
Effects of Methionine Restriction from Different Sources on Sperm Quality in Aging Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Procedures, Diets and Treatments
2.2. Blood and Tissue Preparation
2.3. Measurement of Body Weight and Feed Intake
2.4. Testicular Histopathology Analysis
2.5. Determination of Serum Testosterone Content
2.6. Determination of Sperm Quality
2.7. Determination of Methionine Metabolites in Testis
2.8. Testicular RNA Extraction, Reverse Transcription and Real-Time Fluorescence Quantitative PCR
2.9. Determination of DNA and RNA Methylation Levels in Sperm
2.10. Statistical Analysis
3. Results
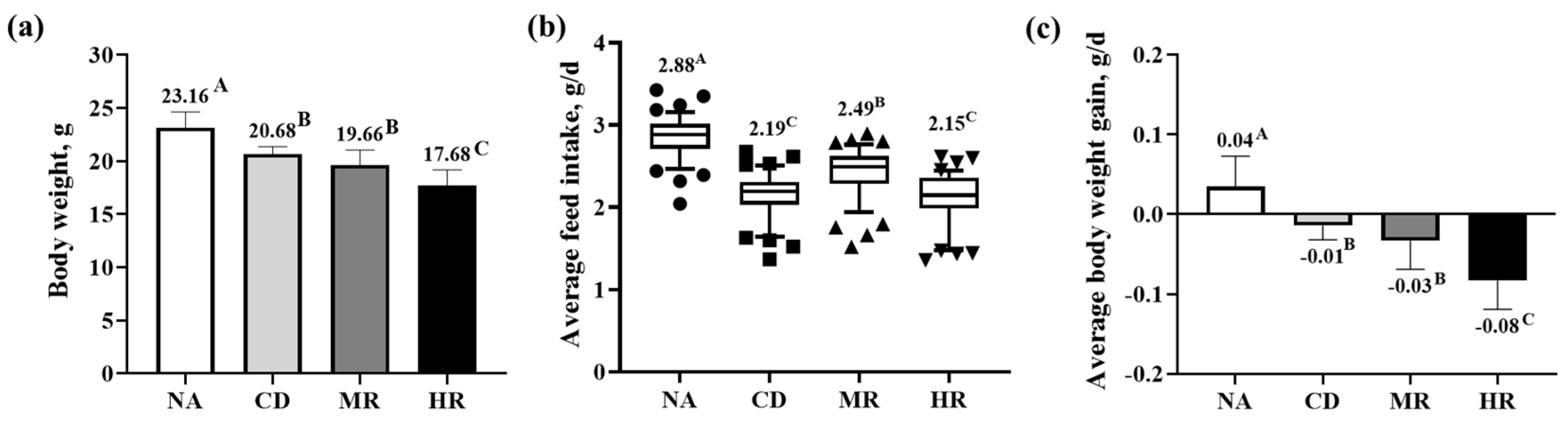
3.1. Growth Performance of Aging Mice with Methionine Restriction
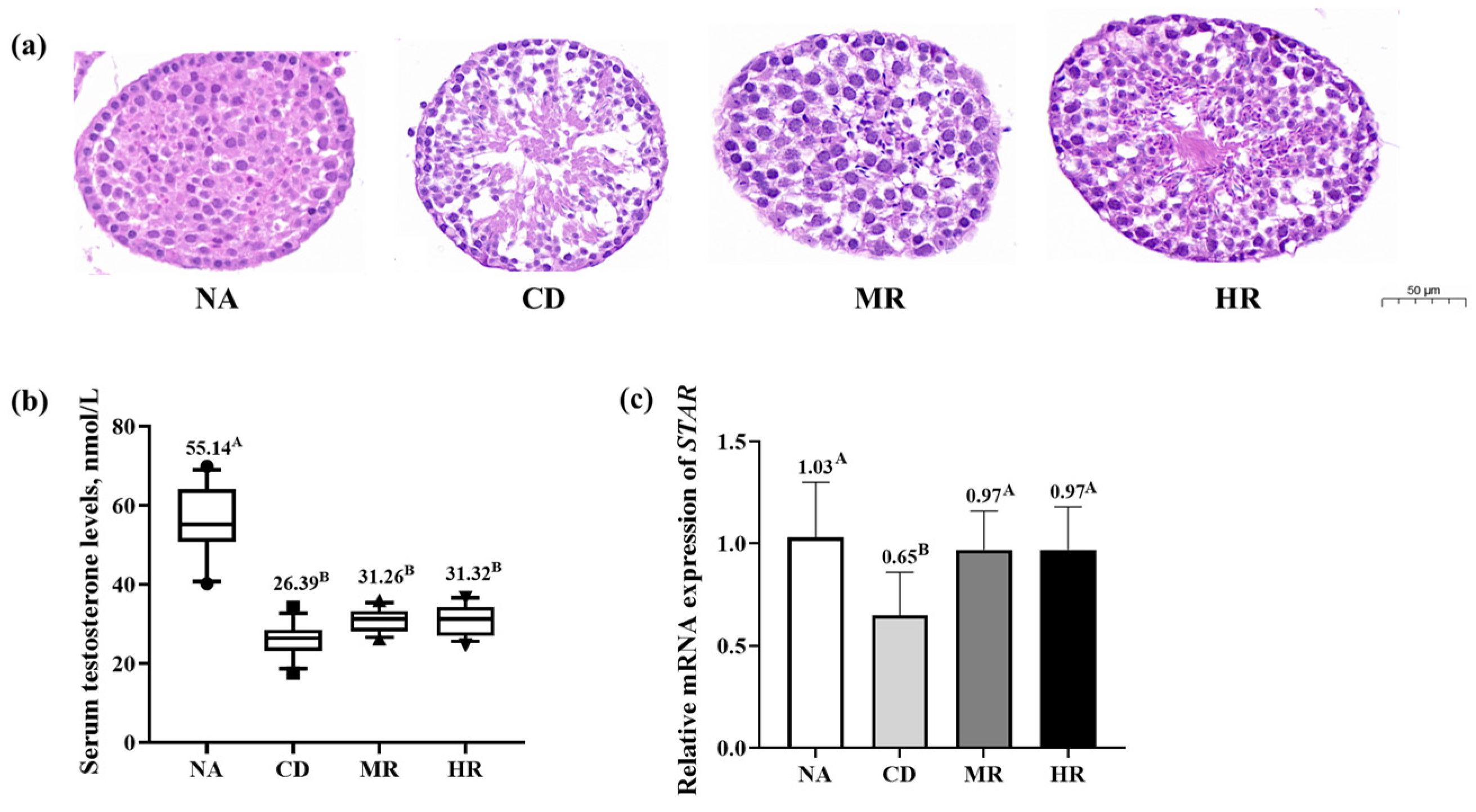
3.2. Testicular Histopathology Analysis and Sex Hormone Levels of Aging Mice with Methionine Restriction
3.3. Sperm Quality of Aging Mice with Methionine Restriction
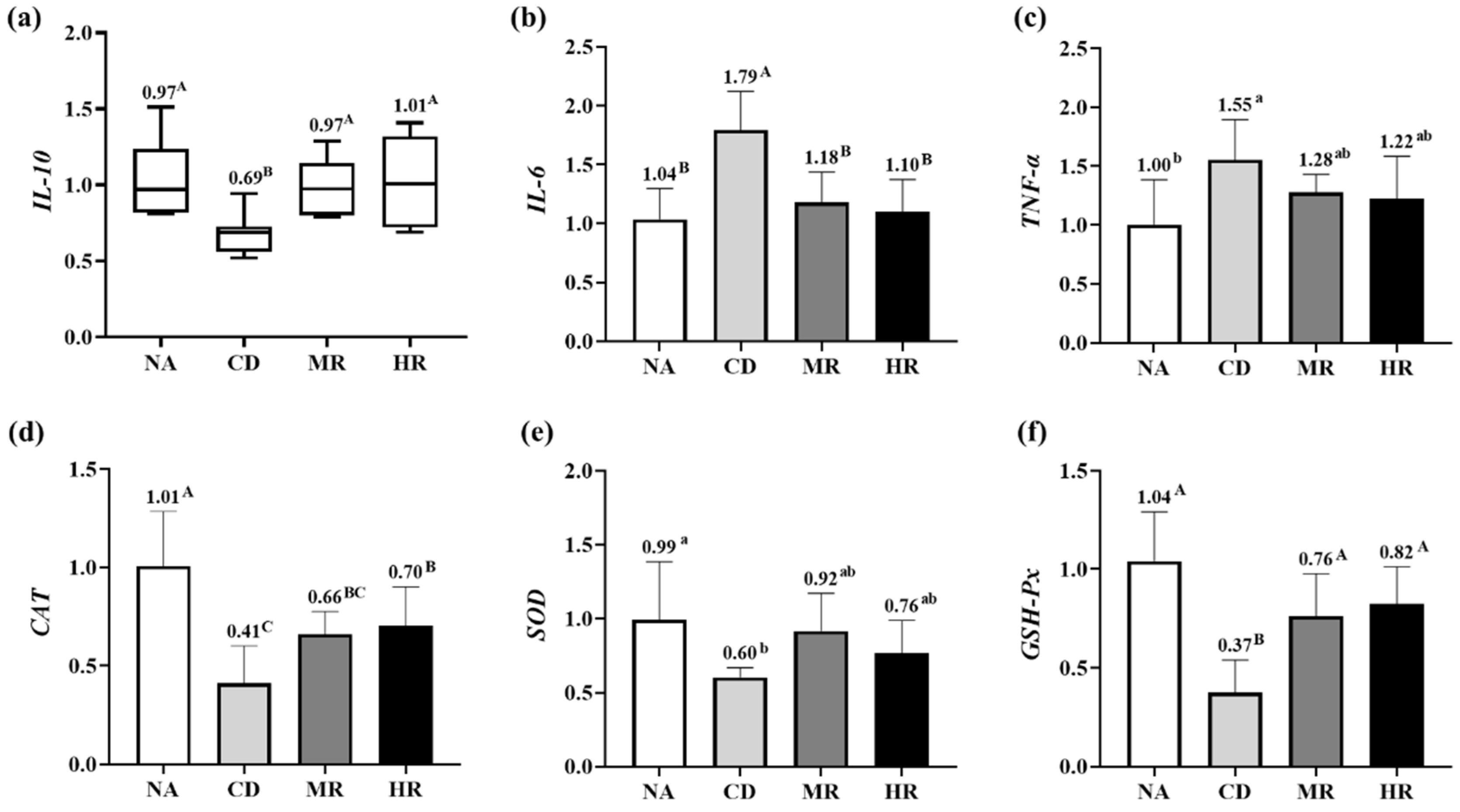
3.4. Levels of Inflammation and Oxidative Stress in Testes of Aging Mice with Methionine Restriction
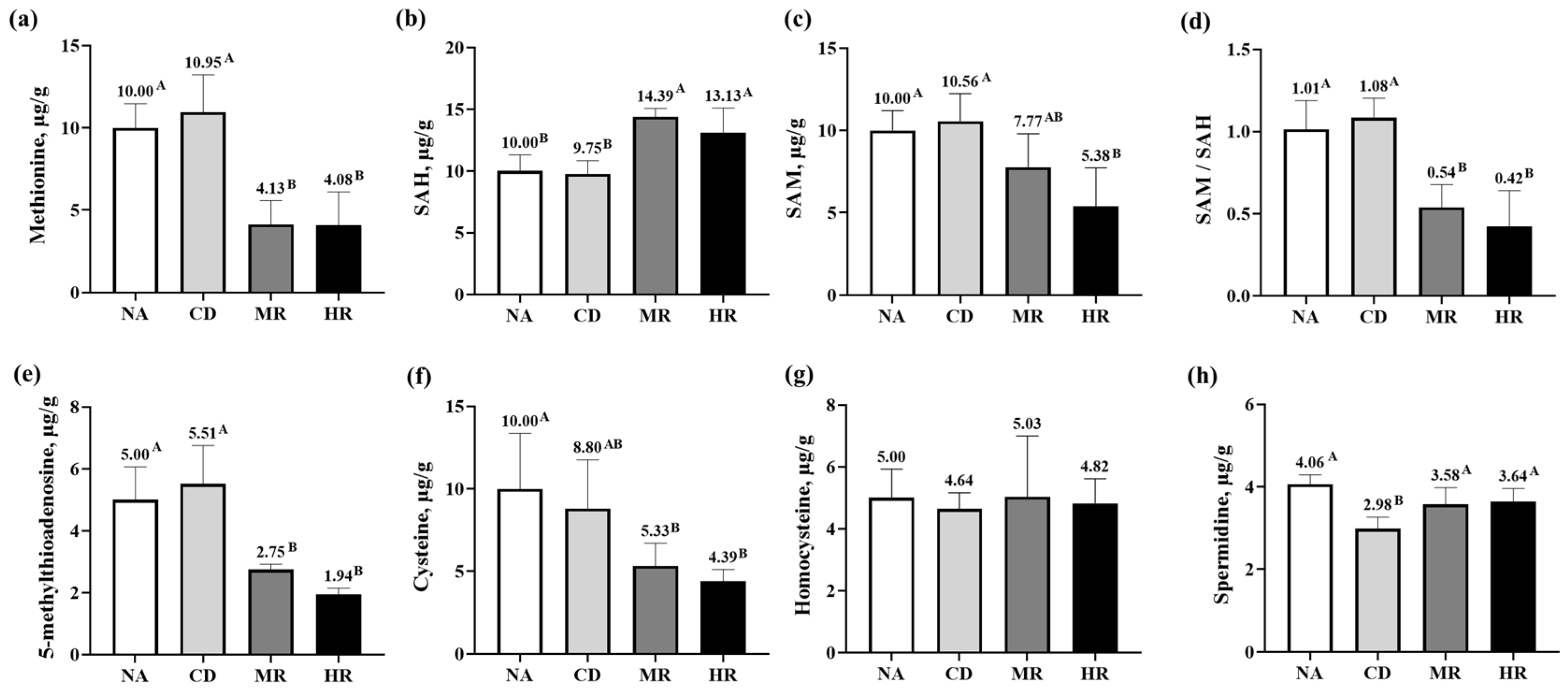
3.5. Methionine Metabolites in Testes of Aging Mice with Methionine Restriction
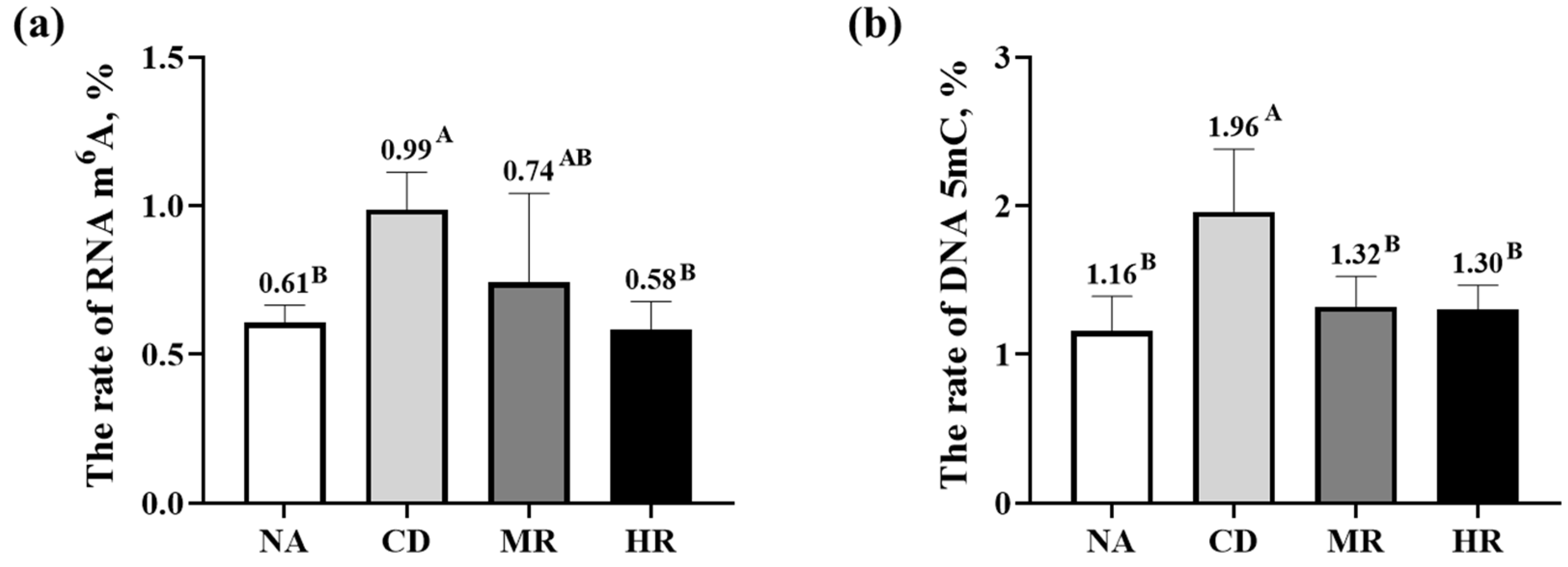
3.6. DNA and RNA Methylation Levels in Sperm of Aging Mice with Methionine Restriction
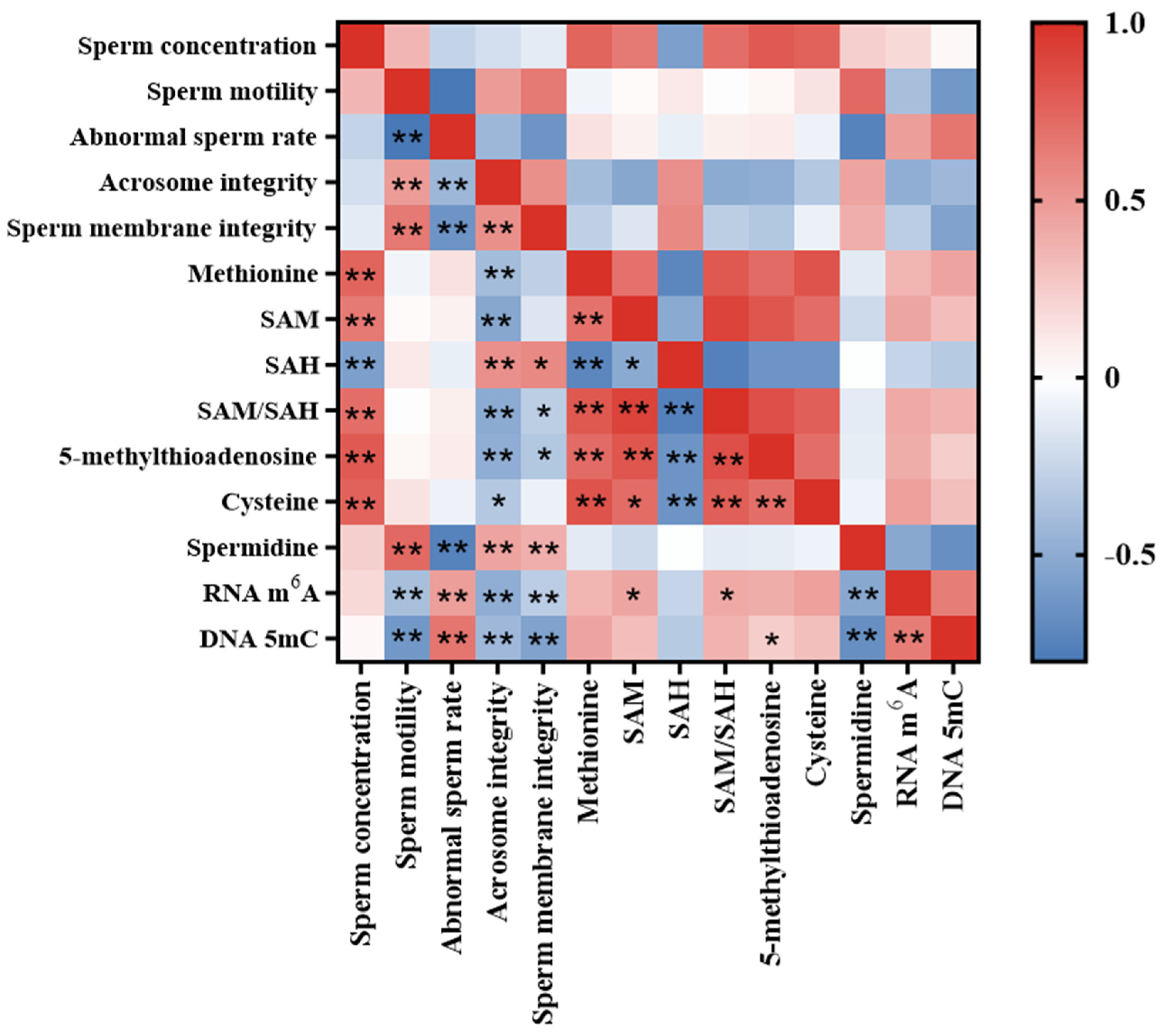
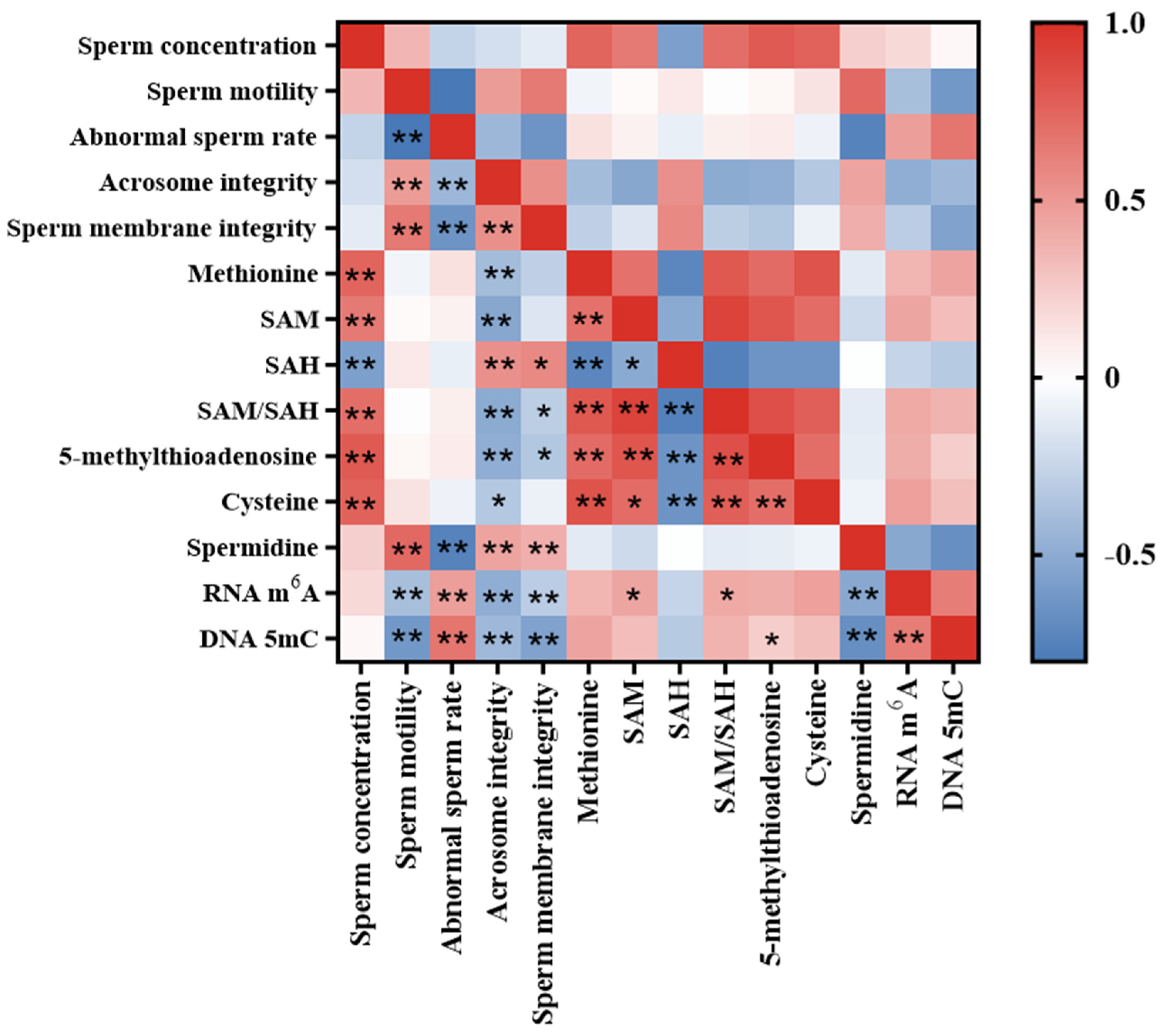
3.7. Correlation Analysis of Sperm Quality Parameters, Methionine Metabolite Level and Sperm RNA m6A and DNA 5mC Level in Aging Mice
4. Discussion
4.1. Effects of Methionine Restriction on Sperm Quality in Aging Mice
4.2. Effects of Methionine Restriction on Inflammation and Oxidative Stress in Testes of Aging Mice
4.3. Effects of Methionine Restriction on Spermidine Content in Testes of Aging Mice
4.4. Effects of Methionine Restriction on Sperm RNA m6A and DNA 5mC Levels in Aging Mice
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, J.A.; Hamilton, B.E.; Ventura, S.J. Births: Final Data for 2011; National vital statistics reports: From the Centers for Disease Control and Prevention, National Center for Health Statistics, National Vital Statistics System; National Center for Health Statistics: Hyattsville, MD, USA, 2013; Volume 62, pp. 1–72.
- Jung, A.; Schuppe, H.C.; Schill, W.B. Comparison of semen quality in older and younger men attending an andrology clinic. Andrologia 2002, 34, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Eskenazi, B.; Wyrobek, A.J.; Sloter, E. The association of age and semen quality in healthy men. Hum. Reprod. 2003, 18, 447–454. [Google Scholar] [CrossRef]
- Chianese, C.; Brilli, S.; Krausz, C. Genomic changes in spermatozoa of the aging male. Adv. Exp. Med. Biol. 2014, 791, 13–26. [Google Scholar] [PubMed]
- Serre, V.; Robaire, B. Paternal age affects fertility and progeny outcome in the Brown Norway rat. Fertil. Steril. 1998, 70, 625–631. [Google Scholar] [CrossRef]
- Zhang, F.; Jiang, L. Neuroinflammation in Alzheimer’s disease. Neuropsychiatr. Dis. Treat. 2015, 11, 243–256. [Google Scholar] [CrossRef]
- Lohan, S.B.; Vitt, K.; Scholz, P. ROS production and glutathione response in keratinocytes after application of β-carotene and VIS/NIR irradiation. Chem-Biol. Interact. 2018, 280, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Vanyushin, B.F.; Nemirovsky, L.E.; Klimenko, V.V. The 5-methylcytosine in DNA of rats. Tissue and age specificity and the changes induced by hydrocortisone and other agents. Gerontologia 1973, 19, 138–152. [Google Scholar] [CrossRef]
- Tammen, S.A.; Dolnikowski, G.G.; Ausman, L.M. Aging and alcohol interact to alter hepatic DNA hydroxymethylation. Alcohol. Clin. Exp. Res. 2014, 38, 2178–2185. [Google Scholar] [CrossRef]
- Jenkins, T.G.; Aston, K.I.; Cairns, B.R. Paternal aging and associated intraindividual alterations of global sperm 5-methylcytosine and 5-hydroxymethylcytosine levels. Fertil. Steril. 2013, 100, 945–951. [Google Scholar] [CrossRef]
- Oakes, C.C.; Smiraglia, D.J.; Plass, C. Aging results in hypermethylation of ribosomal DNA in sperm and liver of male rats. Proc. Natl. Acad. Sci. USA 2003, 100, 1775–1780. [Google Scholar] [CrossRef]
- Barzideh, J.; Scott, R.J.; Aitken, R.J. Analysis of the global methylation status of human spermatozoa and its association with the tendency of these cells to enter apoptosis. Andrologia 2013, 45, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Dixon, T.; Jung, S.; Graff, E.; Forney, L.; Gettys, T.; Wanders, D. Dietary methionine restriction reduces inflammation independent of FGF21 action. Obesity 2019, 27, 78. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.C.; Kaya, A.; Ma, S. Methionine restriction extends lifespan of Drosophila melanogaster under conditions of low amino-acid status. Nat. Commun. 2014, 5, 3592. [Google Scholar] [CrossRef] [PubMed]
- Mattocks, D.A.; Mentch, S.J.; Shneyder, J. Short term methionine restriction increases hepatic global DNA methylation in adult but not young male C57BL/6J mice. Exp. Gerontol. 2017, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wallis, K.F.; Melnyk, S.B.; Miousse, I.R. Sex-specific effects of dietary methionine restriction on the iintestinal microbiome. Nutrients 2020, 12, 781. [Google Scholar] [CrossRef]
- Tamanna, N.; Kroeker, K.; Braun, K.; Banh, S.; Treberg, J.R. The effect of short-term methionine restriction on glutathione synthetic capacity and antioxidant responses at the whole tissue and mitochondrial level in the rat liver. Exp. Gerontol. 2019, 127, 110712. [Google Scholar] [CrossRef]
- Martínez, Y.; Li, X.; Liu, G. The role of methionine on metabolism, oxidative stress, and diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef]
- Xu, Y.; Yang, Y.; Sun, J. Dietary methionine restriction ameliorates the impairment of learning and memory function induced by obesity in mice. Food Funct. 2019, 10, 1411–1425. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, Y.; Sun, J.; Zhang, J.; Guo, H.; Shi, Y.; Cheng, X.; Tang, X.; Le, G. Dietary methionine restriction reduces hepatic steatosis and oxidative stress in high-fat-fed mice by promoting H2S production. Food Funct. 2019, 10, 61–77. [Google Scholar] [CrossRef]
- Fang, Z.; Huang, F.; Luo, J. Effects of dl-2-hydroxy-4-methylthiobutyrate on the first-pass intestinal metabolism of dietary methionine and its extra-intestinal availability. Br. J. Nutr. 2010, 103, 643–651. [Google Scholar] [CrossRef]
- Zuo, F.; Gu, Q.; Peng, J. Effects of Different Methionine Sources on Methionine Metabolism in the IPEC-J2 Cells. BioMed Res. Int. 2019, 2019, 5464906. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cheng, B.; Su, Y.; Li, M.; Ma, S.; Zhang, Y.; Zhang, A.; Cai, S.; Bao, Q.; Wang, S.; et al. The potential role of m6A RNA methylation in the aging process and aging-associated diseases. Front. Genet. 2022, 13, 869950. [Google Scholar] [CrossRef] [PubMed]
- Azman, K.F.; Zakaria, R. D-Galactose-induced accelerated aging model: An overview. Biogerontology 2019, 20, 763–782. [Google Scholar] [CrossRef] [PubMed]
- Kozink, D.M.; Estienne, M.J.; Harper, A.F. Effects of dietary L-carnitine supplementation on semen characteristics in boars. Theriogenology 2004, 61, 1247–1258. [Google Scholar] [CrossRef]
- Zuo, F.; Gu, Q.; Peng, J. Simultaneous Quantification of Methionine-Related Metabolites and Co-factors in IPEC-J2 and PIEC Cells by LC–MS/MS. Chromatographia 2020, 83, 361–371. [Google Scholar] [CrossRef]
- Kitchen, R.R.; Kubista, M.; Tichopad, A. Statistical aspects of quantitative real-time PCR experiment design. Methods 2010, 50, 231–236. [Google Scholar] [CrossRef]
- Song, X.; Bao, M.; Li, D.; Li, Y.M. Advanced glycation in D-galactose induced mouse aging model. Mech. Ageing Dev. 1999, 108, 239–251. [Google Scholar] [CrossRef]
- Bucala, R.; Cerami, A. Advanced glycosylation: Chemistry, biology, and implications for diabetes and aging. Adv. Pharmacol. 1992, 23, 1–34. [Google Scholar]
- Lu, J.; Zheng, Y.L.; Wu, D.M.; Luo, L.; Sun, D.X.; Shan, Q. Ursolic acid ameliorates cognition deficits and attenuates oxidative damage in the brain of senescent mice induced by D-galactose. Biochem. Pharmacol. 2007, 74, 1078–1090. [Google Scholar] [CrossRef]
- Liao, C.H.; Chen, B.H.; Chiang, H.S.; Chen, C.W.; Chen, M.F.; Ke, C.C.; Wang, Y.Y.; Lin, W.N.; Wang, C.C.; Lin, Y.H. Optimizing a male reproductive aging mouse model by D-galactose injection. Int. J. Mol. Sci. 2016, 17, 98. [Google Scholar] [CrossRef]
- Elshorbagy, A.K.; Valdivia-Garcia, M.; Refsum, H. Sulfur amino acids in methionine-restricted rats: Hyperhomocysteinemia. Nutrition 2010, 26, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Bonsall, M.B.; Pizzari, T. Aging and sexual conflict. Science 2007, 316, 383–384. [Google Scholar] [CrossRef] [PubMed]
- Pino, V.; Sanz, A.; Valdés, N. The effects of aging on semen parameters and sperm DNA fragmentation. JBRA Assist. Reprod. 2020, 24, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Noro, T.; Namekata, K.; Kimura, A. Spermidine promotes retinal ganglion cell survival and optic nerve regeneration in adult mice following optic nerve injury. Cell Death Dis. 2015, 6, e1720. [Google Scholar] [CrossRef]
- Qi, Y.; Qiu, Q.; Gu, X. ATM mediates spermidine-induced mitophagy via PINK1 and Parkin regulation in human fibroblasts. Sci. Rep. 2016, 6, 24700. [Google Scholar] [CrossRef]
- Madeo, F.; Eisenberg, T.; Pietrocola, F. Spermidine in health and disease. Science 2018, 359, 2788. [Google Scholar] [CrossRef]
- Matyi, S.; Jackson, J.; Garrett, K. The effect of different levels of dietary restriction on glucose homeostasis and metabolic memory. GeroScience 2018, 40, 139–149. [Google Scholar] [CrossRef]
- Fang, H.; Stone, K.P.; Ghosh, S. The Role of Reduced Methionine in Mediating the Metabolic Responses to Protein Restriction Using Different Sources of Protein. Nutrients 2021, 13, 2609. [Google Scholar] [CrossRef]
- Sun, L.; Sadighi Akha, A.A.; Miller, R.A. Life-span extension in mice by preweaning food restriction and by methionine restriction in middle age. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2009, 64, 711–722. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | 0.86% Methionine, % | 0.17% Methionine, % | 0.17% HMTBA, % |
|---|---|---|---|
| Amino acid mixture 1 | 7.80 | 7.80 | 7.80 |
| L-methione | 0.86 | 0.17 | - |
| HMTBA 2 | - | - | 0.17 |
| L-glutamic acid | - | 0.69 | 0.69 |
| Starch and sucrose | 68.64 | 68.64 | 68.64 |
| Soybean oil | 8.00 | 8.00 | 8.00 |
| Dextrin | 5.00 | 5.00 | 5.00 |
| Cellulose | 5.00 | 5.00 | 5.00 |
| Choline bitartrate | 0.20 | 0.20 | 0.20 |
| Mineral and vitamin mixture | 4.50 | 4.50 | 4.50 |
| Total | 100.00 | 100.00 | 100.00 |
| Gene Symbol | PCR Primer (5′-3′) |
|---|---|
| Steroidogenic acute regulatory protein (STAR) | AACGGGGACGAAGTGCTAAG CCGTGTCTTTTCCAATCCTCTG |
| Interleukin 10 (IL-10) | AGTCTTGGGCTACCTCC |
| CTCGCTTCACTATCTGTCTG | |
| Interleukin 6 (IL-6) | GGCGATGCGTCTAAGGAA |
| GAAGGACTGGGCTGAAAATAAGGGGT | |
| Tumor necrosis factor α (TNF-α) | TAGGCGAGGGAACAAGAG |
| TGGGCAACCGAGTCATAC | |
| Catalase (CAT) | GACAGATCAGATGAAGATGACTGGT |
| GTGAGGAGGACAATTATTTCCAGTT | |
| Superoxide dismutase (SOD) | TCAAGAATTCTGAAATGTGGAAGA |
| GAGCAGAAGTTGCAGATCTTTTAGA | |
| Glutathione peroxidase (GSH-Px) | AAGCTGTGCAAGGTGAGGAA |
| CTTGATGGCATCGGTGATTT | |
| GAPDH | CCTCCAAGGAGTAAGAGCC |
| GTCTGGGATGGAAACTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Li, H.; Miao, Y.; Peng, J.; Wei, H. Effects of Methionine Restriction from Different Sources on Sperm Quality in Aging Mice. Nutrients 2023, 15, 4782. https://doi.org/10.3390/nu15224782
Wu Y, Li H, Miao Y, Peng J, Wei H. Effects of Methionine Restriction from Different Sources on Sperm Quality in Aging Mice. Nutrients. 2023; 15(22):4782. https://doi.org/10.3390/nu15224782
Chicago/Turabian StyleWu, Yinghui, Hao Li, Yueyue Miao, Jian Peng, and Hongkui Wei. 2023. "Effects of Methionine Restriction from Different Sources on Sperm Quality in Aging Mice" Nutrients 15, no. 22: 4782. https://doi.org/10.3390/nu15224782
APA StyleWu, Y., Li, H., Miao, Y., Peng, J., & Wei, H. (2023). Effects of Methionine Restriction from Different Sources on Sperm Quality in Aging Mice. Nutrients, 15(22), 4782. https://doi.org/10.3390/nu15224782