
Maternal Betaine Supplementation Mitigates Maternal High Fat Diet-Induced NAFLD in Offspring Mice through Gut Microbiota


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals Experiment
2.2. Glucose and Insulin Tolerance Tests (GTT/ITT)
2.3. Serum and Liver Biochemical Analysis
2.4. Histopathology Examination
2.5. Quantification of One-Carbon Metabolites in the Serum
2.6. Quantification of SCFAs Content in the Cecal
2.7. 16S rRNA Pyrosequencing Analysis
2.8. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.9. Statistical Analysis
3. Results
3.1. Body Weight, Organ Coefficients and Serum Biochemistry in Dams and Offspring Mice
3.2. Maternal Betaine Intake Attenuated NAFLD in Offspring Mice Induced by MHFD
3.3. Maternal Betaine Intake Altered the Gut Microbiota and Its Related Metabolites in Offspring Mice
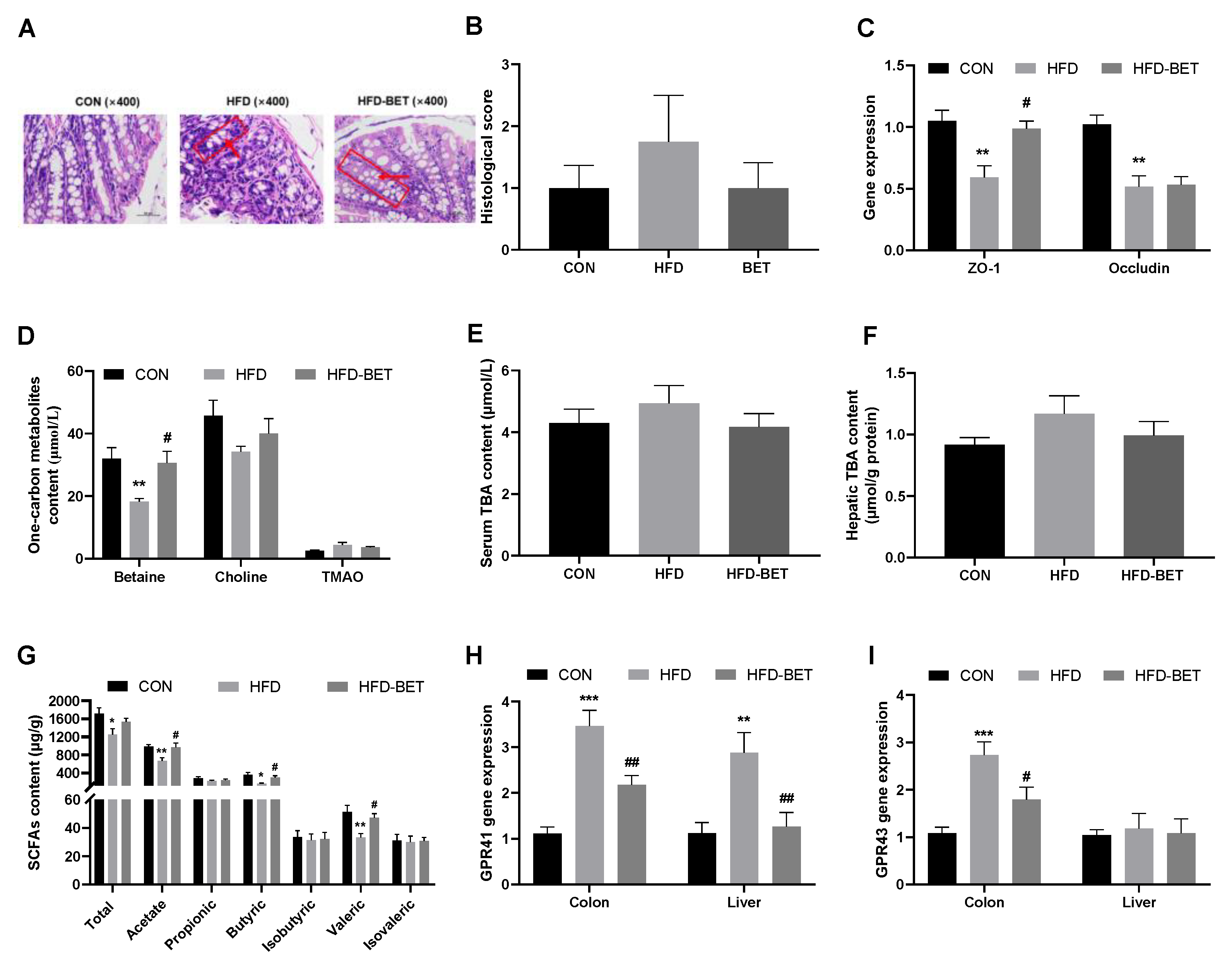
3.3.1. Maternal Betaine Intake Reversed the Disruption of Intestinal Development and Gut Barrier in Offspring Mice Exposed to MHFD
3.3.2. Maternal Betaine Supplementation Reversed Gut Microbiota-Related Metabolites in Offspring Mice Exposed to MHFD
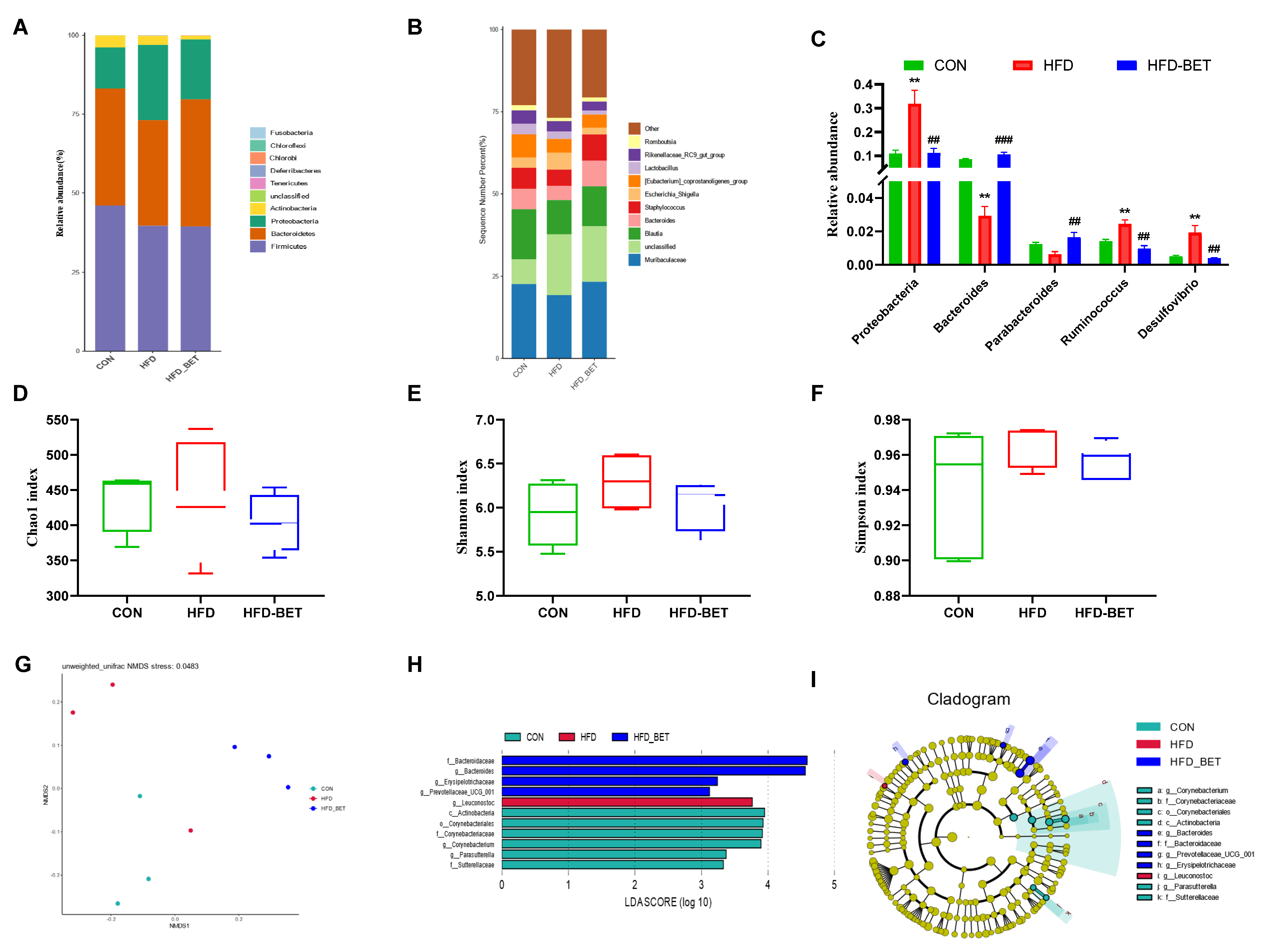
3.3.3. Maternal Betaine Supplementation Altered the Gut Microbiota in Offspring Mice Exposed to MHFD
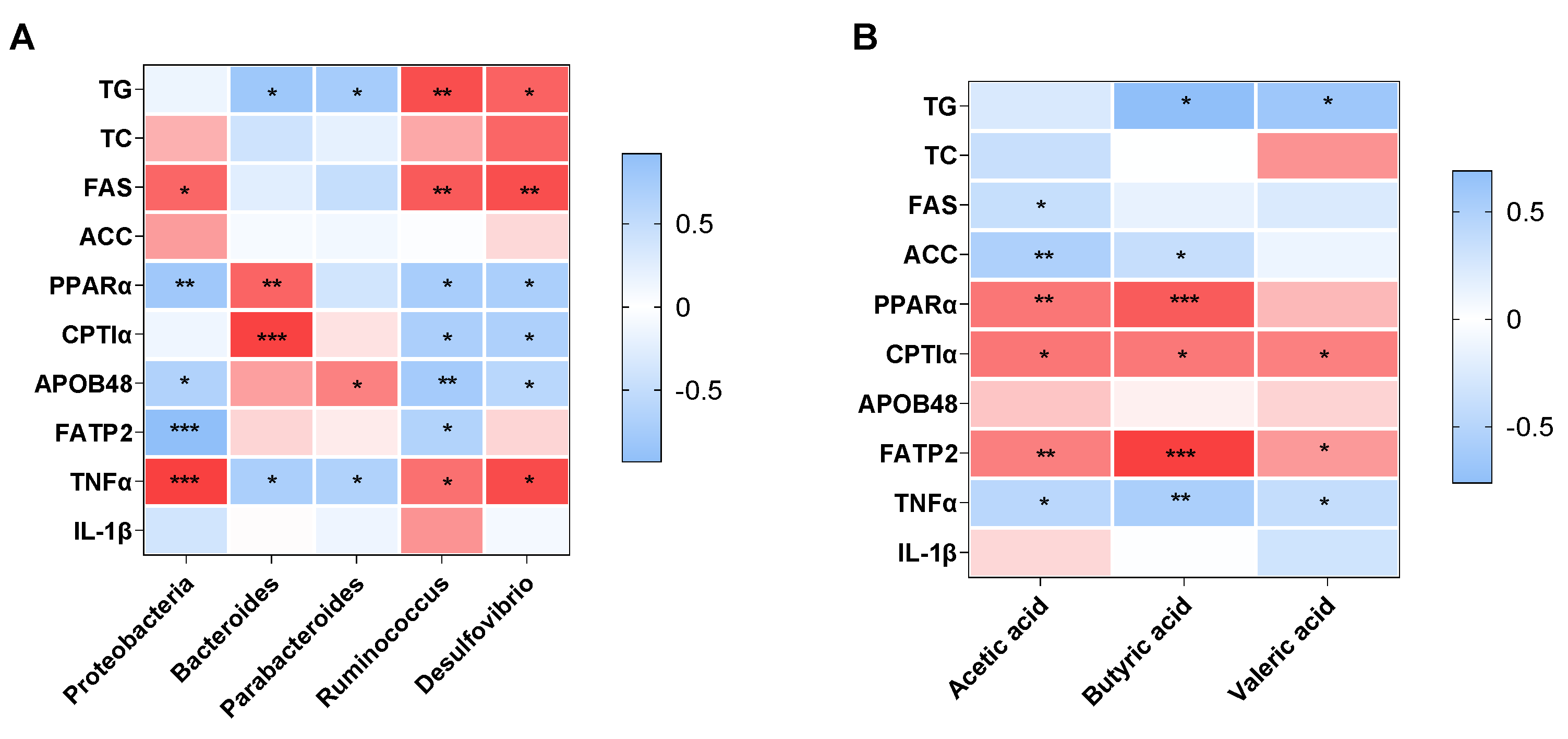
3.3.4. Association of the Altered Gut Microbiota and SCFAs with NAFLD-Related Parameters in Offspring Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Anderson, E.L.; Howe, L.; Jones, H.; Higgins, J.; Lawlor, D.; Fraser, A. The Prevalence of Non-Alcoholic Fatty Liver Disease in Children and Adolescents: A Systematic Review and Meta-Analysis. PLoS ONE 2015, 10, e0140908. [Google Scholar] [CrossRef] [PubMed]
- Younossi, Z.; Tacke, F.; Arrese, M.; Sharma, B.C.; Mostafa, I.; Bugianesi, E.; Wong, V.W.-S.; Yilmaz, Y.; George, J.; Fan, J.; et al. Global Perspectives on Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Hepatology 2019, 69, 2672–2682. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.E. Obesity and Gestational Diabetes Mellitus Pathways for Programming in Mouse, Monkey, and Man—Where Do We Go Next? The 2014 Norbert Freinkel Award Lecture. Diabetes Care 2015, 38, 1402–1411. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, S.R.; Kasmi, K.; Jonscher, K.; Friedman, J. Developmental origins of NAFLD: A womb with a clue. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 81–96. [Google Scholar] [CrossRef]
- Hagström, H.; Simon, T.; Roelstraete, B.; Stephansson, O.; Söderling, J.; Ludvigsson, J. Maternal obesity increases the risk and severity of NAFLD in offspring. J. Hepatol. 2021, 75, 1042–1048. [Google Scholar] [CrossRef]
- Peng, H.; Xu, H.; Wu, J.; Li, J.; Zhou, Y.; Ding, Z.; Siwko, S.; Yuan, X.; Schalinske, K.; Alpini, G.; et al. Maternal high-fat diet disrupted one-carbon metabolism in offspring, contributing to nonalcoholic fatty liver disease. Liver Int. 2021, 41, 1305–1319. [Google Scholar] [CrossRef]
- Li, J.; Huang, J.; Li, J.; Chen, H.; Huang, K.; Zheng, L. Accumulation of endoplasmic reticulum stress and lipogenesis in the liver through generational effects of high fat diets. J. Hepatol. 2012, 56, 900–907. [Google Scholar] [CrossRef]
- Rao, Y.; Kuang, Z.; Li, C.; Guo, S.; Xu, Y.; Zhao, D.; Hu, Y.; Song, B.; Jiang, Z.; Ge, Z.; et al. Gut Akkermansia muciniphila ameliorates metabolic dysfunction-associated fatty liver disease by regulating the metabolism of L-aspartate via gut-liver axis. Gut Microbes 2021, 13, 1–19. [Google Scholar] [CrossRef]
- Tripathi, A.; Debelius, J.; Brenner, D.; Karin, M.; Loomba, R.; Schnabl, B.; Knight, R. The gut-liver axis and the intersection with the microbiome. Nat. Rev. Gastroenterol. Hepatol. 2018, 15, 397–411. [Google Scholar] [CrossRef]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.; Seed, P.; Rawls, J.; David, L.; et al. The severity of nonalcoholic fatty liver disease is associated with gut dysbiosis and shift in the metabolic function of the gut microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef]
- Chen, Y.M.; Liu, Y.; Zhou, R.; Chen, X.; Wang, C.; Tan, X.; Wang, L.; Zheng, R.; Zhang, H.; Ling, W.; et al. Associations of gut-flora-dependent metabolite trimethylamine-N-oxide, betaine and choline with non-alcoholic fatty liver disease in adults. Sci. Rep. 2016, 6, 19076. [Google Scholar] [CrossRef] [PubMed]
- Nimer, N.; Choucair, I.; Wang, Z.; Nemet, I.; Li, L.; Gukasyan, J.; Weeks, T.; Alkhouri, N.; Zein, N.; Tang, W.; et al. Bile acids profile, histopathological indices and genetic variants for non-alcoholic fatty liver disease progression. Metabolism 2021, 116, 154457. [Google Scholar] [CrossRef] [PubMed]
- Sahuri-Arisoylu, M.; Brody, L.; Parkinson, J.; Parkes, H.; Navaratnam, N.; Miller, A.; Thomas, E.; Frost, G.; Bell, J. Reprogramming of hepatic fat accumulation and ‘browning’ of adipose tissue by the short-chain fatty acid acetate. Int. J. Obes. 2016, 40, 955–963. [Google Scholar] [CrossRef] [PubMed]
- Tedelind, S.; Westberg, F.; Kjerrulf, M.; Vidal, A. Anti-inflammatory properties of the short-chain fatty acids acetate and propionate: A study with relevance to inflammatory bowel disease. World J. Gastroenterol. 2007, 13, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Lv, L.; Wu, W.; Li, Y.; Shi, D.; Fang, D.; Guo, F.; Jiang, H.; Yan, R.; Ye, W.; et al. Butyrate Protects Mice Against Methionine-Choline-Deficient Diet-Induced Non-alcoholic Steatohepatitis by Improving Gut Barrier Function, Attenuating Inflammation and Reducing Endotoxin Levels. Front. Microbiol. 2018, 9, 1967. [Google Scholar] [CrossRef]
- Song, L.; Cui, J.; Hu, S.; Wang, R.; Li, H.; Sun, B. Maternal Treatment with Metformin Persistently Ameliorates High-Fat Diet-Induced Metabolic Symptoms and Modulates Gut Microbiota in Rat Offspring. Nutrients 2022, 14, 3612. [Google Scholar] [CrossRef] [PubMed]
- Soderborg, T.K.; Clark, S.; Mulligan, C.; Janssen, R.; Babcock, L.; Ir, D.; Young, B.; Krebs, N.; Lemas, D.; Johnson, L.; et al. The gut microbiota in infants of obese mothers increases inflammation and susceptibility to NAFLD. Nat. Commun. 2018, 9, 4462. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Tan, Y.; Wei, J.; Chang, Y.; Jin, T.; Zhu, H. Betaine supplement alleviates hepatic triglyceride accumulation of apolipoprotein E deficient mice via reducing methylation of peroxisomal proliferator-activated receptor alpha promoter. Lipids Health Dis. 2013, 12, 34. [Google Scholar] [CrossRef]
- Wang, L.J.; Zhang, H.; Zhou, J.; Liu, Y.; Yang, Y.; Chen, X.; Zhu, C.; Zheng, R.; Ling, W.; Zhu, H. Betaine attenuates hepatic steatosis by reducing methylation of the MTTP promoter and elevating genomic methylation in mice fed a high-fat diet. J. Nutr. Biochem. 2014, 25, 329–336. [Google Scholar] [CrossRef]
- Du, J.; Zhang, P.; Luo, J.; Shen, L.; Zhang, S.; Gu, H.; He, J.; Wang, L.; Zhao, X.; Gan, M.; et al. Dietary betaine prevents obesity through gut microbiota-drived microRNA-378a family. Gut Microbes 2021, 13, 1862612. [Google Scholar] [CrossRef]
- van Lee, L.; Tint, M.; Aris, I.; Quah, P.; Fortier, M.; Lee, Y.; Yap, F.; Saw, S.; Godfrey, K.; Gluckman, P.; et al. Prospective associations of maternal betaine status with offspring weight and body composition at birth: The Growing Up in Singapore Towards healthy Outcomes (GUSTO) cohort study. Am. J. Clin. Nutr. 2016, 104, 1327–1333. [Google Scholar] [CrossRef] [PubMed]
- Ribo, S.; Sánchez-Infantes, D.; Martinez-Guino, L.; García-Mantrana, I.; Ramon-Krauel, M.; Tondo, M.; Arning, E.; Nofrarías, M.; Osorio-Conles, Ó.; Fernández-Pérez, A.; et al. Increasing breast milk betaine modulates Akkermansia abundance in mammalian neonates and improves long-term metabolic health. Sci. Transl. Med. 2021, 13, eabb0322. [Google Scholar] [CrossRef]
- Liu, J.; Ding, L.; Zhai, X.; Wang, D.; Xiao, C.; Hui, X.; Sun, T.; Yu, M.; Zhang, Q.; Li, M.; et al. Maternal Dietary Betaine Prevents High-Fat Diet-Induced Metabolic Disorders and Gut Microbiota Alterations in Mouse Dams and Offspring From Young to Adult. Front. Microbiol. 2022, 13, 809642. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Meex, R.; Venema, K.; Blaak, E. Gut microbial metabolites in obesity, NAFLD and T2DM. Nat. Rev. Endocrinol. 2019, 15, 261–273. [Google Scholar] [CrossRef] [PubMed]
- De Minicis, S.; Rychlicki, C.; Agostinelli, L.; Saccomanno, S.; Candelaresi, C.; Trozzi, L.; Mingarelli, E.; Facinelli, B.; Magi, G.; Palmieri, C.; et al. Dysbiosis contributes to fibrogenesis in the course of chronic liver injury in mice. Hepatology 2014, 59, 1738–1749. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.; Brown, M. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Collado, M.C.; Isolauri, E.; Laitinen, K.; Salminen, S. Effect of mother’s weight on infant’s microbiota acquisition, composition, and activity during early infancy: A prospective follow-up study initiated in early pregnancy. Am. J. Clin. Nutr. 2010, 92, 1023–1030. [Google Scholar] [CrossRef]
- Galley, J.D.; Bailey, M.; Dush, C.K.; Schoppe-Sullivan, S.; Christian, L. Maternal obesity is associated with alterations in the gut microbiome in toddlers. PLoS ONE 2014, 9, e113026. [Google Scholar] [CrossRef]
- Mueller, N.T.; Shin, H.; Pizoni, A.; Werlang, I.; Matte, U.; Goldani, M.; Goldani, H.; Dominguez-Bello, M. Birth mode-dependent association between pre-pregnancy maternal weight status and the neonatal intestinal microbiome. Sci. Rep. 2016, 6, 23133. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Z.; Chen, L.; Tang, L.; Wen, S.; Liu, Y.; Yuan, J. Diet induced maternal obesity affects offspring gut microbiota and persists into young adulthood. Food Funct. 2018, 9, 4317–4327. [Google Scholar] [CrossRef]
- Valentini, F.; Rocchi, G.; Vespasiani-Gentilucci, U.; Guarino, M.; Altomare, A.; Carotti, S. The Origins of NAFLD: The Potential Implication of Intrauterine Life and Early Postnatal Period. Cells 2022, 11, 562. [Google Scholar] [CrossRef] [PubMed]
- Joselit, Y.; Nanobashvili, K.; Jack-Roberts, C.; Greenwald, E.; Malysheva, O.; Caudill, M.; Saxena, A.; Jiang, X. Maternal betaine supplementation affects fetal growth and lipid metabolism of high-fat fed mice in a temporal-specific manner. Nutr. Diabetes 2018, 8, 41. [Google Scholar] [CrossRef]
- Zhao, N.; Yang, S.; Jia, Y.; Sun, B.; He, B.; Zhao, R. Maternal betaine supplementation attenuates glucocorticoid-induced hepatic lipid accumulation through epigenetic modification in adult offspring rats. J. Nutr. Biochem. 2018, 54, 105–112. [Google Scholar] [CrossRef]
- Sookoian, S.; Salatino, A.; Castaño, G.; Landa, M.; Fijalkowky, C.; Garaycoechea, M.; Pirola, C. Intrahepatic bacterial metataxonomic signature in non-alcoholic fatty liver disease. Gut 2020, 69, 1483–1491. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Hu, Y.; Ma, H.; Zou, Z.; Xiao, Y.; Yang, Y.; Feng, M.; Li, X.; Ye, X. Rhizoma Coptidis alkaloids alleviate hyperlipidemia in B6 mice by modulating gut microbiota and bile acid pathways. Biochim. Biophys. Acta 2016, 1862, 1696–1709. [Google Scholar] [CrossRef] [PubMed]
- Xiao, S.; Fei, N.; Pang, X.; Shen, J.; Wang, L.; Zhang, B.; Zhang, M.; Zhang, X.; Zhang, C.; Li, M.; et al. A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol. Ecol. 2014, 87, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Coker, O.; Chu, E.; Fu, K.; Lau, H.; Wang, Y.; Chan, A.; Wei, H.; Yang, X.; Sung, J.; et al. Dietary cholesterol drives fatty liver-associated liver cancer by modulating gut microbiota and metabolites. Gut 2021, 70, 761–774. [Google Scholar] [CrossRef] [PubMed]
- Safari, Z.; Gérard, P. The links between the gut microbiome and non-alcoholic fatty liver disease (NAFLD). Cell Mol. Life Sci. 2019, 76, 1541–1558. [Google Scholar] [CrossRef]
- Verdam, F.J.; Fuentes, S.; de Jonge, C.; Zoetendal, E.; Erbil, R.; Greve, J.; Buurman, W.; de Vos, W.; Rensen, S. Human intestinal microbiota composition is associated with local and systemic inflammation in obesity. Obes. Silver Spring 2013, 21, E607–E615. [Google Scholar] [CrossRef]
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandonà, A.; Paci, P.; Capuani, G.; et al. Gut microbiota profiling of pediatric nonalcoholic fatty liver disease and obese patients unveiled by an integrated meta-omics-based approach. Hepatology 2017, 65, 451–464. [Google Scholar] [CrossRef]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e5. [Google Scholar] [CrossRef]
- Yu, J.S.; Youn, G.; Choi, J.; Kim, C.; Kim, B.; Yang, S.; Lee, J.; Park, T.; Kim, B.; Kim, Y.; et al. Lactobacillus lactis and Pediococcus pentosaceus-driven reprogramming of gut microbiome and metabolome ameliorates the progression of non-alcoholic fatty liver disease. Clin. Transl. Med. 2021, 11, e634. [Google Scholar] [CrossRef] [PubMed]
- Kondo, T.; Kishi, M.; Fushimi, T.; Kaga, T. Acetic acid upregulates the expression of genes for fatty acid oxidation enzymes in liver to suppress body fat accumulation. J. Agric. Food Chem. 2009, 57, 5982–5986. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Wang, J.; Wang, F.; Strappe, P.; Liu, W.; Zheng, J.; Zhou, Z.; Zhang, Y. Microbiota fermentation characteristics of acylated starches and the regulation mechanism of short-chain fatty acids on hepatic steatosis. Food Funct. 2021, 12, 8659–8668. [Google Scholar] [CrossRef] [PubMed]
- Cobbina, E.; Akhlaghi, F. Non-alcoholic fatty liver disease (NAFLD)—Pathogenesis, classification, and effect on drug metabolizing enzymes and transporters. Drug Metab. Rev. 2017, 49, 197–211. [Google Scholar] [CrossRef]
- Cobbina, E.; Akhlaghi, F. SCFAs alleviated steatosis and inflammation in mice with NASH induced by MCD. J. Endocrinol. 2020, 245, 425–437. [Google Scholar]
- Zhou, D.; Pan, Q.; Xin, F.; Zhang, R.; He, C.; Chen, G.; Liu, C.; Chen, Y.; Fan, J. Sodium butyrate attenuates high-fat diet-induced steatohepatitis in mice by improving gut microbiota and gastrointestinal barrier. World J. Gastroenterol. 2017, 23, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Vigliotti, C.; Witjes, J.; Le, P.; Holleboom, A.; Verheij, J.; Nieuwdorp, M.; Clément, K. Gut microbiota and human NAFLD: Disentangling microbial signatures from metabolic disorders. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 279–297. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| FAS | GCTGCGGAAACTTCAGGAAAT | AGAGACGTGTCACTCCTGGACTT |
| ACC | CACTGTGAACATGTGGAGG | AGGCTGATGGTGATGACC |
| PPARα | GGGCAAGAGAATCCACGAAG | GTTGTTGCTGGTCTTTCCCG |
| CPT1α | CGCACGGAAGGAAAATGG | TGTGCCCAATATTCCTGG |
| APOB48 | GCATGAGTATGCCAATGGTCTCC | CTGGTTGCCATCTGAAGCCATG |
| FATP2 | TTCAACAGCGGAGACCTCCTGA | CCACGATGTCAGCGACTTCTGT |
| TNFα | GGTGCCTATGTCTCAGCCTCTT | GCCATAGAACTGATGAGAGGGAG |
| IL-1β | TTCAGGCAGGCAGTATCACTC | GAAGGTCCACGGGAAAGACAC |
| ZO-1 | ATCCCACAAGGAGCCATTCC | TAGGGTCACAGTGTGGCAAG |
| Occludin | TTTGCTGTGAAAACCCGAAGA | ACTGTCAACTCTTTCCGCATA |
| GPR41 | TTGCTAAACCTGACCATTTCGG | GATAGGCCACGCTCAGAAAAC |
| GPR43 | ACAGTGGAGGGGACCAAGAT | GGGGACTCTCTACTCGGTGA |
| β-actin | GGGTCAGAAGGACTCCTATG | GTAACAATGCCATGTTCAAT |
| Parameters | CON | HFD | HFD-BET |
|---|---|---|---|
| Liver weight (g) | 1.08 ± 0.06 | 1.31 ± 0.03 * | 1.14 ± 0.03 # |
| Liver/body weight ratio (%) | 6.24 ± 0.13 | 6.64 ± 0.09 * | 5.97 ± 0.11 ### |
| Visceral fat weight (g) | 0.11 ± 0.02 | 0.23 ± 0.02 *** | 0.17 ± 0.02 # |
| Visceral fat /body weight ratio (%) | 0.59 ± 0.08 | 1.14 ± 0.07 *** | 0.85 ± 0.08 # |
| ALT (U/L) | 29.25 ± 2.63 | 42.75 ± 4.55 * | 33.71 ± 1.21 |
| AST (U/L) | 139.25 ± 5.92 | 210.00 ± 18.20 * | 147.43 ± 11.49 # |
| HDL-C (mmol/L) | 1.91 ± 0.21 | 2.10 ± 0.06 | 1.79 ± 0.09 |
| LDL-C (mmol/L) | 0.36 ± 0.02 | 0.39 ± 0.02 | 0.49 ± 0.03 |
| TC (mmol/L) | 1.63 ± 0.07 | 2.55 ± 0.08 ** | 2.18 ± 0.15 |
| TG (mmol/L) | 0.25 ± 0.02 | 0.34 ± 0.02 * | 0.38 ± 0.02 |
| GLU (mmol/L) | 8.15 ± 0.96 | 12.21 ± 2.09 | 10.30 ± 2.84 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, L.; Tan, X.; Liang, X.; Chen, H.; Ou, Q.; Wu, Q.; Yu, X.; Zhao, H.; Huang, Q.; Yi, Z.; et al. Maternal Betaine Supplementation Mitigates Maternal High Fat Diet-Induced NAFLD in Offspring Mice through Gut Microbiota. Nutrients 2023, 15, 284. https://doi.org/10.3390/nu15020284
Sun L, Tan X, Liang X, Chen H, Ou Q, Wu Q, Yu X, Zhao H, Huang Q, Yi Z, et al. Maternal Betaine Supplementation Mitigates Maternal High Fat Diet-Induced NAFLD in Offspring Mice through Gut Microbiota. Nutrients. 2023; 15(2):284. https://doi.org/10.3390/nu15020284
Chicago/Turabian StyleSun, Liuqiao, Xuying Tan, Xiaoping Liang, Hangjun Chen, Qian Ou, Qiongmei Wu, Xinxue Yu, Hanqing Zhao, Qiaoli Huang, Zehua Yi, and et al. 2023. "Maternal Betaine Supplementation Mitigates Maternal High Fat Diet-Induced NAFLD in Offspring Mice through Gut Microbiota" Nutrients 15, no. 2: 284. https://doi.org/10.3390/nu15020284
APA StyleSun, L., Tan, X., Liang, X., Chen, H., Ou, Q., Wu, Q., Yu, X., Zhao, H., Huang, Q., Yi, Z., Wei, J., Wu, F., Zhu, H., & Wang, L. (2023). Maternal Betaine Supplementation Mitigates Maternal High Fat Diet-Induced NAFLD in Offspring Mice through Gut Microbiota. Nutrients, 15(2), 284. https://doi.org/10.3390/nu15020284