
A Multi-Omics and Human Biomonitoring Approach to Assessing the Effectiveness of Fortified Balanced Energy–Protein Supplementation on Maternal and Newborn Health in Burkina Faso: A Study Protocol

 , ,
, ,  , , , , , , , , , , , , ,
, , , , , , , , , , , , ,  and
and
Abstract
:1. Background
2. Methods: Study Design, Biospecimen Collection and Rationale
2.1. Study Setting, Participants and Enrolment Procedures
2.2. Study Supplements
2.3. Biospecimen Collection
2.3.1. Whole Blood (Mother–Infant Dyads)
2.3.2. Plasma (Mothers)
2.3.3. Cord Blood
2.3.4. Urine (Mothers)
2.3.5. Breast Milk (Mothers)
2.3.6. Feces (Mother–Infant Dyads)
2.4. Rationale for the Analysis of Biospecimens with Related Bio-Measurements
2.4.1. Metabolomics
2.4.2. Metagenomics
2.4.3. Proteomics
2.4.4. Breast Milk Characterization
2.5. Human Biomonitoring
2.5.1. Telomere Length and Mitochondrial DNA
2.5.2. Air and Smoke Pollution
2.5.3. Mycotoxins
2.5.4. Environmental Contaminants
2.5.5. Gut Enteropathogens
2.5.6. Fecal Inflammatory Markers
Calprotectin
Short-Chain Fatty Acids
3. Metadata
4. Data Quality Control
5. Ethical Considerations
6. Strengths and Limitations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- UNICEF-WHO. Low Birthweight Estimates: Levels and Trends 2000–2015; World Health Organization: Geneva, Switzerland, 2019; Volume 4, pp. 3–9. Available online: https://www.unicef.org/reports/UNICEF-WHO-low-birthweight-estimates-2019 (accessed on 16 January 2023).
- Benammar, A.; Sermondade, N.; Faure, C.; Dupont, C.; Cedrin-Durnerin, I.; Sifer, C.; Hercberg, S.; Levy, R. Nutrition et fausses couches spontanées: Une revue de la littérature. Gynécologie Obs. Fertil. 2012, 40, 162–169. [Google Scholar] [CrossRef] [PubMed]
- McClure, E.M.; Saleem, S.; Pasha, O.; Goldenberg, R.L. Stillbirth in developing countries: A review of causes, risk factors and prevention strategies. J. Matern.-Fetal Neonatal Med. 2009, 22, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Almond, D.; Chay, K.; Lee, D. The Costs of Low Birth Weight. Q. J. Econ. 2004, 120, 1031–1083. [Google Scholar] [CrossRef]
- WHO. WHO Recommendations on Antenatal Care for a Positive Pregnancy Experience. In WHO Recommendations on Antenatal Care for a Positive Pregnancy Experience; World Health Organization: Geneva, Switzerland, 2016. Available online: https://www.ncbi.nlm.nih.gov/books/NBK409108/ (accessed on 25 January 2022).
- Ota, E.; Tobe-Gai, R.; Mori, R.; Farrar, D. Antenatal dietary advice and supplementation to increase energy and protein intake. Cochrane Database Syst. Rev. 2012, 9, CD000032. [Google Scholar] [CrossRef]
- Gernand, A.D.; Gallagher, K.; Bhandari, N.; Kolsteren, P.; Lee, A.C.; Shafiq, Y.; Taneja, S.; Tielsch, J.M.; Abate, F.W.; Baye, E.; et al. Harmonization of maternal balanced energy-protein supplementation studies for individual participant data (IPD) meta-analyses—Finding and creating similarities in variables and data collection. BMC Pregnancy Childbirth 2023, 23, 107. [Google Scholar] [CrossRef]
- Vanslambrouck, K.; de Kok, B.; Toe, L.C.; De Cock, N.; Ouedraogo, M.; Dailey-Chwalibóg, T.; Hanley-Cook, G.; Ganaba, R.; Lachat, C.; Huybregts, L.; et al. Effect of balanced energy-protein supplementation during pregnancy and lactation on birth outcomes and infant growth in rural Burkina Faso: Study protocol for a randomised controlled trial. BMJ Open 2021, 11, e038393. [Google Scholar] [CrossRef]
- Villar, J.; Altman, D.G.; Purwar, M.; Noble, J.A.; Knight, H.E.; Ruyan, P.; Ismail, L.C.; Barros, F.C.; Lambert, A.; Papageorghiou, A.T.; et al. The objectives, design and implementation of the INTERGROWTH-21st Project. BJOG Int. J. Obstet. Gynaecol. 2013, 120 (Suppl. 2), 9–26. [Google Scholar] [CrossRef]
- De Onis, M.; Branca, F. Childhood stunting: A global perspective. Matern. Child Nutr. 2016, 12 (Suppl. 1), 12–26. [Google Scholar] [CrossRef]
- Argaw, A.; Toe, L.C.; Hanley-Cook, G.; Dailey-Chwalibóg, T.; de Kok, B.; Ouédraogo, L.; Compaoré, A.; Ouédraogo, M.; Sawadogo, A.; Ganaba, R.; et al. Effect of prenatal micronutrient-fortified balanced energy-protein supplementation on maternal and newborn body composition: A sub-study from the MISAME-III randomized controlled efficacy trial in rural Burkina Faso. PLoS Med. 2023, 20, e1004242. [Google Scholar] [CrossRef]
- Argaw, A.; de Kok, B.; Toe, L.C.; Hanley-Cook, G.; Dailey-Chwalibóg, T.; Ouédraogo, M.; Compaoré, A.; Vanslambrouck, K.; Ganaba, R.; Kolsteren, P.; et al. Fortified balanced energy–protein supplementation during pregnancy and lactation and infant growth in rural Burkina Faso: A 2 × 2 factorial individually randomized controlled trial. PLoS Med. 2023, 20, e1004186. [Google Scholar] [CrossRef]
- De Kok, B.; Toe, L.C.; Hanley-Cook, G.; Argaw, A.; Ouédraogo, M.; Compaoré, A.; Vanslambrouck, K.; Dailey-Chwalibóg, T.; Ganaba, R.; Kolsteren, P.; et al. Prenatal fortified balanced energy-Protein supplementation and birth outcomes in rural Burkina Faso: A randomized controlled efficacy trial. PLoS Med. 2022, 19, e1004002. [Google Scholar] [CrossRef]
- Hanley-Cook, G.T.; Bastos-Moreira, Y.; Martens, D.S.; Dailey-Chwalibóg, T.; Toe, L.C.; de Kok, B.; Ouédraogo, L.; Argaw, A.; Tesfamariam, K.; Kolsteren, P.; et al. Prenatal multiple micronutrient-fortified balanced energy-protein supplementation and newborn telomere length and mitochondrial DNA content: A randomized controlled efficacy trial in rural Burkina Faso. Manuscript submitted for publication.
- Perumal, N.; Bassani, D.G.; Roth, D.E. Use and Misuse of Stunting as a Measure of Child Health. J. Nutr. 2018, 148, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Hanley-Cook, G.; Toe, L.C.; Tesfamariam, K.; de Kok, B.; Argaw, A.; Compaoré, A.; Ouédraogo, M.; Dailey-Chwalibóg, T.; Kolsteren, P.; Lachat, C.; et al. Fortified Balanced Energy-Protein Supplementation, Maternal Anemia, and Gestational Weight Gain: A Randomized Controlled Efficacy Trial among Pregnant Women in Rural Burkina Faso. J. Nutr. 2022, 152, 2277–2286. [Google Scholar] [CrossRef] [PubMed]
- De Kok, B.; Argaw, A.; Hanley-Cook, G.; Toe, L.C.; Ouédraogo, M.; Dailey-Chwalibóg, T.; Diop, L.; Becquey, E.; Kolsteren, P.; Lachat, C.; et al. Fortified Balanced Energy-Protein Supplements Increase Nutrient Adequacy without Displacing Food Intake in Pregnant Women in Rural Burkina Faso. J. Nutr. 2021, 151, 3831–3840. [Google Scholar] [CrossRef]
- Hanley-Cook, G.T.; Argaw, A.; de Kok, B.; Toe, L.C.; Dailey-Chwalibóg, T.; Ouédraogo, M.; Kolsteren, P.; Huybregts, L.; Lachat, C. Seasonality and Day-to-Day Variability of Dietary Diversity: Longitudinal Study of Pregnant Women Enrolled in a Randomized Controlled Efficacy Trial in Rural Burkina Faso. J. Nutr. 2022, 152, 2145–2154. [Google Scholar] [CrossRef]
- Ministère de la Santé—Burkina Faso. Politique et Normes en Matière de Santé de la Reproduction; Ministère de la Santé: Ouagadougou, Burkina Faso, 2010. Available online: https://www.prb.org/wp-content/uploads/2018/05/Politiques-et-Normes-en-Matière-de-Santeé-de-la-Reproduction-au-Burkina-Faso-2010.pdf (accessed on 9 July 2023).
- Jones, L.; de Kok, B.; Moore, K.; de Pee, S.; Bedford, J.; Vanslambrouck, K.; Toe, L.C.; Lachat, C.; De Cock, N.; Ouédraogo, M.; et al. Acceptability of 12 fortified balanced energy protein supplements—Insights from Burkina Faso. Matern. Child Nutr. 2021, 17, e13067. [Google Scholar] [CrossRef]
- Bill & Melinda Gates Foundation. Framework and Specifications for the Nutritional Composition of a Food Supplement for Pregnant and Lactating Women (PLW) In Undernourished and Low Income Settings; Gates Open Research: Seattle, WA, USA, 2017; pp. 1–23. [Google Scholar] [CrossRef]
- Allen, L.H.; Carriquiry, A.L.; Murphy, S.P. Perspective: Proposed Harmonized Nutrient Reference Values for Populations. Adv. Nutr. 2020, 11, 469–483. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Belova, L.; Stove, C.; De Boevre, M.; De Saeger, S. Volumetric Absorptive Microsampling as an Alternative Tool for Biomonitoring of Multi-Mycotoxin Exposure in Resource-Limited Areas. Toxins 2021, 13, 345. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Eiden, M.; Morin-Rivron, D.; Christinat, N.; Monteiro, J.P.; Kaput, J.; Masoodi, M. Impact of multi-micronutrient supplementation on lipidemia of children and adolescents. Clin. Nutr. 2020, 39, 2211–2219. [Google Scholar] [CrossRef] [PubMed]
- Weissinger, E.M.; Nguyen-Khoa, T.; Fumeron, C.; Saltiel, C.; Walden, M.; Kaiser, T.; Mischak, H.; Drüeke, T.B.; Lacour, B.; Massy, Z.A. Effects of oral vitamin C supplementation in hemodialysis patients: A proteomic assessment. Proteomics 2006, 6, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Smith, D.L.; Esteves, K.; Drury, S. Telomere length measurement by qPCR—Summary of critical factors and recommendations for assay design. Psychoneuroendocrinology 2019, 99, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Barr, D.B.; Wilder, L.C.; Caudill, S.P.; Gonzalez, A.J.; Needham, L.L.; Pirkle, J.L. Urinary creatinine concentrations in the U.S. population: Implications for urinary biologic monitoring measurements. Environ. Health Perspect. 2005, 113, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Esteban, M.; Castaño, A. Non-invasive matrices in human biomonitoring: A review. Environ. Int. 2009, 35, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Li, Z.; Zhang, W.; Zhang, C.; Zhang, Y.; Mei, H.; Zhuo, N.; Wang, H.; Wang, L.; Wu, D. Comparison of gut microbiota in exclusively breast-fed and formula-fed babies: A study of 91 term infants. Sci. Rep. 2020, 10, 15792. [Google Scholar] [CrossRef]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Fanos, V.; Reali, A. “Omics” in Human Colostrum and Mature Milk: Looking to Old Data with New Eyes. Nutrients 2017, 9, 843. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Fanos, V.; Strigini, F.A.L.; Artini, P.G.; Peroni, D.G. Human Breast Milk: Exploring the Linking Ring Among Emerging Components. Front. Pediatr. 2018, 6, 215. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Peila, C.; Fanos, V.; Coscia, A. Clinical insights gained through metabolomic analysis of human breast milk. Expert Rev. Proteom. 2019, 16, 909–932. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Puddu, M.; Peroni, D.G.; Fanos, V. The Human Breast Milk Metabolome in Overweight and Obese Mothers. Front. Immunol. 2020, 11, 1533. [Google Scholar] [CrossRef]
- Bardanzellu, F.; Reali, A.; Marcialis, M.A.; Fanos, V. Across the “Sweetest” Properties of Human Breast Milk: Focus on Oligosaccharides. Curr. Pediatr. Rev. 2021, 16, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Congiu, M.; Reali, A.; Deidda, F.; Dessì, A.; Bardanzellu, F.; Fanos, V. Breast Milk for Preterm Multiples: More Proteins, Less Lactose. Twin Res. Hum. Genet. 2019, 22, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Dallas, D.C.; Guerrero, A.; Khaldi, N.; Castillo, P.A.; Martin, W.F.; Smilowitz, J.T.; Bevins, C.L.; Barile, D.; German, J.B.; Lebrilla, C.B. Extensive in vivo Human Milk Peptidomics Reveals Specific Proteolysis Yielding Protective Antimicrobial Peptides. J. Proteome Res. 2013, 12, 2295–2304. [Google Scholar] [CrossRef] [PubMed]
- van Herwijnen, M.J.; Zonneveld, M.I.; Goerdayal, S.; Hoen, E.N.N.; Garssen, J.; Stahl, B.; Altelaar, A.M.; Redegeld, F.A.; Wauben, M.H. Comprehensive Proteomic Analysis of Human Milk-derived Extracellular Vesicles Unveils a Novel Functional Proteome Distinct from Other Milk Components. Mol. Cell. Proteom. 2016, 15, 3412–3423. [Google Scholar] [CrossRef]
- Kisuse, J.; La-Ongkham, O.; Nakphaichit, M.; Therdtatha, P.; Momoda, R.; Tanaka, M.; Fukuda, S.; Popluechai, S.; Kespechara, K.; Sonomoto, K.; et al. Urban Diets Linked to Gut Microbiome and Metabolome Alterations in Children: A Comparative Cross-Sectional Study in Thailand. Front. Microbiol. 2018, 9, 1345. [Google Scholar] [CrossRef]
- Nguyen, Q.P.; Karagas, M.R.; Madan, J.C.; Dade, E.; Palys, T.J.; Morrison, H.G.; Pathmasiri, W.W.; McRitche, S.; Sumner, S.J.; Frost, H.R.; et al. Associations between the gut microbiome and metabolome in early life. BMC Microbiol. 2021, 21, 238. [Google Scholar] [CrossRef]
- Gonzalez, C.G.; Zhang, L.; Elias, J.E. From mystery to mechanism: Can proteomics build systems-level understanding of our gut microbes? Expert Rev. Proteom. 2017, 14, 473–476. [Google Scholar] [CrossRef]
- Villar, J.; Ochieng, R.; Gunier, R.B.; Papageorghiou, A.T.; Rauch, S.; McGready, R.; Gauglitz, J.M.; Barros, F.C.; Vatish, M.; Fernandes, M.; et al. Association between fetal abdominal growth trajectories, maternal metabolite signatures early in pregnancy, and childhood growth and adiposity: Prospective observational multinational INTERBIO-21st fetal study. Lancet Diabetes Endocrinol. 2022, 10, 710–719. [Google Scholar] [CrossRef]
- Olm, M.R.; Crits-Christoph, A.; Bouma-Gregson, K.; Firek, B.A.; Morowitz, M.J.; Banfield, J.F. inStrain profiles population microdiversity from metagenomic data and sensitively detects shared microbial strains. Nat. Biotechnol. 2021, 39, 727–736. [Google Scholar] [CrossRef]
- Aramaki, T.; Blanc-Mathieu, R.; Endo, H.; Ohkubo, K.; Kanehisa, M.; Goto, S.; Ogata, H. KofamKOALA: KEGG Ortholog assignment based on profile HMM and adaptive score threshold. Bioinformatics 2019, 36, 2251–2252. [Google Scholar] [CrossRef]
- Mc Ardle, A.; Binek, A.; Moradian, A.; Orgel, B.C.; Rivas, A.; Washington, K.E.; Phebus, C.; Manalo, D.-M.; Go, J.; Venkatraman, V.; et al. Standardized Workflow for Precise Mid- and High-Throughput Proteomics of Blood Biofluids. Clin. Chem. 2021, 68, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Smilowitz, J.T.; Gho, D.S.; Mirmiran, M.; German, J.B.; Underwood, M.A. Rapid measurement of human milk macronutrients in the neonatal intensive care unit: Accuracy and precision of fourier transform mid-infrared spectroscopy. J. Hum. Lact. 2014, 30, 180–189. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; Dror, D.K.; Allen, L.H. Micronutrients in Human Milk: Analytical Methods. Adv. Nutr. 2018, 9 (Suppl. 1), 313S–331S. [Google Scholar] [CrossRef] [PubMed]
- Hampel, D.; York, E.R.; Allen, L.H. Ultra-performance liquid chromatography tandem mass-spectrometry (UPLC-MS/MS) for the rapid, simultaneous analysis of thiamin, riboflavin, flavin adenine dinucleotide, nicotinamide and pyridoxal in human milk. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2012, 903, 7–13. [Google Scholar] [CrossRef]
- Kellman, B.P.; Richelle, A.; Yang, J.-Y.; Chapla, D.; Chiang, A.W.T.; Najera, J.A.; Liang, C.; Fürst, A.; Bao, B.; Koga, N.; et al. Elucidating Human Milk Oligosaccharide biosynthetic genes through network-based multi-omics integration. Nat. Commun. 2022, 13, 2455. [Google Scholar] [CrossRef]
- Ju, H.; Lai, G.; Yan, F. Electrochemiluminescent immunosensing. In Immunosensing for Detection of Protein Biomarkers; Elsevier: Amsterdam, The Netherlands, 2017; pp. 171–206. [Google Scholar] [CrossRef]
- Ramani, S.; Stewart, C.J.; Laucirica, D.R.; Ajami, N.J.; Robertson, B.; Autran, C.A.; Shinge, D.; Rani, S.; Anandan, S.; Hu, L.; et al. Human milk oligosaccharides, milk microbiome and infant gut microbiome modulate neonatal rotavirus infection. Nat. Commun. 2018, 9, 5010. [Google Scholar] [CrossRef]
- Langsdorf, M.; Ömer, G.; Dearth, S.; Röhring, C.; Pham, T.H.; Dallmann, G.; Koal, T. Standardized Quantitative Metabolomics Using the Biocrates MxP® Quant 500 XL Kit across Mass Spectrometer Platforms. 2023. Available online: https://biocrates.com/wp-content/uploads/2023/06/Application-note-Quant500-XL-across-MS-platforms.pdf (accessed on 10 September 2023).
- Semba, R.D.; Shardell, M.; Ashour, F.A.S.; Moaddel, R.; Trehan, I.; Maleta, K.M.; Ordiz, M.I.; Kraemer, K.; Khadeer, M.A.; Ferrucci, L.; et al. Child Stunting is Associated with Low Circulating Essential Amino Acids. EBioMedicine 2016, 6, 246–252. [Google Scholar] [CrossRef]
- Hampel, D.; Shahab-Ferdows, S.; Hossain, M.; Islam, M.M.; Ahmed, T.; Allen, L.H. Validation and Application of Biocrates AbsoluteIDQ® p180 Targeted Metabolomics Kit Using Human Milk. Nutrients 2019, 11, 1733. [Google Scholar] [CrossRef]
- Tanaka, M.; Nakayama, J. Development of the gut microbiota in infancy and its impact on health in later life. Allergol. Int. Off. J. Jpn. Soc. Allergol. 2017, 66, 515–522. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef]
- Gritz, E.C.; Ebhandari, V. The Human Neonatal Gut Microbiome: A Brief Review. Front. Pediatr. 2015, 3, 17. [Google Scholar] [CrossRef]
- Marques, T.M.; Wall, R.; Ross, R.P.; Fitzgerald, G.F.; Ryan, C.A.; Stanton, C. Programming infant gut microbiota: Influence of dietary and environmental factors. Curr. Opin. Biotechnol. 2010, 21, 149–156. [Google Scholar] [CrossRef]
- Gomez-Llorente, C.; Plaza-Diaz, J.; Aguilera, M.; Muñoz-Quezada, S.; Bermudez-Brito, M.; Peso-Echarri, P.; Martinez-Silla, R.; Vasallo-Morillas, M.I.; Campaña-Martin, L.; Vives-Piñera, I.; et al. Three Main Factors Define Changes in Fecal Microbiota Associated With Feeding Modality in Infants. J. Pediatr. Gastroenterol. Nutr. 2013, 57, 461–466. [Google Scholar] [CrossRef]
- Vray, M.; Hedible, B.G.; Adam, P.; Tondeur, L.; Manirazika, A.; Randremanana, R.; Mainassara, H.; Briend, A.; Artaud, C.; von Platen, C.; et al. A multicenter, randomized controlled comparison of three renutrition strategies for the management of moderate acute malnutrition among children aged from 6 to 24 months (the MALINEA project). Trials 2018, 19, 666. [Google Scholar] [CrossRef]
- Adeola, H.A.; Blackburn, J.M.; Rebbeck, T.R.; Zerbini, L.F. Emerging proteomics biomarkers and prostate cancer burden in Africa. Oncotarget 2017, 8, 37991–38007. [Google Scholar] [CrossRef]
- Navarro, S.L.; White, E.; Kantor, E.D.; Zhang, Y.; Rho, J.; Song, X.; Milne, G.L.; Lampe, P.D.; Lampe, J.W. Randomized Trial of Glucosamine and Chondroitin Supplementation on Inflammation and Oxidative Stress Biomarkers and Plasma Proteomics Profiles in Healthy Humans. PLoS ONE 2015, 10, e0117534. [Google Scholar] [CrossRef]
- Boudry, G.; Charton, E.; Le Huerou-Luron, I.; Ferret-Bernard, S.; Le Gall, S.; Even, S.; Blat, S. The Relationship Between Breast Milk Components and the Infant Gut Microbiota. Front. Nutr. 2021, 8, 629740. [Google Scholar] [CrossRef]
- Zhang, S.; Li, T.; Xie, J.; Zhang, D.; Pi, C.; Zhou, L.; Yang, W. Gold standard for nutrition: A review of human milk oligosaccharide and its effects on infant gut microbiota. Microb. Cell Factories 2021, 20, 108. [Google Scholar] [CrossRef]
- Demmelmair, H.; Prell, C.; Timby, N.; Lönnerdal, B. Benefits of Lactoferrin, Osteopontin and Milk Fat Globule Membranes for Infants. Nutrients 2017, 9, 817. [Google Scholar] [CrossRef]
- Miller, G.W.; Jones, D.P. The nature of nurture: Refining the definition of the exposome. Toxicol. Sci. Off. J. Soc. Toxicol. 2014, 137, 1–2. [Google Scholar] [CrossRef]
- Wild, C.P. Complementing the genome with an “exposome”: The outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere measurement by quantitative PCR. Nucleic Acids Res. 2002, 30, e47. [Google Scholar] [CrossRef] [PubMed]
- Cawthon, R.M. Telomere length measurement by a novel monochrome multiplex quantitative PCR method. Nucleic Acids Res. 2009, 37, e21. [Google Scholar] [CrossRef] [PubMed]
- Janssen, B.G.; Munters, E.; Pieters, N.; Smeets, K.; Cox, B.; Cuypers, A.; Fierens, F.; Penders, J.; Vangronsveld, J.; Gyselaers, W.; et al. Placental mitochondrial DNA content and particulate air pollution during in utero life. Environ. Health Perspect. 2012, 120, 1346–1352. [Google Scholar] [CrossRef]
- Martens, D.S.; Plusquin, M.; Gyselaers, W.; De Vivo, I.; Nawrot, T.S. Maternal pre-pregnancy body mass index and newborn telomere length. BMC Med. 2016, 14, 148. [Google Scholar] [CrossRef]
- Martens, D.S.; Janssen, B.G.; Bijnens, E.M.; Clemente, D.B.P.; Vineis, P.; Plusquin, M.; Nawrot, T.S. Association of Parental Socioeconomic Status and Newborn Telomere Length. JAMA Netw. Open 2020, 3, e204057. [Google Scholar] [CrossRef]
- Bové, H.; Bongaerts, E.; Slenders, E.; Bijnens, E.M.; Saenen, N.D.; Gyselaers, W.; Van Eyken, P.; Plusquin, M.; Roeffaers, M.B.J.; Ameloot, M.; et al. Ambient black carbon particles reach the fetal side of human placenta. Nat. Commun. 2019, 10, 3866. [Google Scholar] [CrossRef]
- Saenen, N.D.; Bové, H.; Steuwe, C.; Roeffaers, M.B.J.; Provost, E.B.; Lefebvre, W.; Vanpoucke, C.; Ameloot, M.; Nawrot, T.S. Children’s urinary environmental carbon load: A novel marker reflecting residential ambient air pollution exposure? Am. J. Respir. Crit. Care Med. 2017, 196, 873–881. [Google Scholar] [CrossRef]
- Caballero-Casero, N.; Castro, G.; Bastiaensen, M.; Gys, C.; van Larebeke, N.; Schoeters, G.; Covaci, A. Identification of chemicals of emerging concern in urine of Flemish adolescents using a new suspect screening workflow for LC-QTOF-MS. Chemosphere 2021, 280, 130683. [Google Scholar] [CrossRef]
- Gys, C.; Bamai, Y.A.; Araki, A.; Bastiaensen, M.; Caballero-Casero, N.; Kishi, R.; Covaci, A. Biomonitoring and temporal trends of bisphenols exposure in Japanese school children. Environ. Res. 2020, 191, 110172. [Google Scholar] [CrossRef]
- DeBoer, M.D.; Platts-Mills, J.A.; Scharf, R.J.; McDermid, J.M.; Wanjuhi, A.W.; Gratz, J.; Svensen, E.; Swann, J.R.; Donowitz, J.R.; Jatosh, S.; et al. Early Life Interventions for Childhood Growth and Development in Tanzania (ELICIT): A protocol for a randomised factorial, double-blind, placebo-controlled trial of azithromycin, nitazoxanide and nicotinamide. BMJ Open 2018, 8, e021817. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Kabir, F.; Manneh, J.; Lertsethtakarn, P.; Begum, S.; Gratz, J.; Becker, S.M.; Operario, D.J.; Taniuchi, M.; Janaki, L.; et al. Development and assessment of molecular diagnostic tests for 15 enteropathogens causing childhood diarrhoea: A multicentre study. Lancet Infect. Dis. 2014, 14, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, S.J.; French, J.; Brookes, M.J.; Ford, C.; Gama, R. Between-assay variability of faecal calprotectin enzyme-linked immunosorbent assay kits. Ann. Clin. Biochem. 2013, 50, 53–61. [Google Scholar] [CrossRef]
- Toe, L.C.; Kerckhof, F.-M.; De Bodt, J.; Morel, F.B.; Ouedraogo, J.-B.; Kolsteren, P.; Van de Wiele, T. A prebiotic-enhanced lipid-based nutrient supplement (LNSp) increases Bifidobacterium relative abundance and enhances short-chain fatty acid production in simulated colonic microbiota from undernourished infants. FEMS Microbiol. Ecol. 2020, 96, fiaa105. [Google Scholar] [CrossRef]
- Blackburn, E.H.; Epel, E.S.; Lin, J. Human telomere biology: A contributory and interactive factor in aging, disease risks, and protection. Science 2015, 350, 1193–1198. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Nawrot, T.S.; Van Der Stukken, C.; Tylus, D.; Sleurs, H.; Peusens, M.; Alfano, R.; Langie, S.A.; Plusquin, M.; Martens, D.S. Different epigenetic signatures of newborn telomere length and telomere attrition rate in early life. Aging 2021, 13, 14630–14650. [Google Scholar] [CrossRef] [PubMed]
- Haycock, P.C.; Heydon, E.E.; Kaptoge, S.; Butterworth, A.S.; Thompson, A.; Willeit, P. Leucocyte telomere length and risk of cardiovascular disease: Systematic review and meta-analysis. BMJ 2014, 349, g4227. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Zhan, Y.; Pedersen, N.L.; Fang, F.; Hägg, S. Telomere Length and All-Cause Mortality: A Meta-analysis. Ageing Res. Rev. 2018, 48, 11–20. [Google Scholar] [CrossRef]
- Tsoukalas, D.; Fragkiadaki, P.; Docea, A.O.; Alegakis, A.K.; Sarandi, E.; Vakonaki, E.; Salataj, E.; Kouvidi, E.; Nikitovic, D.; Kovatsi, L.; et al. Association of nutraceutical supplements with longer telomere length. Int. J. Mol. Med. 2019, 44, 218–226. [Google Scholar] [CrossRef]
- Roger, A.J.; Muñoz-Gómez, S.A.; Kamikawa, R. The Origin and Diversification of Mitochondria. Curr. Biol. CB 2017, 27, R1177–R1192. [Google Scholar] [CrossRef]
- Copeland, W.C.; Longley, M.J. Mitochondrial genome maintenance in health and disease. DNA Repair 2014, 19, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Tait, S.W.G.; Green, D.R. Mitochondrial Regulation of Cell Death. Cold Spring Harb. Perspect. Biol. 2013, 5, a008706. [Google Scholar] [CrossRef] [PubMed]
- Gibson, K.; Halliday, J.L.; Kirby, D.M.; Yaplito-Lee, J.; Thorburn, D.R.; Boneh, A. Mitochondrial oxidative phosphorylation disorders presenting in neonates: Clinical manifestations and enzy-matic and molecular diagnoses. Pediatrics 2008, 122, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Kohda, M.; Tokuzawa, Y.; Kishita, Y.; Nyuzuki, H.; Moriyama, Y.; Mizuno, Y.; Hirata, T.; Yatsuka, Y.; Yamashita-Sugahara, Y.; Nakachi, Y.; et al. A Comprehensive Genomic Analysis Reveals the Genetic Landscape of Mitochondrial Respiratory Chain Complex Deficiencies. PLoS Genet. 2016, 12, e1005679. [Google Scholar] [CrossRef]
- Druzhyna, N.M.; Wilson, G.L.; LeDoux, S.P. Mitochondrial DNA repair in aging and disease. Mech. Ageing Dev. 2008, 129, 383–390. [Google Scholar] [CrossRef]
- Priliani, L.; Prado, E.L.; Restuadi, R.; Waturangi, D.E.; Shankar, A.H.; Malik, S.G. Maternal Multiple Micronutrient Supplementation Stabilizes Mitochondrial DNA Copy Number in Pregnant Women in Lombok, Indonesia. J. Nutr. 2019, 149, 1309–1316. [Google Scholar] [CrossRef]
- Sana, A.; Meda, N.; Badoum, G.; Kafando, B.; Bouland, C. Primary Cooking Fuel Choice and Respiratory Health Outcomes among Women in Charge of Household Cooking in Ouagadougou, Burkina Faso: Cross-Sectional Study. Int. J. Environ. Res. Public Health 2019, 16, 1040. [Google Scholar] [CrossRef]
- WHO. National Burden of Disease Estimates; World Health Organization: Geneva, Switzerland, 2007; pp. 1–8. Available online: http://www.who.int/indoorair/publications/fuelforlife/ (accessed on 18 January 2023).
- Pedersen, M.; Giorgis-Allemand, L.; Bernard, C.; Aguilera, I.; Andersen, A.-M.N.; Ballester, F.; Beelen, R.M.J.; Chatzi, L.; Cirach, M.; Danileviciute, A.; et al. Ambient air pollution and low birthweight: A European cohort study (ESCAPE). Lancet Respir. Med. 2013, 1, 695–704. [Google Scholar] [CrossRef]
- Brook, R.D.; Rajagopalan, S.; Pope, C.A.; Brook, J.R.; Bhatnagar, A.; Diez-Roux, A.V.; Holguin, F.; Hong, Y.; Luepker, R.V.; Mittleman, M.A.; et al. Particulate Matter Air Pollution and Cardiovascular Disease: An update to the scientific statement from the american heart association. Circulation 2010, 121, 2331–2378. [Google Scholar] [CrossRef]
- Nawrot, T.S.; Perez, L.; Künzli, N.; Munters, E.; Nemery, B. Public health importance of triggers of myocardial infarction: A comparative risk assessment. Lancet 2011, 377, 732–740. [Google Scholar] [CrossRef]
- Neven, K.Y.; Wang, C.; Janssen, B.G.; Roels, H.A.; Vanpoucke, C.; Ruttens, A.; Nawrot, T.S. Ambient air pollution exposure during the late gestational period is linked with lower placental iodine load in a Belgian birth cohort. Environ. Int. 2021, 147, 106334. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.S.; Brasel, J.M. Toxicity, metabolism, and impact of mycotoxins on humans and animals. Toxicology 2001, 167, 101–134. [Google Scholar] [CrossRef] [PubMed]
- De Kok, B.; Moore, K.; Jones, L.; Vanslambrouck, K.; Toe, L.C.; Ouédraogo, M.; Ganaba, R.; de Pee, S.; Bedford, J.; Lachat, C.; et al. Home consumption of two fortified balanced energy protein supplements by pregnant women in Burkina Faso. Matern. Child Nutr. 2021, 17, e13134. [Google Scholar] [CrossRef] [PubMed]
- Kpoda, D.S.; Bandé, M.; Compaoré, A.M.; Bazié, R.B.S.; Meda, R.N.; Somda, S.; Meda, D.S.; Kpoda, H.B.N.; Somé, S.A.; Sakana, L.; et al. Nutritional, Microbiological, and Toxicological Quality Assessment of Foods Sold in Urban and Suburban Markets in Burkina Faso. Health Secur. 2022, 20, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Gratz, J.; Amour, C.; Kibiki, G.; Becker, S.; Janaki, L.; Verweij, J.J.; Taniuchi, M.; Sobuz, S.U.; Haque, R.; et al. A laboratory-developed TaqMan Array Card for simultaneous detection of 19 enteropathogens. J. Clin. Microbiol. 2013, 51, 472–480. [Google Scholar] [CrossRef]
- Marks, F.; Liu, J.; Soura, A.B.; Gasmelseed, N.; Operario, D.J.; Grundy, B.; Wieser, J.; Gratz, J.; Meyer, C.G.; Im, J.; et al. Pathogens That Cause Acute Febrile Illness Among Children and Adolescents in Burkina Faso, Madagascar, and Sudan. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2021, 73, 1338–1345. [Google Scholar] [CrossRef]
- Medithi, S.; Kasa, Y.D.; Kankipati, V.R.; Kodali, V.; Jee, B.; Jonnalagadda, P.R. Impact of Micronutrient Supplementation on Pesticide Residual, Acetylcholinesterase Activity, and Oxidative Stress Among Farm Children Exposed to Pesticides. Front. Public Health 2022, 10, 872125. [Google Scholar] [CrossRef]
- Moore, C.C.; Jacob, S.T.; Banura, P.; Zhang, J.; Stroup, S.; Boulware, D.R.; Scheld, W.M.; Houpt, E.R.; Liu, J. Etiology of Sepsis in Uganda Using a Quantitative Polymerase Chain Reaction-based TaqMan Array Card. Clin. Infect. Dis. 2018, 68, 266–272. [Google Scholar] [CrossRef]
- Ouedraogo, M.; Toe, A.M.; Ouedraogo, T.; Guissou, P.I. Pesticides in Burkina Faso: Overview of the Situation in a Sahelian African Country. In Pesticides in the Modern World—Pesticides Use and Management; InTech: Houston, TX, USA, 2011. [Google Scholar] [CrossRef]
- Peraica, M.; Richter, D.; Rašić, D. Mycotoxicoses in children. Arh. Za Hig. Rada I Toksikol. 2014, 65, 347–363. [Google Scholar] [CrossRef]
- Richards, L.; Li, M.; van Esch, B.; Garssen, J.; Folkerts, G. The effects of short-chain fatty acids on the cardiovascular system. PharmaNutrition 2016, 4, 68–111. [Google Scholar] [CrossRef]
- Saad-Hussein, A.; Ibrahim, K.S.; Abdalla, M.S.; El-Mezayen, H.; Osman, N.F. Effects of zinc supplementation on oxidant/antioxidant and lipids status of pesticides sprayers. J. Complement. Integr. Med. 2020, 17, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Scheppach, W.; Bartram, H.P.; Richter, F. Role of short-chain fatty acids in the prevention of colorectal cancer. Eur. J. Cancer 1995, 31, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Tesfamariam, K.; Argaw, A.; Hanley-Cook, G.; Gebreyesuse, S.H.; Kolsteren, P.; Belachew, T.; Van de Velde, M.; De Saeger, S.; De Boevre, M.; Lachat, C. Chronic maternal aflatoxin exposure is associated with poor fetal growth trajectories: A prospective cohort study in rural Ethiopia. 2022, unpublished work [Preprint]. [Google Scholar]
- Valadez, J.J.; Brown, L.D.; Vargas, W.V.; Morley, D. Using lot quality assurance sampling to assess measurements for growth monitoring in a developing country’s primary health care system. Int. J. Epidemiol. 1996, 25, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Vidal, A.; Mengelers, M.; Yang, S.; De Saeger, S.; De Boevre, M. Mycotoxin Biomarkers of Exposure: A Comprehensive Review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1127–1155. [Google Scholar] [CrossRef] [PubMed]
- Ware, L.Y.; Durand, N.; Nikiema, P.A.; Alter, P.; Fontana, A.; Montet, D.; Barro, N. Occurrence of mycotoxins in commercial infant formulas locally produced in Ouagadougou (Burkina Faso). Food Control 2017, 73, 518–523. [Google Scholar] [CrossRef]
- FAO. Worldwide Regulations for Mycotoxins in Food and Feed in 2003; FAO: Roma, Italy, 2003; Available online: https://www.fao.org/3/y5499e/y5499e0d.htm (accessed on 1 July 2022).
- Warth, B.; Parich, A.; Atehnkeng, J.; Bandyopadhyay, R.; Schuhmacher, R.; Sulyok, M.; Krska, R. Quantitation of Mycotoxins in Food and Feed from Burkina Faso and Mozambique Using a Modern LC-MS/MS Multitoxin Method. J. Agric. Food Chem. 2012, 60, 9352–9363. [Google Scholar] [CrossRef]
- Lehmann, E.; Oltramare, C.; Dibié, J.-J.N.; Konaté, Y.; de Alencastro, L.F. Assessment of human exposure to pesticides by hair analysis: The case of vegetable-producing areas in Burkina Faso. Environ. Int. 2018, 111, 317–331. [Google Scholar] [CrossRef]
- Lehmann, E.; Turrero, N.; Kolia, M.; Konaté, Y.; de Alencastro, L.F. Dietary risk assessment of pesticides from vegetables and drinking water in gardening areas in Burkina Faso. Sci. Total Environ. 2017, 601–602, 1208–1216. [Google Scholar] [CrossRef]
- Son, D.; Zerbo, F.K.B.; Bonzi, S.; Schiffers, B.; Somda, I.; Schiffers, B.; Legreve, A. Assessment of Tomato (Solanum lycopersicum L.) Producers’ Exposure Level to Pesticides, in Kouka and Toussiana (Burkina Faso). Int. J. Environ. Res. Public Health 2018, 15, 204. [Google Scholar] [CrossRef]
- Diaz, M.H.; Waller, J.L.; Napoliello, R.A.; Islam, S.; Wolff, B.J.; Burken, D.J.; Holden, R.L.; Srinivasan, V.; Arvay, M.; McGee, L.; et al. Optimization of Multiple Pathogen Detection Using the TaqMan Array Card: Application for a Population-Based Study of Neonatal Infection. PLoS ONE 2013, 8, e66183. [Google Scholar] [CrossRef]
- Diaz, M.H.; Waller, J.L.; Theodore, M.J.; Patel, N.; Wolff, B.J.; Benitez, A.J.; Morris, T.; Raghunathan, P.L.; Breiman, R.F.; Whitney, C.G.; et al. Development and Implementation of Multiplex TaqMan Array Cards for Specimen Testing at Child Health and Mortality Prevention Surveillance Site Laboratories. Clin. Infect. Dis. 2019, 69 (Suppl. 4), S311–S321. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, G.A.; Schnabel, K.C.; Erdman, D.D.; Prill, M.M.; Iwane, M.K.; Shelley, L.M.; Whitaker, B.L.; Szilagyi, P.G.; Hall, C.B. Field evaluation of TaqMan Array Card (TAC) for the simultaneous detection of multiple respiratory viruses in children with acute respiratory infection. J. Clin. Virol. 2013, 57, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Jukic, A.; Bakiri, L.; Wagner, E.F.; Tilg, H.; Adolph, T.E. Calprotectin: From biomarker to biological function. Gut 2021, 70, 1978–1988. [Google Scholar] [CrossRef] [PubMed]
- Yui, S.; Nakatani, Y.; Mikami, M. Calprotectin (S100A8/S100A9), an Inflammatory Protein Complex from Neutrophils with a Broad Apoptosis-Inducing Activity. Biol. Pharm. Bull. 2003, 26, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Dale, I.; Brandtzaeg, P.; Fagerhol, M.K.; Scott, H. Distribution of a New Myelomonocytic Antigen (L1) in Human Peripheral Blood Leukocytes: Immunofluorescence and Immunoperoxidase Staining Features in Comparison with Lysozyme and Lactoferrin. Am. J. Clin. Pathol. 1985, 84, 24–34. [Google Scholar] [CrossRef]
- Summerton, C.B.; Longlands, M.G.; Wiener, K.; Shreeve, D.R. Faecal calprotectin: A marker of inflammation throughout the intestinal tract. Eur. J. Gastroenterol. Hepatol. 2002, 14, 841–845. [Google Scholar] [CrossRef]
- Cummings, J.; Macfarlane, G. Collaborative JPEN-Clinical Nutrition Scientific Publications Role of intestinal bacteria in nutrient metabolism. J. Parenter. Enter. Nutr. 1997, 21, 357–365. [Google Scholar] [CrossRef]
- Blad, C.C.; Tang, C.; Offermanns, S. G protein-coupled receptors for energy metabolites as new therapeutic targets. Nat. Rev. Drug Discov. 2012, 11, 603–619. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Diversity, metabolism and microbial ecology of butyrate-producing bacteria from the human large intestine. FEMS Microbiol. Lett. 2009, 294, 1–8. [Google Scholar] [CrossRef]
- Berries, G.; Mnf, W.; Kang, Y.; Yang, G.; Zhang, S.; Ross, C.F.; Zhu, M.-J. Goji Berry Modulates Gut Microbiota and Alleviates Colitis in IL-10-Deficient Mice. Mol. Nutr. Food Res. 2018, 62, e1800535. [Google Scholar] [CrossRef]
- Gomes, S.D.; Oliveira, C.S.; Azevedo-Silva, J.; Casanova, M.R.; Barreto, J.; Pereira, H.; Chaves, S.R.; Rodrigues, L.R.; Casal, M.; Côrte-Real, M.; et al. The Role of Diet Related Short-Chain Fatty Acids in Colorectal Cancer Metabolism and Survival: Prevention and Therapeutic Implications. Curr. Med. Chem. 2020, 27, 4087–4108. [Google Scholar] [CrossRef] [PubMed]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in Gut Microbiota Control Metabolic Endotoxemia-Induced Inflammation in High-Fat Diet-Induced Obesity and Diabetes in Mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef] [PubMed]
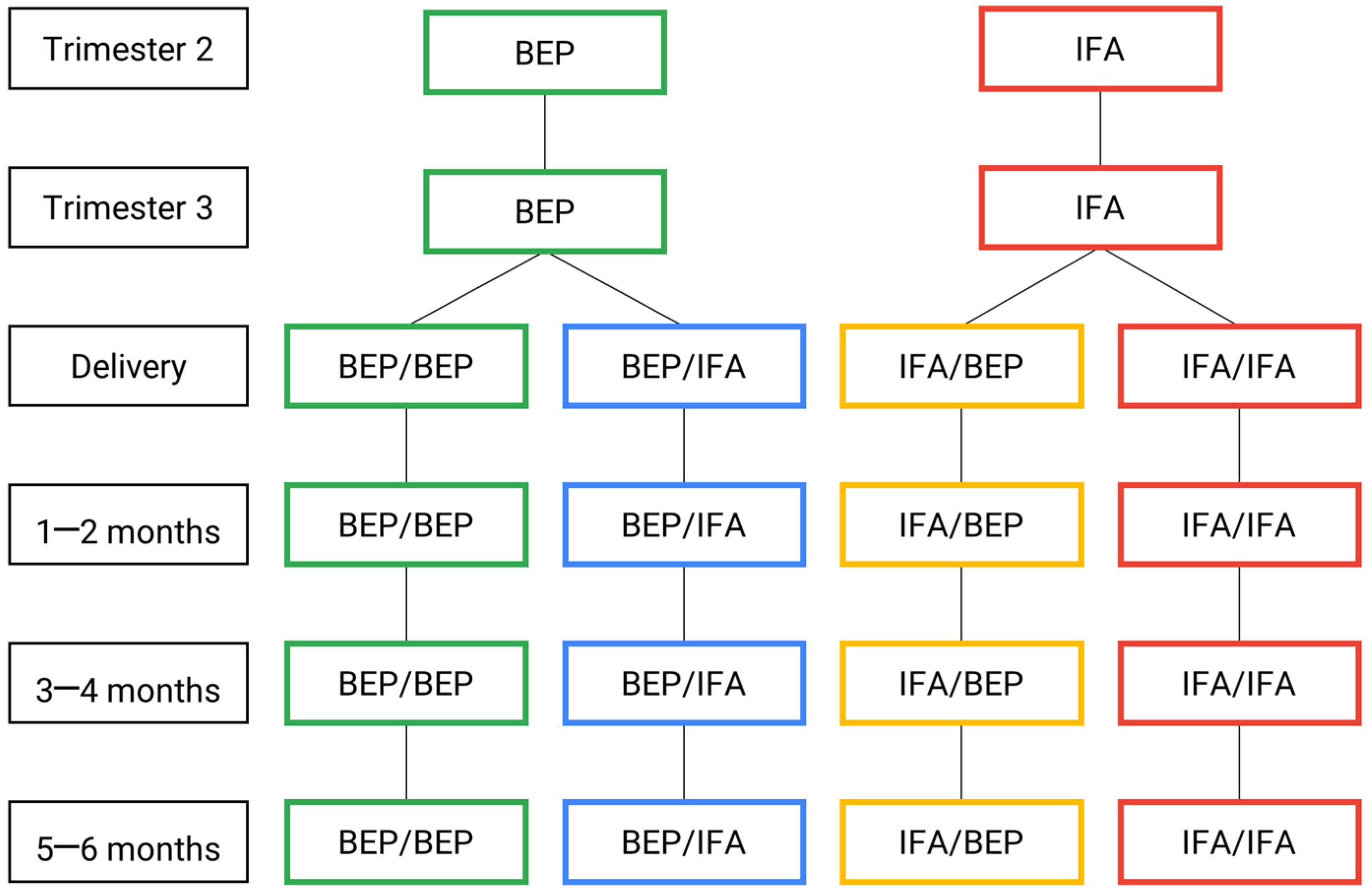
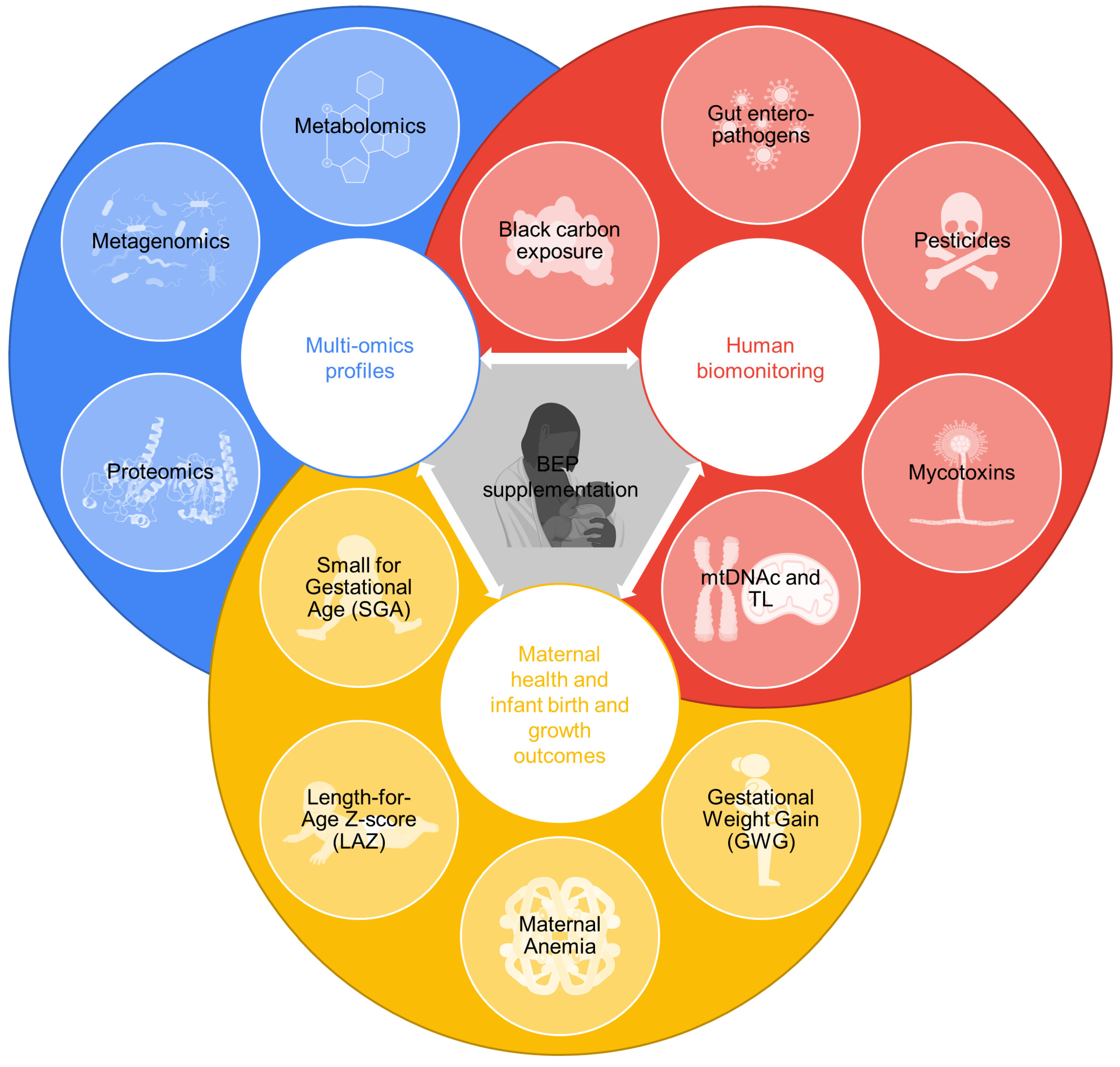

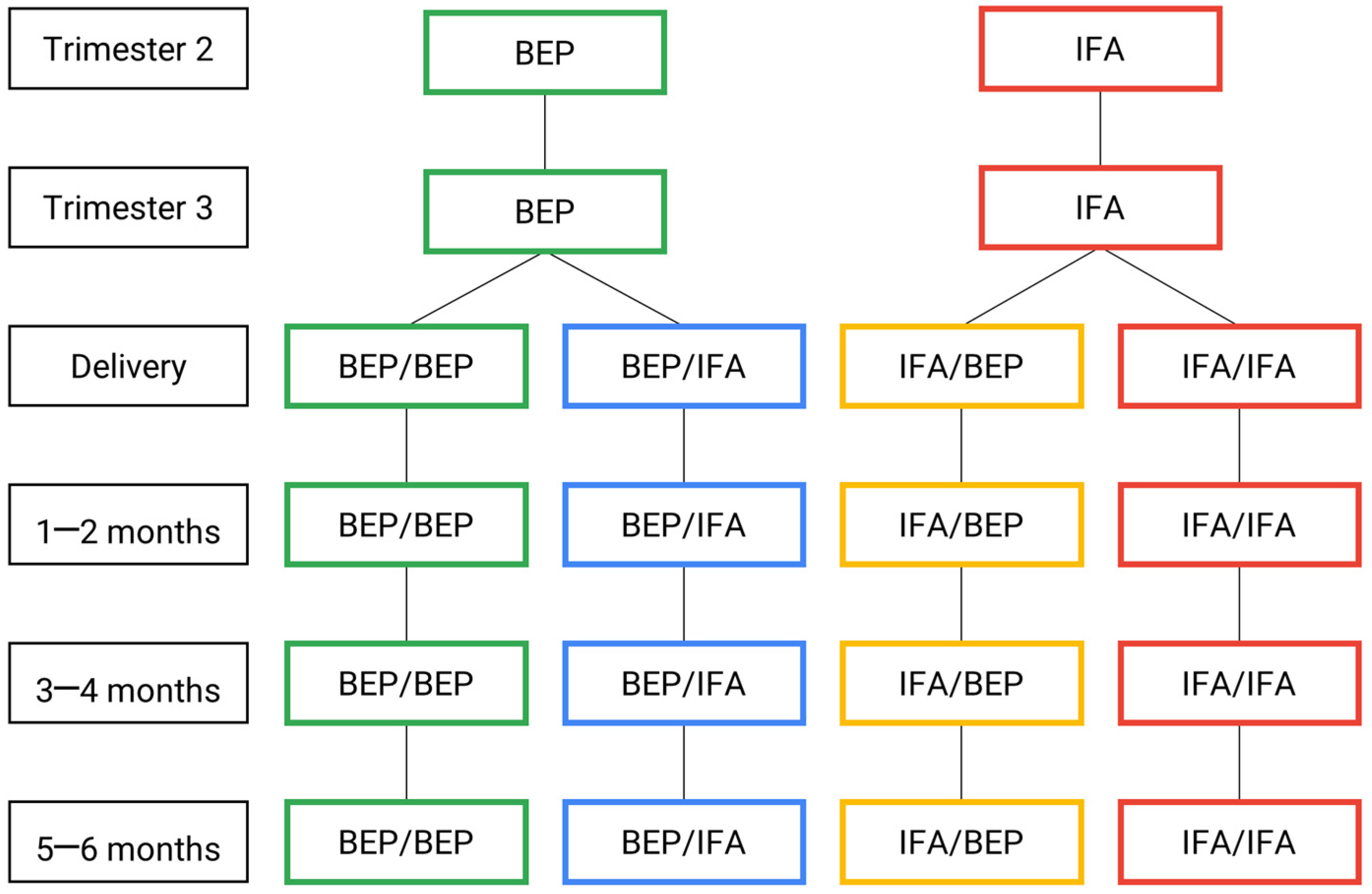{kind=link}
{kind=link}
| tri2 | tri3 | Birth | pn0.5–0.75 | pn12 | pn34 | pn56 | |
|---|---|---|---|---|---|---|---|
| 1. Plasma (metabolome and proteome) | M | M | M | M | |||
| 2. Whole blood (exposome and metabolome) | M | M | I | I | I | D | |
| 3. Cord blood (relative TL, mtDNAc and black carbon) | I | ||||||
| 4. Breast milk (metabolome, metagenomics and proteome) | M | M | M | ||||
| 5. Urine (exposome) | M | M | |||||
| 6. Feces (calprotectin, SCFAs and TAC analysis) | M | M | D | I | D |
| Mean for 72 g (Serving Size) | |
|---|---|
| Total energy (kcal) | 393 |
| Lipids (g) | 26 |
| Linoleic acid (g) | 3.9 |
| α-Linoleic acid (g) | 1.3 |
| Proteins (g) | 14.5 |
| Carbohydrates (g) | 23.3 |
| Calcium (mg) | 500 |
| Copper (mg) | 1.3 |
| Phosphorus (mg) | 418 |
| Iodine (µg) | 250 |
| Iron (mg) | 22 |
| Selenium (µg) | 65 |
| Manganese (mg) | 2.1 |
| Magnesium (mg) | 73 |
| Potassium (mg) | 562 |
| Zinc (mg) | 15 |
| Vitamin A (µg RE) 2 | 770 |
| Thiamin (mg) | 1.4 |
| Riboflavin (mg) | 1.4 |
| Niacin (mg) | 15 |
| Vitamin B5 (mg) | 7 |
| Vitamin B6 (mg) | 1.9 |
| Folic acid (µg) | 400 |
| Vitamin B12 (µg) | 2.6 |
| Vitamin C (mg) | 100 |
| Vitamin D (µg cholecalciferol) 3 | 15 |
| Vitamin E (mg α-tocopherol) 4 | 18 |
| Vitamin K (µg) | 72 |
| Analysis | Sample Matrix | Volume Collected | Analysis | Analytical Technique | Laboratory | Methodology |
|---|---|---|---|---|---|---|
| Metabolome | Capillary whole blood | 10 μL via VAMS | Untargeted | rLC-MS | Sapient Bioanalytics, California, United States | [42] (Villar et al., 2022) |
| Microbiome profile | Feces | 1.8 mL | Untargeted | Quantitative shotgun metagenomics | Stanford University, Stanford, California, United States | [43] (Olm et al., 2021) |
| KofamKOALA | [44] (Aramaki et al., 2020) | |||||
| Proteomics | Plasma | 100 μL | Untargeted | LC-MS/MS | Cedars-Sinai Medical Center, California, United States | [45] (Mc Ardle et al., 2022) |
| Analysis | Sample Matrix | Volume Collected | Analysis | Analytical Technique | Laboratory | Methodology |
|---|---|---|---|---|---|---|
| Macronutrients and fat-soluble vitamins | Breast milk | 7.2 mL | Untargeted | NIR analysis | University of California Davis, United States | [46] (Smilowitz et al., 2014) |
| LC-MS/MS | [47] (Hampel, Dror and Allen, 2018) | |||||
| Water-soluble vitamins | LC-MS/MS | [47] (Hampel, Dror and Allen, 2018) | ||||
| UPLC-MS/MS | [48] (Hampel, York and Allen, 2012) | |||||
| Automated immunoassay | [47] (Hampel, Dror and Allen, 2018) | |||||
| Minerals | ICP-MS | [47] (Hampel, Dror and Allen, 2018) | ||||
| HMOs | LC-MS/MS | Bode Lab, University of California San Diego, United States | [49] (Kellman et al., 2022) | |||
| Proteins | ECL | [50] (Ju, Lai and Yan, 2017) | ||||
| Metabolomics | rLC-MS | Sapient Bioanalytics, United States | [42] (Villar et al., 2022) | |||
| Proteomics | LC-MS/MS | Precision Biomarker Laboratories, United States | [45] (Mc Ardle et al., 2022) | |||
| Microbiome | 16S rRNA | Baylor College of Medicine, Alkek Center for Metagenomics and Microbiome Research, United States | [51] (Ramani et al., 2018) | |||
| Metabolomics (small molecules) | Targeted | LC-MS/MS | Biocrates, Innsbruck, Austria | [52] (Langsdorf et al., 2023) | ||
| Metabolomics (lipids and hexoses) | Targeted | FIA-MS/MS | Biocrates, Innsbruck, Austria | [52] (Langsdorf et al., 2023) |
| Analysis | Sample Matrix | Volume Collected | Analysis | Analytical Technique | Laboratory | Methodology |
|---|---|---|---|---|---|---|
| Telomere length and mitochondrial DNA content | Whole arterial blood from umbilical cord | 200 μL | Targeted | Real-time PCR method | Centre for Environmental Sciences, Hasselt University, Belgium | [68,69,70,71,72] (Cawthon, 2002, 2009; Janssen et al., 2012; Martens et al., 2016, 2020) |
| Black carbon particles | Whole arterial blood from umbilical cord | 250 μL | Targeted | Confocal microscopy | Centre for Environmental Sciences, Hasselt University, Belgium | [73,74] (Saenen et al., 2017; Bové et al., 2019) |
| Mycotoxins | Capillary whole blood | 20 μL VAMS | Targeted | LC-MS/MS | Centre of Excellence in Mycotoxicology and Public Health, Faculty of Pharmaceutical Sciences, Ghent University, Belgium | [23] (Vidal et al., 2021) |
| Pesticides | Urine | 4 mL | Targeted | LC-MS/MS | Toxicological Centre, University of Antwerp, Belgium | [75,76] (Gys et al., 2020; Caballero-Casero et al., 2021) |
| New/emerging contaminants | Untargeted | LC-HRMS | ||||
| Multiple Infection Targets | Feces | 1.8 mL | Targeted | TAC analysis | Institut de Recherche en Sciences de la Santé, Bobo-Dioulasso, Burkina Faso | [77,78] (Liu et al., 2014; Deboer et al., 2018) |
| Calprotectin | Feces | 1.8 mL | Targeted | ELISA | Institut de Recherche en Sciences de la Santé, Bobo-Dioulasso, Burkina Faso | [79] (Whitehead et al., 2013) |
| Short-chain fatty acids | Feces | 1.8 mL | Targeted | GC-FID | Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium | [80] (Toe et al., 2020) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bastos-Moreira, Y.; Ouédraogo, L.; De Boevre, M.; Argaw, A.; de Kok, B.; Hanley-Cook, G.T.; Deng, L.; Ouédraogo, M.; Compaoré, A.; Tesfamariam, K.; et al. A Multi-Omics and Human Biomonitoring Approach to Assessing the Effectiveness of Fortified Balanced Energy–Protein Supplementation on Maternal and Newborn Health in Burkina Faso: A Study Protocol. Nutrients 2023, 15, 4056. https://doi.org/10.3390/nu15184056
Bastos-Moreira Y, Ouédraogo L, De Boevre M, Argaw A, de Kok B, Hanley-Cook GT, Deng L, Ouédraogo M, Compaoré A, Tesfamariam K, et al. A Multi-Omics and Human Biomonitoring Approach to Assessing the Effectiveness of Fortified Balanced Energy–Protein Supplementation on Maternal and Newborn Health in Burkina Faso: A Study Protocol. Nutrients. 2023; 15(18):4056. https://doi.org/10.3390/nu15184056
Chicago/Turabian StyleBastos-Moreira, Yuri, Lionel Ouédraogo, Marthe De Boevre, Alemayehu Argaw, Brenda de Kok, Giles T. Hanley-Cook, Lishi Deng, Moctar Ouédraogo, Anderson Compaoré, Kokeb Tesfamariam, and et al. 2023. "A Multi-Omics and Human Biomonitoring Approach to Assessing the Effectiveness of Fortified Balanced Energy–Protein Supplementation on Maternal and Newborn Health in Burkina Faso: A Study Protocol" Nutrients 15, no. 18: 4056. https://doi.org/10.3390/nu15184056
APA StyleBastos-Moreira, Y., Ouédraogo, L., De Boevre, M., Argaw, A., de Kok, B., Hanley-Cook, G. T., Deng, L., Ouédraogo, M., Compaoré, A., Tesfamariam, K., Ganaba, R., Huybregts, L., Toe, L. C., Lachat, C., Kolsteren, P., De Saeger, S., & Dailey-Chwalibóg, T. (2023). A Multi-Omics and Human Biomonitoring Approach to Assessing the Effectiveness of Fortified Balanced Energy–Protein Supplementation on Maternal and Newborn Health in Burkina Faso: A Study Protocol. Nutrients, 15(18), 4056. https://doi.org/10.3390/nu15184056