
The Integral Role of Magnesium in Muscle Integrity and Aging: A Comprehensive Review
Abstract
1. Introduction
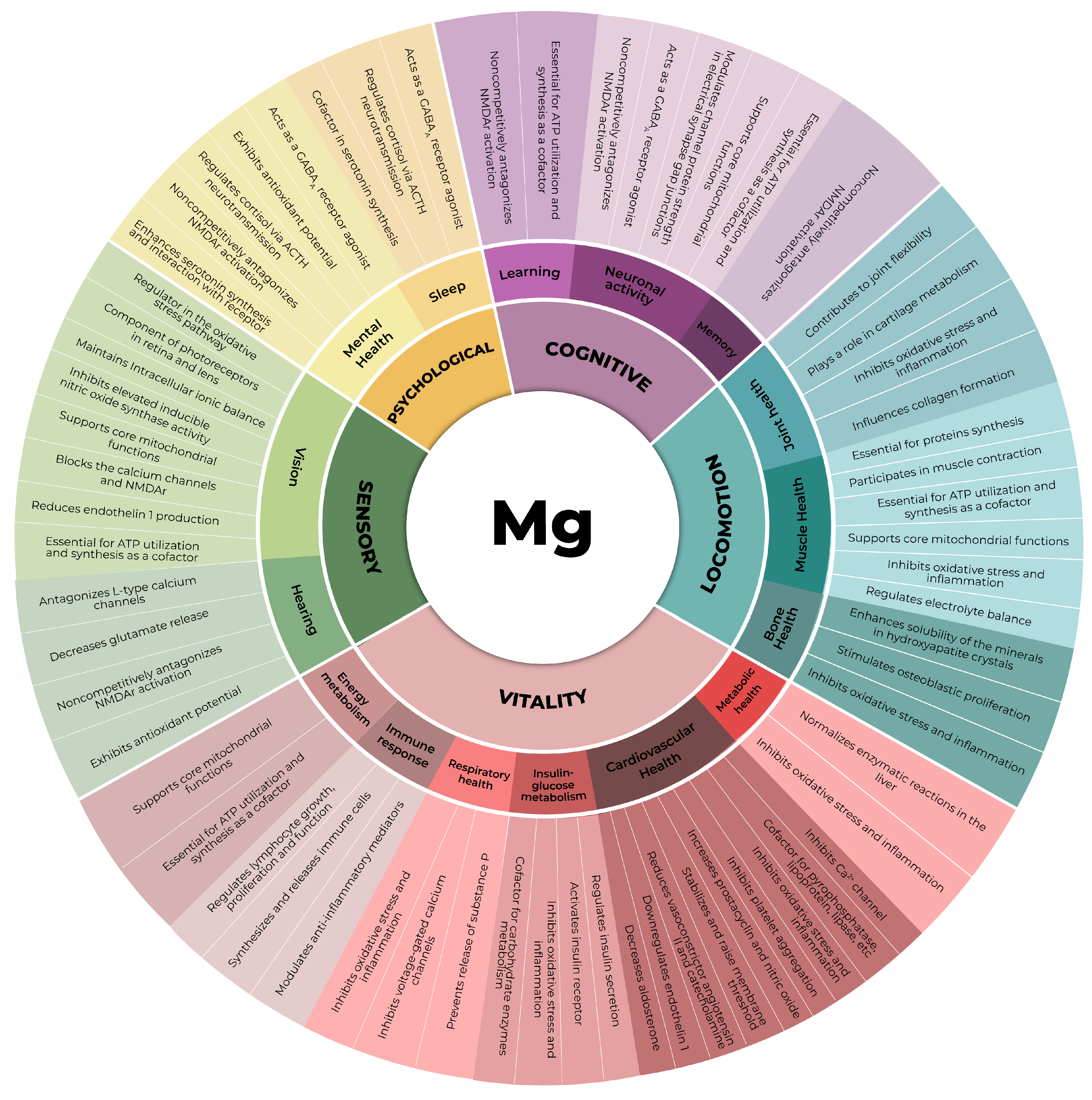
2. Magnesium
3. The Role of Magnesium in Muscle Health
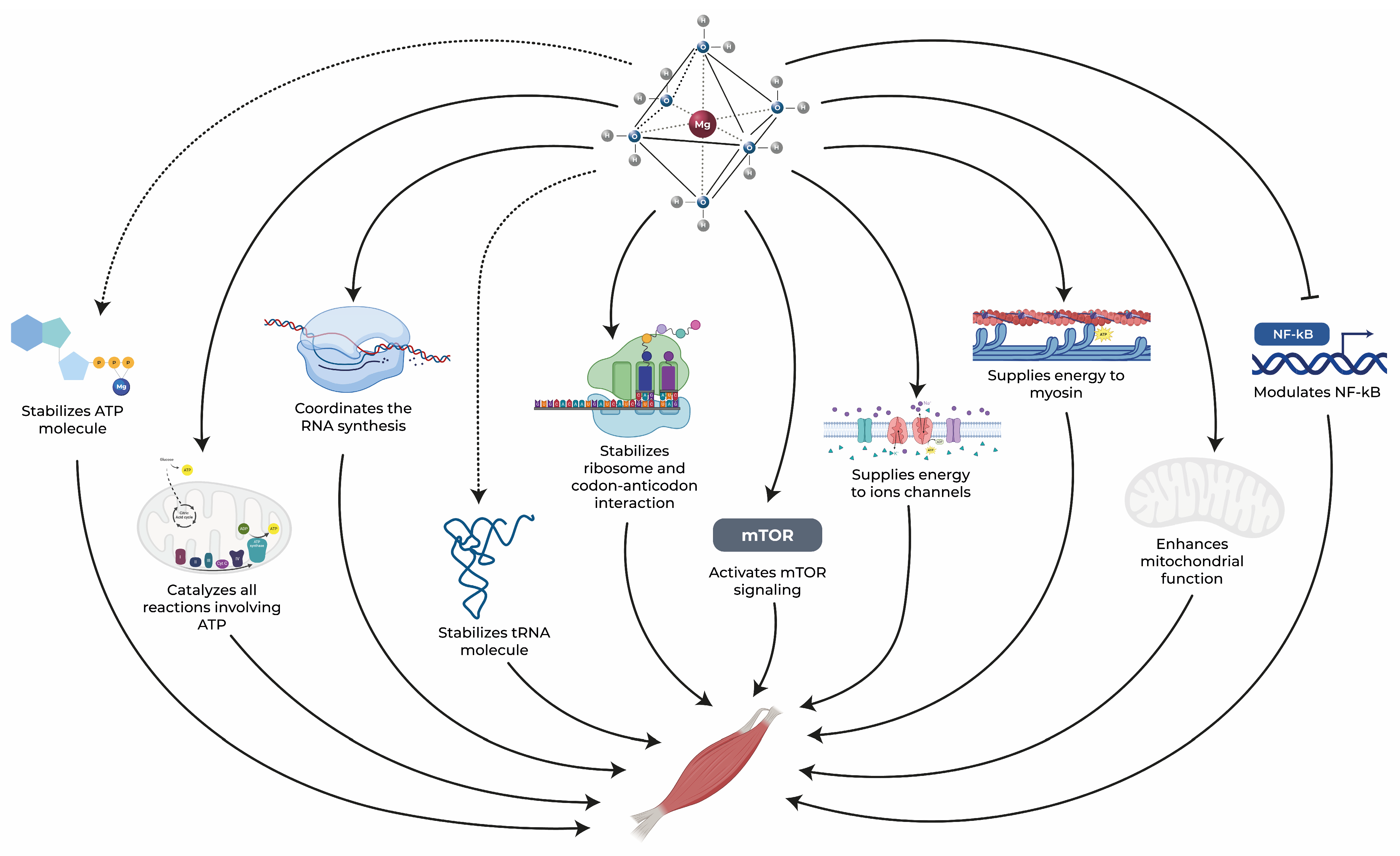
3.1. Energy Metabolism
3.2. Protein Synthesis
3.3. Anti-Inflammatory and Antioxidant Activities
3.4. Muscle Contraction and the Equilibrium of Electrolytes
3.5. Magnesium and Muscle Health: Evidence from Human Studies

{kind=link}
{kind=link}
| Author/Year | Study Design | Subjects (Age) | Intervention Dose and Duration | Source | Main Findings |
|---|---|---|---|---|---|
| (i) Magnesium supplementation in exercise performance | |||||
| Brilla et al., 2003 [86] | Randomized, double-blind, placebo-controlled trial | 35 recreationally active healthy subjects (19–24 y) | Placebo or 800 mg of Mg and 5 g of creatine per day for 2 weeks | MgO + creatine (MgOC) or Mg creatine chelate (MgCC) | MgOC and MgCC showed increases in bw and power. Only MgCC decreased ECW and increased ICW and peak T, suggesting that MgCC affects cellular fluid compartments and may enhance muscle creatine uptake, cellular hydration, and potentially protein synthesis. |
| Brilla and Haley, 1992 [83] | Randomized, double-blind, placebo-controlled trial | 26 untrained subjects (18–30 y) | Mg supplement to achieve an intake of 8 mg/kg bw for 7 weeks | MgO | Significant increase in strength for the Mg group vs. control group, with improvements in absolute quadriceps T, relative T adjusted for bw, and relative T adjusted for LBM. |
| Finstad et al., 2001 [191] | Randomized, double-blind, placebo-controlled, crossover trial | 121 physically active women (21 ± 3 y) | Placebo or 212 mg of Mg for 4 weeks, followed by a 6-week washout period and treatment crossover | MgO | Mg increased resting ionic Mg levels but did not significantly affect performance or recovery indices. |
| Golf et al., 1998 [192] | Randomized, double-blind, placebo-controlled trial | 23 competitive triathletes (29.4 ± 3.3 y) | Placebo or 17 mmol/d of Mg for 4 weeks | Mg orotate | Mg improved performance times in triathlon events, increased serum glucose and higher oxygen uptake, reduced stress responses (lower cortisol levels and leukocyte count), and showed a milder increase in CK post-test, indicating enhanced metabolic efficiency and reduced physiological stress during competition. |
| Kass and Poeira, 2015 [194] | Randomized, double-blind, placebo-controlled, crossover trial | 13 normotensive male and female subjects (38.5 ± 5.3 y) | 300 mg of Mg for 1 week (acute) or 4 weeks (chronic) | Mg citrate | Acute Mg increased bench press performance by 17.7% on day 1, with sustained performance on day 2. Chronic Mg resulted in a 32.1% performance decline on day 2. Both acute and chronic Mg reduced post-exercise SBP on day 2, while only acute Mg reduced DBP. TPR decreased with acute Mg but not with chronic Mg. |
| Kass et al., 2013 [207] | Randomized, controlled, pilot trial | 16 healthy and physically active male subjects (19–24 y) | No treatment or 300 mg of Mg * for 14 days | MgO | Mg reduced resting and post-exercise BP, with a greater effect on resting BP in low-dietary Mg intake individuals and a more pronounced reduction in post-exercise BP in high-dietary Mg intake individuals. No change in performance indicators. |
| Martinez et al., 2017 [211] | Nonrandomized controlled trial | 12 elite (25.3 ± 4.4 y) and 12 recreational (22 ± 3.8 y) male basketball players | No treatment or 400 mg/day of Mg for 32 weeks (measurements in 4 time points, each 8 weeks apart: T1, T2, T3, and T4) | Mg lactate | Serum Mg significantly decreased in T3 but increased after supplementation with T4. Muscle damage markers remained stable, except for creatinine, which decreased post-T2 and then increased in T3 and T4 compared to T2. Mg may prevent tissue damage associated with intense physical activity. |
| Zajac et al., 2020 [199] | Randomized, placebo-controlled trial | 16 elite soccer players (25.6 ± 3.7) | Placebo or 500 mg of Mg (0.07 g/kg bw) for 16 weeks | Mg creatine chelate | Mg improved total time and power in the RAST, enhancing the first and sixth 35 m sprints, with no significant changes in placebo. Mg increased creatinine, lactate, and bicarbonate levels while lowering pH values post-RAST, indicating improved sprint performance and metabolic response. |
| Moslehi et al., 2013 [195] | Randomized, double-blind, placebo-controlled trial | 74 healthy overweight women with BMI 25–30 kg/m² (40–55 y) | Placebo or 250 mg of Mg for 8 weeks | MgO | Mg increased LBM, decreased fat mass, and improved handgrip strength and TGUG vs. baseline. No significant enhancement in knee extension strength. |
| Setaro et al., 2014 [197] | Randomized, double-blind, placebo-controlled trial | 25 professional male volleyball players (Mg: 17.42 ± 1.56 y; C: 17.85 ± 0.99 y) | Placebo or 350 mg of Mg for 4 weeks | MgO | Mg led to decreased lactate production and enhanced plyometric performance, indicating improved alactic anaerobic metabolism. Mg erythrocyte levels, [Mg]U, CK activity, and VO2 max remained within normal ranges. |
| Steward et al., 2019 [197] | Counterbalanced, double-blind, placebo-controlled, crossover study | 9 male recreational runners (27 ± 4 y) | Placebo or 500 mg/day of Mg for 7 days before a 10 km downhill running time trial | MgO + Mg stearate | Mg lowered IL-6 levels, reduced muscle soreness, and improved the recovery of blood glucose post-exercise. No differences in glucose and lactate during the trial or in post-measures of creatine kinase or maximal muscle force. |
| Terblanche et al., 1992 [212] | Double-blind, placebo-controlled trial | 20 marathon runners (25–49 y) | Placebo or 365 mg of Mg for 4 weeks before and 6 weeks after a marathon | Mg-L-aspartate-HCl | In Mg-replete subjects, Mg did not increase muscle or serum Mg concentrations and had no effect on marathon running performance. It also did not influence the extent of muscle damage or the rate of recovery of muscle function post-marathon. |
| Veronese et al., 2014 [208] | Randomized, controlled trial | 139 healthy elderly women (71.5 ± 5.2 y) | No treatment or 300 mg of Mg per day for 12 weeks | MgO | Mg group showed significant improvement in total SPPB score, chair stand times, and 4-m walking speeds vs. control group. No significant differences in secondary outcomes and no serious adverse effects reported. |
| Selsby et al., 2004 [196] | Randomized, double-blind, placebo-controlled trial | 31 weight-trained male subjects (18–24 y) | placebo or 2.5 g of Cr or Mg-creatine chelate (providing 2.5 g of Cr) for 10 days | Mg creatine chelate (MgCC) | Cr and MgCC showed increases in work performed at 70% of 1RM for the bench press vs. placebo. No significant differences between the Cr and MgCC groups in performance tests, suggesting that a low dose of Cr, with or without Mg chelation, can enhance performance. |
| Zorbas et al., 2010 [209] | Randomized, controlled trial | 40 physically healthy male subjects (21.5 ± 3.0 y) | Control subjects (UCS); hypokinetic subjects (UES); control subject + 3.0 mmol of Mg/kg (SCS); hypokinetic subjects + 3.0 mmol of Mg/kg (SES) | MgCl2 | Decreased muscle Mg and increased plasma Mg and urinary/fecal Mg loss in SES and UES groups vs. controls; more pronounced effects in SES. No changes in control groups. |
| (ii) Magnesium supplementation in clinical conditions | |||||
| Aagaard et al., 2005 [190] | Ranadomized, placebo-controlled trial | 59 patients with alcoholic liver disease (34–61 y) | 2 days of infused Mg (30 mmol in 1 L of glucose solution 55 g/L) + 6–7 weeks 12.5 mmol of Mg orally or placebo | MgSO4 (iv) and MgO (oral) | Muscle Mg concentration was 7% higher in the Mg-treated vs. placebo group. Mg had no effect on muscle strength or mass. |
| Gullestad et al., 1992 [193] | Randomized, double-blinded, placebo-controlled trial | 49 chronic alcoholics (28–84 y) | Placebo or 15 mmol Mg for 6 weeks | Mg citrate lactate | Mg significantly reduced liver enzymes (ASAT, ALAT, GOT), slightly increased serum K and Mg, and significantly improved muscle strength. Alcohol consumption remained constant, indicating that the effects were due to Mg supplementation. |
| Bayram et al., 2021 [210] | Prospective, randomized, open-label trial | 209 subjects with acute musculoskeletal low back pain (18–65 y) | NSAID + 365 mg of Mg, NSAID + paracetamol, NSAID | NK | Improvements in functional outcome and musculoskeletal pain intensity from the initial visit to the 4th day with Mg showed greater enhancement than with NSAID alone and NSAID + paracetamol. No significant difference in these improvements or lumbar mobility between groups by the 10th day. |
| Yousef and Al-deeb, 2013 [205] | Randomized, double-blinded, placebo-controlled trial | 80 patients with chronic low back pain (56.4 ± 13.6 y) | Placebo for 6 weeks or iv Mg for 2 weeks followed by oral Mg for 4 weeks. | MgSO4 (iv) and MgO + Mg gluconate (oral) | Significant reduction in pain intensity (7.5 to 4.7) and improvement in lumbar spine flexion, extension, and lateral flexion movements over 6 months in the Mg group. |
| (iii) Magnesium supplementation in nocturnal leg cramps | |||||
| Maor et al., 2017 [206] | Randomized, double-blind, placebo-controlled trial | 88 male and female subjects with NLC (64.9 ± 11.1 y) | Placebo or 520 mg of Mg for 4 weeks | MgO | There was a mean change in weekly NLC of −3.41 in Mg group and −3.03 in placebo group, with no significant difference between groups. No differences were observed in severity and duration of NLC, QoL, or QoS. |
| Dahle et al., 1995 [200] | Randomized, double-blind, placebo-controlled trial | 73 pregnant women with NLC (NK age) | Placebo or 360 mg of Mg for 3 weeks | Mg citrate lactate | Mg decreased NLC vs. baseline and placebo, but did not significantly increase serum Mg levels, excess magnesium being excreted as measured by an increase [Mg]U. |
| Frusso et al., 1999 [201] | Randomized, double-blind, placebo-controlled, crossover trial | 45 male and female subjects with NLC (28–87 y) | Placebo or 1800 mg of Mg for 30 days, followed by a 30-day washout period | Mg citrate | No significant differences between Mg and placebo in any of the evaluated outcomes, including mean number of cramps. |
| Roffe et al., 2002 [202] | Randomized, double-blind, placebo-controlled, crossover trial | 73 male and female subjects with NLC (Mg: 61 ± 11 y, C: 64 ± 10 y) | Placebo or 300 mg of Mg for 6 weeks | Mg citrate | There was a trend towards less NLC on Mg (p = 0.07). No difference in cramp severity and duration between groups. Significantly more subjects thought that the treatment had helped after Mg than after placebo, suggesting that Mg may be effective in NLC. |
| Nygaard et al., 2008 [203] | Randomized, double-blind, placebo-controlled trial | 45 pregnant women with NLC (Mg: 32 ± 4 y, C: 30 ± 3 y) | Placebo or 360 mg of Mg for 2 weeks | Mg citrate lactate | Mg had no significant effect on frequency or intensity of NLC. |
| Supakatisant and Phupong, 2012 [204] | Randomized, double-blind, placebo-controlled trial | 86 pregnant women with NLC (Mg: 29 ± 6 y, C: 29 ± 5 y) | Placebo or 300 mg of Mg for 4 weeks | Mg bisglycinate chelate | Mg resulted in significant lower cramp frequency and intensity vs. placebo. No significant differences in terms of side effects. |
| (iv) Magnesium supplementation and its distribution | |||||
| Wary et al., 1999 [198] | Randomized, double-blind, placebo-controlled trial | 30 young healthy male volunteers (23.7 ± 4.5 y) | Placebo or 12 mmol Mg for 1 month | Mg lactate | Significant change in 24-h [Mg]U after Mg treatment. No differences in other clinical, biological, or Mg status parameters between groups, including intracellular free Mg concentrations of skeletal muscle. |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boss, G.R.; Seegmiller, J.E. Age-related physiological changes and their clinical significance. West. J. Med. 1981, 135, 434–440. [Google Scholar] [PubMed]
- Flint, B.; Tadi, P. Physiology, Aging. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Lee, J.; Kim, H.J. Normal Aging Induces Changes in the Brain and Neurodegeneration Progress: Review of the Structural, Biochemical, Metabolic, Cellular, and Molecular Changes. Front. Aging Neurosci. 2022, 14, 931536. [Google Scholar] [CrossRef] [PubMed]
- WHO. Report of Consortium Meeting 1–2 December 2016—Topic Focus: Frailty and Intrinsic Capacity; World Health Organization: Geneva, Switzerland, 2017; p. 36. [Google Scholar]
- WHO. World Report on Ageing and Health; World Health Organization: Geneva, Switzerland, 2015; p. 246. [Google Scholar]
- WHO. Decade of Healthy Ageing: Baseline Report; World Health Organization: Geneva, Switzerland, 2020; p. 187. [Google Scholar]
- López-Ortiz, S.; Lista, S.; Peñín-Grandes, S.; Pinto-Fraga, J.; Valenzuela, P.L.; Nisticò, R.; Emanuele, E.; Lucia, A.; Santos-Lozano, A. Defining and assessing intrinsic capacity in older people: A systematic review and a proposed scoring system. Ageing Res. Rev. 2022, 79, 101640. [Google Scholar] [CrossRef] [PubMed]
- Leitao, C.; Mignano, A.; Estrela, M.; Fardilha, M.; Figueiras, A.; Roque, F.; Herdeiro, M.T. The Effect of Nutrition on Aging—A Systematic Review Focusing on Aging-Related Biomarkers. Nutrients 2022, 14, 554. [Google Scholar] [CrossRef] [PubMed]
- Bojang, K.P.; Manchana, V. Nutrition and Healthy Aging: A Review. Curr. Nutr. Rep. 2023, 12, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.B.; Silver, R.E.; Das, S.K.; Fielding, R.A.; Gilhooly, C.H.; Jacques, P.F.; Kelly, J.M.; Mason, J.B.; McKeown, N.M.; Reardon, M.A.; et al. Healthy Aging-Nutrition Matters: Start Early and Screen Often. Adv. Nutr. 2021, 12, 1438–1448. [Google Scholar] [CrossRef]
- Bruins, M.J.; Van Dael, P.; Eggersdorfer, M. The Role of Nutrients in Reducing the Risk for Noncommunicable Diseases during Aging. Nutrients 2019, 11, 85. [Google Scholar] [CrossRef]
- Black, M.; Bowman, M. Nutrition and Healthy Aging. Clin. Geriatr. Med. 2020, 36, 655–669. [Google Scholar] [CrossRef]
- Wang, P.; Song, M.; Eliassen, A.H.; Wang, M.; Fung, T.T.; Clinton, S.K.; Rimm, E.B.; Hu, F.B.; Willett, W.C.; Tabung, F.K.; et al. Optimal dietary patterns for prevention of chronic disease. Nat. Med. 2023, 29, 719–728. [Google Scholar] [CrossRef]
- Sanches Machado d’Almeida, K.; Ronchi Spillere, S.; Zuchinali, P.; Correa Souza, G. Mediterranean Diet and Other Dietary Patterns in Primary Prevention of Heart Failure and Changes in Cardiac Function Markers: A Systematic Review. Nutrients 2018, 10, 58. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Veronese, N.; Baiamonte, E.; Guarrera, M.; Parisi, A.; Ruffolo, C.; Tagliaferri, F.; Barbagallo, M. Healthy Aging and Dietary Patterns. Nutrients 2022, 14, 889. [Google Scholar] [CrossRef] [PubMed]
- Śliwińska, S.; Jeziorek, M. The role of nutrition in Alzheimer’s disease. Rocz. Panstw. Zakl. Hig. 2021, 72, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Uusitupa, M.; Khan, T.A.; Viguiliouk, E.; Kahleova, H.; Rivellese, A.A.; Hermansen, K.; Pfeiffer, A.; Thanopoulou, A.; Salas-Salvado, J.; Schwab, U.; et al. Prevention of Type 2 Diabetes by Lifestyle Changes: A Systematic Review and Meta-Analysis. Nutrients 2019, 11, 2611. [Google Scholar] [CrossRef] [PubMed]
- Castro-Espin, C.; Agudo, A. The Role of Diet in Prognosis among Cancer Survivors: A Systematic Review and Meta-Analysis of Dietary Patterns and Diet Interventions. Nutrients 2022, 14, 348. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, M.; Fujita, K.; Nonomura, N. Influence of Diet and Nutrition on Prostate Cancer. Int. J. Mol. Sci. 2020, 21, 1447. [Google Scholar] [CrossRef]
- Fiorentini, D.; Cappadone, C.; Farruggia, G.; Prata, C. Magnesium: Biochemistry, Nutrition, Detection, and Social Impact of Diseases Linked to Its Deficiency. Nutrients 2021, 13, 1136. [Google Scholar] [CrossRef]
- Barbagallo, M.; Veronese, N.; Dominguez, L.J. Magnesium in Aging, Health and Diseases. Nutrients 2021, 13, 463. [Google Scholar] [CrossRef]
- Mathew, A.A.; Panonnummal, R. ‘Magnesium’-the master cation-as a drug-possibilities and evidences. Biometals 2021, 34, 955–986. [Google Scholar] [CrossRef]
- Ajith, T.A. Possible therapeutic effect of magnesium in ocular diseases. J. Basic Clin. Physiol. Pharmacol. 2019, 31, 20190107. [Google Scholar] [CrossRef]
- Cevette, M.J.; Vormann, J.; Franz, K. Magnesium and hearing. J. Am. Acad. Audiol. 2003, 14, 202–212. [Google Scholar] [CrossRef]
- de Baaij, J.H.; Hoenderop, J.G.; Bindels, R.J. Magnesium in man: Implications for health and disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef]
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA receptors in the central nervous system and other organs. Int. Rev. Cytol. 2002, 213, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Murck, H. Magnesium and affective disorders. Nutr. Neurosci. 2002, 5, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Pickering, G.; Mazur, A.; Trousselard, M.; Bienkowski, P.; Yaltsewa, N.; Amessou, M.; Noah, L.; Pouteau, E. Magnesium Status and Stress: The Vicious Circle Concept Revisited. Nutrients 2020, 12, 3672. [Google Scholar] [CrossRef]
- Wolf, F.I.; Cittadini, A. Chemistry and biochemistry of magnesium. Mol. Asp. Med. 2003, 24, 3–9. [Google Scholar] [CrossRef]
- Reddy, S.T.; Soman, S.S.; Yee, J. Magnesium Balance and Measurement. Adv. Chronic Kidney Dis. 2018, 25, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Schwalfenberg, G.K.; Genuis, S.J. The Importance of Magnesium in Clinical Healthcare. Scientifica 2017, 2017, 4179326. [Google Scholar] [CrossRef]
- Struijk, E.A.; Fung, T.T.; Bischoff-Ferrari, H.; Willett, W.C.; Lopez-Garcia, E. Magnesium intake and risk of frailty in older women. In Proceedings of the ASN Nutrition, Boston, MA, USA, 22–25 July 2023. [Google Scholar]
- Reinhart, R.A. Magnesium metabolism. A review with special reference to the relationship between intracellular content and serum levels. Arch. Intern. Med. 1988, 148, 2415–2420. [Google Scholar] [CrossRef]
- Pasternak, K.; Kocot, J.; Horecka, A. Biochemistry of magnesium. J. Elem. 2010, 15, 601–616. [Google Scholar] [CrossRef]
- Pontes, M.H.; Groisman, E.A. Protein synthesis controls phosphate homeostasis. Genes Dev. 2018, 32, 79–92. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, S.; Liu, K.; Abbasi, I.H.R.; Cai, C.; Yao, J. Molecular mechanisms relating to amino acid regulation of protein synthesis. Nutr. Res. Rev. 2019, 32, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Sirago, G.; Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Marzetti, E. Mammalian Target of Rapamycin (mTOR) Signaling at the Crossroad of Muscle Fiber Fate in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 3823. [Google Scholar] [CrossRef]
- Wester, P.O. Electrolyte balance in heart failure and the role for magnesium ions. Am. J. Cardiol. 1992, 70, 44C–49C. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Stubbs, B.; Maggi, S.; Notarnicola, M.; Barbagallo, M.; Firth, J.; Dominguez, L.J.; Caruso, M.G. Dietary Magnesium and Incident Frailty in Older People at Risk for Knee Osteoarthritis: An Eight-Year Longitudinal Study. Nutrients 2017, 9, 1253. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, M.; Belvedere, M.; Dominguez, L.J. Magnesium homeostasis and aging. Magnes. Res. 2009, 22, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Armada, M.J.; Riveiro-Naveira, R.R.; Vaamonde-Garcia, C.; Valcarcel-Ares, M.N. Mitochondrial dysfunction and the inflammatory response. Mitochondrion 2013, 13, 106–118. [Google Scholar] [CrossRef]
- Schuchardt, J.P.; Hahn, A. Intestinal Absorption and Factors Influencing Bioavailability of Magnesium-An Update. Curr. Nutr. Food Sci. 2017, 13, 260–278. [Google Scholar] [CrossRef]
- Dominguez, L.; Veronese, N.; Barbagallo, M. Magnesium and Hypertension in Old Age. Nutrients 2020, 13, 139. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Ketteler, M. Magnesium basics. Clin. Kidney J. 2012, 5, i3–i14. [Google Scholar] [CrossRef]
- Caspi, R.; Billington, R.; Ferrer, L.; Foerster, H.; Fulcher, C.A.; Keseler, I.M.; Kothari, A.; Krummenacker, M.; Latendresse, M.; Mueller, L.A.; et al. The MetaCyc database of metabolic pathways and enzymes and the BioCyc collection of pathway/genome databases. Nucleic Acids Res. 2016, 44, D471–D480. [Google Scholar] [CrossRef]
- Elin, R.J. Assessment of magnesium status for diagnosis and therapy. Magnes. Res. 2010, 23, S194–S198. [Google Scholar] [PubMed]
- Barbagallo, M.; Dominguez, L.J. Magnesium and aging. Curr. Pharm. Des. 2010, 16, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Van Laecke, S. Hypomagnesemia and hypermagnesemia. Acta Clin. Belg. 2019, 74, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Aal-Hamad, A.H.; Al-Alawi, A.M.; Kashoub, M.S.; Falhammar, H. Hypermagnesemia in Clinical Practice. Medicina 2023, 59, 1190. [Google Scholar] [CrossRef] [PubMed]
- Wacker, W.E.C. Magnesium and Man; Harvard University Press: Cambridge, MA, USA, 1980; p. 184. [Google Scholar]
- Bravo, M.; Simón, J.; González-Recio, I.; Martinez-Cruz, L.A.; Goikoetxea-Usandizaga, N.; Martínez-Chantar, M.L. Magnesium and Liver Metabolism Through the Lifespan. Adv. Nutr. 2023, 14, 739–751. [Google Scholar] [CrossRef]
- Romani, A.M. Cellular magnesium homeostasis. Arch. Biochem. Biophys. 2011, 512, 1–23. [Google Scholar] [CrossRef]
- Witkowski, M.; Hubert, J.; Mazur, A. Methods of assessment of magnesium status in humans: A systematic review. Magnes. Res. 2011, 24, 163–180. [Google Scholar] [CrossRef]
- Michalczyk, M.M.; Zajac-Gawlak, I.; Zajac, A.; Pelclova, J.; Roczniok, R.; Langfort, J. Influence of Nutritional Education on the Diet and Nutritional Behaviors of Elderly Women at the University of the Third Age. Int. J. Environ. Res. Public Health 2020, 17, 696. [Google Scholar] [CrossRef]
- Verly Junior, E.; Marchioni, D.M.; Araujo, M.C.; Carli, E.; Oliveira, D.; Yokoo, E.M.; Sichieri, R.; Pereira, R.A. Evolution of energy and nutrient intake in Brazil between 2008–2009 and 2017–2018. Rev. Saude Publica 2021, 55, 5s. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; O’Keefe, J.H.; Wilson, W. Subclinical magnesium deficiency: A principal driver of cardiovascular disease and a public health crisis. Open Heart 2018, 5, e000668. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Barbagallo, M. The Multidomain Nature of Malnutrition in Older Persons. J. Am. Med. Dir. Assoc. 2017, 18, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Rosanoff, A.; Dai, Q.; Shapses, S.A. Essential Nutrient Interactions: Does Low or Suboptimal Magnesium Status Interact with Vitamin D and/or Calcium Status? Adv. Nutr. 2016, 7, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Swaminathan, R. Magnesium metabolism and its disorders. Clin. Biochem. Rev. 2003, 24, 47–66. [Google Scholar] [PubMed]
- Ozsoylu, S.; Hanioğlu, N. Serum magnesium levels in children with vitamin D deficiency rickets. Turk. J. Pediatr. 1977, 19, 89–96. [Google Scholar] [PubMed]
- Uwitonze, A.M.; Razzaque, M.S. Role of Magnesium in Vitamin D Activation and Function. J. Am. Osteopath. Assoc. 2018, 118, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Reddy, V.; Sivakumar, B. Magnesium-dependent vitamin-D-resistant rickets. Lancet 1974, 1, 963–965. [Google Scholar] [CrossRef]
- Erem, S.; Atfi, A.; Razzaque, M.S. Anabolic effects of vitamin D and magnesium in aging bone. J. Steroid Biochem. Mol. Biol. 2019, 193, 105400. [Google Scholar] [CrossRef]
- Venning, G. Recent developments in vitamin D deficiency and muscle weakness among elderly people. BMJ 2005, 330, 524–526. [Google Scholar] [CrossRef]
- Grober, U.; Schmidt, J.; Kisters, K. Magnesium in Prevention and Therapy. Nutrients 2015, 7, 8199–8226. [Google Scholar] [CrossRef]
- Faulkner, J.A.; Larkin, L.M.; Claflin, D.R.; Brooks, S.V. Age-related changes in the structure and function of skeletal muscles. Clin. Exp. Pharmacol. Physiol. 2007, 34, 1091–1096. [Google Scholar] [CrossRef]
- Manrique-Espinoza, B.; Salinas-Rodriguez, A.; Rosas-Carrasco, O.; Gutierrez-Robledo, L.M.; Avila-Funes, J.A. Sarcopenia Is Associated With Physical and Mental Components of Health-Related Quality of Life in Older Adults. J. Am. Med. Dir. Assoc. 2017, 18, 636.e1–636.e5. [Google Scholar] [CrossRef] [PubMed]
- Merchant, R.A.; Chen, M.Z.; Wong, B.L.L.; Ng, S.E.; Shirooka, H.; Lim, J.Y.; Sandrasageran, S.; Morley, J.E. Relationship Between Fear of Falling, Fear-Related Activity Restriction, Frailty, and Sarcopenia. J. Am. Geriatr. Soc. 2020, 68, 2602–2608. [Google Scholar] [CrossRef] [PubMed]
- Brooks, S.V. Current topics for teaching skeletal muscle physiology. Adv. Physiol. Educ. 2003, 27, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Grosicki, G.J.; Zepeda, C.S.; Sundberg, C.W. Single muscle fibre contractile function with ageing. J. Physiol. 2022, 600, 5005–5026. [Google Scholar] [CrossRef] [PubMed]
- Ashique, S.; Kumar, S.; Hussain, A.; Mishra, N.; Garg, A.; Gowda, B.H.J.; Farid, A.; Gupta, G.; Dua, K.; Taghizadeh-Hesary, F. A narrative review on the role of magnesium in immune regulation, inflammation, infectious diseases, and cancer. J. Health Popul. Nutr. 2023, 42, 74. [Google Scholar] [CrossRef] [PubMed]
- Barbagallo, M.; Dominguez, L. Magnesium, Oxidative Stress, and Aging Muscle. In Aging; Academic Press: Cambridge, MA, USA, 2014; pp. 157–166. [Google Scholar] [CrossRef]
- Barclay, C.J. Energy demand and supply in human skeletal muscle. J. Muscle Res. Cell Motil. 2017, 38, 143–155. [Google Scholar] [CrossRef]
- Faria-Pereira, A.; Morais, V.A. Synapses: The Brain’s Energy-Demanding Sites. Int. J. Mol. Sci. 2022, 23, 3627. [Google Scholar] [CrossRef]
- Short, K.R.; Bigelow, M.L.; Kahl, J.; Singh, R.; Coenen-Schimke, J.; Raghavakaimal, S.; Nair, K.S. Decline in skeletal muscle mitochondrial function with aging in humans. Proc. Natl. Acad. Sci. USA 2005, 102, 5618–5623. [Google Scholar] [CrossRef]
- Dirks, A.J.; Hofer, T.; Marzetti, E.; Pahor, M.; Leeuwenburgh, C. Mitochondrial DNA mutations, energy metabolism and apoptosis in aging muscle. Ageing Res. Rev. 2006, 5, 179–195. [Google Scholar] [CrossRef]
- Dominguez, L.J.; Barbagallo, M.; Lauretani, F.; Bandinelli, S.; Bos, A.; Corsi, A.M.; Simonsick, E.M.; Ferrucci, L. Magnesium and muscle performance in older persons: The InCHIANTI study. Am. J. Clin. Nutr. 2006, 84, 419–426. [Google Scholar] [CrossRef]
- Rock, E.; Astier, C.; Lab, C.; Vignon, X.; Gueux, E.; Motta, C.; Rayssiguier, Y. Dietary magnesium deficiency in rats enhances free radical production in skeletal muscle. J. Nutr. 1995, 125, 1205–1210. [Google Scholar] [PubMed]
- Wilson, F.H.; Hariri, A.; Farhi, A.; Zhao, H.; Petersen, K.F.; Toka, H.R.; Nelson-Williams, C.; Raja, K.M.; Kashgarian, M.; Shulman, G.I.; et al. A cluster of metabolic defects caused by mutation in a mitochondrial tRNA. Science 2004, 306, 1190–1194. [Google Scholar] [CrossRef]
- Berg, J.M.T.; Tymoczko, J.L.; Stryer, L. Biochemistry, 5th ed.; W. H. Freeman and Company: New York, NY, USA, 2007. [Google Scholar]
- Dominguez, L.J.; Barbagallo, M.; Di Lorenzo, G.; Drago, A.; Scola, S.; Morici, G.; Caruso, C. Bronchial reactivity and intracellular magnesium: A possible mechanism for the bronchodilating effects of magnesium in asthma. Clin. Sci. 1998, 95, 137–142. [Google Scholar] [CrossRef]
- Saris, N.E.; Mervaala, E.; Karppanen, H.; Khawaja, J.A.; Lewenstam, A. Magnesium: An update on physiological, clinical and analytical aspects. Clin. Chim. Acta 2000, 294, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Brilla, L.R.; Haley, T.F. Effect of magnesium supplementation on strength training in humans. J. Am. Coll. Nutr. 1992, 11, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Brilla, L.; Gunther, K. Effect of magnesium supplementation on exercise time to exhaustion. Med. Exerc. Nutr. Health 1995, 4, 230–233. [Google Scholar]
- Barbagallo, M.; Dominguez, L.J. Magnesium metabolism in type 2 diabetes mellitus, metabolic syndrome and insulin resistance. Arch. Biochem. Biophys. 2007, 458, 40–47. [Google Scholar] [CrossRef]
- Brilla, L.R.; Giroux, M.S.; Taylor, A.; Knutzen, K.M. Magnesium-creatine supplementation effects on body water. Metab. Clin. Exp. 2003, 52, 1136–1140. [Google Scholar] [CrossRef]
- Balch, W.E.; Morimoto, R.I.; Dillin, A.; Kelly, J.W. Adapting proteostasis for disease intervention. Science 2008, 319, 916–919. [Google Scholar] [CrossRef]
- Charmpilas, N.; Daskalaki, I.; Papandreou, M.E.; Tavernarakis, N. Protein synthesis as an integral quality control mechanism during ageing. Ageing Res. Rev. 2015, 23, 75–89. [Google Scholar] [CrossRef]
- Morris, A.L.; Mohiuddin, S.S. Biochemistry, nutrients. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Pontes, M.H.; Sevostyanova, A.; Groisman, E.A. When Too Much ATP Is Bad for Protein Synthesis. J. Mol. Biol. 2015, 427, 2586–2594. [Google Scholar] [CrossRef] [PubMed]
- Hipp, M.S.; Kasturi, P.; Hartl, F.U. The proteostasis network and its decline in ageing. Nat. Rev. Mol. Cell Biol. 2019, 20, 421–435. [Google Scholar] [CrossRef] [PubMed]
- Basisty, N.; Meyer, J.G.; Schilling, B. Protein Turnover in Aging and Longevity. Proteomics 2018, 18, e1700108. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, A.S.; Alexandrov, A.I.; Makarova, N.E.; Gladyshev, V.N.; Dmitriev, S.E. Protein synthesis and quality control in aging. Aging 2018, 10, 4269–4288. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, B.K.; Berger, S.L.; Brunet, A.; Campisi, J.; Cuervo, A.M.; Epel, E.S.; Franceschi, C.; Lithgow, G.J.; Morimoto, R.I.; Pessin, J.E.; et al. Geroscience: Linking aging to chronic disease. Cell 2014, 159, 709–713. [Google Scholar] [CrossRef]
- Douglas, P.M.; Dillin, A. Protein homeostasis and aging in neurodegeneration. J. Cell Biol. 2010, 190, 719–729. [Google Scholar] [CrossRef]
- Vinciguerra, M.; Musaro, A.; Rosenthal, N. Regulation of muscle atrophy in aging and disease. Adv. Exp. Med. Biol. 2010, 694, 211–233. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. Hallmarks of aging: An expanding universe. Cell 2023, 186, 243–278. [Google Scholar] [CrossRef]
- Frontera, W.R.; Ochala, J. Skeletal muscle: A brief review of structure and function. Calcif. Tissue Int. 2015, 96, 183–195. [Google Scholar] [CrossRef]
- Anisimova, A.S.; Meerson, M.B.; Gerashchenko, M.V.; Kulakovskiy, I.V.; Dmitriev, S.E.; Gladyshev, V.N. Multifaceted deregulation of gene expression and protein synthesis with age. Proc. Natl. Acad. Sci. USA 2020, 117, 15581–15590. [Google Scholar] [CrossRef]
- Alber, A.B.; Suter, D.M. Dynamics of protein synthesis and degradation through the cell cycle. Cell Cycle 2019, 18, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.T.; Ang, S.J.; Tsai, S.Y. Sarcopenia: Tilting the Balance of Protein Homeostasis. Proteomics 2020, 20, e1800411. [Google Scholar] [CrossRef] [PubMed]
- Burd, N.A.; Gorissen, S.H.; van Loon, L.J. Anabolic resistance of muscle protein synthesis with aging. Exerc. Sport Sci. Rev. 2013, 41, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Gorissen, S.H.; Pennings, B.; Koopman, R.; Groen, B.B.; Verdijk, L.B.; van Loon, L.J. Aging Is Accompanied by a Blunted Muscle Protein Synthetic Response to Protein Ingestion. PLoS ONE 2015, 10, e0140903. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, S.; Khoutorsky, A.; Mathews, M.B.; Sonenberg, N. Translation deregulation in human disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 791–807. [Google Scholar] [CrossRef] [PubMed]
- Bassler, J.; Hurt, E. Eukaryotic Ribosome Assembly. Annu. Rev. Biochem. 2019, 88, 281–306. [Google Scholar] [CrossRef]
- Akanuma, G. Diverse relationships between metal ions and the ribosome. Biosci. Biotechnol. Biochem. 2021, 85, 1582–1593. [Google Scholar] [CrossRef]
- Schiaffino, S.; Reggiani, C.; Akimoto, T.; Blaauw, B. Molecular Mechanisms of Skeletal Muscle Hypertrophy. J. Neuromuscul. Dis. 2021, 8, 169–183. [Google Scholar] [CrossRef]
- Svetlov, V.; Nudler, E. Basic mechanism of transcription by RNA polymerase II. Biochim. Biophys. Acta 2013, 1829, 20–28. [Google Scholar] [CrossRef][Green Version]
- Petrov, A.S.; Bernier, C.R.; Hsiao, C.; Okafor, C.D.; Tannenbaum, E.; Stern, J.; Gaucher, E.; Schneider, D.; Hud, N.V.; Harvey, S.C.; et al. RNA-magnesium-protein interactions in large ribosomal subunit. J. Phys. Chem. B 2012, 116, 8113–8120. [Google Scholar] [CrossRef]
- Nierhaus, K.H. Mg2+, K+, and the ribosome. J. Bacteriol. 2014, 196, 3817–3819. [Google Scholar] [CrossRef] [PubMed]
- Dever, T.E.; Dinman, J.D.; Green, R. Translation Elongation and Recoding in Eukaryotes. Cold Spring Harb. Perspect. Biol. 2018, 10, a032649. [Google Scholar] [CrossRef] [PubMed]
- Ferriolli, E.; Pessanha, F.P.; Marchesi, J.C. Diabetes and exercise in the elderly. Diabetes Phys. Act. 2014, 60, 122–129. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, Q.; Zhang, Z.; Fu, R.; Zhou, T.; Long, C.; He, T.; Yang, D.; Li, Z.; Peng, S. Magnesium supplementation enhances mTOR signalling to facilitate myogenic differentiation and improve aged muscle performance. Bone 2021, 146, 115886. [Google Scholar] [CrossRef]
- Wang, X.; Proud, C.G. The mTOR pathway in the control of protein synthesis. Physiology 2006, 21, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Garagnani, P.; Parini, P.; Giuliani, C.; Santoro, A. Inflammaging: A new immune-metabolic viewpoint for age-related diseases. Nat. Rev. Endocrinol. 2018, 14, 576–590. [Google Scholar] [CrossRef]
- Franceschi, C.; Bonafe, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- Antuna, E.; Cachan-Vega, C.; Bermejo-Millo, J.C.; Potes, Y.; Caballero, B.; Vega-Naredo, I.; Coto-Montes, A.; Garcia-Gonzalez, C. Inflammaging: Implications in Sarcopenia. Int. J. Mol. Sci. 2022, 23, 5039. [Google Scholar] [CrossRef]
- Meng, S.J.; Yu, L.J. Oxidative stress, molecular inflammation and sarcopenia. Int. J. Mol. Sci. 2010, 11, 1509–1526. [Google Scholar] [CrossRef]
- Meng, Y.; Wu, H.; Yang, Y.; Du, H.; Xia, Y.; Guo, X.; Liu, X.; Li, C.; Niu, K. Relationship of anabolic and catabolic biomarkers with muscle strength and physical performance in older adults: A population-based cross-sectional study. BMC Musculoskelet. Disord. 2015, 16, 202. [Google Scholar] [CrossRef]
- Zembron-Lacny, A.; Dziubek, W.; Wolny-Rokicka, E.; Dabrowska, G.; Wozniewski, M. The Relation of Inflammaging With Skeletal Muscle Properties in Elderly Men. Am. J. Mens Health 2019, 13, 1557988319841934. [Google Scholar] [CrossRef]
- Sanada, F.; Taniyama, Y.; Muratsu, J.; Otsu, R.; Shimizu, H.; Rakugi, H.; Morishita, R. Source of Chronic Inflammation in Aging. Front. Cardiovasc. Med. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Howard, E.E.; Pasiakos, S.M.; Blesso, C.N.; Fussell, M.A.; Rodriguez, N.R. Divergent Roles of Inflammation in Skeletal Muscle Recovery From Injury. Front. Physiol. 2020, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Suetta, C.; Frandsen, U.; Mackey, A.L.; Jensen, L.; Hvid, L.G.; Bayer, M.L.; Petersson, S.J.; Schroder, H.D.; Andersen, J.L.; Aagaard, P.; et al. Ageing is associated with diminished muscle re-growth and myogenic precursor cell expansion early after immobility-induced atrophy in human skeletal muscle. J. Physiol. 2013, 591, 3789–3804. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Zhu, C.F. Causal relationship between insulin resistance and sarcopenia. Diabetol. Metab. Syndr. 2023, 15, 46. [Google Scholar] [CrossRef]
- Webster, J.M.; Kempen, L.; Hardy, R.S.; Langen, R.C.J. Inflammation and Skeletal Muscle Wasting During Cachexia. Front. Physiol. 2020, 11, 597675. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; Li, M.; Chang, M.; Liu, R.; Qiu, J.; Wang, K.; Deng, C.; Shen, Y.; Zhu, J.; Wang, W.; et al. Inflammation: Roles in Skeletal Muscle Atrophy. Antioxidants 2022, 11, 1686. [Google Scholar] [CrossRef]
- Sartori, R.; Romanello, V.; Sandri, M. Mechanisms of muscle atrophy and hypertrophy: Implications in health and disease. Nat. Commun. 2021, 12, 330. [Google Scholar] [CrossRef]
- Merritt, E.K.; Stec, M.J.; Thalacker-Mercer, A.; Windham, S.T.; Cross, J.M.; Shelley, D.P.; Craig Tuggle, S.; Kosek, D.J.; Kim, J.S.; Bamman, M.M. Heightened muscle inflammation susceptibility may impair regenerative capacity in aging humans. J. Appl. Physiol. 2013, 115, 937–948. [Google Scholar] [CrossRef]
- Almoznino-Sarafian, D.; Berman, S.; Mor, A.; Shteinshnaider, M.; Gorelik, O.; Tzur, I.; Alon, I.; Modai, D.; Cohen, N. Magnesium and C-reactive protein in heart failure: An anti-inflammatory effect of magnesium administration? Eur. J. Nutr. 2007, 46, 230–237. [Google Scholar] [CrossRef]
- Simental-Mendia, L.E.; Sahebkar, A.; Rodriguez-Moran, M.; Zambrano-Galvan, G.; Guerrero-Romero, F. Effect of Magnesium Supplementation on Plasma C-reactive Protein Concentrations: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Curr. Pharm. Des. 2017, 23, 4678–4686. [Google Scholar] [CrossRef] [PubMed]
- King, D.E.; Mainous, A.G., 3rd; Geesey, M.E.; Woolson, R.F. Dietary magnesium and C-reactive protein levels. J. Am. Coll. Nutr. 2005, 24, 166–171. [Google Scholar] [CrossRef] [PubMed]
- Steward, C.J.; Zhou, Y.; Keane, G.; Cook, M.D.; Liu, Y.; Cullen, T. One week of magnesium supplementation lowers IL-6, muscle soreness and increases post-exercise blood glucose in response to downhill running. Eur. J. Appl. Physiol. 2019, 119, 2617–2627. [Google Scholar] [CrossRef] [PubMed]
- Chacko, S.A.; Song, Y.; Nathan, L.; Tinker, L.; de Boer, I.H.; Tylavsky, F.; Wallace, R.; Liu, S. Relations of dietary magnesium intake to biomarkers of inflammation and endothelial dysfunction in an ethnically diverse cohort of postmenopausal women. Diabetes Care 2010, 33, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Veronese, N.; Pizzol, D.; Smith, L.; Dominguez, L.J.; Barbagallo, M. Effect of Magnesium Supplementation on Inflammatory Parameters: A Meta-Analysis of Randomized Controlled Trials. Nutrients 2022, 14, 679. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Castiglioni, S.; Locatelli, L.; Zocchi, M.; Mazur, A. Magnesium and inflammation: Advances and perspectives. Semin. Cell Dev. Biol. 2021, 115, 37–44. [Google Scholar] [CrossRef]
- Spasov, A.A.; Iezhitsa, I.N.; Kravchenko, M.S.; Kharitonova, M.V. Study of anti-inflammatory activity of some organic and inorganic magnesium salts in rats fed with magnesium-deficient diet. Vopr. Pitan. 2007, 76, 67–73. [Google Scholar]
- Kharitonova, M.; Iezhitsa, I.; Zheltova, A.; Ozerov, A.; Spasov, A.; Skalny, A. Comparative angioprotective effects of magnesium compounds. J. Trace Elem. Med. Biol. 2015, 29, 227–234. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative stress, inflammation, and cancer: How are they linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Zheltova, A.A.; Kharitonova, M.V.; Iezhitsa, I.N.; Spasov, A.A. Magnesium deficiency and oxidative stress: An update. Biomedicine 2016, 6, 20. [Google Scholar] [CrossRef]
- Gueux, E.; Azais-Braesco, V.; Bussière, L.; Grolier, P.; Mazur, A.; Rayssiguier, Y. Effect of magnesium deficiency on triacylglycerol-rich lipoprotein and tissue susceptibility to peroxidation in relation to vitamin E content. Br. J. Nutr. 1995, 74, 849–856. [Google Scholar] [PubMed]
- Hans, C.P.; Chaudhary, D.P.; Bansal, D.D. Magnesium deficiency increases oxidative stress in rats. Indian J. Exp. Biol. 2002, 40, 1275–1279. [Google Scholar] [PubMed]
- Boparai, R.K.; Kiran, R.; Bansal, D.D. Insinuation of exacerbated oxidative stress in sucrose-fed rats with a low dietary intake of magnesium: Evidence of oxidative damage to proteins. Free Radic. Res. 2007, 41, 981–989. [Google Scholar] [CrossRef]
- Dickens, B.F.; Weglicki, W.B.; Li, Y.S.; Mak, I.T. Magnesium deficiency in vitro enhances free radical-induced intracellular oxidation and cytotoxicity in endothelial cells. FEBS Lett. 1992, 311, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Freedman, A.M.; Mak, I.T.; Stafford, R.E.; Dickens, B.F.; Cassidy, M.M.; Muesing, R.A.; Weglicki, W.B. Erythrocytes from magnesium-deficient hamsters display an enhanced susceptibility to oxidative stress. Am. J. Physiol. 1992, 262, C1371–C1375. [Google Scholar] [CrossRef]
- Weglicki, W.B.; Mak, I.T.; Kramer, J.H.; Dickens, B.F.; Cassidy, M.M.; Stafford, R.E.; Philips, T.M. Role of free radicals and substance P in magnesium deficiency. Cardiovasc. Res. 1996, 31, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Blache, D.; Devaux, S.; Joubert, O.; Loreau, N.; Schneider, M.; Durand, P.; Prost, M.; Gaume, V.; Adrian, M.; Laurant, P.; et al. Long-term moderate magnesium-deficient diet shows relationships between blood pressure, inflammation and oxidant stress defense in aging rats. Free Radic. Biol. Med. 2006, 41, 277–284. [Google Scholar] [CrossRef]
- Calviello, G.; Ricci, P.; Lauro, L.; Palozza, P.; Cittadini, A. Mg deficiency induces mineral content changes and oxidative stress in rats. Biochem. Mol. Biol. Int. 1994, 32, 903–911. [Google Scholar]
- Shivakumar, K.; Kumar, B.P. Magnesium deficiency enhances oxidative stress and collagen synthesis in vivo in the aorta of rats. Int. J. Biochem. Cell Biol. 1997, 29, 1273–1278. [Google Scholar] [CrossRef]
- Kuzniar, A.; Mitura, P.; Kurys, P.; Szymonik-Lesiuk, S.; Florianczyk, B.; Stryjecka-Zimmer, M. The influence of hypomagnesemia on erythrocyte antioxidant enzyme defence system in mice. Biometals 2003, 16, 349–357. [Google Scholar] [CrossRef]
- Mak, I.T.; Komarov, A.M.; Wagner, T.L.; Stafford, R.E.; Dickens, B.F.; Weglicki, W.B. Enhanced NO production during Mg deficiency and its role in mediating red blood cell glutathione loss. Am. J. Physiol. 1996, 271, C385–C390. [Google Scholar] [CrossRef] [PubMed]
- Hans, C.P.; Chaudhary, D.P.; Bansal, D.D. Effect of magnesium supplementation on oxidative stress in alloxanic diabetic rats. Magnes. Res. 2003, 16, 13–19. [Google Scholar] [PubMed]
- Song, Y.; Ridker, P.M.; Manson, J.E.; Cook, N.R.; Buring, J.E.; Liu, S. Magnesium intake, C-reactive protein, and the prevalence of metabolic syndrome in middle-aged and older U.S. women. Diabetes Care 2005, 28, 1438–1444. [Google Scholar] [CrossRef] [PubMed]
- Mazidi, M.; Rezaie, P.; Banach, M. Effect of magnesium supplements on serum C-reactive protein: A systematic review and meta-analysis. Arch. Med. Sci. 2018, 14, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Cernak, I.; Savic, V.; Kotur, J.; Prokic, V.; Kuljic, B.; Grbovic, D.; Veljovic, M. Alterations in magnesium and oxidative status during chronic emotional stress. Magnes. Res. 2000, 13, 29–36. [Google Scholar]
- Guerrero-Romero, F.; Rodriguez-Moran, M. Hypomagnesemia, oxidative stress, inflammation, and metabolic syndrome. Diabetes Metab. Res. Rev. 2006, 22, 471–476. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Morán, M.; Guerrero-Romero, F. Elevated concentrations of TNF-alpha are related to low serum magnesium levels in obese subjects. Magnes. Res. 2004, 17, 189–196. [Google Scholar]
- Maktabi, M.; Jamilian, M.; Asemi, Z. Magnesium-Zinc-Calcium-Vitamin D Co-supplementation Improves Hormonal Profiles, Biomarkers of Inflammation and Oxidative Stress in Women with Polycystic Ovary Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2018, 182, 21–28. [Google Scholar] [CrossRef]
- Afshar Ebrahimi, F.; Foroozanfard, F.; Aghadavod, E.; Bahmani, F.; Asemi, Z. The Effects of Magnesium and Zinc Co-Supplementation on Biomarkers of Inflammation and Oxidative Stress, and Gene Expression Related to Inflammation in Polycystic Ovary Syndrome: A Randomized Controlled Clinical Trial. Biol. Trace Elem. Res. 2018, 184, 300–307. [Google Scholar] [CrossRef]
- Mousavi, R.; Alizadeh, M.; Asghari Jafarabadi, M.; Heidari, L.; Nikbakht, R.; Babaahmadi Rezaei, H.; Karandish, M. Effects of Melatonin and/or Magnesium Supplementation on Biomarkers of Inflammation and Oxidative Stress in Women with Polycystic Ovary Syndrome: A Randomized, Double-Blind, Placebo-Controlled Trial. Biol. Trace Elem. Res. 2022, 200, 1010–1019. [Google Scholar] [CrossRef]
- Tannou, T.; Koeberle, S.; Manckoundia, P.; Aubry, R. Multifactorial immunodeficiency in frail elderly patients: Contributing factors and management. Med. Mal. Infect. 2019, 49, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Killilea, D.W.; Killilea, A.N. Mineral requirements for mitochondrial function: A connection to redox balance and cellular differentiation. Free Radic. Biol. Med. 2022, 182, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Goubern, M.; Rayssiguier, Y.; Miroux, B.; Chapey, M.F.; Ricquier, D.; Durlach, J. Effect of acute magnesium deficiency on the masking and unmasking of the proton channel of the uncoupling protein in rat brown fat. Magnes. Res. 1993, 6, 135–143. [Google Scholar] [PubMed]
- Turlapaty, P.D.; Altura, B.M. Extracellular magnesium ions control calcium exchange and content of vascular smooth muscle. Eur. J. Pharmacol. 1978, 52, 421–423. [Google Scholar] [CrossRef] [PubMed]
- Altura, B.M.; Altura, B.T. Magnesium and cardiovascular biology: An important link between cardiovascular risk factors and atherogenesis. Cell. Mol. Biol. Res. 1995, 41, 347–359. [Google Scholar] [PubMed]
- Malis, C.D.; Bonventre, J.V. Mechanism of calcium potentiation of oxygen free radical injury to renal mitochondria. A model for post-ischemic and toxic mitochondrial damage. J. Biol. Chem. 1986, 261, 14201–14208. [Google Scholar] [CrossRef]
- Kristian, T.; Siesjo, B.K. Calcium in ischemic cell death. Stroke 1998, 29, 705–718. [Google Scholar] [CrossRef]
- Boczkowski, J.; Lisdero, C.L.; Lanone, S.; Samb, A.; Carreras, M.C.; Boveris, A.; Aubier, M.; Poderoso, J.J. Endogenous peroxynitrite mediates mitochondrial dysfunction in rat diaphragm during endotoxemia. FASEB J. 1999, 13, 1637–1646. [Google Scholar] [CrossRef]
- Singh, I.N.; Sullivan, P.G.; Hall, E.D. Peroxynitrite-mediated oxidative damage to brain mitochondria: Protective effects of peroxynitrite scavengers. J. Neurosci. Res. 2007, 85, 2216–2223. [Google Scholar] [CrossRef]
- Yu, H.; Lin, L.; Zhang, Z.; Zhang, H.; Hu, H. Targeting NF-kappaB pathway for the therapy of diseases: Mechanism and clinical study. Signal Transduct. Target. Ther. 2020, 5, 209. [Google Scholar] [CrossRef]
- Shahi, A.; Aslani, S.; Ataollahi, M.; Mahmoudi, M. The role of magnesium in different inflammatory diseases. Inflammopharmacology 2019, 27, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Cleasby, M.E.; Jamieson, P.M.; Atherton, P.J. Insulin resistance and sarcopenia: Mechanistic links between common co-morbidities. J. Endocrinol. 2016, 229, R67–R81. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Frantz, J.D.; Tawa, N.E., Jr.; Melendez, P.A.; Oh, B.C.; Lidov, H.G.; Hasselgren, P.O.; Frontera, W.R.; Lee, J.; Glass, D.J.; et al. IKKbeta/NF-kappaB activation causes severe muscle wasting in mice. Cell 2004, 119, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Bar-Shai, M.; Carmeli, E.; Reznick, A.Z. The role of NF-kappaB in protein breakdown in immobilization, aging, and exercise: From basic processes to promotion of health. Ann. N. Y Acad. Sci. 2005, 1057, 431–447. [Google Scholar] [CrossRef] [PubMed]
- Polkinghorne, E.; Lau, Q.; Cooney, G.J.; Kraegen, E.W.; Cleasby, M.E. Local activation of the IkappaK-NF-kappaB pathway in muscle does not cause insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E316–E325. [Google Scholar] [CrossRef] [PubMed]
- Mourkioti, F.; Kratsios, P.; Luedde, T.; Song, Y.H.; Delafontaine, P.; Adami, R.; Parente, V.; Bottinelli, R.; Pasparakis, M.; Rosenthal, N. Targeted ablation of IKK2 improves skeletal muscle strength, maintains mass, and promotes regeneration. J. Clin. Investig. 2006, 116, 2945–2954. [Google Scholar] [CrossRef]
- Lorenzo, I.; Serra-Prat, M.; Yébenes, J.C. The Role of Water Homeostasis in Muscle Function and Frailty: A Review. Nutrients 2019, 11, 1857. [Google Scholar] [CrossRef]
- Squire, J. Special Issue: The Actin-Myosin Interaction in Muscle: Background and Overview. Int. J. Mol. Sci. 2019, 20, 5715. [Google Scholar] [CrossRef]
- Nielsen, F.H.; Lukaski, H.C. Update on the relationship between magnesium and exercise. Magnes. Res. 2006, 19, 180–189. [Google Scholar]
- Katzberg, H.D. Case Studies in Management of Muscle Cramps. Neurol. Clin. 2020, 38, 679–696. [Google Scholar] [CrossRef]
- Shrimanker, I.; Bhattarai, S. Electrolytes. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. [Google Scholar]
- Tardy, A.L.; Pouteau, E.; Marquez, D.; Yilmaz, C.; Scholey, A. Vitamins and Minerals for Energy, Fatigue and Cognition: A Narrative Review of the Biochemical and Clinical Evidence. Nutrients 2020, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Mendes Netto, R.S. Estado Nutricional em Magnésio Como Determinante da Eficiênciada Suplementação de Creatina em Praticantes de Musculação. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2004. [Google Scholar]
- Bohl, C.H.; Volpe, S.L. Magnesium and exercise. Crit. Rev. Food Sci. Nutr. 2002, 42, 533–563. [Google Scholar] [CrossRef] [PubMed]
- Costello, R.; Wallace, T.C.; Rosanoff, A. Magnesium. Adv. Nutr. 2016, 7, 199–201. [Google Scholar] [CrossRef] [PubMed]
- Lamothe, S.M.; Zhang, S. Chapter Five—Ubiquitination of Ion Channels and Transporters. Prog. Mol. Biol. Transl. Sci. 2016, 141, 161–223. [Google Scholar] [CrossRef]
- Yuen, G.K.; Galice, S.; Bers, D.M. Subcellular localization of Na/K-ATPase isoforms in ventricular myocytes. J. Mol. Cell Cardiol. 2017, 108, 158–169. [Google Scholar] [CrossRef]
- Feng, J.; Wang, H.; Jing, Z.; Wang, Y.; Cheng, Y.; Wang, W.; Sun, W. Role of Magnesium in Type 2 Diabetes Mellitus. Biol. Trace Elem. Res. 2020, 196, 74–85. [Google Scholar] [CrossRef]
- McDonough, A.A.; Fenton, R.A. Potassium homeostasis: Sensors, mediators, and targets. Pflug. Arch. 2022, 474, 853–867. [Google Scholar] [CrossRef]
- Kristensen, M.; Juel, C. Potassium-transporting proteins in skeletal muscle: Cellular location and fibre-type differences. Acta Physiol. 2010, 198, 105–123. [Google Scholar] [CrossRef]
- Aagaard, N.K.; Andersen, H.; Vilstrup, H.; Clausen, T.; Jakobsen, J.; Dørup, I. Magnesium supplementation and muscle function in patients with alcoholic liver disease: A randomized, placebo-controlled trial. Scand. J. Gastroenterol. 2005, 40, 972–979. [Google Scholar] [CrossRef]
- Finstad, E.W.; Newhouse, I.J.; Lukaski, H.C.; McAuliffe, J.E.; Stewart, C.R. The effects of magnesium supplementation on exercise performance. Med. Sci. Sports Exerc. 2001, 33, 493–498. [Google Scholar] [CrossRef]
- Golf, S.W.; Bender, S.; Grüttner, J. On the significance of magnesium in extreme physical stress. Cardiovasc. Drugs Ther. 1998, 12 (Suppl. 2), 197–202. [Google Scholar] [CrossRef]
- Gullestad, L.; Dolva, L.O.; Søyland, E.; Manger, A.T.; Falch, D.; Kjekshus, J. Oral magnesium supplementation improves metabolic variables and muscle strength in alcoholics. Alcohol. Clin. Exp. Res. 1992, 16, 986–990. [Google Scholar] [CrossRef] [PubMed]
- Kass, L.S.; Poeira, F. The effect of acute vs chronic magnesium supplementation on exercise and recovery on resistance exercise, blood pressure and total peripheral resistance on normotensive adults. J. Int. Soc. Sports Nutr. 2015, 12, 19. [Google Scholar] [CrossRef] [PubMed]
- Moslehi, N.; Vafa, M.; Sarrafzadeh, J.; Rahimi-Foroushani, A. Does magnesium supplementation improve body composition and muscle strength in middle-aged overweight women? A double-blind, placebo-controlled, randomized clinical trial. Biol. Trace Elem. Res. 2013, 153, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Selsby, J.T.; DiSilvestro, R.A.; Devor, S.T. Mg2+-creatine chelate and a low-dose creatine supplementation regimen improve exercise performance. J. Strength Cond. Res. 2004, 18, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Setaro, L.; Santos-Silva, P.R.; Nakano, E.Y.; Sales, C.H.; Nunes, N.; Greve, J.M.; Colli, C. Magnesium status and the physical performance of volleyball players: Effects of magnesium supplementation. J. Sports Sci. 2014, 32, 438–445. [Google Scholar] [CrossRef]
- Wary, C.; Brillault-Salvat, C.; Bloch, G.; Leroy-Willig, A.; Roumenov, D.; Grognet, J.M.; Leclerc, J.H.; Carlier, P.G. Effect of chronic magnesium supplementation on magnesium distribution in healthy volunteers evaluated by 31P-NMRS and ion selective electrodes. Br. J. Clin. Pharmacol. 1999, 48, 655–662. [Google Scholar] [CrossRef]
- Zajac, A.; Golas, A.; Chycki, J.; Halz, M.; Michalczyk, M.M. The Effects of Long-Term Magnesium Creatine Chelate Supplementation on Repeated Sprint Ability (RAST) in Elite Soccer Players. Nutrients 2020, 12, 2961. [Google Scholar] [CrossRef]
- Dahle, L.O.; Berg, G.; Hammar, M.; Hurtig, M.; Larsson, L. The effect of oral magnesium substitution on pregnancy-induced leg cramps. Am. J. Obstet. Gynecol. 1995, 173, 175–180. [Google Scholar] [CrossRef]
- Frusso, R.; Zárate, M.; Augustovski, F.; Rubinstein, A. Magnesium for the treatment of nocturnal leg cramps: A crossover randomized trial. J. Fam. Pract. 1999, 48, 868–871. [Google Scholar]
- Roffe, C.; Sills, S.; Crome, P.; Jones, P. Randomised, cross-over, placebo controlled trial of magnesium citrate in the treatment of chronic persistent leg cramps. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2002, 8, CR326–CR330. [Google Scholar]
- Nygaard, I.H.; Valbø, A.; Pethick, S.V.; Bøhmer, T. Does oral magnesium substitution relieve pregnancy-induced leg cramps? Eur. J. Obstet. Gynecol. Reprod. Biol. 2008, 141, 23–26. [Google Scholar] [CrossRef] [PubMed]
- Supakatisant, C.; Phupong, V. Oral magnesium for relief in pregnancy-induced leg cramps: A randomised controlled trial. Matern. Child Nutr. 2015, 11, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Yousef, A.A.; Al-deeb, A.E. A double-blinded randomised controlled study of the value of sequential intravenous and oral magnesium therapy in patients with chronic low back pain with a neuropathic component. Anaesthesia 2013, 68, 260–266. [Google Scholar] [CrossRef]
- Roguin Maor, N.; Alperin, M.; Shturman, E.; Khairaldeen, H.; Friedman, M.; Karkabi, K.; Milman, U. Effect of Magnesium Oxide Supplementation on Nocturnal Leg Cramps: A Randomized Clinical Trial. JAMA Intern. Med. 2017, 177, 617–623. [Google Scholar] [CrossRef] [PubMed]
- Kass, L.S.; Skinner, P.; Poeira, F. A pilot study on the effects of magnesium supplementation with high and low habitual dietary magnesium intake on resting and recovery from aerobic and resistance exercise and systolic blood pressure. J. Sports Sci. Med. 2013, 12, 144–150. [Google Scholar]
- Veronese, N.; Berton, L.; Carraro, S.; Bolzetta, F.; De Rui, M.; Perissinotto, E.; Toffanello, E.D.; Bano, G.; Pizzato, S.; Miotto, F.; et al. Effect of oral magnesium supplementation on physical performance in healthy elderly women involved in a weekly exercise program: A randomized controlled trial. Am. J. Clin. Nutr. 2014, 100, 974–981. [Google Scholar] [CrossRef]
- Zorbas, Y.G.; Kakuris, K.K.; Federenko, Y.F.; Deogenov, V.A. Utilization of magnesium during hypokinesia and magnesium supplementation in healthy subjects. Nutrition 2010, 26, 1134–1138. [Google Scholar] [CrossRef]
- Bayram, S.; Şahin, K.; Anarat, F.B.; Chousein, C.M.; Kocazeybek, E.; Altan, M.; Akgül, T. The effect of oral magnesium supplementation on acute non-specific low back pain: Prospective randomized clinical trial. Am. J. Emerg. Med. 2021, 47, 125–130. [Google Scholar] [CrossRef]
- Córdova Martínez, A.; Fernández-Lázaro, D.; Mielgo-Ayuso, J.; Seco Calvo, J.; Caballero García, A. Effect of magnesium supplementation on muscular damage markers in basketball players during a full season. Magnes. Res. 2017, 30, 61–70. [Google Scholar] [CrossRef]
- Terblanche, S.; Noakes, T.D.; Dennis, S.C.; Marais, D.; Eckert, M. Failure of magnesium supplementation to influence marathon running performance or recovery in magnesium-replete subjects. Int. J. Sport Nutr. 1992, 2, 154–164. [Google Scholar] [CrossRef] [PubMed]


| Location | Magnesium Distribution |
|---|---|
| Bone | 50–60% of total magnesium (24–29 g), with approximately one-third being exchangeable |
| Soft Tissues (muscles and other organs) | 34–39% of total magnesium |
| Blood | Less than 1% of the body’s magnesium |
| Plasma | 60% ionized, 30% bound to albumin, 10% complexed with serum anions (phosphate and citrate) |
| Life Stage | PRI (mg) | AR (mg) | UL * (mg) | RDA-DRI (mg) | DRV-AI (mg) |
|---|---|---|---|---|---|
| Birth to 6 months | - | - | Nd | 30 | - |
| Infants 7–12 months | 80 | Nd | Nd | 75 | 80 |
| Children 1–3 years | 80 | 65 | 250 | 80 | 170 |
| Children 4–6 years | 100 | 85 | 250 | 130 | 230 |
| Children 7–10 years | 150 | 130 | 250 | 240 | 230 |
| Teen boys 11–18 years | 240 | 170–200 | 250 | 410 | 300 |
| Teen girls 11–18 years | 240 | 170–200 | 250 | 360 | 250 |
| Men | 240 | 170 | 250 | 400–420 | 350 |
| Women | 240 | 170 | 250 | 310–320 | 300 |
| Pregnant | 240 | 170 | 250 | 350–400 | 300 |
| Breastfeeding | 240 | 170 | 250 | 310–360 | 300 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, A.C.R.; Vasconcelos, A.R.; Dias, D.D.; Komoni, G.; Name, J.J. The Integral Role of Magnesium in Muscle Integrity and Aging: A Comprehensive Review. Nutrients 2023, 15, 5127. https://doi.org/10.3390/nu15245127
Souza ACR, Vasconcelos AR, Dias DD, Komoni G, Name JJ. The Integral Role of Magnesium in Muscle Integrity and Aging: A Comprehensive Review. Nutrients. 2023; 15(24):5127. https://doi.org/10.3390/nu15245127
Chicago/Turabian StyleSouza, Ana Carolina Remondi, Andrea Rodrigues Vasconcelos, Denise Deo Dias, Geovana Komoni, and José João Name. 2023. "The Integral Role of Magnesium in Muscle Integrity and Aging: A Comprehensive Review" Nutrients 15, no. 24: 5127. https://doi.org/10.3390/nu15245127
APA StyleSouza, A. C. R., Vasconcelos, A. R., Dias, D. D., Komoni, G., & Name, J. J. (2023). The Integral Role of Magnesium in Muscle Integrity and Aging: A Comprehensive Review. Nutrients, 15(24), 5127. https://doi.org/10.3390/nu15245127