
Thyme Extract Alleviates High-Fat Diet-Induced Obesity and Gut Dysfunction


Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Water-Soluble TLE
2.2. Animals
2.3. InAlyzer Analysis
2.4. Biochemical Analysis
2.5. Oral Glucose Tolerance Test
2.6. Colon Length and pH Measurement
2.7. Assessment of Intestinal Permeability
2.8. Western Blotting
2.9. Metabolite Profiling
2.10. Statistical Analysis
3. Results
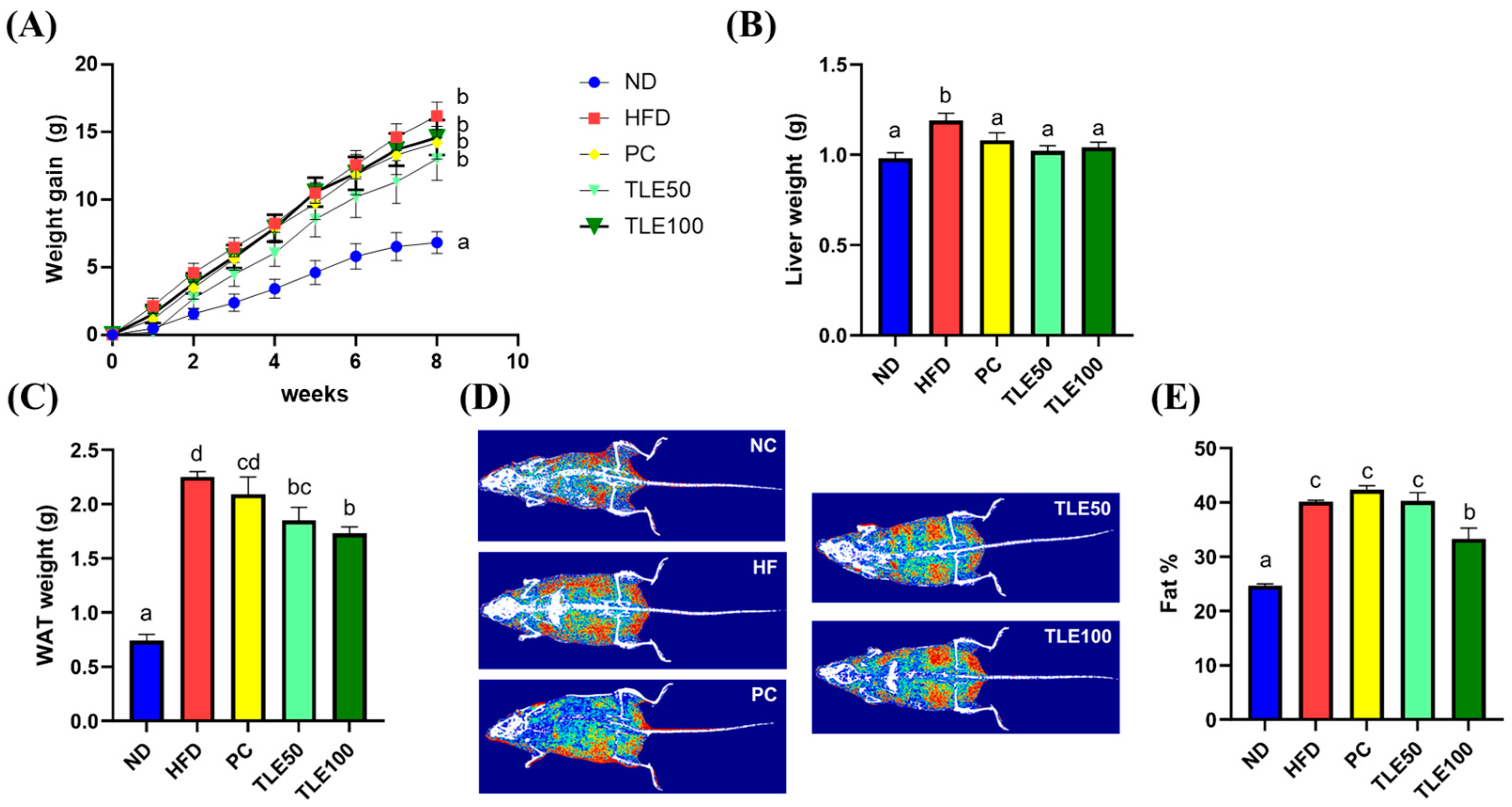
3.1. Changes in Body and Liver Weight
3.2. Effect of TLE on Serum Biochemical Parameters
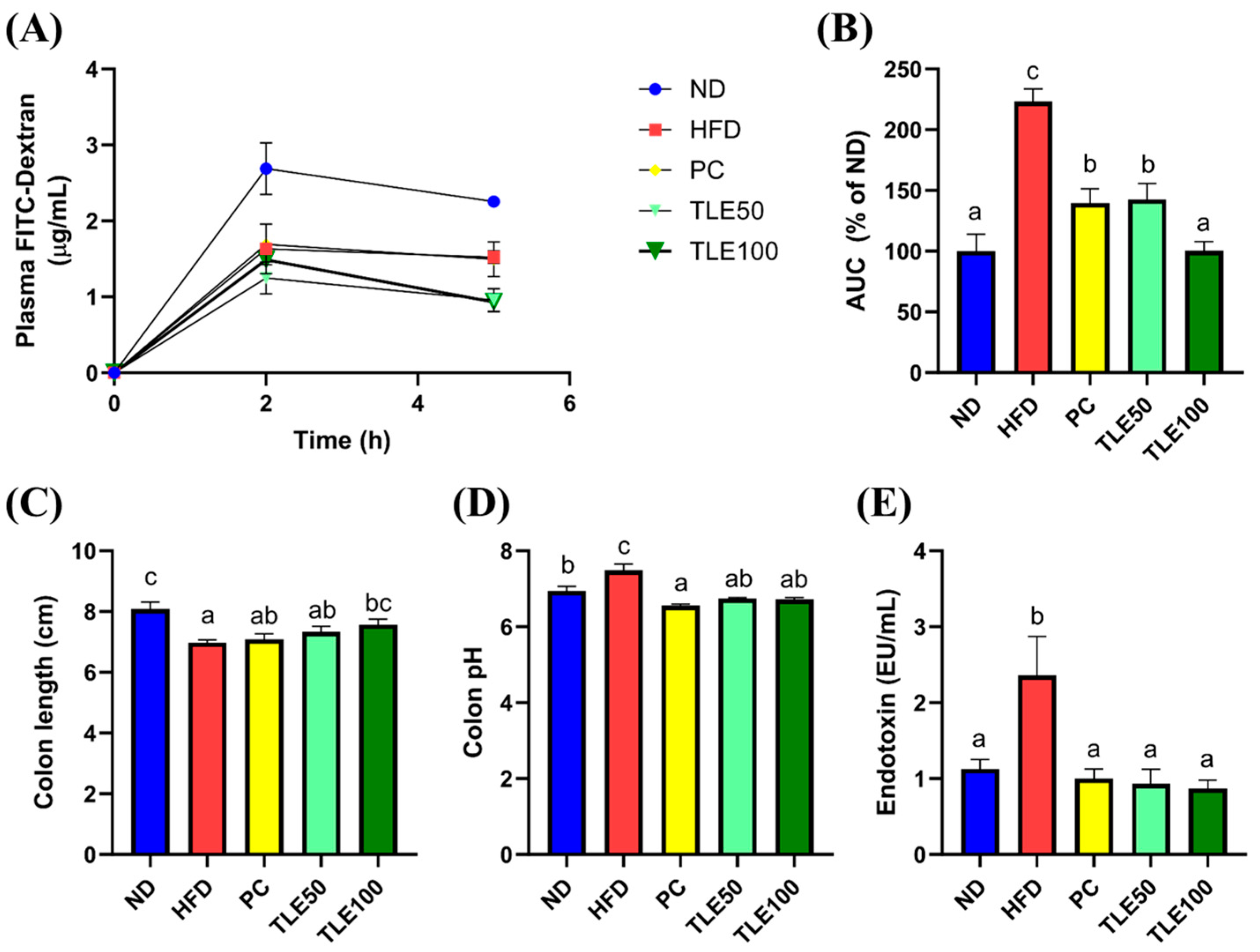
3.3. Effects of TLE on Gut Permeability
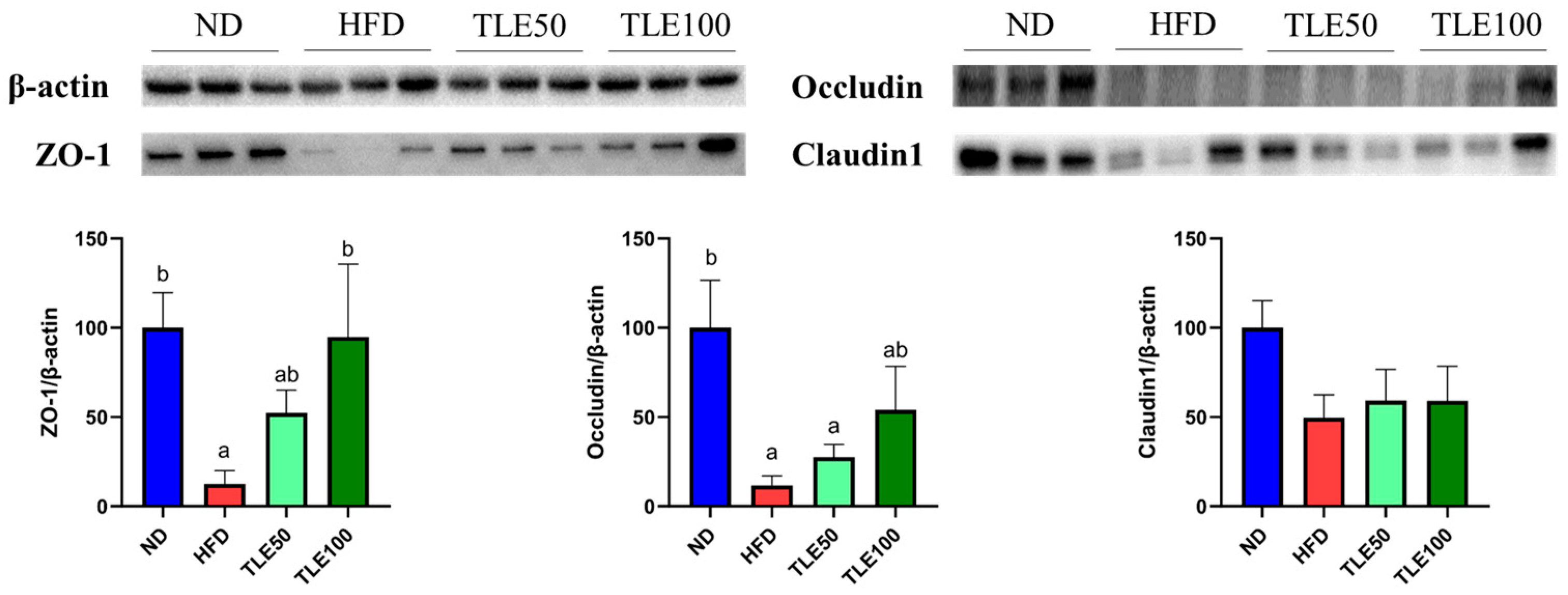
3.4. Effect of TLE on the Expression of Tight Junction Proteins
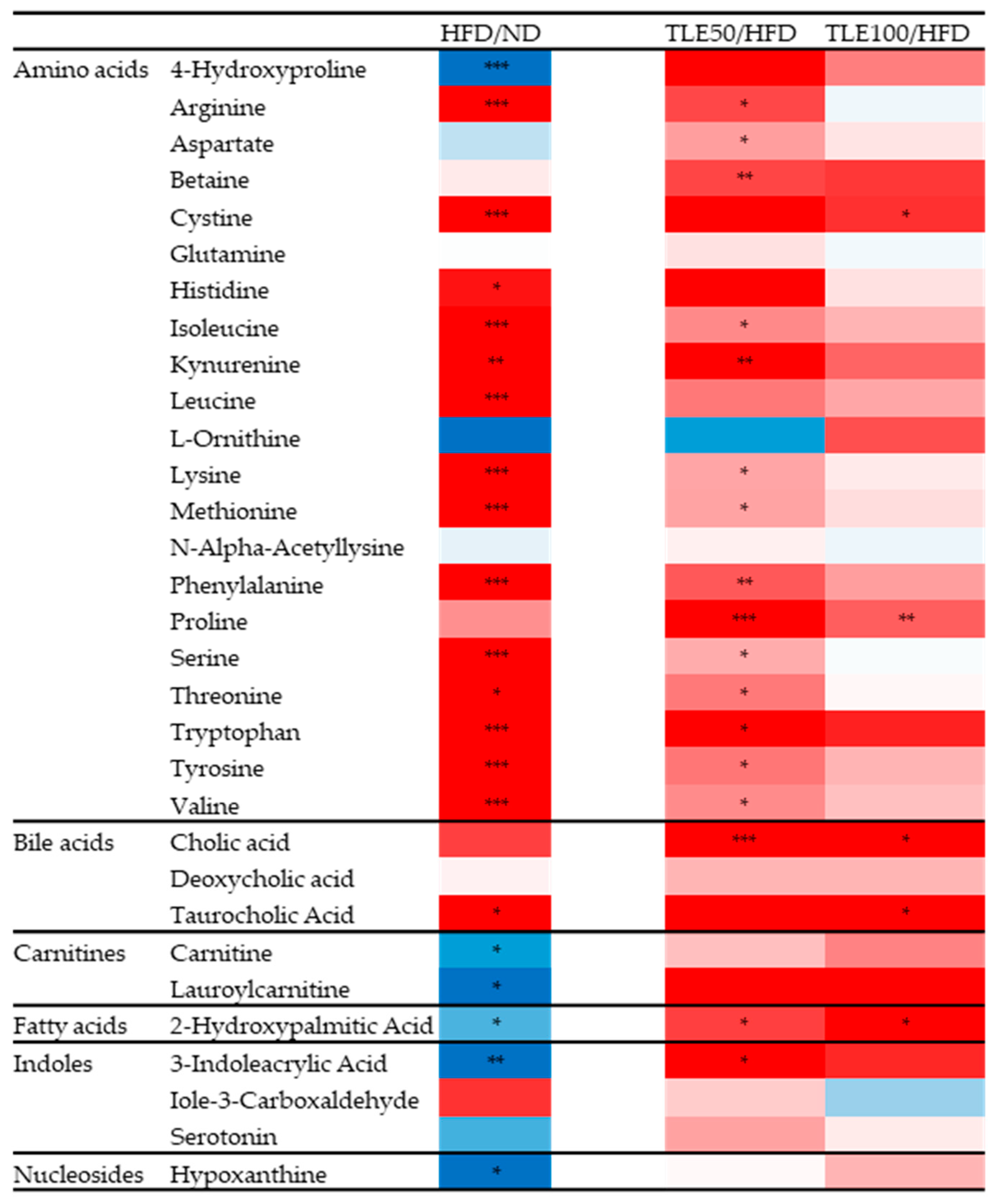
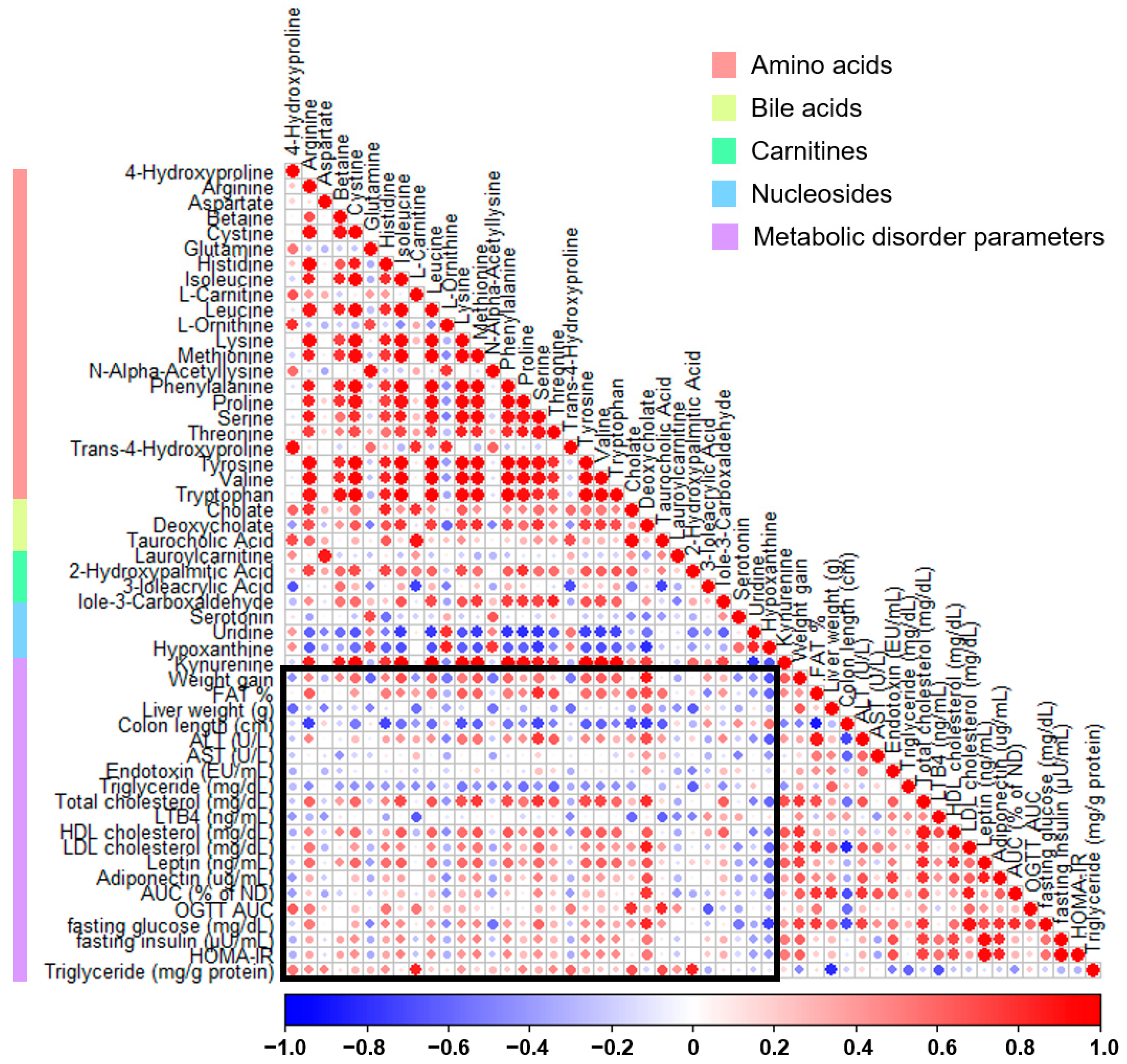
3.5. Correlation between Metabolites and Serum Metabolic Disorder Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ismaili, H.; Milella, L.; Fkih-Tetouani, S.; Ilidrissi, A.; Camporese, A.; Sosa, S.; Altinier, G.; Della Loggia, R.; Aquino, R. In vivo topical anti-inflammatory and in vitro antioxidant activities of two extracts of Thymus satureioides leaves. J. Ethnopharmacol. 2004, 91, 31–36. [Google Scholar] [CrossRef]
- Salehi, B.; Mishra, A.P.; Shukla, I.; Sharifi-Rad, M.; Contreras, M.D.M.; Segura-Carretero, A.; Fathi, H.; Nasrabadi, N.N.; Kobarfard, F.; Sharifi-Rad, J. Thymol, thyme, and other plant sources: Health and potential uses. Phytother. Res. 2018, 32, 1688–1706. [Google Scholar] [CrossRef] [PubMed]
- Almanea, A.; Abd El-Aziz, G.S.; Ahmed, M.M.M. The potential gastrointestinal health benefits of Thymus vulgaris essential oil: A review. Biomed. Pharmacol. J. 2019, 12, 1793–1799. [Google Scholar] [CrossRef]
- Abbaszadeh, S.; Teimouri, H.; Farzan, B. An ethnobotanical study of medicinal plants with antianxiety and antidepressant effects in Shahrekord. Egypt. J. Vet. Sci. 2019, 50, 81–87. [Google Scholar] [CrossRef]
- Nadi, A.; Shiravi, A.A.; Mohammadi, Z.; Aslani, A.; Zeinalian, M. Thymus vulgaris, a natural pharmacy against COVID-19: A molecular review. J. Herb. Med. 2023, 38, 100635. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V. Thymus vulgaris. In Medicinal Spices and Vegetables from Africa; Academic Press: Cambridge, MA, USA, 2017; pp. 599–609. [Google Scholar] [CrossRef]
- Heidari, Z.; Salehzadeh, A.; Sadat Shandiz, S.A.; Tajdoost, S. Anti-cancer and anti-oxidant properties of ethanolic leaf extract of Thymus vulgaris and its bio-functionalized silver nanoparticles. 3 Biotech 2018, 8, 177. [Google Scholar] [CrossRef]
- Ocana, A.; Reglero, G. Effects of Thyme Extract Oils (from Thymus vulgaris, Thymus zygis, and Thymus hyemalis) on Cytokine Production and Gene Expression of oxLDL-Stimulated THP-1-Macrophages. J. Obes. 2012, 2012, 104706. [Google Scholar] [CrossRef]
- Vigo, E.; Cepeda, A.; Gualillo, O.; Perez-Fernandez, R. In-vitro anti-inflammatory effect of Eucalyptus globulus and Thymus vulgaris: Nitric oxide inhibition in J774A.1 murine macrophages. J. Pharm. Pharmacol. 2004, 56, 257–263. [Google Scholar] [CrossRef]
- Uritu, C.M.; Mihai, C.T.; Stanciu, G.D.; Dodi, G.; Alexa-Stratulat, T.; Luca, A.; Leon-Constantin, M.M.; Stefanescu, R.; Bild, V.; Melnic, S.; et al. Medicinal Plants of the Family Lamiaceae in Pain Therapy: A Review. Pain. Res. Manag. 2018, 2018, 7801543. [Google Scholar] [CrossRef]
- Hegazy, A.M.; Abdel-Azeem, A.S.; Zeidan, H.M.; Ibrahim, K.S.; Sayed, E.E. Hypolipidemic and hepatoprotective activities of rosemary and thyme in gentamicin-treated rats. Hum. Exp. Toxicol. 2018, 37, 420–430. [Google Scholar] [CrossRef]
- El-Boshy, M.E.; Refaat, B.; Qasem, A.H.; Khan, A.; Ghaith, M.; Almasmoum, H.; Mahbub, A.; Almaimani, R.A. The remedial effect of Thymus vulgaris extract against lead toxicity-induced oxidative stress, hepatorenal damage, immunosuppression, and hematological disorders in rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 22736–22746. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Marchese, A.; Izadi, M.; Curti, V.; Daglia, M.; Nabavi, S.F. Plants belonging to the genus Thymus as antibacterial agents: From farm to pharmacy. Food Chem. 2015, 173, 339–347. [Google Scholar] [CrossRef]
- Sfeir, J.; Lefrancois, C.; Baudoux, D.; Derbre, S.; Licznar, P. In Vitro Antibacterial Activity of Essential Oils against Streptococcus pyogenes. Evid. Based Complement. Altern. Med. 2013, 2013, 269161. [Google Scholar] [CrossRef] [PubMed]
- Rezatofighi, S.E.; Seydabadi, A.; Seyyed Nejad, S.M. Evaluating the Efficacy of Achillea millefolium and Thymus vulgaris Extracts Against Newcastle Disease Virus in Ovo. Jundishapur J. Microbiol. 2014, 7, e9016. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.G.; Shibamoto, T. Determination of antioxidant potential of volatile extracts isolated from various herbs and spices. J. Agric. Food Chem. 2002, 50, 4947–4952. [Google Scholar] [CrossRef] [PubMed]
- Miladi, H.; Slama, R.B.; Mili, D.; Zouari, S.; Bakhrouf, A.; Ammar, E. Essential oil of Thymus vulgaris L. and Rosmarinus officinalis L.: Gas chromatography-mass spectrometry analysis, cytotoxicity and antioxidant properties and antibacterial activities against foodborne pathogens. Nat. Sci. 2013, 5, 729–739. [Google Scholar] [CrossRef]
- Babaei, M.; Abarghoei, M.E.; Ansari, R.; Vafaei, A.A.; Taherian, A.A.; Akhavan, M.M.; Toussy, G.; Mousavi, S. Antispasmodic effect of hydroalcoholic extract of Thymus vulgaris on the guinea-pig ileum. Nat. Prod. Res. 2008, 22, 1143–1150. [Google Scholar] [CrossRef]
- Begrow, F.; Engelbertz, J.; Feistel, B.; Lehnfeld, R.; Bauer, K.; Verspohl, E.J. Impact of thymol in thyme extracts on their antispasmodic action and ciliary clearance. Planta Med. 2010, 76, 311–318. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; He, W.; Tian, H.; Zhan, P.; Liu, J. Thyme (Thymus vulgaris L.) polyphenols ameliorate DSS-induced ulcerative colitis of mice by mitigating intestinal barrier damage, regulating gut microbiota, and suppressing TLR4/NF-kappaB-NLRP3 inflammasome pathways. Food Funct. 2023, 14, 1113–1132. [Google Scholar] [CrossRef]
- Mehdipour, Z.; Afsharmanesh, M.; Sami, M. Effects of supplemental thyme extract (Thymus vulgaris L.) on growth performance, intestinal microbial populations, and meat quality in Japanese quails. Comp. Clin. Pathol. 2014, 23, 1503–1508. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.H.L.; Kim, Y.; Lee, H.-B.; Lee, E.; Park, J.H.; Park, H.-Y. Corchorus olitorius L. ameliorates alcoholic liver disease by regulating gut-liver axis. J. Funct. Foods 2021, 85, 104648. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.H.L.; Lee, J.E.; Park, M.; Oh, M.J.; Lee, H.B.; Park, J.H.; Jhun, H.; Kim, J.H.; Kang, C.H.; et al. Gellan gum prevents non-alcoholic fatty liver disease by modulating the gut microbiota and metabolites. Food Chem. 2023, 400, 134038. [Google Scholar] [CrossRef]
- Wen, X.-D.; Wang, C.-Z.; Yu, C.; Zhang, Z.; Calway, T.; Wang, Y.; Li, P.; Yuan, C.-S. Salvia miltiorrhiza (dan shen) significantly ameliorates colon inflammation in dextran sulfate sodium induced colitis. Am. J. Chin. Med. 2013, 41, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- Manting, L.; Haihong, Z.; Jing, L.; Shaodong, C.; Yihua, L. The model of rat lipid metabolism disorder induced by chronic stress accompanying high-fat-diet. Lipids Health Dis. 2011, 10, 153. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.; Gholami, A.M.; Berry, D.; Desmarchelier, C.; Hahne, H.; Loh, G.; Mondot, S.; Lepage, P.; Rothballer, M.; Walker, A.; et al. High-fat diet alters gut microbiota physiology in mice. ISME J. 2014, 8, 295–308. [Google Scholar] [CrossRef]
- Frank, S.M.; Webster, J.; McKenzie, B.; Geldsetzer, P.; Manne-Goehler, J.; Andall-Brereton, G.; Houehanou, C.; Houinato, D.; Gurung, M.S.; Bicaba, B.W.; et al. Consumption of Fruits and Vegetables Among Individuals 15 Years and Older in 28 Low- and Middle-Income Countries. J. Nutr. 2019, 149, 1252–1259. [Google Scholar] [CrossRef]
- Gao, M.; Ma, Y.; Liu, D. High-fat diet-induced adiposity, adipose inflammation, hepatic steatosis and hyperinsulinemia in outbred CD-1 mice. PLoS ONE 2015, 10, e0119784. [Google Scholar] [CrossRef]
- Fachini-Queiroz, F.C.; Kummer, R.; Estevao-Silva, C.F.; Carvalho, M.D.; Cunha, J.M.; Grespan, R.; Bersani-Amado, C.A.; Cuman, R.K. Effects of Thymol and Carvacrol, Constituents of Thymus vulgaris L. Essential Oil, on the Inflammatory Response. Evid. Based Complement. Altern. Med. 2012, 2012, 657026. [Google Scholar] [CrossRef]
- Oliva Mde, L.; Carezzano, M.E.; Giuliano, M.; Daghero, J.; Zygadlo, J.; Bogino, P.; Giordano, W.; Demo, M. Antimicrobial activity of essential oils of Thymus vulgaris and Origanum vulgare on phytopathogenic strains isolated from soybean. Plant Biol. 2015, 17, 758–765. [Google Scholar] [CrossRef]
- Sanchez, G.; Aznar, R. Evaluation of Natural Compounds of Plant Origin for Inactivation of Enteric Viruses. Food Environ. Virol. 2015, 7, 183–187. [Google Scholar] [CrossRef]
- Kohiyama, C.Y.; Yamamoto Ribeiro, M.M.; Mossini, S.A.; Bando, E.; Bomfim Nda, S.; Nerilo, S.B.; Rocha, G.H.; Grespan, R.; Mikcha, J.M.; Machinski, M., Jr. Antifungal properties and inhibitory effects upon aflatoxin production of Thymus vulgaris L. by Aspergillus flavus Link. Food Chem. 2015, 173, 1006–1010. [Google Scholar] [CrossRef]
- Al-Menhali, A.; Al-Rumaihi, A.; Al-Mohammed, H.; Al-Mazrooey, H.; Al-Shamlan, M.; AlJassim, M.; Al-Korbi, N.; Eid, A.H. Thymus vulgaris (thyme) inhibits proliferation, adhesion, migration, and invasion of human colorectal cancer cells. J. Med. Food 2015, 18, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Mihailovic-Stanojevic, N.; Miloradovic, Z.; Ivanov, M.; Bugarski, B.; Jovovic, D.; Karanovic, D.; Vajic, U.J.; Komes, D.; Grujic-Milanovic, J. Upregulation of Heme Oxygenase-1 in Response to Wild Thyme Treatment Protects against Hypertension and Oxidative Stress. Oxid. Med. Cell Longev. 2016, 2016, 1458793. [Google Scholar] [CrossRef] [PubMed]
- Aazza, S.; Lyoussi, B.; Megias, C.; Cortes-Giraldo, I.; Vioque, J.; Figueiredo, A.C.; Miguel, M.G. Anti-oxidant, anti-inflammatory and anti-proliferative activities of Moroccan commercial essential oils. Nat. Prod. Commun. 2014, 9, 587–594. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili-Mahani, S.; Falahi, F.; Yaghoobi, M.M. Proapoptotic and Antiproliferative Effects of Thymus caramanicus on Human Breast Cancer Cell Line (MCF-7) and Its Interaction with Anticancer Drug Vincristine. Evid. Based Complement. Altern. Med. 2014, 2014, 893247. [Google Scholar] [CrossRef] [PubMed]
- Giarratana, F.; Muscolino, D.; Beninati, C.; Giuffrida, A.; Panebianco, A. Activity of Thymus vulgaris essential oil against Anisakis larvae. Exp. Parasitol. 2014, 142, 7–10. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.S.; Landau, J.M.; Huang, M.T.; Newmark, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef]
- Soliman, M.M.; Aldhahrani, A.; Metwally, M.M.M. Hepatoprotective effect of Thymus vulgaris extract on sodium nitrite-induced changes in oxidative stress, antioxidant and inflammatory marker expression. Sci. Rep. 2021, 11, 5747. [Google Scholar] [CrossRef]
- Salzman, N.H.; Underwood, M.A.; Bevins, C.L. Paneth cells, defensins, and the commensal microbiota: A hypothesis on intimate interplay at the intestinal mucosa. Semin. Immunol. 2007, 19, 70–83. [Google Scholar] [CrossRef]
- Jin, B.R.; Chung, K.S.; Cheon, S.Y.; Lee, M.; Hwang, S.; Noh Hwang, S.; Rhee, K.J.; An, H.J. Rosmarinic acid suppresses colonic inflammation in dextran sulphate sodium (DSS)-induced mice via dual inhibition of NF-kappaB and STAT3 activation. Sci. Rep. 2017, 7, 46252. [Google Scholar] [CrossRef]
- Melo, M.L.; Brito, G.A.; Soares, R.C.; Carvalho, S.B.; Silva, J.V.; Soares, P.M.; Vale, M.L.; Souza, M.H.; Cunha, F.Q.; Ribeiro, R.A. Role of cytokines (TNF-alpha, IL-1beta and KC) in the pathogenesis of CPT-11-induced intestinal mucositis in mice: Effect of pentoxifylline and thalidomide. Cancer Chemother. Pharmacol. 2008, 61, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Kirpich, I.A.; Marsano, L.S.; McClain, C.J. Gut-liver axis, nutrition, and non-alcoholic fatty liver disease. Clin. Biochem. 2015, 48, 923–930. [Google Scholar] [CrossRef] [PubMed]
- Backhed, F.; Manchester, J.K.; Semenkovich, C.F.; Gordon, J.I. Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. USA 2007, 104, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, J.C.; Matheus, V.A.; Oliveira, R.B.; Tada, S.F.S.; Collares-Buzato, C.B. High-Fat Diet Induces Disruption of the Tight Junction-Mediated Paracellular Barrier in the Proximal Small Intestine Before the Onset of Type 2 Diabetes and Endotoxemia. Dig. Dis. Sci. 2021, 66, 3359–3374. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.B.; Oh, M.J.; Jhun, H.; Choi, S.Y.; Park, H.Y. Polysaccharide fraction from greens of Raphanus sativus alleviates high fat diet-induced obesity. Food Chem. 2021, 343, 128395. [Google Scholar] [CrossRef] [PubMed]
- Heintz-Buschart, A.; Wilmes, P. Human Gut Microbiome: Function Matters. Trends Microbiol. 2018, 26, 563–574. [Google Scholar] [CrossRef]
- Heintz-Buschart, A.; May, P.; Laczny, C.C.; Lebrun, L.A.; Bellora, C.; Krishna, A.; Wampach, L.; Schneider, J.G.; Hogan, A.; de Beaufort, C.; et al. Integrated multi-omics of the human gut microbiome in a case study of familial type 1 diabetes. Nat. Microbiol. 2016, 2, 16180. [Google Scholar] [CrossRef]
- Zierer, J.; Jackson, M.A.; Kastenmuller, G.; Mangino, M.; Long, T.; Telenti, A.; Mohney, R.P.; Small, K.S.; Bell, J.T.; Steves, C.J.; et al. The fecal metabolome as a functional readout of the gut microbiome. Nat. Genet. 2018, 50, 790–795. [Google Scholar] [CrossRef]
- Riecan, M.; Paluchova, V.; Lopes, M.; Brejchova, K.; Kuda, O. Branched and linear fatty acid esters of hydroxy fatty acids (FAHFA) relevant to human health. Pharmacol. Ther. 2022, 231, 107972. [Google Scholar] [CrossRef]
- Zhang, B.; Jiang, M.; Zhao, J.; Song, Y.; Du, W.; Shi, J. The Mechanism Underlying the Influence of Indole-3-Propionic Acid: A Relevance to Metabolic Disorders. Front. Endocrinol. 2022, 13, 841703. [Google Scholar] [CrossRef]
- Kuipers, F.; Bloks, V.W.; Groen, A.K. Beyond intestinal soap--bile acids in metabolic control. Nat. Rev. Endocrinol. 2014, 10, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Huang, D.; Xu, X.; Wu, X.; Liu, L.; Niu, W.; Lu, L.; Zhou, H. An elevated deoxycholic acid level induced by high-fat feeding damages intestinal stem cells by reducing the ileal IL-22. Biochem. Biophys. Res. Commun. 2021, 579, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Patti, M.E.; Houten, S.M.; Bianco, A.C.; Bernier, R.; Larsen, P.R.; Holst, J.J.; Badman, M.K.; Maratos-Flier, E.; Mun, E.C.; Pihlajamaki, J.; et al. Serum bile acids are higher in humans with prior gastric bypass: Potential contribution to improved glucose and lipid metabolism. Obesity 2009, 17, 1671–1677. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Ibelles, P.; Gutierrez-Vidal, R.; Calixto-Tlacomulco, S.; Delgado-Coello, B.; Mas-Oliva, J. Hepatic Accumulation of Hypoxanthine: A Link between Hyperuricemia and Nonalcoholic Fatty Liver Disease. Arch. Med. Res. 2021, 52, 692–702. [Google Scholar] [CrossRef]
- Furuhashi, M.; Koyama, M.; Higashiura, Y.; Murase, T.; Nakamura, T.; Matsumoto, M.; Sakai, A.; Ohnishi, H.; Tanaka, M.; Saitoh, S.; et al. Differential regulation of hypoxanthine and xanthine by obesity in a general population. J. Diabetes Investig. 2020, 11, 878–887. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Compound | Precursor | Product | CE | RT (min) |
|---|---|---|---|---|---|
| Amino acids | 4-Hydroxyproline | 132.1 | 86.2 | 14 | 0.9 |
| Arginine | 175.1 | 70.2 | 20 | 0.87 | |
| Aspartate | 134.1 | 74.1 | 14 | 0.93 | |
| Betaine | 118.1 | 59.1 | 16 | 0.95 | |
| Cystine | 241.1 | 152 | 12 | 0.88 | |
| Glutamine | 148.1 | 84.2 | 14 | 0.92 | |
| Histidine | 156.1 | 110.2 | 14 | 0.86 | |
| Isoleucine | 132.1 | 86.2 | 10 | 1.43 | |
| Leucine | 132.1 | 86.2 | 12 | 1.55 | |
| L-Ornithine | 133.1 | 70.2 | 14 | 0.87 | |
| Lysine | 147.1 | 84.2 | 14 | 0.84 | |
| Methionine | 150.1 | 104.1 | 12 | 1.23 | |
| N-Alpha-Acetyllysine | 189.3 | 84.1 | 20 | 0.84 | |
| Phenylalanine | 166.1 | 120.1 | 14 | 2.17 | |
| Proline | 117 | 71.1 | 14 | 0.94 | |
| Serine | 106 | 60.2 | 10 | 0.9 | |
| Threonine | 120 | 74.1 | 8 | 0.92 | |
| Tryptophan | 205.1 | 146.1 | 18 | 3.16 | |
| Tyrosine | 182.1 | 136.1 | 12 | 1.29 | |
| Valine | 118.1 | 72.2 | 10 | 1.25 | |
| Kynurenine | 209.1 | 146.1 | 18 | 2.23 | |
| Bile acids | Cholic acid | 407.3 | 343.3 | 30 | 12.71 |
| Deoxycholic acid | 391.3 | 345.2 | 28 | 14.52 | |
| Taurocholic Acid | 514.3 | 80.1 | 58 | 9.89 | |
| Carnitines | L-Carnitine | 162.1 | 60.2 | 16 | 0.94 |
| Lauroylcarnitine | 344.4 | 85 | 24 | 13.14 | |
| Fatty acids | 2-Hydroxypalmitic Acid | 271.3 | 225.2 | 20 | 17.83 |
| Indoles | 3-Indoleacrylic Acid | 188.1 | 115.1 | 28 | 7.94 |
| Indole-3-Carboxaldehyde | 146.1 | 118.1 | 14 | 6.33 | |
| Serotonin | 177.1 | 115.1 | 24 | 1.74 | |
| Purines | Hypoxanthine | 137 | 110.1 | 20 | 1.29 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.R.; Lee, H.-B.; Oh, M.-J.; Kim, Y.; Park, H.-Y. Thyme Extract Alleviates High-Fat Diet-Induced Obesity and Gut Dysfunction. Nutrients 2023, 15, 5007. https://doi.org/10.3390/nu15235007
Lee YR, Lee H-B, Oh M-J, Kim Y, Park H-Y. Thyme Extract Alleviates High-Fat Diet-Induced Obesity and Gut Dysfunction. Nutrients. 2023; 15(23):5007. https://doi.org/10.3390/nu15235007
Chicago/Turabian StyleLee, Yu Ra, Hye-Bin Lee, Mi-Jin Oh, Yoonsook Kim, and Ho-Young Park. 2023. "Thyme Extract Alleviates High-Fat Diet-Induced Obesity and Gut Dysfunction" Nutrients 15, no. 23: 5007. https://doi.org/10.3390/nu15235007
APA StyleLee, Y. R., Lee, H.-B., Oh, M.-J., Kim, Y., & Park, H.-Y. (2023). Thyme Extract Alleviates High-Fat Diet-Induced Obesity and Gut Dysfunction. Nutrients, 15(23), 5007. https://doi.org/10.3390/nu15235007