
The Different Ways Multi-Strain Probiotics with Different Ratios of Bifidobacterium and Lactobacillus Relieve Constipation Induced by Loperamide in Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Probiotics
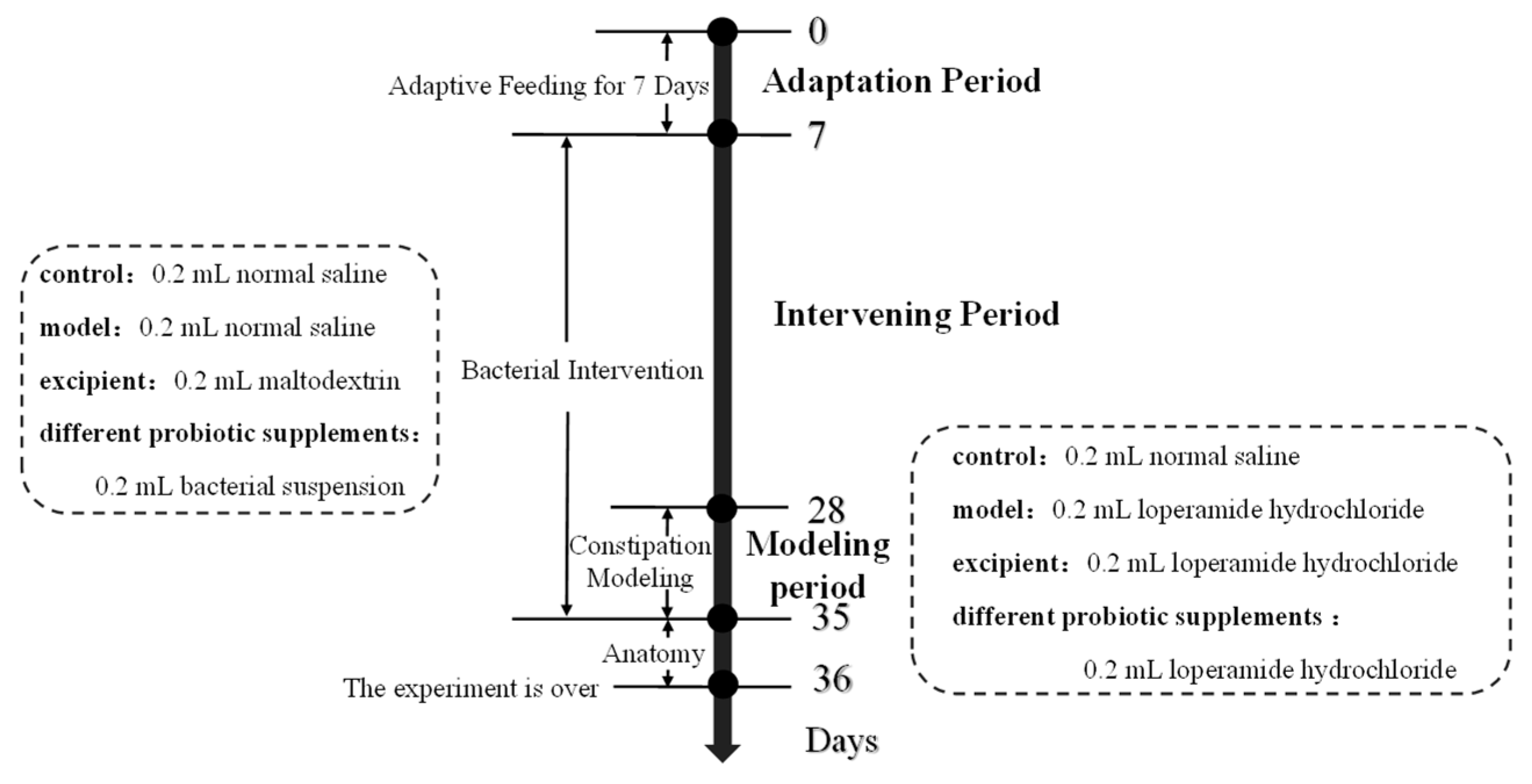
2.3. Animals and Experimental Design
2.4. The Collection of Mouse Tissues and Serum
2.5. Experiment on Regulating the Function of Intestinal Flora
2.6. Gastrointestinal Indices
2.6.1. Faecal Water Content
2.6.2. The Time of the First Black Stool Defecation
2.6.3. Small Intestine Propulsion Rate
2.7. Biochemical Analyses
2.8. The Determination of SCFA (Short-Chain Fatty Acid) Contents in Faeces
2.9. 16S rDNA Sequencing and Bioinformatics Analysis
2.10. Statistical Analyses
3. Results
3.1. The Effect of Multi-Strain Probiotic Intervention on the Body Weight of Constipated Mice Was Not Significant
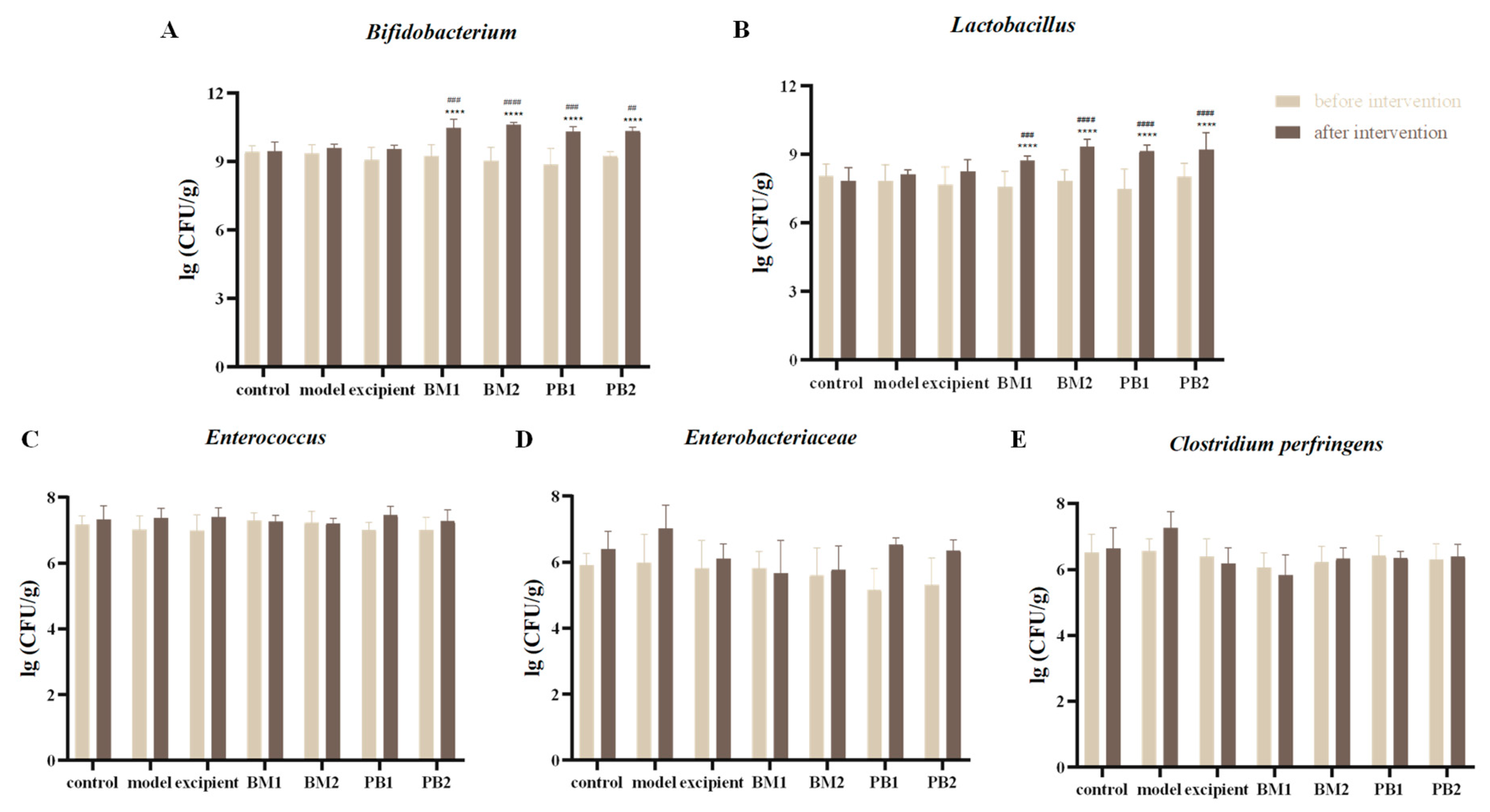
3.2. Four Multi-Strain Probiotics Increased the Number of Beneficial Bacteria in the Intestines of Constipated Mice and Regulated Their Intestinal Flora
3.3. Three Multi-Strain Probiotics Relieve Constipation
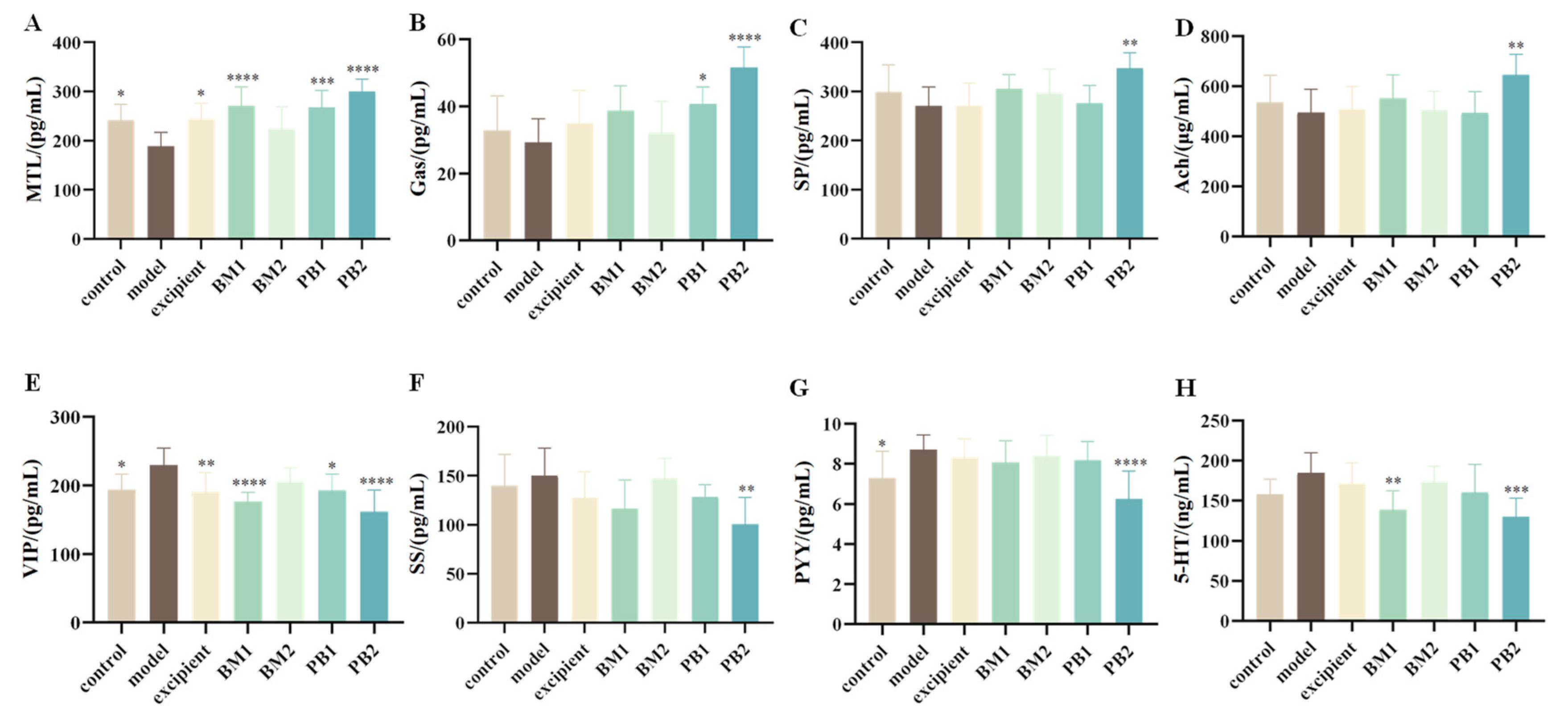
3.4. Different Multi-Strain Probiotics Modulated the Levels of Neurotransmitters and Inflammatory Factors in Mice Serum to Different Degrees
3.4.1. The Effects of Different Probiotic Supplements on Serum Neurotransmitters in Mice
3.4.2. The Effect of Different Probiotic Supplements on Serum Inflammatory Factors in Mice
3.5. Multi-Strain Probiotics Significantly Upregulated the Content of SCFAs in the Faeces of Constipated Mice
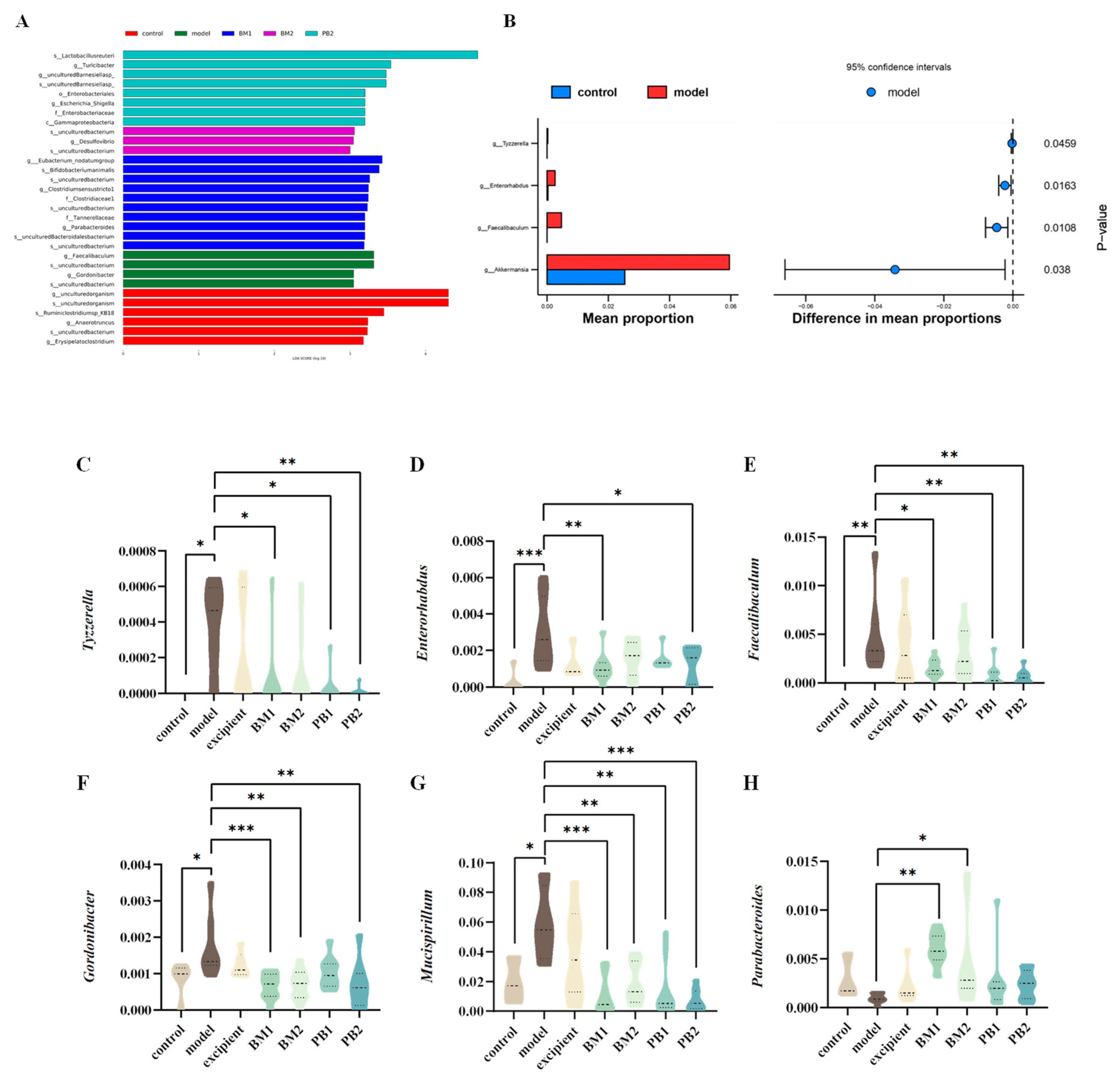
3.6. Different Multi-Strain Probiotics Improve Constipation Symptoms by Changing the Compositional Structure of Intestinal Flora
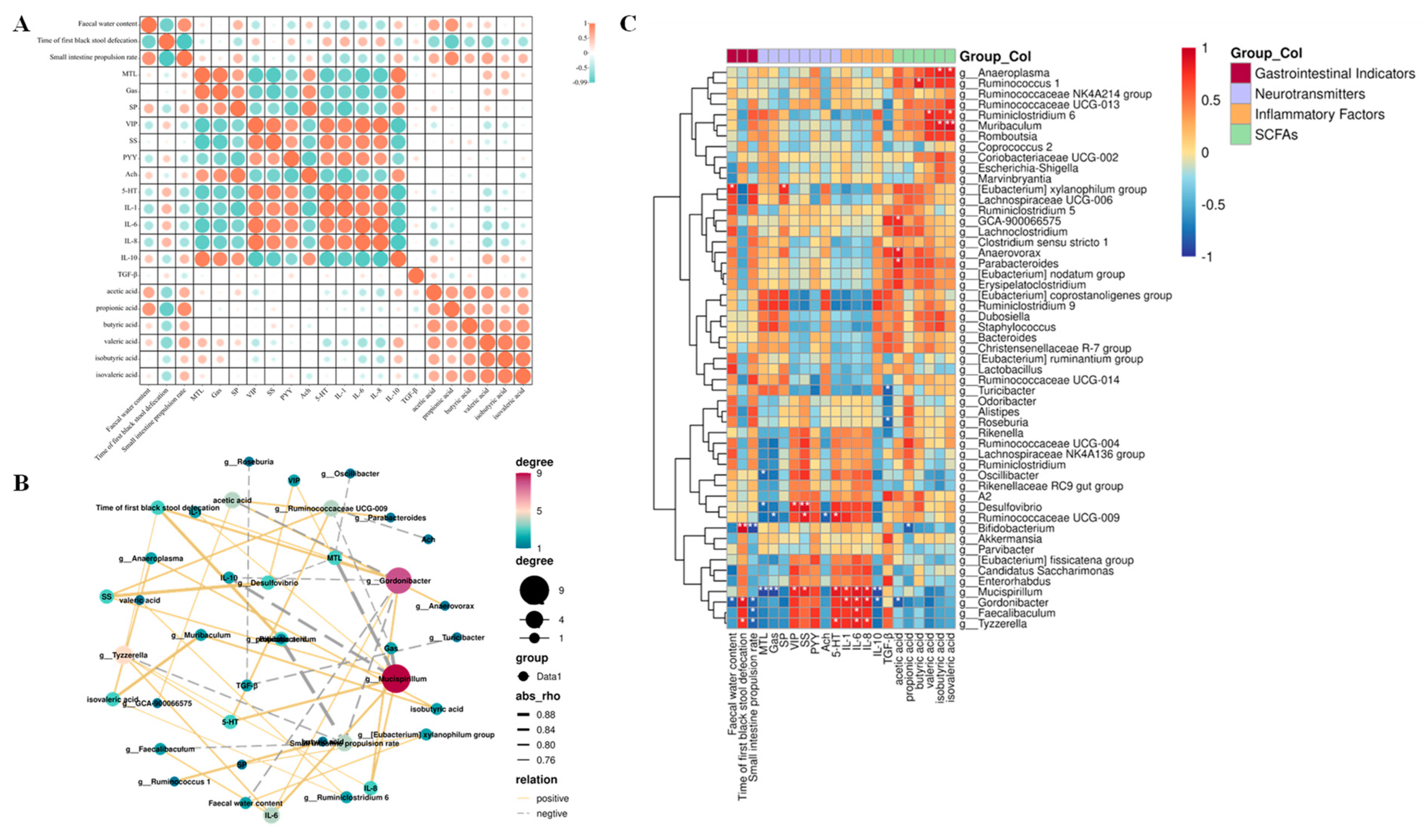
3.7. Correlation Analysis Showed That Constipation Relief by Different Multi-Strain Probiotics Was Associated with Changes in Intestinal Flora and SCFAs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohkusa, T.; Koido, S.; Nishikawa, Y.; Sato, N. Gut Microbiota and Chronic Constipation: A Review and Update. Front. Med. 2019, 6, 19. [Google Scholar] [CrossRef] [PubMed]
- Jicheng, W.; Xiaoye, B.; Chuantao, P.; Zhongjie, Y.; Bohai, L.; Wenyi, Z.; Zhihong, S.; Heping, Z. Fermented milk containing Lactobacillus casei Zhang and Bifidobacterium animalis ssp. lactis V9 alleviated constipation symptoms through regulation of intestinal microbiota, inflammation, and metabolic pathways-ScienceDirect. J. Dairy Sci. 2020, 103, 11025–11038. [Google Scholar] [CrossRef]
- Mostafa, S.M.; Bhandari, S.; Ritchie, G.; Gratton, N.; Wenstone, R. Constipation and its implications in the critically ill patient. Br. J. Anaesth. 2003, 91, 815–819. [Google Scholar] [CrossRef]
- Ghoshal, U.C. Chronic constipation in Rome IV era: The Indian perspective. Indian J. Gastroenterol. 2017, 36, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Linsheng, H.; Qi, Z.; Xiao, Q.; Huanlong, Q. Microbial treatment in chronic constipation. Sci. China (Life Sci.) 2018, 61, 744–752. [Google Scholar] [CrossRef]
- Johanson, J.F.; Kralstein, J. Chronic constipation: A survey of the patient perspective. Aliment. Pharmacol. Ther. 2007, 25, 599–608. [Google Scholar] [CrossRef]
- Omer, A.; Quigley, E.M.M. An update on prucalopride in the treatment of chronic constipation. Ther. Adv. Gastroenterol. 2017, 10, 877–887. [Google Scholar] [CrossRef] [PubMed]
- Chey, W.D.; Drossman, D.A.; Johanson, J.F.; Scott, C.; Panas, R.M.; Ueno, R. Safety and patient outcomes with lubiprostone for up to 52 weeks in patients with irritable bowel syndrome with constipation. Aliment. Pharmacol. Ther. 2012, 35, 587–599. [Google Scholar] [CrossRef]
- Araujo, M.M.; Botelho, P.B. Probiotics, prebiotics, and synbiotics in chronic constipation: Outstanding aspects to be considered for the current evidence. Front. Nutr. 2022, 9, 935830. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Dimidi, E.; Christodoulides, S.; Mark, S.S.; Whelan, K. Mechanisms of Action of Probiotics and the Gastrointestinal Microbiota on Gut Motility and Constipation. Adv. Nutr. 2017, 8, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, A.; Szajewska, H. Systematic review of randomised controlled trials: Probiotics for functional constipation. World J. Gastroenterol. 2010, 16, 69. [Google Scholar] [CrossRef] [PubMed]
- Dharmani, P.; Srivastava, V.; Kissoon-Singh, V.; Chadee, K. Role of intestinal mucins in innate host defense mechanisms against pathogens. J. Innate Immun. 2009, 1, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Bunnett, N.W. Neuro-humoral signalling by bile acids and the TGR5 receptor in the gastrointestinal tract. J. Physiol. 2014, 592, 2943–2950. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wang, L.; Dahiya, S.; Beier, U.H.; Han, R.; Samanta, A.; Bergman, J.; Sotomayor, E.M.; Seto, E.; Kozikowski, A.P. Histone/protein deacetylase 11 targeting promotes Foxp3+ Treg function. Sci. Rep. 2017, 7, 8626. [Google Scholar] [CrossRef]
- Postler, T.S.; Ghosh, S. Understanding the Holobiont: How Microbial Metabolites Affect Human Health and Shape the Immune System. Cell Metab. 2017, 25, 110–130. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Kitazawa, H.; Iwabuchi, N.; Xiao, J.Z.; Miyaji, K.; Iwatsuki, K.; Saito, T. Immunostimulatory oligodeoxynucleotide from Bifidobacterium longum suppresses Th2 immune responses in a murine model. Clin. Exp. Immunol. 2006, 145, 130–138. [Google Scholar] [CrossRef]
- Dimidi, E.; Scott, S.M.; Whelan, K. Probiotics and constipation: Mechanisms of action, evidence for effectiveness and utilisation by patients and healthcare professionals. Proc. Nutr. Soc. 2020, 79, 147–157. [Google Scholar] [CrossRef]
- Chengcheng, Z.; Jinchi, J.; Fengwei, T.; Jianxin, Z.; Hao, Z.; Qixiao, Z.; Wei, C. Meta-analysis of randomized controlled trials of the effects of probiotics on functional constipation in adults. Clin. Nutr. 2020, 39, 2960–2969. [Google Scholar] [CrossRef]
- MacPherson, C.W.; Shastri, P.; Mathieu, O.; Tompkins, T.A.; Burguiere, P. Genome-Wide Immune Modulation of TLR3-Mediated Inflammation in Intestinal Epithelial Cells Differs between Single and Multi-Strain Probiotic Combination. PLoS ONE 2017, 12, e0169847. [Google Scholar] [CrossRef]
- Jeong, J.-J.; Ganesan, R.; Jin, Y.-J.; Park, H.J.; Min, B.H.; Jeong, M.K.; Yoon, S.J.; Choi, M.R.; Choi, J.; Moon, J.H.; et al. Multi-strain probiotics alleviate loperamide-induced constipation by adjusting the microbiome, serotonin, and short-chain fatty acids in rats. Front. Microbiol. 2023, 14, 1174968. [Google Scholar] [CrossRef] [PubMed]
- Yerlett, N.; Petrof, G.; Bageta, M.; Balboa, P.L.; Martinez, A.E. The impact of using a multistrain probiotic supplement on gastrointestinal function in children and adolescents with severe recessive dystrophic epidermolysis bullosa. Clin. Exp. Dermatol. 2023, 48, 188–192. [Google Scholar] [CrossRef]
- Cheng, S.; Li, H.; Ding, Y.; Huo, J.; Zheng, Y.; Jiang, Y.; Zhang, Y.; Man, C. The Probiotic Combination of Lacticaseibacillus paracasei JY062 and Lactobacillus gasseri JM1 Alleviates Gastrointestinal Motility Disorder via Improving Gut Microbiota. Nutrients 2023, 15, 839. [Google Scholar] [CrossRef] [PubMed]
- Ayob, N.; Nawawi, K.N.M.; Nor, M.H.M.; Ali, R.A.R.; Ahmad, H.F.; Oon, S.F.; Mokhtar, N.M. The Effects of Probiotics on Small Intestinal Microbiota Composition, Inflammatory Cytokines and Intestinal Permeability in Patients with Non-Alcoholic Fatty Liver Disease. Biomedicines 2023, 11, 640. [Google Scholar] [CrossRef]
- Tian, P.; Zou, R.; Wang, L.; Chen, Y.; Qian, X.; Zhao, J.; Zhang, H.; Qian, L.; Wang, Q.; Wang, G.; et al. Multi-Probiotics ameliorate Major depressive disorder and accompanying gastrointestinal syndromes via serotonergic system regulation. J. Adv. Res. 2023, 45, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.-Y.; Lin, J.-H.; Kuo, Y.-W.; Chiang, P.-F.R.; Ho, H.-H. Probiotics and their Metabolites Reduce Oxidative Stress in Middle-Aged Mice. Curr. Microbiol. 2022, 79, 104. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, W.; Gan, D. Alleviation of slow transit constipation by probiotics complex. Food Ferment. Ind. 2022, 48, 95–100. [Google Scholar] [CrossRef]
- Lin, W.; Kuo, Y.; Chen, C.; Hsu, Y.; Huang, Y.; Hsu, C.; Lin, J.; Lin, C.; Lin, C.; Yi, T.; et al. The Function of Mixed Postbiotic PE0401 in Improving Intestinal Health via Elevating Anti-inflammation, Anti-oxidation, Epithelial Tight Junction Gene Expression and Promoting Beneficial Bacteria Growth. J. Pure Appl. Microbiol. 2022, 16, 1771–1782. [Google Scholar] [CrossRef]
- Bo, Z.; Lynch, B.; Jian, Z.; Yu, G.; Mak, A. Lactobacillus rhamnosus MP108: Toxicological evaluation. J. Food Sci. 2021, 86, 228–241. [Google Scholar] [CrossRef]
- Ouyang, B.; Cui, S.; Mao, B.; Tang, X.; Ma, F.; Zhao, J.; Zhang, H.; Chen, W. Preparation of the acid-resistant freeze-dried Lactobacillus casei CCFM711. Food Ferment. Ind. 2020, 46, 62–68. Available online: https://www.webofscience.com/wos/alldb/full-record/CSCD:6768516 (accessed on 28 September 2023).
- Chai, M.; Wang, L.; Li, X.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Different Bifidobacterium bifidum strains change the intestinal flora composition of mice via different mechanisms to alleviate loperamide-induced constipation. Food Funct. 2021, 12, 6058–6069. [Google Scholar] [CrossRef]
- Li, Y.; Shen, M.; Wang, P. Study on the effects and mechanism of Rehmanniae Radix water extract on sleep in rats. Nat. Prod. Res. Dev. 2022, 34, 1911–1918+1977. [Google Scholar] [CrossRef]
- Wang, L.; Yang, S.; Wang, J.; Wang, G.; Zhang, H.; Zhao, J.; Chen, W. Lactobacillus paracasei LPC-F relieved constipation by promoting the proliferation of interstitial cells of Cajal. Food Ferment. Ind. 2022, 48, 1–9. [Google Scholar] [CrossRef]
- Wang, G.; Yang, S.; Sun, S.; Si, Q.; Wang, L.; Zhang, Q.; Wu, G.; Zhao, J.; Zhang, H.; Chen, W. Lactobacillus rhamnosusStrains Relieve Loperamide-Induced Constipation via Different Pathways Independent of Short-Chain Fatty Acids. Front. Cell. Infect. Microbiol. 2020, 10, 423. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, L.; Xu, Q.; Jiang, T.; Fang, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Bifidobacteria exert species-specific effects on constipation in BALB/c mice. Food Funct. 2017, 8, 3587–3600. [Google Scholar] [CrossRef]
- Wang, L.; Chen, C.; Cui, S.; Lee, Y.-k.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Adhesive Bifidobacterium Induced Changes in Cecal Microbiome Alleviated Constipation in Mice. Front. Microbiol. 2019, 10, 1721. [Google Scholar] [CrossRef]
- Vandenplas, Y.; Benninga, M. Probiotics and Functional Gastrointestinal Disorders in Children. J. Pediatr. Gastroenterol. Nutr. 2009, 48, S107–S109. [Google Scholar] [CrossRef] [PubMed]
- Flach, J.; Koks, M.; van der Waal, M.B.; Claassen, E.; Larsen, O.F.A. Economic potential of probiotic supplementation in institutionalized elderly with chronic constipation. Pharmanutrition 2018, 6, 198–206. [Google Scholar] [CrossRef]
- Rondanelli, M.; Giacosa, A.; Faliva, M.A.; Perna, S.; Allieri, F.; Castellazzi, A.M. Review on microbiota and effectiveness of probiotics use in older. World J. Clin. Cases 2015, 3, 156–162. [Google Scholar] [CrossRef]
- Isabel Martinez-Martinez, M.; Calabuig-Tolsa, R.; Cauli, O. The effect of probiotics as a treatment for constipation in elderly people: A systematic review. Arch. Gerontol. Geriatr. 2017, 71, 142–149. [Google Scholar] [CrossRef]
- Makizaki, Y.; Uemoto, T.; Yokota, H.; Yamamoto, M.; Tanaka, Y.; Ohno, H. Improvement of loperamide-induced slow transit constipation by Bifidobacterium bifidum G9-1 is mediated by the correction of butyrate production and neurotransmitter profile due to improvement in dysbiosis. PLoS ONE 2021, 16, e0248584. [Google Scholar] [CrossRef]
- Chuan, L.; Shao-Ping, N.; Ke-Xue, Z.; Tao, X.; Chang, L.; Joshua, G.; Ming-Yong, X. Effect of Lactobacillus plantarum NCU116 on loperamide-induced constipation in mice. Int. J. Food Sci. Nutr. 2015, 66, 533–538. [Google Scholar] [CrossRef]
- Yue, X.; Jianxin, Z.; Hao, Z.; Qixiao, Z.; Wei, C. Mining Lactobacillus and Bifidobacterium for organisms with long-term gut colonization potential. Clin. Nutr. 2020, 39, 1315–1323. [Google Scholar] [CrossRef]
- Waitzberg, D.L.; Logullo, L.C.; Bittencourt, A.E.; Torrinhas, R.S.; Shiroma, G.M.; Paulino, N.P.; Teixeira-da-Silva, M.L. Effect of synbiotic in constipated adult women-A randomized, double-blind, placebo-controlled study of clinical response. Clin. Nutr. 2013, 32, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Larsen, C.N.; Nielsen, S.; Kaestel, P.; Brockmann, E.; Bennedsen, M.; Christensen, H.R.; Eskesen, D.C.; Jacobsen, B.L.; Michaelsen, K.F. Dose-response study of probiotic bacteria Bifidobacterium animalis subsp lactis BB-12 and Lactobacillus paracasei subsp paracasei CRL-341 in healthy young adults. Eur. J. Clin. Nutr. 2006, 60, 1284–1293. [Google Scholar] [CrossRef] [PubMed]
- Chengcheng, Z.; Yan, Z.; Jinchi, J.; Leilei, Y.; Fengwei, T.; Jianxin, Z.; Hao, Z.; Wei, C.; Qixiao, Z. Identification of the key characteristics of Bifidobacterium longum strains for the alleviation of ulcerative colitis. Food Funct. 2021, 12, 3476–3492. [Google Scholar] [CrossRef]
- Setchell, K.D.R.; Brown, N.M.; Zimmer-Nechemias, L.; Brashear, W.T.; Wolfe, B.E.; Kirschner, A.S.; Heubi, J.E. Evidence for lack of absorption of soy isoflavone glycosides in humans, supporting the crucial role of intestinal metabolism for bioavailability. Am. J. Clin. Nutr. 2002, 76, 447–453. [Google Scholar] [CrossRef]
- King, S.K.; Sutcliffe, J.R.; Ong, S.Y.; Lee, M.; Koh, T.L.; Wong, S.Q.; Farmer, P.J.; Peck, C.J.; Stanton, M.P.; Keck, J.; et al. Substance P and vasoactive intestinal peptide are reduced in right transverse colon in pediatric slow-transit constipation. Neurogastroenterol. Motil. 2010, 22, 883-e234. [Google Scholar] [CrossRef]
- El-Salhy, M.; Mazzawi, T.; Gundersen, D.; Hatlebakk, J.G.; Hausken, T. The role of peptide YY in gastrointestinal diseases and disorders (Review). Int. J. Mol. Med. 2013, 31, 275–282. [Google Scholar] [CrossRef]
- Shurong, Y.; Huiyue, Z.; Xiaobing, W.; Shanshan, S.; Qian, S.; Qiuxiang, Z.; Linlin, W.; Gang, W.; Jianxin, Z.; Hao, Z.; et al. Analysis on the different effects of Lactobacillus paracasei in relieving constipation induced by loperamide in mice. Food Ferment. Ind. 2020, 46, 25–31. [Google Scholar] [CrossRef]
- Bo, Q.; Lian, Z.; Shuobo, Z.; Shengyi, H.; Yiqiu, F.; Furong, B.; Bjorn, B.; Lanjuan, L.; Mingfei, Y. Prevention of Loperamide-Induced Constipation in Mice and Alteration of 5-Hydroxytryotamine Signaling by Ligilactobacillus salivarius Li01. Nutrients 2022, 14, 4083. [Google Scholar] [CrossRef]
- Liu, W.; Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. The effects of Bacillus coagulans B. C-39 compound microecological preparation against constipation in mice. Food Ferment. Ind. 2019, 45, 85–91. Available online: https://www.webofscience.com/wos/alldb/full-record/CSCD:6539898 (accessed on 1 March 2022).
- Wang, L.; Gourcerol, G.; Yuan, P.-Q.; Wu, S.V.; Million, M.; Larauche, M.; Tache, Y. Peripheral peptide YY inhibits propulsive colonic motor function through Y-2 receptor in conscious mice. Am. J. Physiol.-Gastrointest. Liver Physiol. 2010, 298, G45–G56. [Google Scholar] [CrossRef] [PubMed]
- Bihui, L.; Xin, Y.; Liujin, G.; Jing, Z.; Xianrong, Z.; Ruokun, Y.; Xin, Z. Inhibitory effect of Lactobacillus plantarum CQPC02 isolated from chinese sichuan pickles (Paocai) on constipation in mice. J. Food Qual. 2019, 2019, 9781914. [Google Scholar] [CrossRef]
- Kellow, J.E.; Delvaux, M.; Azpiroz, F.; Camilleri, M.; Quigley, E.M.; Thompson, D.G. Principles of applied neurogastroenterology: Physiology/motility-sensation. Gut 2019, 45 (Suppl. S2), II17–II24. Available online: https://webofscience.clarivate.cn/wos/alldb/full-record/MEDLINE:10457040 (accessed on 12 April 2023).
- Cirali, C.; Ulusoy, E.; Kume, T.; Arslan, N. Elevated serum neopterin levels in children with functional constipation: Association with systemic proinflammatory cytokines. World J. Pediatr. 2018, 14, 448–453. [Google Scholar] [CrossRef]
- Mokhtare, M.; Alimoradzadeh, R.; Agah, S.; Mirmiranpour, H.; Khodabandehloo, N. The Association between Modulating Inflammatory Cytokines and Constipation of Geriatrics in Iran. Middle East J. Dig. Dis. 2017, 9, 228–234. [Google Scholar] [CrossRef]
- Shadnoush, M.; Hosseini, R.S.; Mehrabi, Y.; Delpisheh, A.; Alipoor, E.; Faghfoori, Z.; Mohammadpour, N.; Moghadam, J.Z. Probiotic yogurt Affects Pro- and Anti-inflammatory Factors in Patients with Inflammatory Bowel Disease. Iran. J. Pharm. Res. 2013, 12, 929–936. Available online: https://www.webofscience.com/wos/alldb/full-record/WOS:000336728300038 (accessed on 23 April 2022). [PubMed]
- Xiang, N.; Zhao, J.; Chang, S.; Li, S.; Liu, S.; Wang, C. In Vitro Fecal Fermentation of Euphorbia humifusa-Derived Polysaccharides and Their Protective Effect against Ulcerative Colitis in Mice. Foods 2023, 12, 751. [Google Scholar] [CrossRef]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Gomes Neto, J.C.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle and Horizontal Gene Transfer-Mediated Evolution of Mucispirillum schaedleri, a Core Member of the Murine Gut Microbiota. Msystems 2017, 2, e00171-16. [Google Scholar] [CrossRef]
- Pei, L.-Y.; Ke, Y.-S.; Zhao, H.-H.; Wang, L.; Jia, C.; Liu, W.-Z.; Fu, Q.-H.; Shi, M.-N.; Cui, J.; Li, S.-C. Role of colonic microbiota in the pathogenesis of ulcerative colitis. BMC Gastroenterol. 2019, 19, 10. [Google Scholar] [CrossRef]
- Lecomte, V.; Kaakoush, N.O.; Maloney, C.A.; Raipuria, M.; Huinao, K.D.; Mitchell, H.M.; Morris, M.J. Changes in Gut Microbiota in Rats Fed a High Fat Diet Correlate with Obesity-Associated Metabolic Parameters. PLoS ONE 2015, 10, e0126931. [Google Scholar] [CrossRef] [PubMed]
- de Meij, T.G.J.; de Groot, E.F.J.; Eck, A.; Budding, A.E.; Kneepkens, C.M.F.; Benninga, M.A.; van Bodegraven, A.A.; Savelkoul, P.H.M. Characterization of Microbiota in Children with Chronic Functional Constipation. PLoS ONE 2016, 11, e0164731. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Das, C.; Mande, S.S. In Silico Analysis of Putrefaction Pathways in Bacteria and Its Implication in Colorectal Cancer. Front. Microbiol. 2017, 8, 2166. [Google Scholar] [CrossRef]
- Chen, M.; Fan, H.-N.; Chen, X.-Y.; Yi, Y.-C.; Zhang, J.; Zhu, J.-S. Alterations in the saliva microbiome in patients with gastritis and small bowel inflammation. Microb. Pathog. 2022, 165, 105491. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Zhang, P.; Shen, L.; Niu, L.; Tan, Y.; Chen, L.; Zhao, Y.; Bai, L.; Hao, X.; Li, X.; et al. Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2020, 21, 6356. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Containing Strain | Strain Combination |
|---|---|---|
| BM1 | B. infantis BLI-02 B. animalis BB-115 L. rhamnosus MP108 | BB-115 + MP108 + BLI-02 |
| BM2 | B. infantis BLI-02 B. animalis BB-115 L. rhamnosus MP108 L. paracasei CCFM2711 | BB-115 + MP108 + BLI-02 + CCFM2711 |
| PB1 | postbiotic PE0401 B. infantis BLI-02 B. animalis BB-115 L. rhamnosus MP108 | PE0401 + BB-115 + MP108 + BLI-02 |
| PB2 | postbiotic PE0401 B. infantis BLI-02 B. animalis BB-115 L. rhamnosus MP108 L. paracasei CCFM2711 | PE0401 + BB-115 + MP108 + BLI-02 + CCFM2711 |
| Group | Gavage Substance | Gavage Concentration | Gavage Volume |
|---|---|---|---|
| control | normal saline | - | 0.2 mL |
| model | loperamide hydrochloride | 15 mg/(kg·bw) | 0.2 mL |
| excipient | maltodextrin | 500 mg/(kg·bw) | 0.2 mL |
| BM1 | BB-115 + MP108 + BLI-02 | 4.1 × 108 CFU/g | 0.2 mL |
| BM2 | BB-115 + MP108 + BLI-02 + CCFM2711 | 4.1 × 108 CFU/g | 0.2 mL |
| PB1 | PE0401 + BB-115 + MP108 + BLI-02 | 4.1 × 108 CFU/g | 0.2 mL |
| PB2 | PE0401 + BB-115 + MP108 + BLI-02 + CCFM2711 | 4.1 × 108 CFU/g | 0.2 mL |
| Group | Initial Weight | Final Weight | p |
|---|---|---|---|
| control | 23.16 ± 1.06 | 25.04 ± 1.66 | - |
| model | 23.99 ± 1.52 | 26.14 ± 2.29 | <0.05 |
| excipient | 24.44 ± 1.17 | 25.78 ± 1.61 | - |
| BM1 | 22.89 ± 0.79 | 24.74 ± 1.19 | - |
| BM2 | 23.86 ± 1.05 | 26.07 ± 1.21 | <0.05 |
| PB1 | 24.77 ± 1.61 | 26.81 ± 1.15 | - |
| PB2 | 22.82 ± 0.65 | 24.49 ± 1.08 | - |
| Factors | Model | BM1 | BM2 | PB2 | Figures | |
|---|---|---|---|---|---|---|
| Gastrointestinal Indices | Faecal water content | ↓ | ↑ | ↑ | ↑ | Figure 3A |
| Time of first black stool defecation | ↑ | ↓ | ↓ | ↓ | Figure 3B | |
| Small intestine propulsion rate | ↓ | ↑ | - | - | Figure 3C | |
| Neurotransmitters | MTL↓, VIP↑, PYY↑ | MTL↑, VIP↓ | - | MTL↑, VIP↓, PYY↓ | Figure 4 | |
| Inflammatory Factors | IL-1↑, IL-6↑, IL-8↑ | IL-1↓, IL-6↓, IL-8↓ | IL-6↓, IL-8↓ | IL-1↓, IL-6↓, IL-8↓ | Figure 5 | |
| SCFAs | AA↓, PA↓, BA↓, IVA↓ | AA↑, PA↑, IVA↑ | AA↑, PA↑, BA↑, IVA↑ | AA↑ | Figure 6A | |
| Intestinal Flora | Tyzzerella↑, Enterorhabdus↑, Faecalibaculum↑, Gordonibacter↑, Mucispirillum↑ | Parabacteroides↑, Tyzzerella↓, Enterorhabdus↓, Faecalibaculum↓, Gordonibacter↓, Mucispirillum↓ | Parabacteroides↑, Gordonibacter↓, Mucispirillum↓ | Tyzzerella↓, Enterorhabdus↓, Faecalibaculum↓, Gordonibacter↓, Mucispirillum↓ | Figure 7 and Figure 8 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Wang, L.; Liu, X.; Wang, G.; Guo, X.; Liu, X.; Zhao, J.; Chen, W. The Different Ways Multi-Strain Probiotics with Different Ratios of Bifidobacterium and Lactobacillus Relieve Constipation Induced by Loperamide in Mice. Nutrients 2023, 15, 4230. https://doi.org/10.3390/nu15194230
Zhang C, Wang L, Liu X, Wang G, Guo X, Liu X, Zhao J, Chen W. The Different Ways Multi-Strain Probiotics with Different Ratios of Bifidobacterium and Lactobacillus Relieve Constipation Induced by Loperamide in Mice. Nutrients. 2023; 15(19):4230. https://doi.org/10.3390/nu15194230
Chicago/Turabian StyleZhang, Chenyue, Linlin Wang, Xiaoming Liu, Gang Wang, Xinmei Guo, Xuecong Liu, Jianxin Zhao, and Wei Chen. 2023. "The Different Ways Multi-Strain Probiotics with Different Ratios of Bifidobacterium and Lactobacillus Relieve Constipation Induced by Loperamide in Mice" Nutrients 15, no. 19: 4230. https://doi.org/10.3390/nu15194230
APA StyleZhang, C., Wang, L., Liu, X., Wang, G., Guo, X., Liu, X., Zhao, J., & Chen, W. (2023). The Different Ways Multi-Strain Probiotics with Different Ratios of Bifidobacterium and Lactobacillus Relieve Constipation Induced by Loperamide in Mice. Nutrients, 15(19), 4230. https://doi.org/10.3390/nu15194230