch-Ferré, M.; Lee, C.H.; Estruch, R.; Clish, C.B.; Ros, E. Protective Effects of the Mediterranean Diet on Type 2 Diabetes and Metabolic Syndrome. J. Nutr. 2016, 146, 920S–927S. [Google Scholar] [CrossRef]
- Ramzan, F.; Vickers, M.H.; Mithen, R.F. Epigenetics, Microrna and Metabolic Syndrome: A Comprehensive Review. Int. J. Mol. Sci. 2021, 22, 5047. [Google Scholar] [CrossRef]
- Marques-Rocha, J.L.; Milagro, F.I.; Mansego, M.L.; Zulet, M.A.; Bressan, J.; Martínez, J.A. Expression of Inflammation-Related MiRNAs in White Blood Cells from Subjects with Metabolic Syndrome after 8 Wk of Following a Mediterranean Diet-Based Weight Loss Program. Nutrition 2016, 32, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Gillies, P.J.; Bhatia, S.K.; Belcher, L.A.; Hannon, D.B.; Thompson, J.T.; van den Heuvel, J.P. Regulation of Inflammatory and Lipid Metabolism Genes by Eicosapentaenoic Acid-Rich Oil. J. Lipid Res. 2012, 53, 1679–1689. [Google Scholar] [CrossRef]
- Ide, T. Interaction of Fish Oil and Conjugated Linoleic Acid in Affecting Hepatic Activity of Lipogenic Enzymes and Gene Expression in Liver and Adipose Tissue. Diabetes 2005, 54, 412–423. [Google Scholar] [CrossRef]
- Ortega, F.J.; Cardona-Alvarado, M.I.; Mercader, J.M.; Moreno-Navarrete, J.M.; Moreno, M.; Sabater, M.; Fuentes-Batllevell, N.; Ramírez-Chávez, E.; Ricart, W.; Molina-Torres, J.; et al. Circulating Profiling Reveals the Effect of a Polyunsaturated Fatty Acid-Enriched Diet on Common MicroRNAs. J. Nutr. Biochem. 2015, 26, 1095–1101. [Google Scholar] [CrossRef]
- Castro-Barquero, S.; Ruiz-León, A.M.; Sierra-Pérez, M.; Estruch, R.; Casas, R. Dietary Strategies for Metabolic Syndrome: A Comprehensive Review. Nutrients 2020, 12, 2983. [Google Scholar] [CrossRef]
- Lefevre, M.; Redman, L.M.; Heilbronn, L.K.; Smith, J.V.; Martin, C.K.; Rood, J.C.; Greenway, F.L.; Williamson, D.A.; Smith, S.R.; Ravussin, E. Caloric Restriction Alone and with Exercise Improves CVD Risk in Healthy Non-Obese Individuals. Atherosclerosis 2009, 203, 206–213. [Google Scholar] [CrossRef]
- Most, J.; Gilmore, L.A.; Smith, S.R.; Han, H.; Ravussin, E.; Redman, L.M. Significant Improvement in Cardiometabolic Health in Healthy Nonobese Individuals during Caloric Restriction-Induced Weight Loss and Weight Loss Maintenance. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E396–E405. [Google Scholar] [CrossRef]
- Wan, R.; Camandola, S.; Mattson, M.P. Intermittent Food Deprivation Improves Cardiovascular and Neuroendocrine Responses to Stress in Rats. J. Nutr. 2003, 133, 1921–1929. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, E.J.; Kim, Y.N.; Choi, C.; Lee, B.H. Comparison of the Chemical Compositions and Nutritive Values of Various Pumpkin (Cucurbitaceae) Species and Parts. Nutr. Res. Pract. 2012, 6, 21–27. [Google Scholar] [CrossRef]
- Abdelkader, C.; Cherif, F.Z.H.; Elius, E.A.E.; Lucchesi, D.; Pucci, L.; Yahia, D.A. Pumpkin Seed Proteins (Cucurbita Pepo, L.) Protect against Diet-Induced Metabolic Syndrome by Improving Insulin Resistance and Markers of Oxidative Stress and Inflammation in Rats. Biologia 2022, 77, 2677–2687. [Google Scholar] [CrossRef]
- Akrami, A.; Makiabadi, E.; Askarpour, M.; Zamani, K.; Hadi, A.; Mokari-Yamchi, A.; Babajafari, S.; Faghih, S.; Hojhabrimanesh, A. A Comparative Study of the Effect of Flaxseed Oil and Sunflower Oil on the Coagulation Score, Selected Oxidative and Inflammatory Parameters in Metabolic Syndrome Patients. Clin. Nutr. Res. 2020, 9, 63–72. [Google Scholar] [CrossRef]
- Dewell, A.; Marvasti, F.F.; Harris, W.S.; Tsao, P.; Gardner, C.D. Low- and High-Dose Plant and Marine (n-3) Fatty Acids Do Not Affect Plasma Inflammatory Markers in Adults with Metabolic Syndrome. J. Nutr. 2011, 141, 2166–2171. [Google Scholar] [CrossRef]
- Din, J.N.; Archer, R.M.; Harding, S.A.; Sarma, J.; Lyall, K.; Flapan, A.D.; Newby, D.E. Effect of ω-3 Fatty Acid Supplementation on Endothelial Function, Endogenous Fibrinolysis and Platelet Activation in Male Cigarette Smokers. Heart 2013, 99, 168–174. [Google Scholar] [CrossRef]
- Sangouni, A.A.; Alizadeh, M.; Jamalzehi, A.; Parastouei, K. Effects of Garlic Powder Supplementation on Metabolic Syndrome Components, Insulin Resistance, Fatty Liver Index, and Appetite in Subjects with Metabolic Syndrome: A Randomized Clinical Trial. Phytother. Res. 2021, 35, 4433–4441. [Google Scholar] [CrossRef]
- Rezazadeh, K.; Aliashrafi, S.; Asghari-Jafarabadi, M.; Ebrahimi-Mameghani, M. Antioxidant Response to Artichoke Leaf Extract Supplementation in Metabolic Syndrome: A Double-Blind Placebo-Controlled Randomized Clinical Trial. Clin. Nutr. 2018, 37, 790–796. [Google Scholar] [CrossRef]
- Li, Y.; Tran, V.H.; Kota, B.P.; Nammi, S.; Duke, C.C.; Roufogalis, B.D. Preventative Effect of Zingiber Officinale on Insulin Resistance in a High-Fat High-Carbohydrate Diet-Fed Rat Model and Its Mechanism of Action. Basic Clin. Pharmacol. Toxicol. 2014, 115, 209–215. [Google Scholar] [CrossRef]
- Sarriá, B.; Martínez-López, S.; Sierra-Cinos, J.L.; García-Diz, L.; Mateos, R.; Bravo-Clemente, L. Regularly Consuming a Green/Roasted Coffee Blend Reduces the Risk of Metabolic Syndrome. Eur. J. Nutr. 2018, 57, 269–278. [Google Scholar] [CrossRef]
- Wu, G.; Sun, X.; Cheng, H.; Xu, S.; Li, D.; Xie, Z. Large Yellow Tea Extract Ameliorates Metabolic Syndrome by Suppressing Lipogenesis through SIRT6/SREBP1 Pathway and Modulating Microbiota in Leptin Receptor Knockout Rats. Foods 2022, 11, 1638. [Google Scholar] [CrossRef]
- Lee, M.; Lim, M.; Kim, J. Fruit and Vegetable Consumption and the Metabolic Syndrome: A Systematic Review and Dose-Response Meta-Analysis. Br. J. Nutr. 2019, 122, 723–733. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, D.Z. Associations of Vegetable and Fruit Consumption with Metabolic Syndrome. A Meta-Analysis of Observational Studies. Public Health Nutr. 2018, 21, 1693–1703. [Google Scholar] [CrossRef]
- Tian, Y.; Su, L.; Wang, J.; Duan, X.; Jiang, X. Fruit and Vegetable Consumption and Risk of the Metabolic Syndrome: A Meta-Analysis. Public Health Nutr. 2018, 21, 756–765. [Google Scholar] [CrossRef]
- Pap, N.; Fidelis, M.; Azevedo, L.; do Carmo, M.A.V.; Wang, D.; Mocan, A.; Pereira, E.P.R.; Xavier-Santos, D.; Sant’Ana, A.S.; Yang, B.; et al. Berry Polyphenols and Human Health: Evidence of Antioxidant, Anti-Inflammatory, Microbiota Modulation, and Cell-Protecting Effects. Curr. Opin. Food Sci. 2021, 42, 167–186. [Google Scholar] [CrossRef]
- Simão, T.N.C.; Lozovoy, M.A.B.; Simão, A.N.C.; Oliveira, S.R.; Venturini, D.; Morimoto, H.K.; Miglioranza, L.H.S.; Dichi, I. Reduced-Energy Cranberry Juice Increases Folic Acid and Adiponectin and Reduces Homocysteine and Oxidative Stress in Patients with the Metabolic Syndrome. Br. J. Nutr. 2013, 110, 1885–1894. [Google Scholar] [CrossRef]
- Aboonabi, A.; Meyer, R.R.; Gaiz, A.; Singh, I. Anthocyanins in Berries Exhibited Anti-Atherogenicity and Antiplatelet Activities in a Metabolic Syndrome Population. Nutr. Res. 2020, 76, 82–93. [Google Scholar] [CrossRef]
- Carresi, C.; Gliozzi, M.; Musolino, V.; Scicchitano, M.; Scarano, F.; Bosco, F.; Nucera, S.; Maiuolo, J.; Macrì, R.; Ruga, S.; et al. The Effect of Natural Antioxidants in the Development of Metabolic Syndrome: Focus on Bergamot Polyphenolic Fraction. Nutrients 2020, 12, 1504. [Google Scholar] [CrossRef]
- Salerno, R.; Casale, F.; Calandruccio, C.; Procopio, A. Characterization of Flavonoids in Citrus Bergamia (Bergamot) Polyphenolic Fraction by Liquid Chromatography–High Resolution Mass Spectrometry (LC/HRMS). PharmaNutrition 2016, 4, S1–S7. [Google Scholar] [CrossRef]
- Mollace, V.; Sacco, I.; Janda, E.; Malara, C.; Ventrice, D.; Colica, C.; Visalli, V.; Muscoli, S.; Ragusa, S.; Muscoli, C.; et al. Hypolipemic and Hypoglycaemic Activity of Bergamot Polyphenols: From Animal Models to Human Studies. Fitoterapia 2011, 82, 309–316. [Google Scholar] [CrossRef]
- Cicero, A.F.G.; Fogacci, F.; Bove, M.; Giovannini, M.; Borghi, C. Three-Arm, Placebo-Controlled, Randomized Clinical Trial Evaluating the Metabolic Effect of a Combined Nutraceutical Containing a Bergamot Standardized Flavonoid Extract in Dyslipidemic Overweight Subjects. Phytother. Res. 2019, 33, 2094–2101. [Google Scholar] [CrossRef]
- Arabi, S.M.; Bahrami, L.S.; Milkarizi, N.; Nematy, M.; Kalmykov, V.; Sahebkar, A. Impact of Walnut Consumption on Cardio Metabolic and Anthropometric Parameters in Metabolic Syndrome Patients: GRADE-Assessed Systematic Review and Dose-Response Meta-Analysis of Data from Randomized Controlled Trials. Pharmacol. Res. 2022, 178, 106190. [Google Scholar] [CrossRef]
- Hwang, H.J.; Liu, Y.; Kim, H.S.; Lee, H.; Lim, Y.; Park, H. Daily Walnut Intake Improves Metabolic Syndrome Status and Increases Circulating Adiponectin Levels: Randomized Controlled Crossover Trial. Nutr. Res. Pract. 2019, 13, 105–114. [Google Scholar] [CrossRef]
- Maphosa, Y.; Jideani, V.A. The Role of legumes in human nutrition. In Functional Food—Improve Health through Adequate Food; BoD—Books on Demand: Norderstedt, Germany, 2017. [Google Scholar]
- Reverri, E.J.; Randolph, J.M.; Steinberg, F.M.; Tissa Kappagoda, C.; Edirisinghe, I.; Burton-Freeman, B.M. Black Beans, Fiber, and Antioxidant Capacity Pilot Study: Examination of Whole Foods vs. Functional Components on Postprandial Metabolic, Oxidative Stress, and Inflammation in Adults with Metabolic Syndrome. Nutrients 2015, 7, 6139–6154. [Google Scholar] [CrossRef]
- Azadbakht, L.; Kimiagar, M.; Mehrabi, Y.; Esmaillzadeh, A.; Padyab, M.; Hu, F.B.; Willett, W.C. Soy Inclusion in the Diet Improves Features of the Metabolic Syndrome: A Randomized Crossover Study in Postmenopausal Women. Am. J. Clin. Nutr. 2007, 85, 735–741. [Google Scholar] [CrossRef]
- Ruscica, M.; Pavanello, C.; Gandini, S.; Gomaraschi, M.; Vitali, C.; Macchi, C.; Morlotti, B.; Aiello, G.; Bosisio, R.; Calabresi, L.; et al. Effect of Soy on Metabolic Syndrome and Cardiovascular Risk Factors: A Randomized Controlled Trial. Eur. J. Nutr. 2018, 57, 499–511. [Google Scholar] [CrossRef]
- Guo, H.; Ding, J.; Liang, J.; Zhang, Y. Associations of Whole Grain and Refined Grain Consumption With Metabolic Syndrome. A Meta-Analysis of Observational Studies. Front. Nutr. 2021, 8, 695620. [Google Scholar] [CrossRef]
- Song, S.J.; Lee, J.E.; Song, W.O.; Paik, H.Y.; Song, Y.J. Carbohydrate Intake and Refined-Grain Consumption Are Associated with Metabolic Syndrome in the Korean Adult Population. J. Acad Nutr. Diet. 2014, 114, 54–62. [Google Scholar] [CrossRef]
- Sahyoun, N.R.; Jacques, P.F.; Zhang, X.L.; Juan, W.; McKeown, N.M. Whole-Grain Intake Is Inversely Associated with the Metabolic Syndrome and Mortality in Older Adults. Am. J. Clin. Nutr. 2006, 83, 124–131. [Google Scholar] [CrossRef]
- Esmaillzadeh, A.; Mirmiran, P.; Azizi, F. Whole-Grain Consumption and the Metabolic Syndrome: A Favorable Association in Tehranian Adults. Eur. J. Clin. Nutr. 2005, 59, 353–362. [Google Scholar] [CrossRef]
- Liu, S.; Manson, J.E.; Stamfer, M.J.; Hu, F.B.; Giovannucci, E.; Colditz, G.A.; Hennekens, C.H.; Willett, W.C. A Prospective Study of Whole-Grain Intake and Risk of Type 2 Diabetes Mellitus in US Women. Am. J. Public Health 2000, 90, 1409. [Google Scholar]
- Radhika, G.; van Dam, R.M.; Sudha, V.; Ganesan, A.; Mohan, V. Refined Grain Consumption and the Metabolic Syndrome in Urban Asian Indians (Chennai Urban Rural Epidemiology Study 57). Metabolism 2009, 58, 675–681. [Google Scholar] [CrossRef]
- Giacco, R.; Lappi, J.; Costabile, G.; Kolehmainen, M.; Schwab, U.; Landberg, R.; Uusitupa, M.; Poutanen, K.; Pacini, G.; Rivellese, A.A.; et al. Effects of Rye and Whole Wheat versus Refined Cereal Foods on Metabolic Risk Factors: A Randomised Controlled Two-Centre Intervention Study. Clin. Nutr. 2013, 32, 941–949. [Google Scholar] [CrossRef]
- Giacco, R.; Costabile, G.; della Pepa, G.; Anniballi, G.; Griffo, E.; Mangione, A.; Cipriano, P.; Viscovo, D.; Clemente, G.; Landberg, R.; et al. A Whole-Grain Cereal-Based Diet Lowers Postprandial Plasma Insulin and Triglyceride Levels in Individuals with Metabolic Syndrome. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 837–844. [Google Scholar] [CrossRef]
- Huang, L.; Wang, H.; Wang, Z.; Zhang, J.; Zhang, B.; Ding, G. Regional Disparities in the Association between Cereal Consumption and Metabolic Syndrome: Results from the China Health and Nutrition Survey. Nutrients 2019, 11, 764. [Google Scholar] [CrossRef]
- Kang, Y.; Lee, K.; Lee, J.; Kim, J. Grain Subtype and the Combination of Grains Consumed Are Associated with the Risk of Metabolic Syndrome: Analysis of a Community-Based Prospective Cohort. J. Nutr. 2020, 150, 118–127. [Google Scholar] [CrossRef]
- Zhou, L.; Hu, S.; Rong, S.; Mo, X.; Wang, Q.; Yin, J.; Peng, X.; Li, X.; Chen, L.; Shan, Z.; et al. DHPPA, a Major Plasma Alkylresorcinol Metabolite Reflecting Whole-Grain Wheat and Rye Intake, and Risk of Metabolic Syndrome: A Case-Control Study. Eur. J. Nutr. 2022, 61, 3247–3254. [Google Scholar] [CrossRef]
- Yang, L.; Chen, J.H.; Lv, J.; Wu, Q.; Xu, T.; Zhang, H.; Liu, Q.H.; Yang, H.K. Rice Protein Improves Adiposity, Body Weight and Reduces Lipids Level in Rats through Modification of Triglyceride Metabolism. Lipids Health Dis. 2012, 11, 118–127. [Google Scholar] [CrossRef]
- Bahrami, A.; Khalesi, S.; Makiabadi, E.; Alibeyk, S.; Hajigholam-Saryazdi, M.; Hejazi, E. Adherence to the Mediterranean Diet and the Risk of Lung Cancer: A Systematic Review and Dose-Response Meta-Analysis of Observational Studies. Nutr. Rev. 2022, 80, 1118–1128. [Google Scholar] [CrossRef]
- Gorzynik-Debicka, M.; Przychodzen, P.; Cappello, F.; Kuban-Jankowska, A.; Gammazza, A.M.; Knap, N.; Wozniak, M.; Gorska-Ponikowska, M. Potential Health Benefits of Olive Oil and Plant Polyphenols. Int. J. Mol. Sci. 2018, 19, 686. [Google Scholar] [CrossRef]
- Violi, F.; Loffredo, L.; Pignatelli, P.; Angelico, F.; Bartimoccia, S.; Nocella, C.; Cangemi, R.; Petruccioli, A.; Monticolo, R.; Pastori, D.; et al. Extra Virgin Olive Oil Use Is Associated with Improved Post-Prandial Blood Glucose and LDL Cholesterol in Healthy Subjects. Nutr. Diabetes 2015, 5, e172. [Google Scholar] [CrossRef]
- Venturini, D.; Simão, A.N.C.; Urbano, M.R.; Dichi, I. Effects of Extra Virgin Olive Oil and Fish Oil on Lipid Profile and Oxidative Stress in Patients with Metabolic Syndrome. Nutrition 2015, 31, 834–840. [Google Scholar] [CrossRef]
- Raatz, S.K.; Silverstein, J.T.; Jahns, L.; Picklo, M.J. Issues of Fish Consumption for Cardiovascular Disease Risk Reduction. Nutrients 2013, 5, 1081–1097. [Google Scholar] [CrossRef] [PubMed]
- Rylander, C.; Sandanger, T.M.; Engeset, D.; Lund, E. Consumption of Lean Fish Reduces the Risk of Type 2 Diabetes Mellitus: A Prospective Population Based Cohort Study of Norwegian Women. PLoS ONE 2014, 9, e89845. [Google Scholar] [CrossRef]
- Kim, Y.S.; Xun, P.; He, K. Fish Consumption, Long-Chain Omega-3 Polyunsaturated Fatty Acid Intake and Risk of Metabolic Syndrome: A Meta-Analysis. Nutrients 2015, 7, 2085–2100. [Google Scholar] [CrossRef]
- Li, N.; Jia, M.; Deng, Q.; Wang, Z.; Huang, F.; Hou, H.; Xu, T. Effect of Low-Ratio n-6/n-3 PUFA on Blood Lipid Level: A Meta-Analysis. Hormones 2021, 20, 697–706. [Google Scholar] [CrossRef]
- Jiang, X.; Yang, Z.; Wang, S.; Deng, S. “Big Data” Approaches for Prevention of the Metabolic Syndrome. Front. Genet. 2022, 13, 810152. [Google Scholar] [CrossRef]
- Guo, X.F.; Li, X.; Shi, M.; Li, D. N-3 Polyunsaturated Fatty Acids and Metabolic Syndrome Risk: A Meta-Analysis. Nutrients 2017, 9, 703. [Google Scholar] [CrossRef]
- Coltell, O.; Sorlí, J.V.; Asensio, E.M.; Barragán, R.; González, J.I.; Giménez-Alba, I.M.; Zanón-Moreno, V.; Estruch, R.; Ramírez-Sabio, J.B.; Pascual, E.C.; et al. Genome-Wide Association Study for Serum Omega-3 and Omega-6 Polyunsaturated Fatty Acids: Exploratory Analysis of the Sex-Specific Effects and Dietary Modulation in Mediterranean Subjects with Metabolic Syndrome. Nutrients 2020, 12, 310. [Google Scholar] [CrossRef]
- Lai, Y.H.L.; Petrone, A.B.; Pankow, J.S.; Arnett, D.K.; North, K.E.; Ellison, R.C.; Hunt, S.C.; Djoussé, L. Association of Dietary Omega-3 Fatty Acids with Prevalence of Metabolic Syndrome: The National Heart, Lung, and Blood Institute Family Heart Study. Clin. Nutr. 2013, 32, 966–969. [Google Scholar] [CrossRef]
- Denisenko, Y.; Novgorodtseva, T.; Zhukova, N.; Gvozdenko, T.; Antonyuk, M.; Knyshova, V.; Nazarenko, A. Metabolic Syndrome: Modification of the Fatty Acid Composition and Glucose-Insulin Homeostasis. Br. J. Med. Med. Res. 2015, 8, 975–987. [Google Scholar] [CrossRef]
- Huang, T.; Bhulaidok, S.; Cai, Z.; Xu, T.; Xu, F.; Wahlqvist, M.L.; Li, D. Plasma Phospholipids N-3 Polyunsaturated Fatty Acid Is Associated with Metabolic Syndrome. Mol. Nutr. Food Res. 2010, 54, 1628–1635. [Google Scholar] [CrossRef]
- Dai, X.W.; Chen, Y.M.; Zeng, F.F.; Sun, L.L.; Chen, C.G.; Su, Y.X. Association between N-3 Polyunsaturated Fatty Acids in Erythrocytes and Metabolic Syndrome in Chinese Men and Women. Eur. J. Nutr. 2016, 55, 981–989. [Google Scholar] [CrossRef]
- Chien, K.L.; Chao, C.L.; Kuo, C.H.; Lin, H.J.; Liu, P.H.; Chen, P.R.; Hsu, H.C.; Lee, B.C.; Lee, Y.T.; Chen, M.F. Plasma Fatty Acids and the Risk of Metabolic Syndrome in Ethnic Chinese Adults in Taiwan. Lipids Health Dis. 2011, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Zaribaf, F.; Falahi, E.; Barak, F.; Heidari, M.; Keshteli, A.H.; Yazdannik, A.; Esmaillzadeh, A. Fish Consumption Is Inversely Associated with the Metabolic Syndrome. Eur. J. Clin. Nutr. 2014, 68, 474–480. [Google Scholar] [CrossRef]
- Mirmiran, P.; Asghari, G.; Dizavi, A.; Farhadnejad, H.; Azizi, F. The Association between Fish Consumption and Risk of Metabolic Syndrome in Adults: Tehran Lipid and Glucose Study. Int. J. Vitam. Nutr. Res. 2019, 89, 192–199. [Google Scholar] [CrossRef]
- Mirmiran, P.; Hosseinpour-Niazi, S.; Naderi, Z.; Bahadoran, Z.; Sadeghi, M.; Azizi, F. Association between Interaction and Ratio of ω-3 and ω-6 Polyunsaturated Fatty Acid and the Metabolic Syndrome in Adults. Nutrition 2012, 28, 856–863. [Google Scholar] [CrossRef]
- Kim, Y.S.; Xun, P.; Iribarren, C.; van Horn, L.; Steffen, L.; Daviglus, M.L.; Siscovick, D.; Liu, K.; He, K. Intake of Fish and Long-Chain Omega-3 Polyunsaturated Fatty Acids and Incidence of Metabolic Syndrome among American Young Adults: A 25-Year Follow-up Study. Eur. J. Nutr. 2016, 55, 1707–1716. [Google Scholar] [CrossRef]
- Kim, K.Y.; Park, J.S. Impact of Fish Consumption by Subjects with Prediabetes on the Metabolic Risk Factors: Using Data in the 2015 (6th) Korea National Health and Nutrition Examination Surveys. Nutr. Res. Pract. 2018, 12, 233–242. [Google Scholar] [CrossRef]
- Tørris, C.; Molin, M.; Småstuen, M.C. Associations between Fish Consumption and Metabolic Syndrome. A Large Cross-Sectional Study from the Norwegian Tromsø Study: Tromsø 4. Diabetol. Metab. Syndr. 2016, 8, 18. [Google Scholar] [CrossRef]
- Tørris, C.; Molin, M.; Småstuen, M.C. Lean Fish Consumption Is Associated with Beneficial Changes in the Metabolic Syndrome Components: A 13-Year Follow-up Study from the Norwegian Tromsø Study. Nutrients 2017, 9, 247. [Google Scholar] [CrossRef]
- Poudyal, H.; Panchal, S.K.; Ward, L.C.; Brown, L. Effects of ALA, EPA and DHA in High-Carbohydrate, High-Fat Diet-Induced Metabolic Syndrome in Rats. J. Nutr. Biochem. 2013, 24, 1041–1052. [Google Scholar] [CrossRef]
- Tardivo, A.P.; Nahas-Neto, J.; Orsatti, C.L.; Dias, F.B.; Poloni, P.F.; Schmitt, E.B.; Nahas, E.A.P. Effects of Omega-3 on Metabolic Markers in Postmenopausal Women with Metabolic Syndrome. Climacteric 2015, 18, 290–298. [Google Scholar] [CrossRef]
- Lee, T.C.; Ivester, P.; Hester, A.G.; Sergeant, S.; Case, L.D.; Morgan, T.; Kouba, E.O.; Chilton, F.H. The Impact of Polyunsaturated Fatty Acid-Based Dietary Supplements on Disease Biomarkers in a Metabolic Syndrome/Diabetes Population. Lipids Health Dis. 2014, 13, 196. [Google Scholar] [CrossRef]
- Ortega, J.F.; Morales-Palomo, F.; Fernandez-Elias, V.; Hamouti, N.; Bernardo, F.J.; Martin-Doimeadios, R.C.; Nelson, R.K.; Horowitz, J.F.; Mora-Rodriguez, R. Dietary Supplementation with Omega-3 Fatty Acids and Oleate Enhances Exercise Training Effects in Patients with Metabolic Syndrome. Obesity 2016, 24, 1704–1711. [Google Scholar] [CrossRef]
- Vázquez, C.; Botella-Carretero, J.I.; Corella, D.; Fiol, M.; Lage, M.; Lurbe, E.; Richart, C.; Fernández-Real, J.M.; Fuentes, F.; Ordóñez, A.; et al. White Fish Reduces Cardiovascular Risk Factors in Patients with Metabolic Syndrome: The WISH-CARE Study, a Multicenter Randomized Clinical Trial. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 328–335. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant Polyphenols as Dietary Antioxidants in Human Health and Disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Ghani, U. Chapter three—Polyphenols. In Alpha-Glucosidase Inhibitors, 1st ed.; Ghani, U., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 61–100. [Google Scholar]
- Shahidi, F.; Ambigaipalan, P. Phenolics and Polyphenolics in Foods, Beverages and Spices: Antioxidant Activity and Health Effects—A Review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of Dietary Polyphenols on Metabolic Syndrome Features in Humans: A Systematic Review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef]
- Parhiz, H.; Roohbakhsh, A.; Soltani, F.; Rezaee, R.; Iranshahi, M. Antioxidant and Anti-Inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Updated Review of Their Molecular Mechanisms and Experimental Models. Phytother. Res. 2015, 29, 323–331. [Google Scholar] [CrossRef]
- Barth, S.W.; Koch, T.C.L.; Watzl, B.; Dietrich, H.; Will, F.; Bub, A. Moderate Effects of Apple Juice Consumption on Obesity-Related Markers in Obese Men: Impact of Diet-Gene Interaction on Body Fat Content. Eur. J. Nutr. 2012, 51, 841–850. [Google Scholar] [CrossRef]
- Medina-Remón, A.; Tresserra-Rimbau, A.; Pons, A.; Tur, J.A.; Martorell, M.; Ros, E.; Buil-Cosiales, P.; Sacanella, E.; Covas, M.I.; Corella, D.; et al. Effects of Total Dietary Polyphenols on Plasma Nitric Oxide and Blood Pressure in a High Cardiovascular Risk Cohort. The PREDIMED Randomized Trial. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 60–67. [Google Scholar] [CrossRef]
- Macready, A.L.; George, T.W.; Chong, M.F.; Alimbetov, D.S.; Jin, Y.; Vidal, A.; Spencer, J.P.E.; Kennedy, O.B.; Tuohy, K.M.; Minihane, A.M.; et al. Flavonoid-Rich Fruit and Vegetables Improve Microvascular Reactivity and Inflammatory Status in Men at Risk of Cardiovascular Disease-FLAVURS: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2014, 99, 479–489. [Google Scholar] [CrossRef]
- Qin, W.; Ren, B.; Wang, S.; Liang, S.; He, B.; Shi, X.; Wang, L.; Liang, J.; Wu, F. Apigenin and Naringenin Ameliorate PKCβII-Associated Endothelial Dysfunction via Regulating ROS/Caspase-3 and NO Pathway in Endothelial Cells Exposed to High Glucose. Vascul. Pharmacol. 2016, 85, 39–49. [Google Scholar] [CrossRef]
- Duarte, S.; Arango, D.; Parihar, A.; Hamel, P.; Yasmeen, R.; Doseff, A.I. Apigenin Protects Endothelial Cells from Lipopolysaccharide (LPS)-Induced Inflammation by Decreasing Caspase-3 Activation and Modulating Mitochondrial Function. Int. J. Mol. Sci. 2013, 14, 17664–17679. [Google Scholar] [CrossRef]
- Wang, Z.M.; Zhao, D.; Nie, Z.L.; Zhao, H.; Zhou, B.; Gao, W.; Wang, L.S.; Yang, Z.J. Flavonol Intake and Stroke Risk: A Meta-Analysis of Cohort Studies. Nutrition 2014, 30, 518–523. [Google Scholar] [CrossRef]
- McCullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid Intake and Cardiovascular Disease Mortality in a Prospective Cohort of US Adults. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar] [CrossRef]
- Cassidy, A.; Bertoia, M.; Chiuve, S.; Flint, A.; Forman, J.; Rimm, E.B. Habitual Intake of Anthocyanins and Flavanones and Risk of Cardiovascular Disease in Men. Am. J. Clin. Nutr. 2016, 104, 587–594. [Google Scholar] [CrossRef]
- Dower, J.I.; Geleijnse, J.M.; Hollman, P.C.H.; Soedamah-Muthu, S.S.; Kromhout, D. Dietary Epicatechin Intake and 25-y Risk of Cardiovascular Mortality: The Zutphen Elderly Study. Am. J. Clin. Nutr. 2016, 104, 58–64. [Google Scholar] [CrossRef]
- Lajous, M.; Rossignol, E.; Fagherazzi, G.; Perquier, F.; Scalbert, A.; Clavel-Chapelon, F.; Boutron-Ruault, M.C. Flavonoid Intake and Incident Hypertension in Women. Am. J. Clin. Nutr. 2016, 103, 1091–1098. [Google Scholar] [CrossRef]
- Mastroiacovo, D.; Kwik-Uribe, C.; Grassi, D.; Necozione, S.; Raffaele, A.; Pistacchio, L.; Righetti, R.; Bocale, R.; Lechiara, M.C.; Marini, C.; et al. Cocoa Flavanol Consumption Improves Cognitive Function, Blood Pressure Control, and Metabolic Profile in Elderly Subjects: The Cocoa, Cognition, and Aging (CoCoA) Study-A Randomized Controlled Trial. Am. J. Clin. Nutr. 2015, 101, 538–548. [Google Scholar] [CrossRef]
- Ahuja, V.; Miura, K.; Vishnu, A.; Fujiyoshi, A.; Evans, R.; Zaid, M.; Miyagawa, N.; Hisamatsu, T.; Kadota, A.; Okamura, T.; et al. Significant Inverse Association of Equol-Producer Status with Coronary Artery Calcification but Not Dietary Isoflavones in Healthy Japanese Men. Br. J. Nutr. 2017, 117, 260–266. [Google Scholar] [CrossRef]
- Bertoia, M.L.; Rimm, E.B.; Mukamal, K.J.; Hu, F.B.; Willett, W.C.; Cassidy, A. Dietary Flavonoid Intake and Weight Maintenance: Three Prospective Cohorts of 124 086 US Men and Women Followed for up to 24 Years. BMJ 2016, 352, i17. [Google Scholar] [CrossRef]
- Huang, K.; Liang, X.C.; Zhong, Y.L.; He, W.Y.; Wang, Z. 5-Caffeoylquinic Acid Decreases Diet-Induced Obesity in Rats by Modulating PPARα and LXRα Transcription. J. Sci. Food Agric. 2015, 95, 1903–1910. [Google Scholar] [CrossRef]
- Ong, K.W.; Hsu, A.; Tan, B.K.H. Anti-Diabetic and Anti-Lipidemic Effects of Chlorogenic Acid Are Mediated by Ampk Activation. Biochem. Pharmacol. 2013, 85, 1341–1351. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, M.; Liu, D. Chlorogenic Acid Improves High Fat Diet-Induced Hepatic Steatosis and Insulin Resistance in Mice. Pharm. Res. 2015, 32, 1200–1209. [Google Scholar] [CrossRef]
- Jin, S.; Chang, C.; Zhang, L.; Liu, Y.; Huang, X.; Chen, Z. Chlorogenic Acid Improves Late Diabetes through Adiponectin Receptor Signaling Pathways in Db/Db Mice. PLoS ONE 2015, 10, e0120842. [Google Scholar] [CrossRef]
- Wan, C.W.; Wong, C.N.Y.; Pin, W.K.; Wong, M.H.Y.; Kwok, C.Y.; Chan, R.Y.K.; Yu, P.H.F.; Chan, S.W. Chlorogenic Acid Exhibits Cholesterol Lowering and Fatty Liver Attenuating Properties by Up-Regulating the Gene Expression of PPAR-α in Hypercholesterolemic Rats Induced with a High-Cholesterol Diet. Phytother. Res. 2013, 27, 545–551. [Google Scholar] [CrossRef]
- Bondonno, C.P.; Mubarak, A.; Liu, A.H.; Considine, M.J.; Rich, L.; Mas, E.; Croft, K.D.; Hodgson, J.M. Acute Effects of Chlorogenic Acid on Nitric Oxide Status, Endothelial Function and Blood Pressure in Healthy Volunteers: A Randomised Trial. Free Radic. Biol. Med. 2012, 53, 9130–9136. [Google Scholar] [CrossRef]
- Hewlings, S.J.; Kalman, D.S. Curcumin: A Review of Its Effects on Human Health. Foods 2017, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Amalraj, A.; Pius, A.; Gopi, S.; Gopi, S. Biological Activities of Curcuminoids, Other Biomolecules from Turmeric and Their Derivatives—A Review. J. Tradit Complement. Med. 2017, 7, 205–233. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic Roles of Curcumin: Lessons Learned from Clinical Trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef]
- Goel, A.; Kunnumakkara, A.B.; Aggarwal, B.B. Curcumin as “Curecumin”: From Kitchen to Clinic. Biochem. Pharmacol. 2008, 75, 787–809. [Google Scholar] [CrossRef] [PubMed]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A Review of Curcumin and Its Derivatives as Anticancer Agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef]
- Wu, L.Y.; Chen, C.W.; Chen, L.K.; Chou, H.Y.; Chang, C.L.; Juan, C.C. Curcumin Attenuates Adipogenesis by Inducing Preadipocyte Apoptosis and Inhibiting Adipocyte Differentiation. Nutrients 2019, 11, 2307. [Google Scholar] [CrossRef]
- Di Pierro, F.; Bressan, A.; Ranaldi, D.; Rapacioli, G.; Giacomelli, L.; Bertuccioli, A. Potential Role of Bioavailable Curcumin in Weight Loss and Omental Adipose Tissue Decrease: Preliminary Data of a Randomized, Controlled Trial in Overweight People with Metabolic Syndrome. Preliminary Study. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4195–4202. [Google Scholar]
- Panahi, Y.; Hosseini, M.S.; Khalili, N.; Naimi, E.; Majeed, M.; Sahebkar, A. Antioxidant and Anti-Inflammatory Effects of Curcuminoid-Piperine Combination in Subjects with Metabolic Syndrome: A Randomized Controlled Trial and an Updated Meta-Analysis. Clin. Nutr. 2015, 34, 1101–1108. [Google Scholar] [CrossRef]
- El-Moselhy, M.A.; Taye, A.; Sharkawi, S.S.; El-Sisi, S.F.I.; Ahmed, A.F. The Antihyperglycemic Effect of Curcumin in High Fat Diet Fed Rats. Role of TNF-α and Free Fatty Acids. Food Chem. Toxicol. 2011, 49, 1129–1140. [Google Scholar] [CrossRef]
- Thota, R.N.; Acharya, S.H.; Garg, M.L. Curcumin and/or Omega-3 Polyunsaturated Fatty Acids Supplementation Reduces Insulin Resistance and Blood Lipids in Individuals with High Risk of Type 2 Diabetes: A Randomised Controlled Trial. Lipids Health Dis. 2019, 18, 31. [Google Scholar] [CrossRef]
- Thota, R.N.; Rosato, J.I.; Dias, C.B.; Burrows, T.L.; Martins, R.N.; Garg, M.L. Dietary Supplementation with Curcumin Reduce Circulating Levels of Glycogen Synthase Kinase-3Β and Islet Amyloid Polypeptide in Adults with High Risk of Type 2 Diabetes and Alzheimer’s Disease. Nutrients 2020, 12, 1032. [Google Scholar] [CrossRef]
- Bateni, Z.; Rahimi, H.R.; Hedayati, M.; Afsharian, S.; Goudarzi, R.; Sohrab, G. The Effects of Nano-Curcumin Supplementation on Glycemic Control, Blood Pressure, Lipid Profile, and Insulin Resistance in Patients with the Metabolic Syndrome: A Randomized, Double-Blind Clinical Trial. Phytother. Res. 2021, 35, 3945–3953. [Google Scholar] [CrossRef]
- Auger, F.; Martin, F.; Pétrault, O.; Samaillie, J.; Hennebelle, T.; Trabelsi, M.S.; Bailleul, F.; Staels, B.; Bordet, R.; Duriez, P. Risperidone-Induced Metabolic Dysfunction Is Attenuated by Curcuma Longa Extract Administration in Mice. Metab. Brain Dis. 2018, 33, 63–77. [Google Scholar] [CrossRef]
- Kassaian, N.; Feizi, A.; Aminorroaya, A.; Amini, M. Probiotic and Synbiotic Supplementation Could Improve Metabolic Syndrome in Prediabetic Adults: A Randomized Controlled Trial. Diabetes Metab. Syndr. Clin. Res. Rev. 2019, 13, 2991–2996. [Google Scholar] [CrossRef] [PubMed]
- Tenorio-Jiménez, C.; Martínez-Ramírez, M.J.; del Castillo-Codes, I.; Arraiza-Irigoyen, C.; Tercero-Lozano, M.; Camacho, J.; Chueca, N.; García, F.; Olza, J.; Plaza-Díaz, J.; et al. Lactobacillus Reuteri V3401 Reduces Inflammatory Biomarkers and Modifies the Gastrointestinal Microbiome in Adults with Metabolic Syndrome: The PROSIR Study. Nutrients 2019, 11, 1761. [Google Scholar] [CrossRef]
- Cunningham, M.; Azcarate-Peril, M.A.; Barnard, A.; Benoit, V.; Grimaldi, R.; Guyonnet, D.; Holscher, H.D.; Hunter, K.; Manurung, S.; Obis, D.; et al. Shaping the Future of Probiotics and Prebiotics. Trends Microbiol. 2021, 29, 667–685. [Google Scholar] [CrossRef]
- Kootte, R.S.; Levin, E.; Salojärvi, J.; Smits, L.P.; Hartstra, A.V.; Udayappan, S.D.; Hermes, G.; Bouter, K.E.; Koopen, A.M.; Holst, J.J.; et al. Improvement of Insulin Sensitivity after Lean Donor Feces in Metabolic Syndrome Is Driven by Baseline Intestinal Microbiota Composition. Cell Metab. 2017, 26, 611–619. [Google Scholar] [CrossRef]
- Moles, L.; Otaegui, D. The Impact of Diet on Microbiota Evolution and Human Health. Is Diet. an Adequate Tool for Microbiota Modulation? Nutrients 2020, 12, 1654. [Google Scholar] [CrossRef]
- Oriach, C.S.; Robertson, R.C.; Stanton, C.; Cryan, J.F.; Dinan, T.G. Food for Thought: The Role of Nutrition in the Microbiota-Gut-Brain Axis. Clin. Nutr. Exp. 2016, 6, 25–38. [Google Scholar] [CrossRef]
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment Dominates over Host Genetics in Shaping Human Gut Microbiota. Nature 2018, 555, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Costea, P.I.; Hildebrand, F.; Manimozhiyan, A.; Bäckhed, F.; Blaser, M.J.; Bushman, F.D.; de Vos, W.M.; Ehrlich, S.D.; Fraser, C.M.; Hattori, M.; et al. Enterotypes in the Landscape of Gut Microbial Community Composition. Nat. Microbiol. 2017, 3, 8–16. [Google Scholar] [CrossRef]
- Goodrich, J.K.; Davenport, E.R.; Beaumont, M.; Jackson, M.A.; Knight, R.; Ober, C.; Spector, T.D.; Bell, J.T.; Clark, A.G.; Ley, R.E. Genetic Determinants of the Gut Microbiome in UK Twins. Cell Host Microbe 2016, 19, 731–743. [Google Scholar] [CrossRef]
- Garcia-Rios, A.; Gomez-Delgado, F.J.; Garaulet, M.; Alcala-Diaz, J.F.; Delgado-Lista, F.J.; Marin, C.; Rangel-Zuñiga, O.A.; Rodriguez-Cantalejo, F.; Gomez-Luna, P.; Ordovas, J.M.; et al. Beneficial Effect of CLOCK Gene Polymorphism Rs1801260 in Combination with Low-Fat Diet on Insulin Metabolism in the Patients with Metabolic Syndrome. Chronobiol. Int. 2014, 31, 401–408. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Milagro, F.I.; Allayee, H.; Chmurzynska, A.; Choi, M.S.; Curi, R.; de Caterina, R.; Ferguson, L.R.; Goni, L.; Kang, J.X.; et al. Guide for Current Nutrigenetic, Nutrigenomic, and Nutriepigenetic Approaches for Precision Nutrition Involving the Prevention and Management of Chronic Diseases Associated with Obesity. J. Nutrigenet. Nutri. 2017, 10, 43–62. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Experimental Model | Detection Method | Markers for Diagnosis | Metabolites Regulation in MetS or Its Correlated Pathologies | Ref. |
|---|---|---|---|---|
| Human serum (overweight adults) | NMR | CH3 lipids | ↑ | [47] |
| CH2 lipids | ↑ | |||
| CH2-CH= lipids | ↑ | |||
| Lactate | ↑ | |||
| Alanine | ↓ | |||
| Glucose | ↑ | |||
| Choline | ↓ | |||
| Human, Zucker rat, and mice urine (subjects affected by T2D) | NMR | Creatinine | ↓ | [48] |
| N-acetyl group (glycoproteins) | ↓ | |||
| Allantoin | ↓ | |||
| Glutamate | ↓ | |||
| Glutamine | ↓ | |||
| Histidine | ↑ | |||
| BCAA (valine, leucine, | ↑ | |||
| and isoleucine) | ||||
| N-butyrate | ↑ | |||
| Citrate | ↑ | |||
| Lactate | ↑ | |||
| Sprague–Dawley mice urine | NMR | Lactate | ↑ | [49] |
| Acetone/acetoacetate | ↑ | |||
| Pyruvate | ↑ | |||
| Human serum, patients affected by MetS and HUA (hyperuricemia) | NMR | Glutamine, | ↓ | [50] |
| Trimethylamine (TMA) | ↓ | |||
| Isoleucine | ↓ | |||
| Alanine | ↓ | |||
| Lysine | ↓ | |||
| Lipids | ↑ | |||
| 3-Hydroxybutyrate | ↓ | |||
| Glutamate | ↓ | |||
| Glucose | ↑ | |||
| Citrate | ↓ | |||
| Proline | ↓ | |||
| Glycine | ↓ | |||
| Tyrosine | ↓ | |||
| Triglycerides | ↑ | |||
| 1-Methylhistidine | ↓ | |||
| Human plasma (T2D and obesity) | NMR | BCAAs (branched-chain amino acids) | ↑ | [51] |
| AAAs (aromatic amino acids) | ↑ | |||
| Alanine | ↑ | |||
| Isoleucine | ↑ | |||
| Phenylalanine | ↑ | |||
| Tyrosine | ↑ | |||
| Glutamate/glutamine | ↑ | |||
| Aspartate/asparagine | ↑ | |||
| Arginine | ↑ | |||
| Tryptophan | ↑ | |||
| α-Methyl butyryl carnitine | ↑ | |||
| Iso valeryl carnitine | ↑ | |||
| α-Hydroxybutyrate | ↑ | |||
| Glycine | ↓ | |||
| Betaine | ↓ | |||
| Acylcarnitine | ↑ | |||
| Long-chain FAs | ↑ | |||
| Five-week-old male Sprague–Dawley rats’ urine | NMR | Acetate | ↑ | [52] |
| Leucine | ↑ | |||
| Lysine | ↑ | |||
| Glucose | ↑ | |||
| Citrate | ↓ | |||
| 2-Oxoglutarate | ↓ | |||
| Hippurate | ↓ | |||
| Allantoin | ↑ | |||
| Creatinine | ↓ | |||
| Trigonelline | ↓ | |||
| Tryptophan (TRP) | ↓ | |||
| 3-Hydroxybutyrate (3-HB) | ↑ | |||
| Dimethylamine (DMA) | ↓ | |||
| Succinate | ↓ | |||
| Acetoacetate | ↓ | |||
| Human plasma (patients affected by T2D and CVD) | NMR | Valine | ↑ | [53] |
| Leucine | ↑ | |||
| Isoleucine | ↑ | |||
| Human serum (patients affected by obesity, IR, and T2D) | NMR | Isoleucine | ↑ | [54] |
| Leucine | ↑ | |||
| Valine | ↑ | |||
| Phenylalanine | ↑ | |||
| Tyrosine | ↑ | |||
| Alanine | ↑ | |||
| Histidine | ↓ | |||
| Glutamine | ↓ | |||
| Human serum (IR, glycemia, and T2D) | NMR | Alanine | ↑ | [55] |
| Lactate | ↑ | |||
| Pyruvate | ↑ | |||
| Tyrosine | ↑ | |||
| BCAAs | ↑ | |||
| Leucine | ↑ | |||
| Isoleucine | ↑ | |||
| Valine | ↑ | |||
| Phenylalanine | ↑ | |||
| AAAs | ↑ | |||
| Male Sprague Dawley rats urine | NMR | Creatinine | ↑ | [56] |
| Creatine | ↑ | |||
| Arginine | ↑ | |||
| Aspartate | ↑ | |||
| C57BL/6J (B6) and leptin-deficient ob/ob mice (obese) serum and urine | NMR | Acetoacetate, acetone, citrate, fumarate, | [57] | |
| 2-oxoglutarate, succinate, trimethylamine (TMA), and | ||||
| 3-hydroxybutyrate are up-regulated | ||||
| for urine sample and acetoacetate, | ||||
| acetone, succinate, carnitine, VLDL/LDL cholesterol, and | ||||
| TMAO for serum | ||||
| Human serum (overweight and patients with obesity and with or without MetS) | NMR | BCAAs | ↑ | [58] |
| AAAs | ↑ | |||
| Orosomucoid and fatty acids | ↑ | |||
| Human plasma (patients affected by MetS) | LC/GC-MS | Hydroxypalmitic acid | ↓ | [59] |
| Cholesterol | ↓ | |||
| Sphingosine-1-phosphate | ↓ | |||
| Lactic acid | ↑ | |||
| Alanine | ↑ | |||
| Cysteine | ↑ | |||
| Lysine | ↑ | |||
| Cystine | ↑ | |||
| Glutamic acid | ↑ | |||
| Valine | ↑ | |||
| Proline | ↑ | |||
| Aspartic acid | ↑ | |||
| Tryptophan | ↑ | |||
| Tyrosine | ↑ | |||
| Phenylalanine | ↑ | |||
| Urea | ↑ | |||
| Uric acid | ↑ | |||
| Sorbitol | ↑ | |||
| Human serum (patients affected by MetS) | NMR | C14:0 | ↑ | [60] |
| C16:0 | ↑ | |||
| C18:0 | ↑ | |||
| C18:1n-9c | ↑ | |||
| C18:2n-6c | ↑ | |||
| Human plasma (T2D) | LC-MS | 14 sphingolipids including ceramides (d18:1/18:1, d18:1/20:0, d18:1/20:1, and d18:1/22:1), saturated sphingomyelins (SMs) (C34:0, C38:0, and C40:0), unsaturated SMs (C34:1, C36:1, and C42:3), and hydroxyl-SMs (C34:1, C38:3) are positively associated with incident T2D | [61] | |
| Human plasma (T2D, obesity and MetS) | LC | Ceramides (d18:1/16:0, d18:1/18:0, d18:1/20:0, d18:1/22:0, and d18:1/24:0) and SMs (d18:1/18:0, d18:1/18:1, and, d18:1/20:0) are associated with obesity and MetS | [62] | |
| Ceramides (d18:1/24:1) are associated with triglyceride change | ||||
| SMs (C36:0 and d18:0/24:0) are associated with glucose change | ||||
| Ceramides (C18:0, C20:0, and C24:1) are associated with cardiovascular disease and T2D | ||||
| Human serum (MetS, T2D, and CVD) | GC-MS | 2-Hydroxybutyric acid | ↑ | [63] |
| Inositol | ↑ | |||
| D-glucose | ↑ | |||
| Human urine (MetS) | LC-MS | Metabolites in patients with MetS with respect to control | [64] | |
| Indole-3-acetic acid | ||||
| Indole-3-acetic acid-O-glucuronide | ||||
| N-(indol-3-ylacetyl) glutamine Indole-3-carbaldehyde | ||||
| Hydroxyhexanoycarnitine | ||||
| Indole-3-carboxylic acid | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ambroselli, D.; Masciulli, F.; Romano, E.; Catanzaro, G.; Besharat, Z.M.; Massari, M.C.; Ferretti, E.; Migliaccio, S.; Izzo, L.; Ritieni, A.; et al. New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food. Nutrients 2023, 15, 640. https://doi.org/10.3390/nu15030640
Ambroselli D, Masciulli F, Romano E, Catanzaro G, Besharat ZM, Massari MC, Ferretti E, Migliaccio S, Izzo L, Ritieni A, et al. New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food. Nutrients. 2023; 15(3):640. https://doi.org/10.3390/nu15030640
Chicago/Turabian StyleAmbroselli, Donatella, Fabrizio Masciulli, Enrico Romano, Giuseppina Catanzaro, Zein Mersini Besharat, Maria Chiara Massari, Elisabetta Ferretti, Silvia Migliaccio, Luana Izzo, Alberto Ritieni, and et al. 2023. "New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food" Nutrients 15, no. 3: 640. https://doi.org/10.3390/nu15030640
APA StyleAmbroselli, D., Masciulli, F., Romano, E., Catanzaro, G., Besharat, Z. M., Massari, M. C., Ferretti, E., Migliaccio, S., Izzo, L., Ritieni, A., Grosso, M., Formichi, C., Dotta, F., Frigerio, F., Barbiera, E., Giusti, A. M., Ingallina, C., & Mannina, L. (2023). New Advances in Metabolic Syndrome, from Prevention to Treatment: The Role of Diet and Food. Nutrients, 15(3), 640. https://doi.org/10.3390/nu15030640