

Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin

 ,
,  ,
, 
 and
and 
Abstract
1. Introduction
2. Sources of Apigenin
3. Pharmacological Actions of Apigenin
4. Therapeutic Potential of Apigenin in Cardiovascular Disorders
4.1. Atherosclerosis
4.2. Cerebrovascular Diseases
4.3. Myocardial Ischemia
4.4. Cardiovascular Complications of Diabetes
4.5. Hypertension
4.6. Drug-Induced Cardiotoxicity and Cardiomyopathy
4.7. Aortic Aneurysm
4.8. Cardiac Hypertrophy
5. Pharmacokinetics and Toxicity of Apigenin
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABCA1 | ATP-binding cassette A1 |
| BCS | Biopharmaceutics classification system |
| GSK-3β | Glycogen synthase kinase-3β |
| HBMVEC | Human brain microvascular endothelial cells |
| HAEC | Human aortic endothelial cells |
| HUVEC | Human umbilical vein endothelial cells |
| HO-1 | Heme oxygenase-1 |
| HMGCR | 3-hydroxy-3-methylglutaryl coenzyme A reductase |
| ICAM-1 | Intercellular adhesion molecule-1 |
| JAK2-STAT3 | Janus kinase 2/signal transducer and activator of transcription |
| LDH | Lactate dehydrogenase |
| MCP-1 | Monocyte chemoattractant protein-1 |
| MAPK | Mitogen-activated protein kinase |
| NF-κB | Nuclear factor-kappa B |
| Nrf2 | Nuclear factor-erythroid factor 2-related factor 2 |
| Npc1l1 | Niemann-Pick C1-like 1 |
| NQO1 | NAD(P)H quinone dehydrogenase 1 |
| OGD-R | Oxygen and glucose deprivation and re-oxygenation |
| PAI-2 | Plasminogen activator inhibitor-2 |
| SREBP-2 | Sterol regulatory element binding protein-2 |
| TNFα | Tumor necrosis factor-α |
| TLR | Toll like receptor |
| VEGF | Vascular endothelial growth factor |
| VCAM-1 | Vascular cell adhesion molecule-1 |
References
- Wang, M.; Firrman, J.; Liu, L.; Yam, K. A Review on Flavonoid Apigenin: Dietary Intake, ADME, Antimicrobial Effects, and Interactions with Human Gut Microbiota. BioMed Res. Int. 2019, 2019, 7010467. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Wang, F.; Zhou, R.; Song, X.; Xie, M. Apigenin: A Current Review on Its Beneficial Biological Activities. J. Food Biochem. 2017, 41, e12376. [Google Scholar] [CrossRef]
- Tamer, F.; Tullemans, B.M.E.; Kuijpers, M.J.E.; Claushuis, T.A.M.; Heemskerk, J.W.M. Nutrition Phytochemicals Affecting Platelet Signaling and Responsiveness: Implications for Thrombosis and Hemostasis. Thromb. Haemost. 2021, 122, 879–894. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Khan, H.; D’onofrio, G.; Šamec, D.; Shirooie, S.; Dehpour, A.R.; Argüelles, S.; Habtemariam, S.; Sobarzo-Sanchez, E. Apigenin as Neuroprotective Agent: Of Mice and Men. Pharmacol. Res. 2018, 128, 359–365. [Google Scholar] [CrossRef]
- Salehi, B.; Venditti, A.; Sharifi-Rad, M.; Kręgiel, D.; Sharifi-Rad, J.; Durazzo, A.; Lucarini, M.; Santini, A.; Souto, E.; Novellino, E.; et al. The Therapeutic Potential of Apigenin. Int. J. Mol. Sci. 2019, 20, 1305. [Google Scholar] [CrossRef] [PubMed]
- Alam, W.; Rocca, C.; Khan, H.; Hussain, Y.; Aschner, M.; De Bartolo, A.; Amodio, N.; Angelone, T.; Cheang, W.S. Current Status and Future Perspectives on Therapeutic Potential of Apigenin: Focus on Metabolic-Syndrome-Dependent Organ Dysfunction. Antioxidants 2021, 10, 1643. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, J.K.; Shankar, E.; Gupta, S. Chamomile: A Herbal Medicine of the Past with a Bright Future (Review). Mol. Med. Rep. 2010, 3, 895–901. [Google Scholar] [CrossRef]
- Shankar, E.; Goel, A.; Gupta, K.; Gupta, S. Plant Flavone Apigenin: An Emerging Anticancer Agent. Curr. Pharmacol. Rep. 2017, 3, 423–446. [Google Scholar] [CrossRef]
- Database on Polyphenol Content in Foods. Phenol-Explorer. Available online: http://phenol-explorer.eu/ (accessed on 3 May 2022).
- Flavonoids: USDA ARS. Available online: https://www.ars.usda.gov/northeast-area/beltsville-md-bhnrc/beltsville-human-nutrition-research-center/methods-and-application-of-food-composition-laboratory/mafcl-site-pages/flavonoids/ (accessed on 1 May 2022).
- Thiruvengadam, M.; Venkidasamy, B.; Subramanian, U.; Samynathan, R.; Ali Shariati, M.; Rebezov, M.; Girish, S.; Thangavel, S.; Dhanapal, A.R.; Fedoseeva, N.; et al. Bioactive Compounds in Oxidative Stress-Mediated Diseases: Targeting the NRF2/ARE Signaling Pathway and Epigenetic Regulation. Antioxidants 2021, 10, 1859. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, J.; Wang, Y.; Fa, X. Apigenin Attenuates Copper-Mediated β-Amyloid Neurotoxicity through Antioxidation, Mitochondrion Protection and MAPK Signal Inactivation in an AD Cell Model. Brain Res. 2013, 1492, 33–45. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Wang, H. Protective Roles of Apigenin Against Cardiometabolic Diseases: A Systematic Review. Front. Nutr. 2022, 9, 875826. [Google Scholar] [CrossRef]
- Jung, U.J.; Cho, Y.-Y.; Choi, M.-S. Apigenin Ameliorates Dyslipidemia, Hepatic Steatosis and Insulin Resistance by Modulating Metabolic and Transcriptional Profiles in the Liver of High-Fat Diet-Induced Obese Mice. Nutrients 2016, 8, 305. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, B. Apigenin Protects Ovalbumin-Induced Asthma through the Regulation of Th17 Cells. Fitoterapia 2013, 91, 298–304. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.-T.; Xiao, L.; Zhu, L.; Wang, Q.; Yan, T. Anti-Inflammatory Effects of Apigenin in Lipopolysaccharide-Induced Inflammatory in Acute Lung Injury by Suppressing COX-2 and NF-KB Pathway. Inflammation 2014, 37, 2085–2090. [Google Scholar] [CrossRef]
- Lv, X.; Qiu, M.; Chen, D.; Zheng, N.; Jin, Y.; Wu, Z. Apigenin Inhibits Enterovirus 71 Replication through Suppressing Viral IRES Activity and Modulating Cellular JNK Pathway. Antivir. Res. 2014, 109, 30–41. [Google Scholar] [CrossRef]
- Qian, S.; Fan, W.; Qian, P.; Zhang, D.; Wei, Y.; Chen, H.; Li, X. Apigenin Restricts FMDV Infection and Inhibits Viral IRES Driven Translational Activity. Viruses 2015, 7, 1613–1626. [Google Scholar] [CrossRef]
- Bahoran, T.; Soobrattee, M.A.; Luximon-Ramma, V.; Aruoma, O.I. Free Radicals and Antioxidants in Cardiovascular Health and Disease. Internet J. Med. Update 2007, 1, 25–41. [Google Scholar] [CrossRef]
- Zhang, K.; Song, W.; Li, D.; Jin, X. Apigenin in the Regulation of Cholesterol Metabolism and Protection of Blood Vessels. Exp. Ther. Med. 2017, 13, 1719–1724. [Google Scholar] [CrossRef]
- Wang, Q.; Zeng, P.; Liu, Y.; Wen, G.; Fu, X.; Sun, X. Inhibition of Autophagy Ameliorates Atherogenic Inflammation by Augmenting Apigenin-Induced Macrophage Apoptosis. Int. Immunopharmacol. 2015, 27, 24–31. [Google Scholar] [CrossRef]
- Wong, T.Y.; Tan, Y.Q.; Lin, S.; Leung, L.K. Apigenin and Luteolin Display Differential Hypocholesterolemic Mechanisms in Mice Fed a High-Fat Diet. Biomed. Pharmacother. 2017, 96, 1000–1007. [Google Scholar] [CrossRef]
- Wong, T.Y.; Tan, Y.Q.; Lin, S.; Leung, L.K. Co-Administrating Apigenin in a High-Cholesterol Diet Prevents Hypercholesterolaemia in Golden Hamsters. J. Pharm. Pharmacol. 2018, 70, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, Y.; Du, C.; Wang, L.; Xiao, Y. Effects of Apigenin on the Expression of LOX-1, Bcl-2, and Bax in Hyperlipidemia Rats. Chem. Biodivers. 2021, 18, e2100049. [Google Scholar] [CrossRef] [PubMed]
- Yamagata, K.; Hashiguchi, K.; Yamamoto, H.; Tagami, M. Dietary Apigenin Reduces Induction of LOX-1 and NLRP3 Expression, Leukocyte Adhesion, and Acetylated Low-Density Lipoprotein Uptake in Human Endothelial Cells Exposed to Trimethylamine-N-Oxide. J. Cardiovasc. Pharmacol. 2019, 74, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Zeng, P.; Liu, B.; Wang, Q.; Fan, Q.; Diao, J.-X.; Tang, J.; Fu, X.-Q.; Sun, X.-G. Apigenin Attenuates Atherogenesis through Inducing Macrophage Apoptosis via Inhibition of AKT Ser473 Phosphorylation and Downregulation of Plasminogen Activator Inhibitor-2. Oxidative Med. Cell. Longev. 2015, 2015, 379538. [Google Scholar] [CrossRef]
- Clayton, Z.S.; Hutton, D.A.; Brunt, V.E.; VanDongen, N.S.; Ziemba, B.P.; Casso, A.G.; Greenberg, N.T.; Mercer, A.N.; Rossman, M.J.; Campisi, J.; et al. Apigenin Restores Endothelial Function by Ameliorating Oxidative Stress, Reverses Aortic Stiffening, and Mitigates Vascular Inflammation with Aging. Am. J. Physiol. Heart Circ. Physiol. 2021, 321, H185–H196. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.S.; Sabu, V.; Sindhu, G.; Rauf, A.A.; Helen, A. Isolation, Identification and Characterization of Apigenin from Justicia Gendarussa and Its Anti-Inflammatory Activity. Int. Immunopharmacol. 2018, 59, 157–167. [Google Scholar] [CrossRef]
- Morgan, M.J.; Liu, Z. Crosstalk of Reactive Oxygen Species and NF-ΚB Signaling. Cell Res. 2011, 21, 103–115. [Google Scholar] [CrossRef]
- Ren, K.; Jiang, T.; Zhou, H.-F.; Liang, Y.; Zhao, G.-J. Apigenin Retards Atherogenesis by Promoting ABCA1-Mediated Cholesterol Efflux and Suppressing Inflammation. Cell Physiol. Biochem. 2018, 47, 2170–2184. [Google Scholar] [CrossRef]
- Jeong, Y.-J.; Choi, Y.-J.; Choi, J.-S.; Kwon, H.-M.; Kang, S.-W.; Bae, J.-Y.; Lee, S.-S.; Kang, J.-S.; Han, S.J.; Kang, Y.-H. Attenuation of Monocyte Adhesion and Oxidised LDL Uptake in Luteolin-Treated Human Endothelial Cells Exposed to Oxidised LDL. Br. J. Nutr. 2007, 97, 447–457. [Google Scholar] [CrossRef]
- Kuriakose, D.; Xiao, Z. Pathophysiology and Treatment of Stroke: Present Status and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 7609. [Google Scholar] [CrossRef]
- Cai, M.; Ma, Y.; Zhang, W.; Wang, S.; Wang, Y.; Tian, L.; Peng, Z.; Wang, H.; Qingrong, T. Apigenin-7-O-β-D-(-6″-p-Coumaroyl)-Glucopyranoside Treatment Elicits Neuroprotective Effect against Experimental Ischemic Stroke. Int. J. Biol. Sci. 2016, 12, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xu, S.; Duan, H.; Zhu, Z.; Yang, Z.; Cao, J.; Zhao, Y.; Huang, Z.; Wu, Q.; Duan, J. A Novel, Highly-Water-Soluble Apigenin Derivative Provides Neuroprotection Following Ischemia in Male Rats by Regulating the ERK/Nrf2/HO-1 Pathway. Eur. J. Pharmacol. 2019, 855, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, S.; Sun, S.; Lu, Y.; Gao, K.; Guo, C.; Li, R.; Li, W.; Zhao, X.; Tang, H.; et al. Apigenin-7-O-β-D-(-6″-p-Coumaroyl)-Glucopyranoside Treatment Elicits a Neuroprotective Effect through GSK-3β Phosphorylation-Mediated Nrf2 Activation. Aging 2020, 12, 23872–23888. [Google Scholar] [CrossRef] [PubMed]
- Tu, F.; Pang, Q.; Chen, X.; Huang, T.; Liu, M.; Zhai, Q. Angiogenic Effects of Apigenin on Endothelial Cells after Hypoxia-Reoxygenation via the Caveolin-1 Pathway. Int. J. Mol. Med. 2017, 40, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Pang, Q.; Zhao, Y.; Chen, X.; Zhao, K.; Zhai, Q.; Tu, F. Apigenin Protects the Brain against Ischemia/Reperfusion Injury via Caveolin-1/VEGF In Vitro and In Vivo. Oxidative Med. Cell. Longev. 2018, 2018, 7017204. [Google Scholar] [CrossRef] [PubMed]
- Tu, F.; Pang, Q.; Huang, T.; Zhao, Y.; Liu, M.; Chen, X. Apigenin Ameliorates Post-Stroke Cognitive Deficits in Rats Through Histone Acetylation-Mediated Neurochemical Alterations. Med. Sci. Monit. 2017, 23, 4004–4013. [Google Scholar] [CrossRef]
- Ling, C.; Lei, C.; Zou, M.; Cai, X.; Xiang, Y.; Xie, Y.; Li, X.; Huang, D.; Wang, Y. Neuroprotective Effect of Apigenin against Cerebral Ischemia/Reperfusion Injury. J. Int. Med. Res. 2020, 48, 030006052094585. [Google Scholar] [CrossRef]
- Han, Y.; Zhang, T.; Su, J.; Zhao, Y.; Wang, C.; Li, X. Apigenin Attenuates Oxidative Stress and Neuronal Apoptosis in Early Brain Injury Following Subarachnoid Hemorrhage. J. Clin. Neurosci. 2017, 40, 157–162. [Google Scholar] [CrossRef]
- Zhang, T.; Su, J.; Guo, B.; Wang, K.; Li, X.; Liang, G. Apigenin Protects Blood–Brain Barrier and Ameliorates Early Brain Injury by Inhibiting TLR4-Mediated Inflammatory Pathway in Subarachnoid Hemorrhage Rats. Int. Immunopharmacol. 2015, 28, 79–87. [Google Scholar] [CrossRef]
- Fraccarollo, D.; Galuppo, P.; Bauersachs, J. Novel Therapeutic Approaches to Post-Infarction Remodelling. Cardiovasc. Res. 2012, 94, 293–303. [Google Scholar] [CrossRef]
- Hu, J.; Li, Z.; Xu, L.; Sun, A.; Fu, X.; Zhang, L.; Jing, L.; Lu, A.; Dong, Y.; Jia, Z. Protective Effect of Apigenin on Ischemia/Reperfusion Injury of the Isolated Rat Heart. Cardiovasc. Toxicol. 2015, 15, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yang, J.; Hu, J.; Li, X.; Zhang, X.; Li, Z. Apigenin Attenuates Myocardial Ischemia/Reperfusion Injury via the Inactivation of P38 Mitogen-Activated Protein Kinase. Mol. Med. Rep. 2015, 12, 6873–6878. [Google Scholar] [CrossRef]
- Chen, C.; He, H.; Luo, Y.; Zhou, M.; Yin, D.; He, M. Involvement of Bcl-2 Signal Pathway in the Protective Effects of Apigenin on Anoxia/Reoxygenation-Induced Myocardium Injury. J. Cardiovasc. Pharmacol. 2016, 67, 152–163. [Google Scholar] [CrossRef]
- Quan, W.; Ma, S.; Zhu, Y.; Shao, Q.; Hou, J.; Li, X. Apigenin-7-O-β-d-(6″-p-Coumaroyl)-Glucopyranoside Reduces Myocardial Ischaemia/Reperfusion Injury in an Experimental Model via Regulating the Inflammation Response. Pharm. Biol. 2020, 58, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, L.; Xiao, Y. Apigenin Protects against Ischemia-/Hypoxia-Induced Myocardial Injury by Mediating Pyroptosis and Apoptosis. In Vitro Cell. Dev.Biol. Anim. 2020, 56, 307–312. [Google Scholar] [CrossRef]
- Feng, Y.; Lu, Y.; Liu, D.; Zhang, W.; Liu, J.; Tang, H.; Zhu, Y. Apigenin-7-O-β-d-(-6″-p-Coumaroyl)-Glucopyranoside Pretreatment Attenuates Myocardial Ischemia/Reperfusion Injury via Activating AMPK Signaling. Life Sci. 2018, 203, 246–254. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, Y.; Lin, L.; Zhou, J. Apigenin Suppresses the Apoptosis of H9C2 Rat Cardiomyocytes Subjected to Myocardial Ischemia-Reperfusion Injury via Upregulation of the PI3K/Akt Pathway. Mol. Med. Rep. 2018, 18, 1560–1570. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Di, S.; Hu, W.; Feng, Y.; Zhou, Q.; Gong, B.; Tang, X.; Liu, J.; Zhang, W.; Xi, M.; et al. A New Flavonoid Glycoside (APG) Isolated from Clematis Tangutica Attenuates Myocardial Ischemia/Reperfusion Injury via Activating PKCε Signaling. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 701–711. [Google Scholar] [CrossRef]
- Wang, P.; Sun, J.; Lv, S.; Xie, T.; Wang, X. Apigenin Alleviates Myocardial Reperfusion Injury in Rats by Downregulating MiR-15b. Med. Sci. Monit. 2019, 25, 2764–2776. [Google Scholar] [CrossRef]
- Huang, H.; Lai, S.; Luo, Y.; Wan, Q.; Wu, Q.; Wan, L.; Qi, W.; Liu, J. Nutritional Preconditioning of Apigenin Alleviates Myocardial Ischemia/Reperfusion Injury via the Mitochondrial Pathway Mediated by Notch1/Hes1. Oxidative Med. Cell. Longev. 2019, 2019, 7973098. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, H.; Liu, Z.; Ma, Z.; An, D.; Xu, D. Apigenin Attenuates Myocardial Infarction-Induced Cardiomyocyte Injury by Modulating Parkin-Mediated Mitochondrial Autophagy. J. Biosci. 2020, 45, 75. [Google Scholar] [CrossRef] [PubMed]
- Ge, Z.-D.; Lian, Q.; Mao, X.; Xia, Z. Current Status and Challenges of NRF2 as a Potential Therapeutic Target for Diabetic Cardiomyopathy. Int. Heart J. 2019, 60, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-J.; Fan, Y.-L.; Liao, H.-H.; Liu, Y.; Chen, S.; Ma, Z.-G.; Zhang, N.; Yang, Z.; Deng, W.; Tang, Q.-Z. Apigenin Alleviates STZ-Induced Diabetic Cardiomyopathy. Mol. Cell. Biochem. 2017, 428, 9–21. [Google Scholar] [CrossRef]
- Mahajan, U.; Chandrayan, G.; Patil, C.; Arya, D.; Suchal, K.; Agrawal, Y.; Ojha, S.; Goyal, S. The Protective Effect of Apigenin on Myocardial Injury in Diabetic Rats Mediating Activation of the PPAR-γ Pathway. Int. J. Mol. Sci. 2017, 18, 756. [Google Scholar] [CrossRef]
- Qin, W.; Ren, B.; Wang, S.; Liang, S.; He, B.; Shi, X.; Wang, L.; Liang, J.; Wu, F. Apigenin and Naringenin Ameliorate PKCβII-Associated Endothelial Dysfunction via Regulating ROS/Caspase-3 and NO Pathway in Endothelial Cells Exposed to High Glucose. Vasc. Pharmacol. 2016, 85, 39–49. [Google Scholar] [CrossRef]
- Cao, Y.; Xie, L.; Liu, K.; Liang, Y.; Dai, X.; Wang, X.; Lu, J.; Zhang, X.; Li, X. The Antihypertensive Potential of Flavonoids from Chinese Herbal Medicine: A Review. Pharmacol. Res. 2021, 174, 105919. [Google Scholar] [CrossRef]
- Crowley, S.D. The Cooperative Roles of Inflammation and Oxidative Stress in the Pathogenesis of Hypertension. Antioxid. Redox Signal. 2014, 20, 102–120. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-L.; Yu, X.-J.; Hu, H.-B.; Yang, Q.-W.; Liu, K.-L.; Chen, Y.-M.; Zhang, Y.; Zhang, D.-D.; Tian, H.; Zhu, G.-Q.; et al. Apigenin Improves Hypertension and Cardiac Hypertrophy Through Modulating NADPH Oxidase-Dependent ROS Generation and Cytokines in Hypothalamic Paraventricular Nucleus. Cardiovasc. Toxicol. 2021, 21, 721–736. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Fang, X.; Shi, J.; Li, X.; Xie, M.; Liu, X. Apigenin Attenuates Pulmonary Hypertension by Inducing Mitochondria-Dependent Apoptosis of PASMCs via Inhibiting the Hypoxia Inducible Factor 1α–KV1.5 Channel Pathway. Chem. Biol. Interact. 2020, 317, 108942. [Google Scholar] [CrossRef]
- Paredes, M.; Romecín, P.; Atucha, N.; O’Valle, F.; Castillo, J.; Ortiz, M.; García-Estañ, J. Beneficial Effects of Different Flavonoids on Vascular and Renal Function in L-NAME Hypertensive Rats. Nutrients 2018, 10, 484. [Google Scholar] [CrossRef] [PubMed]
- Paredes, M.; Romecín, P.; Atucha, N.; O’Valle, F.; Castillo, J.; Ortiz, M.; García-Estañ, J. Moderate Effect of Flavonoids on Vascular and Renal Function in Spontaneously Hypertensive Rats. Nutrients 2018, 10, 1107. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Gao, P.; Pu, Y.; Li, Q.; Yang, T.; Zhang, H.; Xiong, S.; Cui, Y.; Li, L.; Ma, X.; et al. Activation of TRPV4 by Dietary Apigenin Antagonizes Renal Fibrosis in Deoxycorticosterone Acetate (DOCA)–Salt-Induced Hypertension. Clin. Sci. 2017, 131, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Hortal, M.D.; Varela-López, A.; Romero-Márquez, J.M.; Rivas-García, L.; Speranza, L.; Battino, M.; Quiles, J.L. Role of Flavonoids against Adriamycin Toxicity. Food Chem. Toxicol. 2020, 146, 111820. [Google Scholar] [CrossRef]
- Sahu, R.; Dua, T.K.; Das, S.; De Feo, V.; Dewanjee, S. Wheat Phenolics Suppress Doxorubicin-Induced Cardiotoxicity via Inhibition of Oxidative Stress, MAP Kinase Activation, NF-ΚB Pathway, PI3K/Akt/MTOR Impairment, and Cardiac Apoptosis. Food Chem. Toxicol. 2019, 125, 503–519. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.F.R.; Rakhshan, K.; Aboutaleb, N.; Nikbakht, F.; Naderi, N.; Bakhshesh, M.; Azizi, Y. Apigenin Attenuates Doxorubicin Induced Cardiotoxicity via Reducing Oxidative Stress and Apoptosis in Male Rats. Life Sci. 2019, 232, 116623. [Google Scholar] [CrossRef]
- Thangaiyan, R.; Robert, B.M.; Arjunan, S.; Govindasamy, K.; Nagarajan, R.P. Preventive Effect of Apigenin against Isoproterenol-Induced Apoptosis in Cardiomyoblasts. J. Biochem. Mol. Toxicol. 2018, 32, e22213. [Google Scholar] [CrossRef]
- Li, D.; Ma, J.; Wang, L.; Xin, S. Apigenin Prevent Abdominal Aortic Aneurysms Formation by Inhibiting the NF-KB Signaling Pathway. J. Cardiovasc. Pharmacol. 2020, 75, 11. [Google Scholar]
- Feng, W.; Ying, Z.; Ke, F.; Mei-Lin, X. Apigenin Suppresses TGF-Β1-Induced Cardiac Fibroblast Differentiation and Collagen Synthesis through the Downregulation of HIF-1α Expression by MiR-122-5p. Phytomedicine 2021, 83, 153481. [Google Scholar] [CrossRef]
- Wang, F.; Fan, K.; Zhao, Y.; Xie, M.-L. Apigenin Attenuates TGF-Β1-Stimulated Cardiac Fibroblast Differentiation and Extracellular Matrix Production by Targeting MiR-155-5p/c-Ski/Smad Pathway. J. Ethnopharmacol. 2021, 265, 113195. [Google Scholar] [CrossRef]
- Tang, D.; Chen, K.; Huang, L.; Li, J. Pharmacokinetic Properties and Drug Interactions of Apigenin, a Natural Flavone. Expert Opin. Drug Metab. Toxicol. 2017, 13, 323–330. [Google Scholar] [CrossRef]
- DeRango-Adem, E.F.; Blay, J. Does Oral Apigenin Have Real Potential for a Therapeutic Effect in the Context of Human Gastrointestinal and Other Cancers? Front. Pharmacol. 2021, 12, 681477. [Google Scholar] [CrossRef] [PubMed]


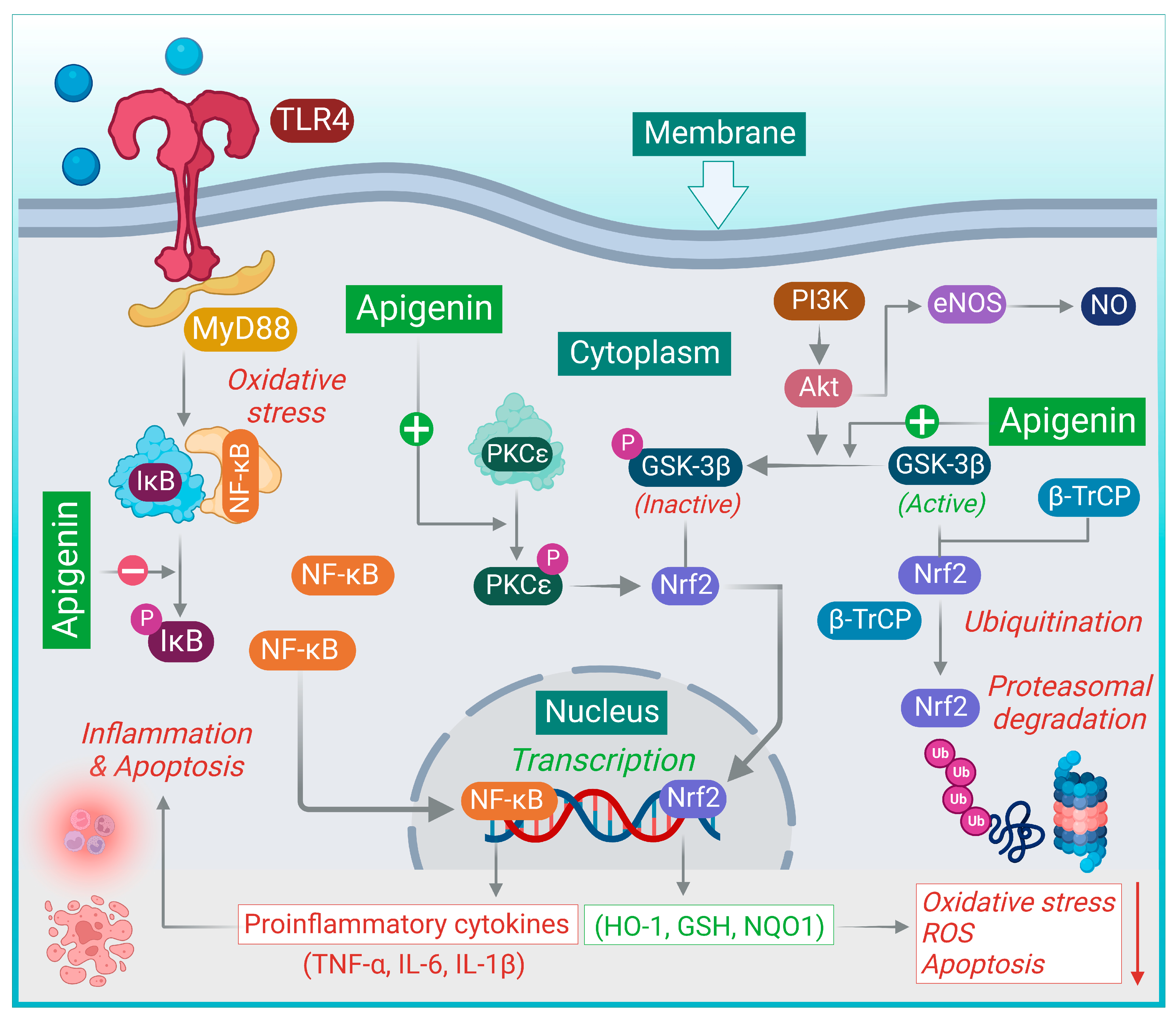{kind=link}
| Source (s) | Glycoside | Quantity (mg/100 g or mg/100 mL) |
|---|---|---|
| Olive oil, extra virgin | Apigenin aglycone | 1.17 |
| Common sage, fresh | Apigenin aglycone | 2.40 |
| Italian oregano, fresh | Apigenin aglycone | 3.50 |
| Rosemary, fresh | Apigenin aglycone | 0.55 |
| Buckwheat, whole grain flour | Apigenin 6-C-glucoside (Isovitexin) | 0.90 |
| Common wheat, whole grain flour | Apigenin 6,8-C-arabinoside-C-glucoside | 30.47 |
| Lentils, whole, raw | Apigenin 7-O-glucoside | 0.18 |
| Globe artichoke, heads, raw | Apigenin 7-O-glucuronide | 7.40 |
| Orange, pure juice | Apigenin 6,8-di-C-glucoside (Vicenin 2) | 5.53 |
| Celery (Seeds) | Apigenin 7-O-apiosyl-glucoside (Apiin) | 111.00 |
| Celery (Leaves) | Apigenin 7-O-apiosyl-glucoside (Apiin) | 8.37 |
| Parsley | Apigenin 7-O-apiosyl-glucoside | 4503.50 (dried) |
| 215.46 (fresh) | ||
| Kumquats | Apigenin 7-O-neohesperidoside | 21.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, S.D.; Jha, N.K.; Jha, S.K.; Sadek, B.; Ojha, S. Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin. Nutrients 2023, 15, 385. https://doi.org/10.3390/nu15020385
Thomas SD, Jha NK, Jha SK, Sadek B, Ojha S. Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin. Nutrients. 2023; 15(2):385. https://doi.org/10.3390/nu15020385
Chicago/Turabian StyleThomas, Shilu Deepa, Niraj Kumar Jha, Saurabh Kumar Jha, Bassem Sadek, and Shreesh Ojha. 2023. "Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin" Nutrients 15, no. 2: 385. https://doi.org/10.3390/nu15020385
APA StyleThomas, S. D., Jha, N. K., Jha, S. K., Sadek, B., & Ojha, S. (2023). Pharmacological and Molecular Insight on the Cardioprotective Role of Apigenin. Nutrients, 15(2), 385. https://doi.org/10.3390/nu15020385