
Protective Effect of Anthocyanins against Neurodegenerative Diseases through the Microbial-Intestinal-Brain Axis: A Critical Review


 and
and
Abstract
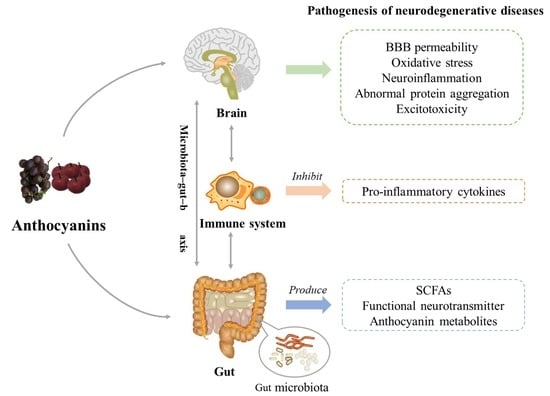
1. Introduction
2. Protective Effects of Anthocyanins on Neurodegenerative Diseases
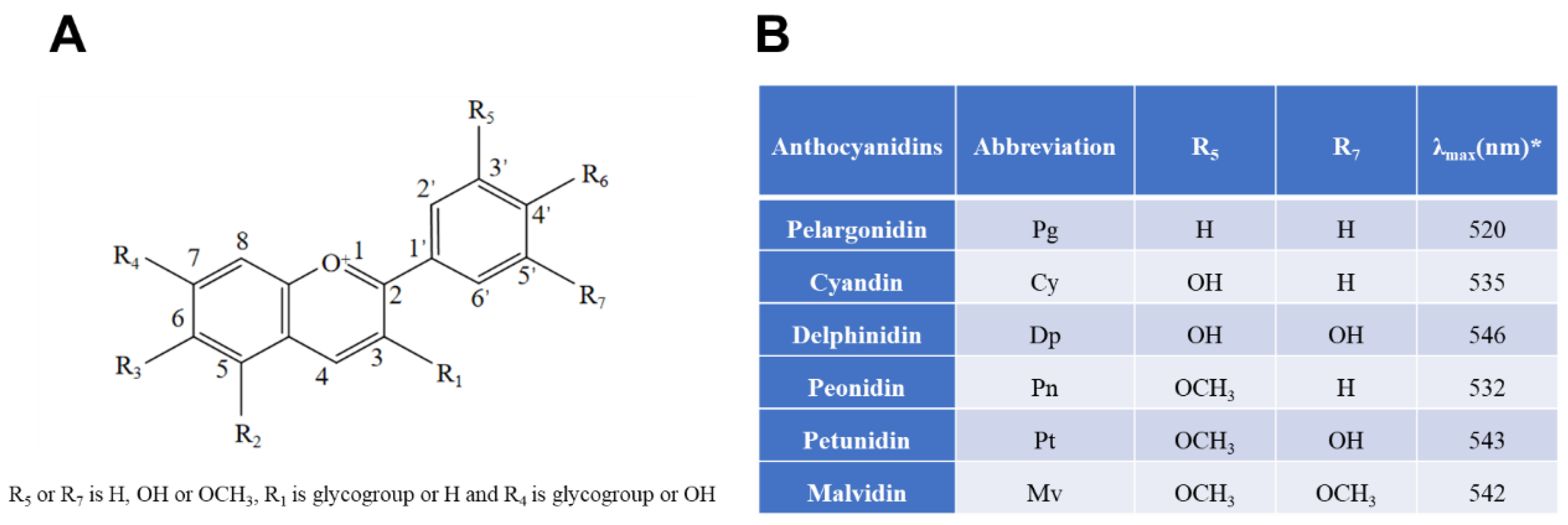
2.1. Neuroprotective Effects of Anthocyanins on Neurotoxicities Induced by Oxidative Stress and Neuroinflammatory Response
2.2. The Penetration of Anthocyanins through the Blood-Brain Barrier
3. Interactions between Anthocyanins and Gut Microbiota
3.1. Metabolism of Anthocyanins by Gut Microbiota
3.2. Modulation of Gut Microbiota by Anthocyanins
3.3. Physiological Activity Related to Modulation of Microbiota
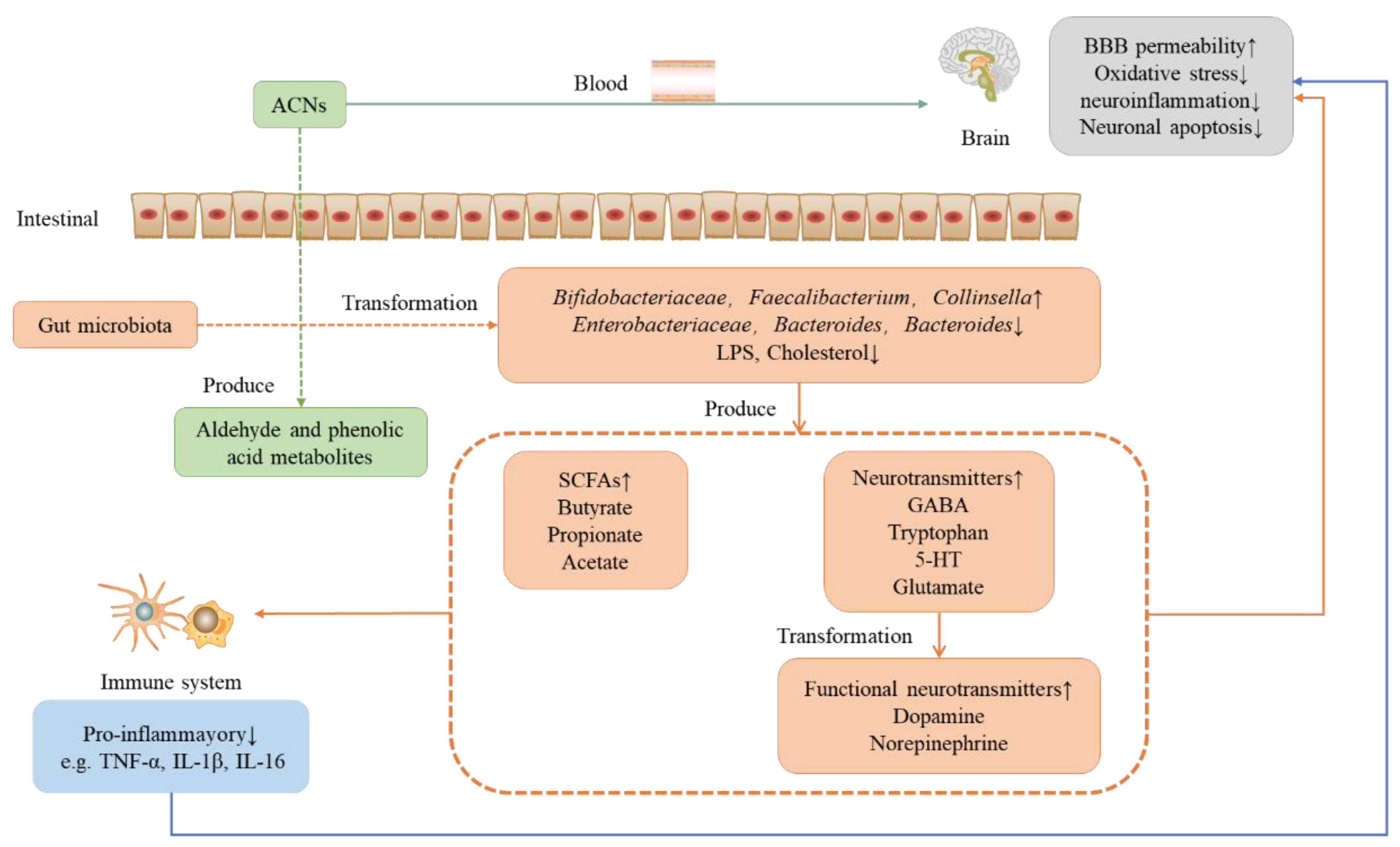
4. Protective Effect of Anthocyanins on Neurodegenerative Diseases under the Microbial-Gut-Brain Axis System
4.1. The Microbiota-Gut-Brain Axis
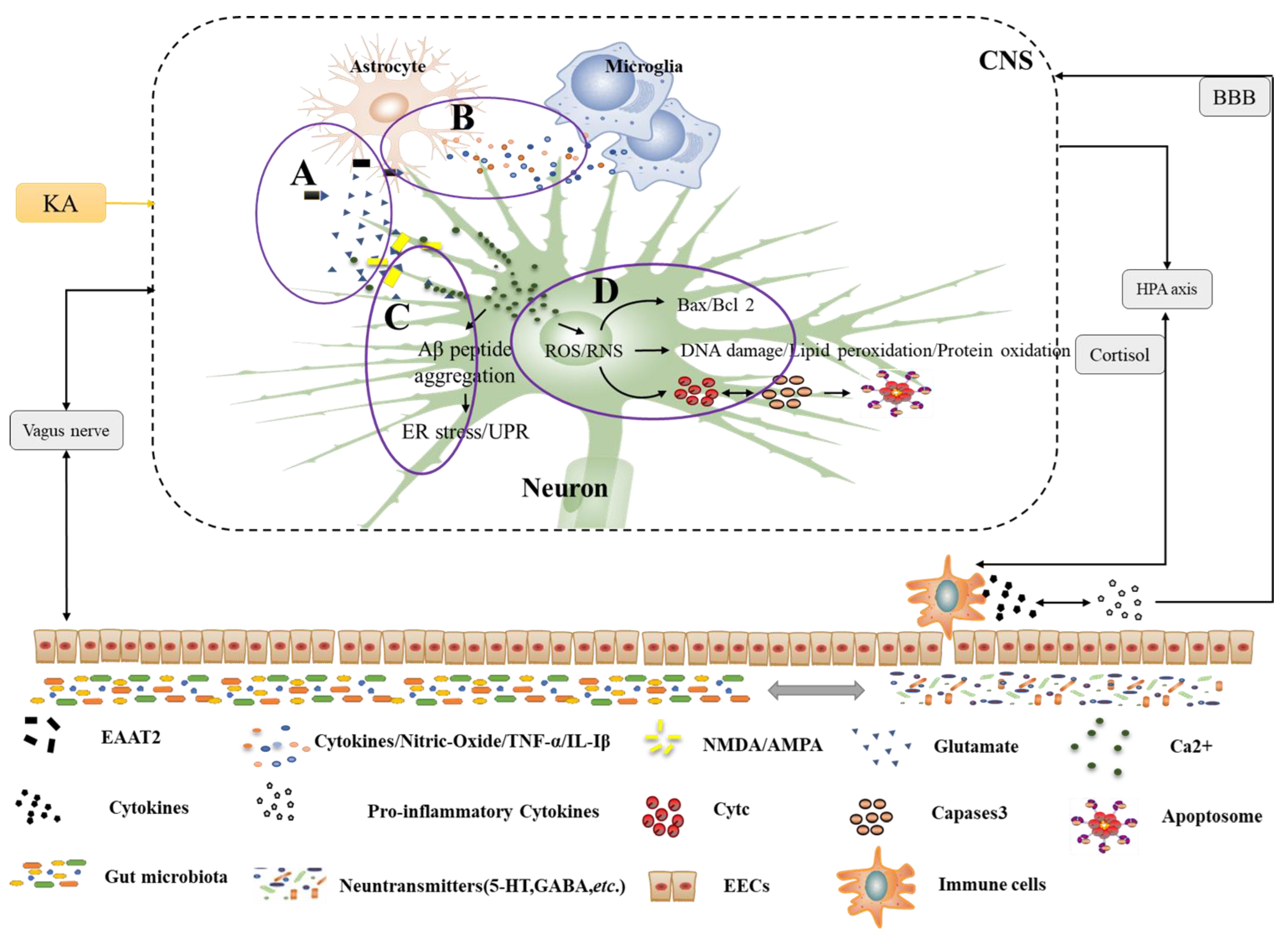
4.1.1. Neuronal Pathways
4.1.2. Neural Immune Pathways
4.1.3. Chemical Signaling Pathways
4.2. The Role of the Microbiota-Gut-Brain Axis in Neuropsychiatric Disorders
4.2.1. Alzheimer Disease
4.2.2. Parkinson’s Disease
4.2.3. Autism Spectrum Disorder
4.2.4. Anxiety and Depression
4.2.5. Schizophrenia
4.2.6. Bipolar Disorder
4.3. The Protective Effect of Anthocyanins on the Nervous System under the Microbial-Entero-Brain Axis System
5. Conclusions and Expectations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rashid, K.; Wachira, F.N.; Nyabuga, J.N.; Wanyonyi, B.; Murilla, G.; Isaac, A.O. Kenyan purple tea anthocyanins ability to cross the blood brain barrier and reinforce brain antioxidant capacity in mice. Nutr. Neurosci. 2014, 17, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Du, J.; Du, L.; Luo, Q.; Xiong, J. Anti-fatigue activity of purified anthocyanins prepared from purple passion fruit (P. edulis Sim) epicarp in mice. J. Funct. Foods 2020, 65, 103725. [Google Scholar] [CrossRef]
- Szymanowska, U.; Baraniak, B. Antioxidant and Potentially Anti-Inflammatory Activity of Anthocyanin Fractions from Pomace Obtained from Enzymatically Treated Raspberries. Antioxidants 2019, 8, 299. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tikunov, Y.; Schouten, R.E.; Marcelis, L.F.M.; Visser, R.G.F.; Bovy, A. Anthocyanin Biosynthesis and Degradation Mechanisms in Solanaceous Vegetables: A Review. Front. Chem. 2018, 6, 52. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Giusti, M. Anthocyanins: Natural Colorants with Health-Promoting Properties. Annu. Rev. Food Sci. Technol. 2010, 1, 163–187. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Wang, P.; Hu, X.; Fang, C. The gut microbiota: A treasure for human health. Biotechnol. Adv. 2016, 34, 1210–1224. [Google Scholar] [CrossRef]
- Eckburg, P.; Bik, E.; Bernstein, C.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.; Nelson, K.; Relman, D. Diversity of the Human Intestinal Microbial Flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, B.; Wu, J.; Jiang, X.; Tang, H. Modulation of Gut Microbiota in Pathological States. Engineering 2017, 3, 83–89. [Google Scholar] [CrossRef]
- Joseph, S.; Curtis, M.A. Microbial transitions from health to disease. Periodontology 2021, 86, 201–209. [Google Scholar] [CrossRef]
- Agirman, G.; Hsiao, E.Y. SnapShot: The microbiota-gut-brain axis. Cell 2021, 184, 2524. [Google Scholar] [CrossRef]
- Fröhlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jačan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Fröhlich, E.; et al. Cognitive impairment by antibiotic-induced gut dysbiosis: Analysis of gut microbiota-brain communication. Brain Behav. Immun. 2016, 56, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Sang, H.; Pothoulakis, C.; Mayer, E. Principles and clinical implications of the brain-gut-enteric microbiota axis. Nat. Rev. Gastroenterol. Hepatol. 2009, 6, 306–314. [Google Scholar] [CrossRef]
- Cryan, J.; Mahony, S. The microbiome-gut-brain axis: From bowel to behavior. Neurogastroenterol. Motil. 2011, 23, 187–192. [Google Scholar] [CrossRef]
- Long-Smith, C.; O’Riordan, K.J.; Clarke, G.; Stanton, C.; Cryan, J.F. Microbiota-Gut-Brain Axis: New Therapeutic Opportunities. Annu. Rev. Pharmacol. 2020, 60, 477–502. [Google Scholar] [CrossRef]
- Radi, E.; Formichi, P.; Battisti, C.; Federico, A. Apoptosis and Oxidative Stress in Neurodegenerative Diseases. J. Alzheimer Dis. 2014, 42, S125. [Google Scholar] [CrossRef] [PubMed]
- Winter, A.N.; Brenner, M.C.; Noelle, P.; Michael, S.; Caleb, B.; Yingyot, A.; Linseman, D.A. Comparison of the Neuroprotective and Anti-Inflammatory Effects of the Anthocyanin Metabolites, Protocatechuic Acid and 4-Hydroxybenzoic Acid. Oxid. Med. Cell Longev. 2017, 2017, 6297080. [Google Scholar] [CrossRef]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kamil, K.K. Redox- and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef]
- Subhramanyam, C.S.; Wang, C.; Hu, Q.; Dheen, S.T. Microglia-mediated neuroinflammation in neurodegenerative diseases. Semin. Cell Dev. Biol. 2019, 94, 112–120. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Zhang, X.; Qu, S. Glutamate, Glutamate Transporters, and Circadian Rhythm Sleep Disorders in Neurodegenerative Diseases. ACS Chem. Neurosci. 2018, 10, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.W.; Lin, T.Y.; Wang, S.J. 11-Keto-β-boswellic acid attenuates glutamate release and kainic acid-induced excitotoxicity in the rat hippocampus. Planta Med. 2020, 86, 434–441. [Google Scholar] [CrossRef]
- Huo, L.; Du, X.; Li, X.; Liu, S.; Xu, Y. The Emerging Role of Neural Cell-Derived Exosomes in Intercellular Communication in Health and Neurodegenerative Diseases. Front. Neurosci. 2021, 15, 738442. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, A.; Ramassamy, C. Innovative anthocyanin/anthocyanidin formulation protects SK-N-SH cells against the amyloid-β peptide-induced toxicity: Relevance to Alzheimer’s disease. Cent. Nerv. Syst. Agents Med. Chem. (Former. Curr. Med. Chem. -Cent. Nerv. Syst. Agents) 2016, 16, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.; Ali, T.; Kim, M.W.; Jo, M.H.; Chung, J.I.; Kim, M.O. Anthocyanins Improve Hippocampus-Dependent Memory Function and Prevent Neurodegeneration via JNK/Akt/GSK3β Signaling in LPS-Treated Adult Mice. Mol. Neurobiol. 2018, 56, 671–687. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Park, H.Y.; Kim, M.O. Anthocyanins protect against kainic acid-induced excitotoxicity and apoptosis via ROS-activated AMPK pathway in hippocampal neurons. Cns Neurosci. Ther. 2014, 20, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Song, N.; Zhang, L.; Chen, W.; Zhu, H.; Deng, W.; Han, Y.; Guo, J.; Qin, C. Cyanidin 3- O -β-glucopyranoside activates peroxisome proliferator-activated receptor-γ and alleviates cognitive impairment in the APP swe /PS1 ΔE9 mouse model. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2016, 1862, 1786–1800. [Google Scholar] [CrossRef]
- Wei, J.; Wu, H.; Zhang, H.; Fang, L.; Chen, S.; Hou, B. Anthocyanins inhibit high glucose-induced renal tubular cell apoptosis caused by oxidative stress in db/db mice. Int. J. Mol. Med. 2018, 41, 1608–1618. [Google Scholar] [CrossRef]
- Gosselet, F.; Loiola, R.A.; Roig, A.; Rosell, A.; Culot, M. Central nervous system delivery of molecules across the blood-brain barrier. Neurochem. Int. 2021, 144, 104952. [Google Scholar] [CrossRef]
- Talavéra, S.; Felgines, C.; Texier, O.; Besson, C.; Gil-Izquierdo, A.; Lamaison, J.L.; Rémésy, C. Anthocyanin Metabolism in Rats and Their Distribution to Digestive Area, Kidney, and Brain. J. Agric. Food Chem. 2005, 53, 3902–3908. [Google Scholar] [CrossRef]
- Kalt, W.; Blumberg, J.B.; Mcdonald, J.E.; Vinqvist-Tymchuk, M.R.; Fillmore, S.; Graf, B.A.; O’Leary, J.; Milbury, P.E. Identification of anthocyanins in the liver, eye, and brain of blueberry-fed pigs. J. Agric. Food Chem. 2008, 56, 705–712. [Google Scholar] [CrossRef]
- Shimazu, R.; Anada, M.; Miyaguchi, A.; Nomi, Y.; Matsumoto, H. Evaluation of Blood-Brain Barrier Permeability of Polyphenols, Anthocyanins, and Their Metabolites. J. Agric. Food Chem. 2021, 69, 11676–11686. [Google Scholar] [CrossRef]
- Faria, A.; Fernandes, I.; Norberto, S.; Mateus, N.; Calhau, C. Interplay between Anthocyanins and Gut Microbiota. J. Agric. Food Chem. 2014, 62, 6898–6902. [Google Scholar] [CrossRef] [PubMed]
- Ticinesi, A.; Tana, C.; Nouvenne, A.; Prati, B.; Lauretani, F.; Meschi, T. Gut microbiota, cognitive frailty and dementia in older individuals: A systematic review. Clin. Interv. Aging 2018, 13, 1497–1511. [Google Scholar] [CrossRef]
- Gao, K.; Pi, Y.; Mu, C.; Peng, Y.; Huang, Z.; Zhu, W. Antibiotics—Induced modulation of large intestinal microbiota altered aromatic amino acid profile and expression of neurotransmitters in the hypothalamus of piglets. J. Neurochem. 2018, 146, 219–324. [Google Scholar] [CrossRef]
- Jameson, K.G.; Olson, C.A.; Kazmi, S.A.; Hsiao, E.Y. Toward Understanding Microbiome-Neuronal Signaling. Mol. Cell 2020, 78, 577–583. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Wang, L.; Bai, W.; Chen, W.; Chen, F.; Shu, C. Unveiling the Metabolic Modulatory Effect of Anthocyanin and Gut Microbiota Involvement; Springer: Singapore, 2021. [Google Scholar]
- Felgines, C.; Talaveéra, S.v.; Gonthier, M.-P.; Texier, O.; Scalbert, A.; Lamaison, J.-L.; Reémeésy, C. Strawberry anthocyanins are recovered in urine as glucuro-and sulfoconjugates in humans. J. Nutr. 2003, 133, 1296–1301. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Li, Y.; Hou, D.-X.; Wu, S. The Effects and Mechanisms of Cyanidin-3-Glucoside and Its Phenolic Metabolites in Maintaining Intestinal Integrity. Antioxidants 2019, 8, 479. [Google Scholar] [CrossRef]
- Pojer, E.; Mattivi, F.; Johnson, D.; Stockley, C.S. The Case for Anthocyanin Consumption to Promote Human Health: A Review. Compr. Rev. Food Sci. Food Saf. 2013, 12, 483–508. [Google Scholar] [CrossRef]
- Lila, M.A.; Burton-Freeman, B.; Grace, M.; Kalt, W. Unraveling Anthocyanin Bioavailability for Human Health. Annu. Rev. Food Sci. Technol. 2016, 7, 375–393. [Google Scholar] [CrossRef]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef]
- Clemente-Postigo, M.; Queipo-Ortuno, M.I.; Boto-Ordonez, M.; Coin-Araguez, L.; Roca-Rodriguez, M.; Delgado-Lista, J.; Cardona, F.; Andres-Lacueva, C.; Tinahones, F.J. Effect of acute and chronic red wine consumption on lipopolysaccharide concentrations. Am. J. Clin. Nutr. 2013, 97, 1053–1061. [Google Scholar] [CrossRef]
- Fernandes, I.; Faria, A.; Freitas, V.D.; Calhau, C.; Mateus, N. Multiple-approach studies to assess anthocyanin bioavailability. Phytochem. Rev. 2015, 14, 899–919. [Google Scholar] [CrossRef]
- González-Barrio, R.; Edwards, C.A.; Crozier, A. Colonic Catabolism of Ellagitannins, Ellagic Acid, and Raspberry Anthocyanins: In Vivo and In Vitro Studies. Drug Metab. Dispos. Biol. Fate Chem. 2011, 39, 1680. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Q.; Zhao, T.; Zhang, Z.; Mao, G.; Feng, W.; Wu, X.; Yang, L. Biotransformation and metabolism of three mulberry anthocyanin monomers by rat gut microflora. Food Chem. 2017, 237, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Liu, C.; Gao, W.; Li, Z.; Qin, G.; Qi, S.; Jiang, H.; Li, X.; Liu, M.; Yan, F.; et al. Anthocyanins Are Converted into Anthocyanidins and Phenolic Acids and Effectively Absorbed in the Jejunum and Ileum. J. Agric. Food Chem. 2021, 69, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Zhou, W.; Xu, W.; Peng, Y.; Yan, Y.; Lu, L.; Mi, J.; Zeng, X.; Cao, Y. The Main Anthocyanin Monomer from Lycium ruthenicum Murray Fruit Mediates Obesity via Modulating the Gut Microbiota and Improving the Intestinal Barrier. Foods 2021, 11, 98. [Google Scholar] [CrossRef]
- Yu, W.; Gao, J.; Hao, R.; Yang, J.; Wei, J. Effects of simulated digestion on black chokeberry (Aronia melanocarpa (Michx.) Elliot) anthocyanins and intestinal flora. J. Food Sci. Technol. 2021, 58, 1511–1523. [Google Scholar] [CrossRef] [PubMed]
- Coman, M.M.; Oancea, A.M.; Verdenelli, M.C.; Cecchini, C.; Bahrim, G.E.; Orpianesi, C.; Cresci, A.; Silvi, S. Polyphenol content and in vitro evaluation of antioxidant, antimicrobial and prebiotic properties of red fruit extracts. Eur. Food Res. Technol. 2018, 244, 735–745. [Google Scholar] [CrossRef]
- Chen, L.; Jiang, B.; Zhong, C.; Guo, J.; Zhang, L.; Mu, T.; Zhang, Q.; Bi, X. Chemoprevention of colorectal cancer by black raspberry anthocyanins involved the modulation of gut microbiota and SFRP2 demethylation. Carcinogenesis 2018, 39, 471–481. [Google Scholar] [CrossRef]
- Podsędek, A.; Redzynia, M.; Klewicka, E.; Koziołkiewicz, M. Matrix Effects on the Stability and Antioxidant Activity of Red Cabbage Anthocyanins under Simulated Gastrointestinal Digestion. BioMed Res. Int. 2014, 2014, 365738. [Google Scholar] [CrossRef]
- Mayta-Apaza, A.C.; Pottgen, E.; De Bodt, J.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Van de Wiele, T.; Lee, S.-O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, P.; Zhu, Y.; Lou, Q.; He, S. Antioxidant and prebiotic activity of five peonidin-based anthocyanins extracted from purple sweet potato (Ipomoea batatas (L.) Lam.). Sci. Rep. 2018, 8, 5018. [Google Scholar] [CrossRef]
- Peng, Y.; Yan, Y.; Wan, P.; Chen, C.; Chen, D.; Zeng, X.; Cao, Y. Prebiotic effects in vitro of anthocyanins from the fruits of Lycium ruthenicum Murray on gut microbiota compositions of feces from healthy human and patients with inflammatory bowel disease. LWT 2021, 149, 111829. [Google Scholar] [CrossRef]
- Li, S.; Wang, T.; Wu, B.; Fu, W.; Xu, B.; Pamuru, R.R.; Kennett, M.; Vanamala, J.K.P.; Reddivari, L. Anthocyanin-containing purple potatoes ameliorate DSS-induced colitis in mice. J. Nutr. Biochem. 2021, 93, 108616. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Oruna-Concha, M.J.; Kolida, S.; Walton, G.E.; Pascual-Teresa, S.D. Metabolism of Anthocyanins by Human Gut Microflora and Their Influence on Gut Bacterial Growth. J. Agric. Food Chem. 2012, 60, 3882–3890. [Google Scholar] [CrossRef]
- Zheng, F.; Wang, Y.Z.; Wu, Y.X.; Zhang, M.Y.; Li, F.T.; He, Y.; Wen, L.k.; Yue, H. Effect of stabilization malvids anthocyanins on the gut microbiota in mice with oxidative stress. J. Food Biochem. 2021, 45, 4892–4902. [Google Scholar] [CrossRef]
- Zhu, Y.; Sun, H.; He, S.; Lou, Q.; Yu, M.; Tang, M.; Tu, L. Metabolism and prebiotics activity of anthocyanins from black rice (Oryza sativa L.) in vitro. PLoS ONE 2018, 13, e0195754. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Y.; Wu, Z.; Weng, P. The Modulatory Effect of Anthocyanins from Purple Sweet Potato on Human Intestinal Microbiota In Vitro. J. Agric. Food Chem. 2016, 64, 2582–2590. [Google Scholar] [CrossRef]
- Flores, G.; del Castillo, M.L.R.; Costabile, A.; Klee, A.; Guergoletto, K.B.; Gibson, G.R. In vitro fermentation of anthocyanins encapsulated with cyclodextrins: Release, metabolism and influence on gut microbiota growth. J. Funct. Foods 2015, 16, 50–57. [Google Scholar] [CrossRef]
- González-Domínguez, R.; Sayago, A.; Santos-Martín, M.; Fernández-Recamales, Á. High-Throughput Method for Wide-Coverage and Quantitative Phenolic Fingerprinting in Plant-Origin Foods and Urine Samples. J. Agric. Food Chem. 2022, 70, 7796–7804. [Google Scholar] [CrossRef]
- Zhou, L.; Xie, M.; Yang, F.; Liu, J. Antioxidant activity of high purity blueberry anthocyanins and the effects on human intestinal microbiota. LWT 2020, 117, 108621. [Google Scholar] [CrossRef]
- Boto- Ordóñez, M.; Urpi-Sarda, M.; Queipo-Ortu?o, M.I.; Tulipani, S.; Tinahones, F.J.; Andres-Lacueva, C. High levels of Bifidobacteria are associated with increased levels of anthocyanin microbial metabolites: A randomized clinical trial. Food Funct. 2014, 5, 1932–1938. [Google Scholar] [CrossRef]
- Blanco, G.; Ruiz, L.; Ruasmadiedo, P.; Fdezriverola, F.; Margolles, A. Revisiting the Metabolic Capabilities of Bifidobacterium longum susbp. longum and Bifidobacterium longum subsp. infantis from a Glycoside Hydrolase Perspective. Microorganisms 2020, 8, 723. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.S.; Kang, I.; Lee, Y.; Habib, M.A.; Choi, B.J.; Kang, J.S.; Park, D.-S.; Kim, Y.-S. Bifidobacterium longum subsp. infantis YB0411 Inhibits Adipogenesis in 3T3-L1 Pre-adipocytes and Reduces High-Fat-Diet-Induced Obesity in Mice. J. Agric. Food Chem. 2021, 69, 6032–6042. [Google Scholar] [CrossRef] [PubMed]
- Cisowska, A.; Wojnicz, D.; Hendrich, A.B. Anthocyanins as antimicrobial agents of natural plant origin. Nat. Prod. Commun. 2011, 6, 1934578X1100600136. [Google Scholar] [CrossRef]
- Chen, Z.; Chen, X.; Zhai, Y.; Liu, K.; Chen, J.; Yuan, F. Effects of black rice anthocyanins on the behavior and intestinal microbiota of mice with chronic unpredictable mild stress. IOP Conf. Ser. Earth Environ. Sci. 2021, 792, 012005–012006. [Google Scholar] [CrossRef]
- Si, X.; Bi, J.; Chen, Q.; Cui, H.; Bao, Y.; Tian, J.; Shu, C.; Wang, Y.; Tan, H.; Zhang, W.; et al. Effect of Blueberry Anthocyanin-Rich Extracts on Peripheral and Hippocampal Antioxidant Defensiveness: The Analysis of the Serum Fatty Acid Species and Gut Microbiota Profile. J. Agric. Food Chem. 2021, 69, 3658–3666. [Google Scholar] [CrossRef]
- Yan, Y.; Peng, Y.; Tang, J.; Mi, J.; Lu, L.; Li, X.; Ran, L.; Zeng, X.; Cao, Y. Effects of anthocyanins from the fruit of Lycium ruthenicum Murray on intestinal microbiota. J. Funct. Foods 2018, 48, 533–541. [Google Scholar] [CrossRef]
- Hosomi, K.; Kunisawa, J. The Specific Roles of Vitamins in the Regulation of Immunosurveillance and Maintenance of Immunologic Homeostasis in the Gut. Immune Netw. 2017, 17, 13. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Mo, Q.; Liu, T.; Fu, A.; Ruan, S.; Zhong, H.; Tang, J.; Zhao, M.; Li, Y.; Zhu, S.; Cai, H.; et al. Novel Gut Microbiota Patterns Involved in the Attenuation of Dextran Sodium Sulfate-Induced Mouse Colitis Mediated by Glycerol Monolaurate via Inducing Anti-inflammatory Responses. mBio 2021, 12, e02148-21. [Google Scholar] [CrossRef]
- Hair, R.; Sakaki, J.R.; Chun, O.K. Anthocyanins, Microbiome and Health Benefits in Aging. Molecules 2021, 26, 537. [Google Scholar] [CrossRef]
- Feng, Y.; Duan, Y.; Xu, Z.; Na, L.; Zhu, B. An examination of data from the American Gut Project reveals that the dominance of the genus Bifidobacterium is associated with the diversity and robustness of the gut microbiota. MicrobiologyOpen 2019, 8, e939. [Google Scholar] [CrossRef] [PubMed]
- Morais, C.A.; Oyama, L.M.; Conrado, R.D.M.; Rosso, V.D.; Nascimento, C.D.; Pisani, L.P. Polyphenols-rich fruit in maternal diet modulates inflammatory markers and the gut microbiota and improves colonic expression of ZO-1 in offspring. Food Res. Int. 2015, 77, 186–193. [Google Scholar] [CrossRef]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A.; et al. Dietary Flavonoids from Modified Apple Reduce Inflammation Markers and Modulate Gut Microbiota in Mice. J. Nutr. 2013, 144, 146–154. [Google Scholar] [CrossRef]
- Krga, I.; Monfoulet, L.E.; Konic-Ristic, A.; Mercier, S.; Glibetic, M.; Morand, C.; Milenkovic, D. Anthocyanins and their gut metabolites reduce the adhesion of monocyte to TNFα-activated endothelial cells at physiologically relevant concentrations. Arch. Biochem. Biophys. 2016, 599, 51–59. [Google Scholar] [CrossRef]
- Wang, D.; Xia, M.; Yan, X.; Li, D.; Wang, L.; Xu, Y.; Jin, T.; Ling, W. Gut microbiota metabolism of anthocyanin promotes reverse cholesterol transport in mice via repressing miRNA-10b. Circ. Res. 2012, 111, 967. [Google Scholar] [CrossRef]
- Yang, L.A.; Jlf, A.; Ke, Y.; Shj, B.; Ying, G.C. Ameliorative effect of purified anthocyanin from Lycium ruthenicum on atherosclerosis in rats through synergistic modulation of the gut microbiota and NF-κB/SREBP-2 pathways. J. Funct. Foods 2019, 59, 223–233. [Google Scholar] [CrossRef]
- Eisenstein, M. Microbiome: Bacterial broadband. Nature 2016, 533, S104–S106. [Google Scholar] [CrossRef]
- Bogunovic, M.; Davé, S.; Tilstra, J.; Chang, D.; Harpaz, N.; Xiong, H.; Mayer, L.; Plevy, S. Enteroendocrine cells express functional Toll-like receptors. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G1770–G1783. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Fang, F.; Pedersen, N.L.; Tillander, A.; Ludvigsson, J.F.; Ekbom, A. Vagotomy and Parkinson disease A Swedish register-based matched-cohort study. Neurology 2017, 88, 1996–2002. [Google Scholar] [CrossRef]
- Bercik, P.; Park, A.J.; Sinclair, D.; Khoshdel, A.; Lu, J.; Huang, X.; Deng, Y.; Blennerhassett, P.A.; Fahnestock, M.; Moine, D. The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut-brain communication. Neurogastroenterol. Motil. 2011, 23, 1132–1139. [Google Scholar] [CrossRef]
- Wang, S.; Yu, Y.; Adeli, K. Role of Gut Microbiota in Neuroendocrine Regulation of Carbohydrate and Lipid Metabolism via the Microbiota-Gut-Brain-Liver Axis. Microorganisms 2020, 8, 527. [Google Scholar] [CrossRef] [PubMed]
- De Vadder, F.; Grasset, E.; Mannerås Holm, L.; Karsenty, G.; Macpherson, A.J.; Olofsson, L.E.; Bäckhed, F. Gut microbiota regulates maturation of the adult enteric nervous system via enteric serotonin networks. Proc. Natl. Acad. Sci. USA 2018, 115, 6458–6463. [Google Scholar] [CrossRef] [PubMed]
- Diamond, B.; Huerta, P.T.; Tracey, K.; Volpe, B.T. It takes guts to grow a brain: Increasing evidence of the important role of the intestinal microflora in neuro- and immune-modulatory functions during development and adulthood. BioEssays 2011, 33, 588–591. [Google Scholar] [CrossRef] [PubMed]
- Lenz, K.M.; Nelson, L.H. Microglia and Beyond: Innate Immune Cells As Regulators of Brain Development and Behavioral Function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Angelis, A.L.H.D.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Rothhammer, V.; Mascanfroni, I.; Bunse, L.; Takenaka, M.; Kenison, J.; Mayo, L.; Chao, C.-C.; Patel, B.; Yan, R.; Blain, M. Type I interferons and microbial metabolites of tryptophan modulate astrocyte activity and central nervous system inflammation via the aryl hydrocarbon receptor. Nat. Med. 2016, 22, 586–597. [Google Scholar] [CrossRef]
- Powell, N.; Walker, M.M.; Talley, N.J. The mucosal immune system: Master regulator of bidirectional gut-brain communications. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 143–159. [Google Scholar] [CrossRef]
- Logsdon, A.F.; Erickson, M.A.; Rhea, E.M.; Salameh, T.S.; Banks, W.A. Gut reactions: How the blood-brain barrier connects the microbiome and the brain. Exp. Biol. Med. 2017, 243, 159–165. [Google Scholar] [CrossRef]
- Zhao, Y.; Jaber, V.; Lukiw, W.J. Secretory products of the human GI tract microbiome and their potential impact on Alzheimer’s disease (AD): Detection of lipopolysaccharide (LPS) in AD hippocampus. Front. Cell Infect. Microbiol. 2017, 7, 318. [Google Scholar] [CrossRef]
- O’Mahony, L.; Mccarthy, J.; Kelly, P.; Hurley, G.; Luo, F.; Chen, K.; O’Sullivan, G.; Kiely, B.; Collins, J.; Shanahan, F. Lactobacillus and bifidobacterium in irritable bowel syndrome: Symptom responses and relationship to cytokine profiles. Gastroenterology 2005, 128, 541–551. [Google Scholar] [CrossRef]
- Wei, H.; Chadman, K.K.; McCloskey, D.P.; Sheikh, A.M.; Malik, M.; Brown, W.T.; Li, X. Brain IL-6 elevation causes neuronal circuitry imbalances and mediates autism-like behaviors. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2012, 1822, 831–842. [Google Scholar] [CrossRef]
- Deshmukh, H.S.; Liu, Y.; Menkiti, O.R.; Mei, J.; Dai, N.; O’Leary, C.E.; Oliver, P.M.; Kolls, J.K.; Weiser, J.N.; Worthen, G.S. The microbiota regulates neutrophil homeostasis and host resistance to Escherichia coli K1 sepsis in neonatal mice. Nat. Med. 2014, 20, 524–530. [Google Scholar] [CrossRef] [PubMed]
- Stephanie, W.; Sebastian, P.; Claudia, P.; Herbert, R.; Ulrich, B.; Armin, S. The Granulocyte-colony stimulating factor has a dual role in neuronal and vascular plasticity. Front. Cell Dev. Biol. 2015, 3, 48. [Google Scholar] [CrossRef]
- Yang, L.; Lin, H.; Lin, W.; Xu, X. Exercise Ameliorates Insulin Resistance of Type 2 Diabetes through Motivating Short-Chain Fatty Acid-Mediated Skeletal Muscle Cell Autophagy. Biology 2020, 9, 203. [Google Scholar] [CrossRef] [PubMed]
- David, R.-C.; Patricia, R.-M.; Abelardo, M.; Miguel, G.; G, d.l.R.-G.C.; Nuria, S. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Rafiki, A.; Boulland, J.; Halestrap, A.; Ottersen, O.; Bergersen, L. Highly differential expression of the monocarboxylate transporters MCT2 and MCT4 in the developing rat brain. Neuroscience 2003, 122, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Spohn, S.; Mawe, G. Non-conventional features of peripheral serotonin signalling—The gut and beyond. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 412–420. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.; Terekhova, D.; Liu, J.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Mazzoli, R.; Pessione, E. The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef]
- Młynarska, E.; Gadzinowska, J.; Tokarek, J.; Forycka, J.; Szuman, A.; Franczyk, B.; Rysz, J. The Role of the Microbiome-Brain-Gut Axis in the Pathogenesis of Depressive Disorder. Nutrients 2022, 14, 1921. [Google Scholar] [CrossRef]
- Shaaban, S.Y.; El Gendy, Y.G.; Mehanna, N.S.; El-Senousy, W.M.; El-Feki, H.S.A.; Saad, K.; El-Asheer, O.M. The role of probiotics in children with autism spectrum disorder: A prospective, open-label study. Nutr. Neurosci. 2017, 21, 676–681. [Google Scholar] [CrossRef]
- Sudo, N.; Chida, Y.; Aiba, Y.; Sonoda, J.; Oyama, N.; Yu, X.-N.; Kubo, C.; Koga, Y. Postnatal microbial colonization programs the hypothalamic-pituitary-adrenal system for stress response in mice: Commensal microbiota and stress response. J. Physiol. 2004, 558, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Tamtaji, O.R.; Heidari-Soureshjani, R.; Mirhosseini, N.; Kouchaki, E.; Bahmani, F.; Aghadavod, E.; Tajabadi-Ebrahimi, M.; Asemi, Z. Probiotic and selenium co-supplementation, and the effects on clinical, metabolic and genetic status in Alzheimer’s disease: A randomized, double-blind, controlled trial. Clin. Nutr. 2019, 38, 2569–2575. [Google Scholar] [CrossRef]
- Bonfili, L.; Cecarini, V.; Cuccioloni, M.; Angeletti, M.; Berardi, S.; Scarpona, S.; Rossi, G.; Eleuteri, A. SLAB51 Probiotic Formulation Activates SIRT1 Pathway Promoting Antioxidant and Neuroprotective Effects in an AD Mouse Model. Mol. Neurobiol. 2018, 55, 7987–8000. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Taghizadeh, M.; Daneshvar Kakhaki, R.; Kouchaki, E.; Bahmani, F.; Borzabadi, S.; Oryan, S.; Mafi, A.; Asemi, Z. Clinical and metabolic response to probiotic administration in people with Parkinson’s disease: A randomized, double-blind, placebo-controlled trial. Clin. Nutr. 2019, 38, 1031–1035. [Google Scholar] [CrossRef]
- Barichella, M.; Severgnini, M.; Cilia, R.; Cassani, E.; Bolliri, C.; Caronni, S.; Ferri, V.; Cancello, R.; Ceccarani, C.; Faierman, S.; et al. Unraveling gut microbiota in Parkinson’s disease and atypical parkinsonism. Mov. Disord. 2019, 34, 396–405. [Google Scholar] [CrossRef]
- Kang, D.-W.; Adams, J.B.; Gregory, A.C.; Borody, T.; Chittick, L.; Fasano, A.; Khoruts, A.; Geis, E.; Maldonado, J.; McDonough-Means, S.; et al. Microbiota Transfer Therapy alters gut ecosystem and improves gastrointestinal and autism symptoms: An open-label study. Microbiome 2017, 5, 10. [Google Scholar] [CrossRef]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Sheu, J.C.; Venkatachalam, A.; Runge, J.K.; Luna, R.A.; Calarge, C.A. Gut microbiome in adolescent depression. J. Affect. Disord. 2021, 292, 500–507. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Zeng, B.; Liu, M.; Chen, J.; Peng, X. The gut microbiome from patients with schizophrenia modulates the glutamate-glutamine-GABA cycle and schizophrenia-relevant behaviors in mice. Sci. Adv. 2019, 5, eaau8317. [Google Scholar] [CrossRef]
- Severance, E.G.; Gressitt, K.L.; Stallings, C.R.; Katsafanas, E.; Schweinfurth, L.A.; Christina, L.; Adamos, M.B.; Sweeney, K.M.; Origoni, A.E.; Khushalani, S. Probiotic normalization of Candida albicans in schizophrenia: A randomized, placebo-controlled, longitudinal pilot study. Brain Behav. Immun. 2016, 62, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Annamaria, P.; Sabrina, M.; Karl, K.; Bettina, H.; Nina, D.; Susanne, B.; Armin, B.; Frederike, F.; Martina, P.; Robert, Q. A step ahead: Exploring the gut microbiota in inpatients with bipolar disorder during a depressive episode. Bipolar Disord. 2018, 21, 40–49. [Google Scholar] [CrossRef]
- Lu, Q.; Lai, J.; Lu, H.; Ng, C.; Hu, S. Gut Microbiota in Bipolar Depression and Its Relationship to Brain Function: An Advanced Exploration. Front. Psychiatry 2019, 10, 784. [Google Scholar] [CrossRef]
- Alzheimer’s, A. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2019, 15, 321–387. [Google Scholar] [CrossRef]
- Elmira, A.; Zatollah, A.; Reza, D.K.; Fereshteh, B.; Ebrahim, K.; Reza, T.O.; Ali, H.G.; Mahmoud, S. Effect of Probiotic Supplementation on Cognitive Function and Metabolic Status in Alzheimer’s Disease: A Randomized, Double-Blind and Controlled Trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef]
- Cryan, J.; O’Riordan, K.; Cowan, C.; Sandhu, K.; Dinan, T. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Tran, L.; Greenwood-Van, M.B. Age-Associated Remodeling of the Intestinal Epithelial Barrier. J. Gerontol. 2013, 68, 1045–1056. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 2016, 167, 1469–1480. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Zhu, Y.; Zhou, Z.; Jia, X.; Xu, Y.; Yang, Q.; Cui, C.; Shen, Y. Neuroprotective effects of fecal microbiota transplantation on MPTP-induced Parkinson’s disease mice: Gut microbiota, glial reaction and TLR4/TNF-α signaling pathway. Brain Behav. Immun. 2018, 70, 48–60. [Google Scholar] [CrossRef]
- Hasegawa, S.; Goto, S.; Tsuji, H.; Okuno, T.; Asahara, T.; Nomoto, K.; Shibata, A.; Fujisawa, Y.; Minato, T.; Okamoto, A. Intestinal Dysbiosis and Lowered Serum Lipopolysaccharide-Binding Protein in Parkinson’s Disease. PLoS ONE 2015, 10, e0142164. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhou, J. The microbiota-gut-brain axis and its potential therapeutic role in autism spectrum disorder. Neuroscience 2016, 324, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Liang, J.; Liang, M.; Dai, J.; Wang, J.; Luo, Z. Altered Gut Microbiota in Chinese Children With Autism Spectrum Disorders. Front. Cell Infect. Microbiol. 2019, 9, 40. [Google Scholar] [CrossRef]
- Adams, J.B.; Johansen, L.J.; Powell, L.D.; Quig, D.; Rubin, R.A. Gastrointestinal flora and gastrointestinal status in children with autism—Comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 2011, 11, 22. [Google Scholar] [CrossRef]
- Janik, R.; Thomason, L.; Stanisz, A.; Forsythe, P.; Stanisz, G. Magnetic resonance spectroscopy reveals oral Lactobacillus promotion of increases in brain GABA, N-acetyl aspartate and glutamate. NeuroImage 2015, 125, 988–995. [Google Scholar] [CrossRef]
- Kroner, Z. Vitamins and Minerals: Fact Versus Fiction; ABC-CLIO: Santa Barbara, CA, USA, 2018. [Google Scholar]
- Ferrari, A.J.; Charlson, F.J.; Norman, R.E.; Patten, S.B.; Freedman, G.; Murray, C.J.L.; Vos, T.; Whiteford, H.A. Burden of Depressive Disorders by Country, Sex, Age, and Year: Findings from the Global Burden of Disease Study 2010. PLoS Med. 2013, 10, e1001547. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.A.; Baker, G.B.; Dursun, S.M. The Relationship Between the Gut Microbiome-Immune System-Brain Axis and Major Depressive Disorder. Front. Neurol. 2021, 12, 721126. [Google Scholar] [CrossRef]
- Molteni, R.; Macchi, F.; Zecchillo, C.; Dell”Agli, M.; Colombo, E.; Calabrese, F.; Guidotti, G.; Racagni, G.; Riva, M.A. Modulation of the inflammatory response in rats chronically treated with the antidepressant agomelatine. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2013, 23, 1645–1655. [Google Scholar] [CrossRef]
- Naseribafrouei, A.; Hestad, K.; Avershina, E.; Sekelja, M.; Linlkken, A.; Wilson, R.; Rudi, K. Correlation between the human fecal microbiota and depression. Neurogastroenterol. Motil. 2014, 26, 1155–1162. [Google Scholar] [CrossRef]
- Jiang, H.; Ling, Z.; Zhang, Y.; Mao, H.; Ma, Z.; Yin, Y.; Wang, W.; Tang, W.; Tan, Z.; Shi, J. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav. Immun. 2015, 48, 186–194. [Google Scholar] [CrossRef]
- Borelli, C.M.; Solari, H. Schizophrenia. JAMA 2019, 322, 1322. [Google Scholar] [CrossRef] [PubMed]
- Genedi, M.; Janmaat, I.E.; Haarman, B.B.C.M.; Sommer, I.E.C. Dysregulation of the gut-brain axis in schizophrenia and bipolar disorder: Probiotic supplementation as a supportive treatment in psychiatric disorders. Curr. Opin. Psychiatry 2019, 32, 185–195. [Google Scholar] [CrossRef]
- Zhu, F.; Ju, Y.; Wang, W.; Wang, Q.; Ma, X. Metagenome-wide association of gut microbiome features for schizophrenia. Nat. Commun. 2020, 11, 1612–1621. [Google Scholar] [CrossRef] [PubMed]
- Coello, K.; Hansen, T.H.; Sørensen, N.; Munkholm, K.; Kessing, L.V.; Pedersen, O.; Vinberg, M. Gut microbiota composition in patients with newly diagnosed bipolar disorder and their unaffected first-degree relatives. Brain Behav. Immun. 2018, 75, 112–118. [Google Scholar] [CrossRef]
- Marques, C.; Fernandes, I.; Meireles, M.; Faria, A.; Spencer, J.P.; Mateus, N.; Calhau, C. Gut microbiota modulation accounts for the neuroprotective properties of anthocyanins. Sci. Rep. 2018, 8, 11341. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.J.; Cryan, J.F.; Dinan, T.G.; Clarke, G. Kynurenine pathway metabolism and the microbiota-gut-brain axis. Neuropharmacology 2017, 112, 399–412. [Google Scholar] [CrossRef]
- O’Mahony, S.M.; Clarke, G.; Borre, Y.E.; Dinan, T.G.; Cryan, J.F. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav. Brain Res. 2015, 277, 32–48. [Google Scholar] [CrossRef]
- Wu, H.Q.; Pereira, E.; Bruno, J.P.; Pellicciari, R.; Schwarcz, R. The Astrocyte-Derived α7 Nicotinic Receptor Antagonist Kynurenic Acid Controls Extracellular Glutamate Levels in the Prefrontal Cortex. J. Mol. Neurosci. 2010, 40, 204–210. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Whitfield-Cargile, C.M.; Cohen, N.D.; Chapkin, R.S.; Weeks, B.R.; Davidson, L.A.; Goldsby, J.S.; Hunt, C.L.; Steinmeyer, S.H.; Menon, R.; Suchodolski, J.S. The microbiota-derived metabolite indole decreases mucosal inflammation and injury in a murine model of NSAID enteropathy. Gut Microbes 2016, 7, 246–261. [Google Scholar] [CrossRef]
- Stasi, C.; Sadalla, S.; Milani, S. The Relationship Between the Serotonin Metabolism, Gut-Microbiota and the Gut-Brain Axis. Curr. Drug Metab. 2019, 20, 646–655. [Google Scholar] [CrossRef] [PubMed]
- Pan, P.; Lam, V.; Salzman, N.; Huang, Y.W.; Wang, L.S. Black Raspberries and Their Anthocyanin and Fiber Fractions Alter the Composition and Diversity of Gut Microbiota in F-344 Rats. Nutr. Cancer 2017, 69, 943. [Google Scholar] [CrossRef] [PubMed]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. γ-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Liu, J.; Hao, W.; He, Z.; Kwek, E.; Zhu, H.; Ma, N.; Ma, K.Y.; Chen, Z.Y. Blueberry and cranberry anthocyanin extracts reduce bodyweight and modulate gut microbiota in C57BL/6J mice fed with a high-fat diet. Eur. J. Nutr. 2021, 60, 2735–2746. [Google Scholar] [CrossRef]
- Yang, L.L.; Millischer, V.; Rodin, S.; Macfabe, D.F.; Villaescusa, J.C.; Lavebratt, C. Enteric shor—Chain fatty acids promote proliferation of human neural progenitor cells. J. Neurochem. 2020, 154, 635–646. [Google Scholar] [CrossRef]
- Park, J.; Qin, W.; Qi, W.; Yang, M.; Chang, H.K. Bidirectional regulatory potentials of short-chain fatty acids and their G-protein-coupled receptors in autoimmune neuroinflammation. Sci. Rep. 2019, 9, 8837. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, T.J.; Gates, E.J.; Ranger, A.L.; Klegeris, A. Short-chain fatty acids (SCFAs) alone or in combination regulate select immune functions of microglia-like cells. Mol. Cell Neurosci. 2020, 105, 103493. [Google Scholar] [CrossRef] [PubMed]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Toth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Cueva, C.; Alba, C.; Rodriguez, J.M.; de Pascual-Teresa, S.; Jones, J.; Caturla, N.; Victoria Moreno-Arribas, M.; Bartolomé, B. Gut microbiome-modulating properties of a polyphenol-enriched dietary supplement comprised of hibiscus and lemon verbena extracts. Monitoring of phenolic metabolites. J. Funct. Foods 2022, 91, 105016. [Google Scholar] [CrossRef]
- Khan, M.S.; Ikram, M.; Park, J.S.; Park, T.J.; Kim, M.O. Gut microbiota, its role in induction of Alzheimer’s disease pathology, and possible therapeutic interventions: Special focus on anthocyanins. Cells 2020, 9, 853. [Google Scholar] [CrossRef] [PubMed]
- Morissette, A.; Kropp, C.; Songpadith, J.P.; Moreira, R.J.; Marette, A. Blueberry proanthocyanidins and anthocyanins improve metabolic health through a gut microbiota-dependent mechanism in diet-induced obese mice. AJP Endocrinol. Metab. 2020, 318, E965–E980. [Google Scholar] [CrossRef] [PubMed]
- Fox, M.; Knorr, D.A.; Haptonstall, K.M. Alzheimer’s disease and symbiotic microbiota: An evolutionary medicine perspective. Ann. N. York Acad. Sci. 2019, 1449, 3–24. [Google Scholar] [CrossRef] [PubMed]
- Marta, S.; Katarzyna, D.Y.; Satler, D.B.; Donata, K.; Ewa, B.; Jerzy, L. The Gut Microbiome Alterations and Inflammation-Driven Pathogenesis of Alzheimer’s Disease—A Critical Review. Mol. Neurobiol. 2018, 56, 1841–1851. [Google Scholar] [CrossRef]
- Thevaranjan, N.; Puchta, A.; Schulz, C.; Naidoo, A.; Szamosi, J.C.; Verschoor, C.P.; Loukov, D.; Schenck, L.P.; Jury, J.; Foley, K.P.; et al. Age-Associated Microbial Dysbiosis Promotes Intestinal Permeability, Systemic Inflammation, and Macrophage Dysfunction. Cell Host Microbe 2018, 23, 570. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Su, J.; Gao, X.; Yang, H.; Weng, R.; Ni, W.; Gu, Y. The microbiota-gut-brain axis participates in chronic cerebral hypoperfusion by disrupting the metabolism of short-chain fatty acids. Microbiome 2022, 10, 62. [Google Scholar] [CrossRef]
- Hu, R.; Wu, S.; Li, B.; Tan, J.; Yan, J.; Wang, Y.; Tang, Z.; Liu, M.; Fu, C.; Zhang, H.; et al. Dietary ferulic acid and vanillic acid on inflammation, gut barrier function and growth performance in lipopolysaccharide-challenged piglets. Anim. Nutr. 2022, 8, 144–152. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sources | Ac | Subject | Microbiota Sources | Intervention | Microbial Modulation Effects | Ref |
|---|---|---|---|---|---|---|
| Lycium ruthenicum Murray Fruit | Pg | in vivo: mice | Male C57BL/6J mice | standard diet, lipid rich food, HFD, and oral P3G (100 mg/kg body weight). fecal microbiota transplantation | ↑Bifidobacteriaceae, Helicobacteraceae, Deferribacteraceae ↓Firmicutes, Lactobacillaceae, Streptococcaceae, Erysipelotrichaceae | [46] |
| Black chokeberry (Aronia melanocarpa (Michx.) Elliot) | Cy | In vitro | 5 healthy volunteers who had not taken antibiotics for at least three months (3 males and 2 females, aged 22 to 28-years-old) | anthocyanin (1 g/L), tea polyphenol (1 g/L) mixed in growth medium 9 mL and fecal slurry suspension 1mL and adjusted in gastrointestinal digestion | ↑Bacteroides, Bififidobacterium, Blautia, Faecalibacterium ↓Prevotella, Megamonas, Escherichia/Shigella | [47] |
| Plums Italian red grapes Elderberry fruits | Cy | in vitro | strains | Rogosa, Sharpe broth (mMRS) fermentation by L. rhamnosus IMC 501®, L. para-casei IMC 502®, SYNBIO® and L. plantarum IMC 509, the fruit extracts (10 g/L) conduct growth studies CP: MRS broth Medium (containing glucose at a concentration of 10 g/L) | ↑Lactobacillus rhamnosus ↓Bacillus cereus; Staphylococcus aureus; Escherichia coli | [48] |
| Black raspberry | Cy | In vivo: C57BL/6J mice | C57BL/6J mice (five-week-old, 18–20 g) | Normal diet; contains no anthocyanins diet; BRB anthocyanins in the diet, 3.5 µmol/g (LBA), and 7.0 µmol/g (MBA) | ↑Eubacterium rectale; Faecalibacterium prausnitzii; Lactobacillus ↓Desulfovibrio spp.; Enterococcus spp. | [49] |
| Red cabbage | Cy | In vitro | Fecal samples were collected from 14 healthy volunteers (4 males and 10 females, aged 17 to 52-years-old) | Fecal suspension (1:10, W/V) was mixed with an extract rich in anthocyanins and sterile medium to simulated gastrointestinal fermentation | ↓Lactobacillus spp. Clostridium spp. Bacteroides spp. Enterococcus spp. Enterobacteriaceae | [50] |
| Cherries polyphenols | Cy | In vivo In vitro | 10 healthy volunteers who had not taken antibiotics for at least 3 months (5 males and 5 females, aged 23 to 30-years-old) | In vivo: consume 8 oz of juice daily for five days In vitro incubations were performed by mimicking gastric, intestine, and colon conditions | ↑Bacteroides, Collinsella, Firmicutes, Enterobacteriaceae, Bilophila | [51] |
| Purple sweet potato | Pn | In vitro | strains | / | ↑Bifidobacterium spp.; Lactobacillus acidophilus ↓Staphylococcus aureus, Salmonella typhimurium | [52] |
| Lycium ruthenicum Murray | Pt | In vitro | Fecal samples were collected from 4 healthy volunteers and 3 IBD patientshealthy volunteers (3 males and 1 female, aged 20 to 27-years-old) IBD patients (2 males and 1 female, aged 31 to 56-years-old). | Fecal samples were treated to obtain fecal suspension and then mixed with autoclave base growth nutrient medium (Mixed with P3G (1.0 g/L), ACN (1.0 g/L), and glucose (10.0 g/L), respectively) for in vitro fermentation | ↑Collinsella, Bifidobacterium, Streptococcus Lactobacillus plantarum ST-III ↓Escherichia, Shigella | [53] |
| Purple Potato | Pt | In vivo: C57BL/6 mice | C57BL/6 mice (male, four-week-old) | AIN-93G diet, DSS diet, DSS + 15% purple potato in standard diet, DSS + 25% purple potato in standard diet | ↑Bififidobacterium spp., Lactobacillus spp., relative abundance of A. muciniphila | [54] |
| Grape | Mv | In vitro | Fecal samples were collected from 3 healthy volunteers who had not taken antibiotics in at least 6 months. | To mimic colon fermentation, fecal content was incorporated in fecal suspension (1:10) malvidin-3-glucoside (20 and 200 mg/L), gallic acid (150 and 1000 mg/L), and enocianin (4850 and 48,500 mg/L), then injected in the vessels containing fecal slurry. | ↑Bifidobacterium spp. Lactobacillus-Enterococcus spp. ↓Bacteroides spp. Clostridium | [55] |
| Vitis Amurensis Rupr of “Beibinghong” | Mv | In vivo: mice | Feces of experimental male ICR mice | Intraperitoneal injected of normal saline, given D- gal (500 mg/kg bw) once a day, given intragastric administration of malvids anthocyanins and stabilization malvids anthocyanins of 50 mg/kg bw every day, respectively | ↑ Lactobacillus, Alloprevotella ↓ Bacteroides, Alistipes | [56] |
| Black rice | Cy, Pn | In vitro | strains | / | ↑Bifidobacteria; Lactobacillus | [57] |
| Purple sweet potato | Cy, Pg | In vitro | Fecal from 8 healthy volunteers (4 males and 4 females, age 25 to 30-years-old) | Anthocyanin samples or fructooligosaccharide (FOS) (prebiotic) were combined at a final concentration of 1% (w/v) with autoclaved nutrient basal growth medium for in vitro fermentation. | ↑Bifidobacterium spp.; Lactobacillus/ Enterococcus spp. ↓Bacteroides-Prevotella; Clostridium histolyticum | [58] |
| Anthocyanins supplied by Extrasynthese | Cy, Dp | In vitro | Fecal from 3 healthy volunteers, who had not ingested antibiotics for at least 6 months. | Fecal samples were treated to obtain fecal suspension (10%, w/w) and then mixed with sterile medium and encapsulated anthocyanins to obtain fecal slurry (20 mg/L). The positive control was added with prebiotics, while the negative control was not. | ↑Bifidobacteria; Lactobacilli ↓Clostridium histolyticum | [59] |
| Red wine: dealcoholized red Wine | Dp, Mv | In vivo (randomized cross-over-controlled trial (three consecutive periods of 20 days each) | 9 males, age 45 to 50-years-old | The participants were allowed to drink after 20 days abstaining, DRW (272 mL/d, containing 30 g ethanol), or RW (272 mL/d, containing 30 g ethanol), | ↑Bifidobacterium; ↓Enterococcus, Eggerthella lenta | [60] |
| Blueberry | Mv, Pt | In vitro | Taken fecal content from three participants who have not taken antibiotics for 25 weeks | Fecal samples were treated to obtain fecal suspension (10%, w/v) and mixed with high pressure growth nutrient medium with or without BA (10.0 g /L) simulated fermentation. | ↑Actinobacteria, Alloprevotella, Faecalibacterium, Bififidobacterium, Streptococcus | [61] |
| NDs | Subject | Study Design | Species Intervention | Consequences | Ref |
|---|---|---|---|---|---|
| AD | AD patients AD selenium probiotic group (n = 27); age 78.56 ± 8.0 AD selenium group (n = 26); age 78.86 ± 10.2 AD placebo group (n = 26); age 76.26 ± 8.1 | Randomized, double blind, and controlled clinical trial Microbial interventions | Microbial interventions: L. acidophilus, B. bififidum, and B. longum (2 × 109 CFU/g) | Patients with AD had improved cognitive function (increased MMSE scores) and good results for specific inflammatory and oxidative stress markers such as TAC and GSH | [105] |
| AD | Three transgenic mouse models of AD: B6;129-Psen1tm1Mpm Tg (APPSwe, tauP301L) 1Lfa/J (namely, 3xTg-AD) and control wild-type animals. | Analysis of RARβ Acetylation Redox Enzyme Activity Assays Western Blotting Analyses Oxyblot Analysis | SLAB51: Formulation of lactic acid bacteria and Hypertrendella | SLAB51 Activates SIRT1 Pathway in AD Mice, increases antioxidant enzyme activity-SOD, GPX, and reduces peroxidation levels of proteins and lipids | [106] |
| PD | PD patients PD probiotic group (n = 30); age 68.26 ± 7.8 PD placebo group (n = 30); age 67.76 ± 10.2 | Randomized, double-blind, placebo-controlled trialMicrobial interventions | Microbial interventions: L. acidophilus, B. bififidum, L. reuteri, and L. fermentum (each 2 × 109 CFU/g) | PD Patients have a more beneficial MDS-UPDRS score. | [107] |
| PD | PD patients at different stages (n = 237) PSP (n = 22) MSA (n = 22) HC (n = 113) | Prospective observational case-control studies 16S rRNA gene sequencing | / | PD (compared to HC): Lower levels of Lachnospiraceae PSP (compared to PD): Lactobacillaceae similar, Streptococcaceae were reduced. MSA: Lachnospiraceae were not lower, Prevotellaceae were reduced | [108] |
| ASD | ASD patients (n = 30); age 5 to 9-years-old | Prospective, open-label study. Microbial interventions. Evaluation of GI flora using quantitative real-time PCR | Microbial interventions: Lactobacillus acidophilus, Lactobacillus rhamnosus, and Bifidobacteria longum (1 × 108 CFU/g) | In terms of the severity of autism (assessed by ATEC) and gastrointestinal symptoms (assessed by 6-GSI), the colony counts at the level of Bifidobacterium and lactic acid bacteria increased, weight loss decreased, autism levels improved | [103] |
| ASD | ASD patients (n = 18); age 7 to 16-years-old | Open-label clinical trial FMT (Two weeks of antibiotic therapy) and MTT treatment | / | After treatment, the gastrointestinal symptoms of patients were reduced by 80% and ASD behavior symptoms were significantly improved. MTT: ↑Bifidobacterium, Prevotella, Desulfovibrio. All remained improved 8 weeks after treatment ended. | [109] |
| Anxiety/Depression | Male C57BL/6J mice | Prebiotics (fruco-oligosaccharides (Fos) and galacto-oligosaccharides (Gos)) administrationFos, Gos, Fos+Gos for 3 weeks | / | Antidepressant and anxiolytic effect. Reduced stress-induced corticosterone release and promotes the normalization of the intestinal microbiota | [110] |
| Anxiety/Depression | 160 participants:110 MDD patients, 27 healthy controls, and 23 psychiatric controls. Participants (68 males and 92 females, age 20.0 ± 1.9, 29% taking SSRIs) MDD patients (60% in acute episode) | Longitudinal study underwent a diagnostic evaluation and provided a stool sample. 16S rRNA gene sequencing | / | Whether it is in the period of major depression or remission, and whether it is using SSRIs, it is not related to different bacterial composition. | [111] |
| SCZ | Human studies: SCZ patients’ group (n = 63) HC group (n = 69) no significant differences in age, sex, or body mass index Animal studies: GF mice | Human studies: provided a stool sample and 16S rRNA gene sequencing. Animal studies: FMT experiments, behavioral tests, Y-maze | / | The reduction in microbial diversity in SCZ patients, which at the same time led to lower glutamic acid and higher glutamine and GABA in mice, suggests that the SCZ microbiome itself can alter neurochemical and neurological function in ways that may be relevant to SCZ pathology | [112] |
| SCZ | SCZ probiotic group (n = 30), 22 males and 8 females, age 44.66 ± 11.4; SCZ placebo group (n = 26), 15 males and 11 females, age 48.11 ± 9.6 | Longitudinal, double blinded, and placebo controlled | Microbiota intervention | It was found that there was a relationship between candida albicans seropositivity and more serious positive mental symptoms. However, administration of probiotics can normalize the antibody levels of candida albicans. | [113] |
| BD | BD patients’ group (n = 32), 18 males and 14 females, age 41.3 ± 14.7 HC group (n = 10) | Provided a stool sample and 16S rRNA gene | / | An inverse relationship was observed between illness period and micro alpha diversity (r = 0.408, p = 0.07) HC (n = 10) | [114] |
| BD | BD patients’ group (n = 36); HC group (n = 27) no significant differences in age, sex, or body mass index | Rating Scale Assessment for Mood Symptoms Fecal Bacterial Population Determination | BD subjects were treated with quetiapine (300 mg/d) for four weeks. | Quetiapine treatment was effective for depression. After treatment, MARS score decreased, while the levels of rectal eubacteria, Bifidobacterium, and B/E increased. | [115] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, H.; Xu, J.; Yang, M.; Hussain, M.; Liu, X.; Feng, F.; Guan, R. Protective Effect of Anthocyanins against Neurodegenerative Diseases through the Microbial-Intestinal-Brain Axis: A Critical Review. Nutrients 2023, 15, 496. https://doi.org/10.3390/nu15030496
Zhong H, Xu J, Yang M, Hussain M, Liu X, Feng F, Guan R. Protective Effect of Anthocyanins against Neurodegenerative Diseases through the Microbial-Intestinal-Brain Axis: A Critical Review. Nutrients. 2023; 15(3):496. https://doi.org/10.3390/nu15030496
Chicago/Turabian StyleZhong, Hao, Jie Xu, Mengyu Yang, Muhammad Hussain, Xiaofeng Liu, Fengqin Feng, and Rongfa Guan. 2023. "Protective Effect of Anthocyanins against Neurodegenerative Diseases through the Microbial-Intestinal-Brain Axis: A Critical Review" Nutrients 15, no. 3: 496. https://doi.org/10.3390/nu15030496
APA StyleZhong, H., Xu, J., Yang, M., Hussain, M., Liu, X., Feng, F., & Guan, R. (2023). Protective Effect of Anthocyanins against Neurodegenerative Diseases through the Microbial-Intestinal-Brain Axis: A Critical Review. Nutrients, 15(3), 496. https://doi.org/10.3390/nu15030496