
Nutritional Composition and Anti-Type 2 Diabetes Mellitus Potential of Femur Bone Extracts from Bovine, Chicken, Sheep, and Goat: Phytochemical and In Vivo Studies

 , , , , ,
, , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of the Bone Extract
2.3. Proximate Analysis
2.4. FAME Preparation
2.5. FAME GC-MS Analysis
2.6. Analysis of Amino Acids
2.7. Mineral Analysis
2.8. Animals
2.8.1. Animal Ethical Statement
2.8.2. Acute Toxicity Study
2.8.3. In Vitro DPPH Radical Scavenging Activity Assay of the Investigated Bone Extracts
2.8.4. Induction of T2DM and Experimental Design
2.8.5. Sample Preparation
2.8.6. Estimation of Blood Glucose Profiles Parameters
2.8.7. Estimation of Lipid Profile Parameters
2.8.8. Estimation of Hepatic Functions Markers
2.8.9. Estimation of Kidney Functions Markers
2.8.10. Estimation of Proinflammatory Markers
2.8.11. Estimation of Oxidative Stress Markers
2.8.12. Histopathological Investigation
2.8.13. Histopathological Lesion Scoring
2.9. Statistical Analysis
3. Results
3.1. Proximate Composition
3.2. Fatty Acid Composition
3.3. Amino Acid Content
3.4. Mineral Contents
3.5. In Vitro DPPH Radical Scavenging Activity Assay of Bovine (12 Months), Chicken (4 Months), Sheep (13 Months), and Goat (9 Months) Femur Bone Extracts
3.6. Estimation of Blood Glucose Profiles Parameters
3.7. Estimation of Lipid Profiles Parameters
3.8. Estimation of Hepatic Functions Markers
3.9. Estimation of Kidney Functions Markers
3.10. Estimation of Proinflammatory Markers
3.11. Estimation of Oxidative Stress Markers
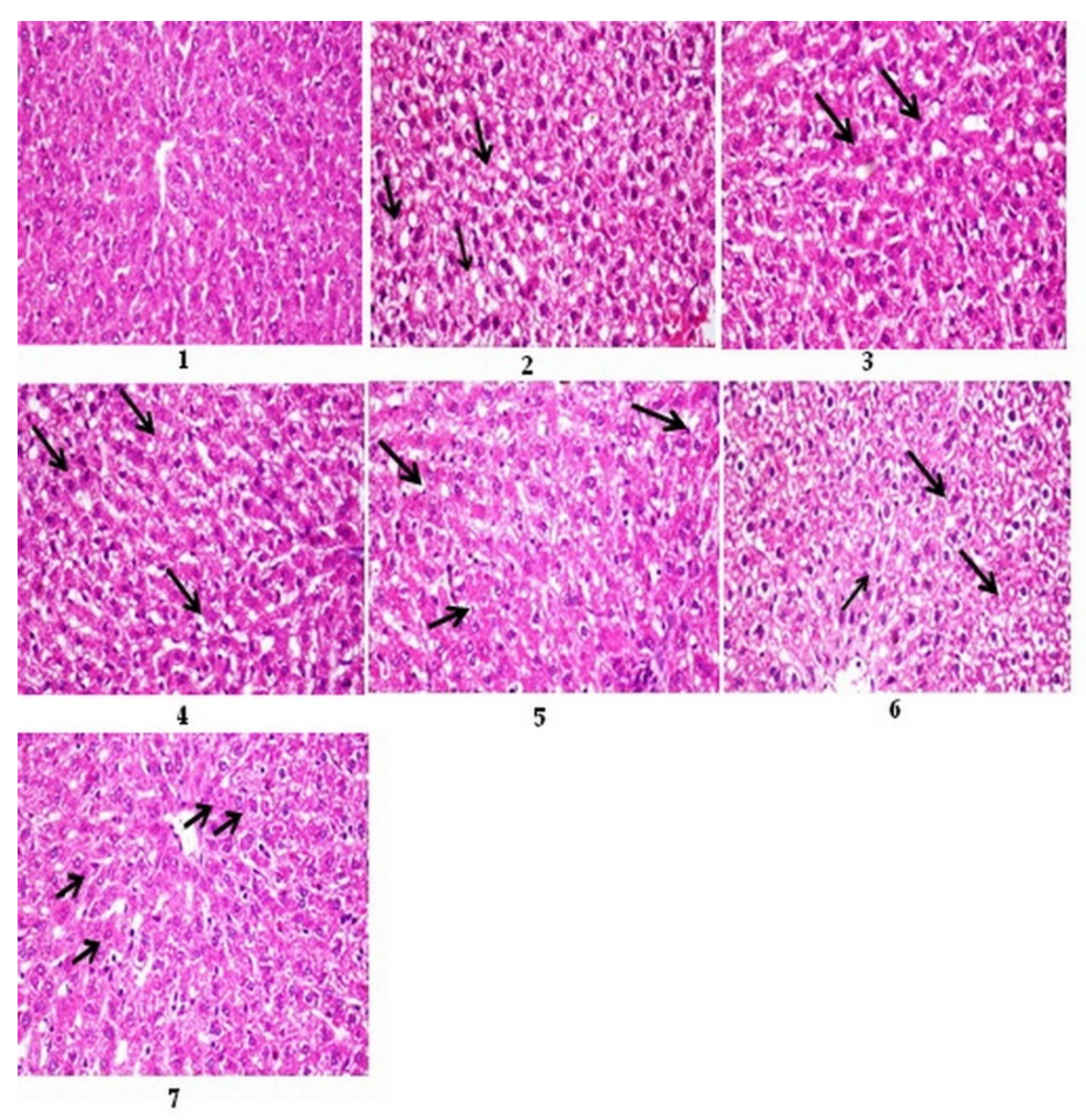
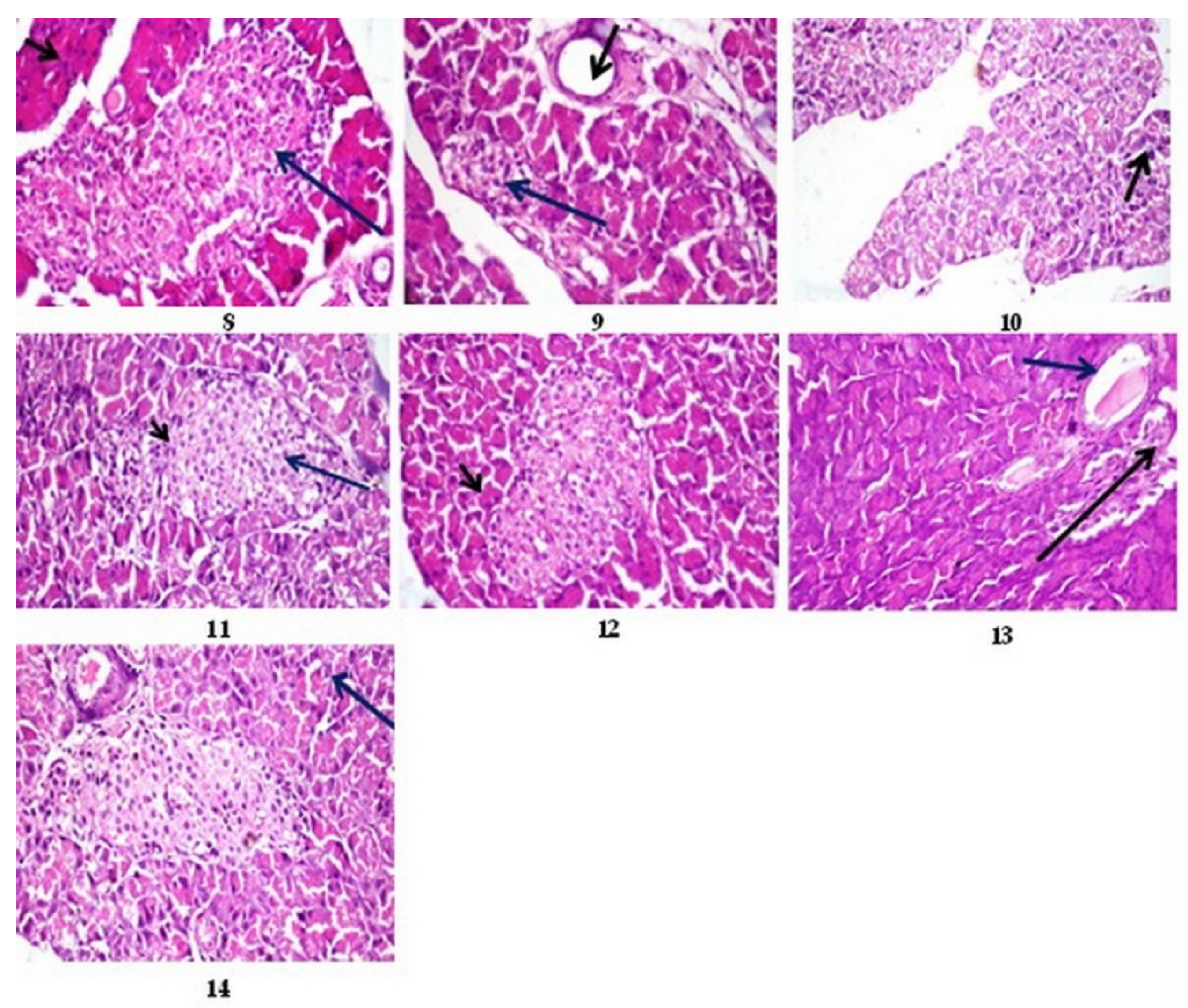
3.12. Histopathological Results
3.12.1. Histopathological Results of Liver
3.12.2. Histopathological Results of Pancreas
3.12.3. Histopathological Lesion Scoring
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
List of Abbreviations
References
- Chalamaiah, M.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Li, F.; Jia, D.; Yao, K. Amino acid composition and functional properties of collagen polypeptide from Yak (Bos grunniens) bone. LWT-Food Sci. Technol. 2009, 42, 945–949. [Google Scholar] [CrossRef]
- Reid, I.; Ibbertson, H. Calcium supplements in the prevention of steroid-induced osteoporosis. Am. J. Clin. Nutr. 1986, 44, 287–290. [Google Scholar] [CrossRef]
- Kim, D.; Kim, H.-J.; Chae, H.-S.; Park, N.-G.; Kim, Y.-B.; Jang, A. Anti-oxidation and anti-wrinkling effects of Jeju horse leg bone hydrolysates. Korean J. Food Sci. Anim. Resour. 2014, 34, 844. [Google Scholar] [CrossRef]
- Hopkins, A.B. Chicken soup cure may not be a myth. Nurse Pract. 2003, 28, 16. [Google Scholar]
- Saketkhoo, K.; Januszkiewicz, A.; Sackner, M.A. Effects of drinking hot water, cold water, and chicken soup on nasal mucus velocity and nasal airflow resistance. Chest 1978, 74, 408–410. [Google Scholar] [CrossRef]
- Monro, J.; Leon, R.; Puri, B. The risk of lead contamination in bone broth diets. Med. Hypotheses 2013, 80, 389–390. [Google Scholar] [CrossRef]
- Rosen, H.; Salemme, H.; Zeind, A.; Moses, A.; Shapiro, A.; Greenspan, S. Chicken soup revisited: Calcium content of soup increases with duration of cooking. Calcif. Tissue Int. 1994, 54, 486–488. [Google Scholar] [CrossRef]
- Roden, M.; Shulman, G.I. The integrative biology of type 2 diabetes. Nature 2019, 576, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Gæde, P.; Vedel, P.; Larsen, N.; Jensen, G.V.; Parving, H.-H.; Pedersen, O. Multifactorial intervention and cardiovascular disease in patients with type 2 diabetes. N. Engl. J. Med. 2003, 348, 383–393. [Google Scholar] [CrossRef] [PubMed]
- The Emerging Risk Factors Collaboration. Diabetes mellitus, fasting blood glucose concentration, and risk of vascular disease: A collaborative meta-analysis of 102 prospective studies. Lancet 2010, 375, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.; D’Alonzo, D.; De Fenza, M.; De Gregorio, E.; Tamanini, A.; Lippi, G.; Dechecchi, M.C.; Guaragna, A. Synthesis and therapeutic applications of iminosugars in cystic fibrosis. Int. J. Mol. Sci. 2020, 21, 3353. [Google Scholar] [CrossRef] [PubMed]
- Yahaya, T.O.; Yusuf, A.B.; Danjuma, J.K.; Usman, B.M.; Ishiaku, Y.M. Mechanistic links between vitamin deficiencies and diabetes mellitus: A review. Egypt. J. Basic Appl. Sci. 2021, 8, 189–202. [Google Scholar] [CrossRef]
- Santos, R.K.F.; Brandão-Lima, P.N.; Tete, R.M.D.D.; Freire, A.R.S.; Pires, L.V. Vitamin D ratio and glycaemic control in individuals with type 2 diabetes mellitus: A systematic review. Diabetes/Metab. Res. Rev. 2018, 34, e2969. [Google Scholar] [CrossRef]
- Martini, L.A.; Catania, A.S.; Ferreira, S.R. Role of vitamins and minerals in prevention and management of type 2 diabetes mellitus. Nutr. Rev. 2010, 68, 341–354. [Google Scholar] [CrossRef]
- Hales, C.N.; Barker, D.J. The thrifty phenotype hypothesis: Type 2 diabetes. Br. Med. Bull. 2001, 60, 5–20. [Google Scholar] [CrossRef]
- Barker, D.J. The malnourished baby and infant: Relationship with Type 2 diabetes. Br. Med. Bull. 2001, 60, 69–88. [Google Scholar] [CrossRef][Green Version]
- Iacucci, M.; Ghosh, S. Looking beyond symptom relief: Evolution of mucosal healing in inflammatory bowel disease. Ther. Adv. Gastroenterol. 2011, 4, 129–143. [Google Scholar] [CrossRef]
- Papi, C.; Fascì-Spurio, F.; Rogai, F.; Settesoldi, A.; Margagnoni, G.; Annese, V. Mucosal healing in inflammatory bowel disease: Treatment efficacy and predictive factors. Dig. Liver Dis. 2013, 45, 978–985. [Google Scholar] [CrossRef]
- Górska-Warsewicz, H.; Laskowski, W.; Kulykovets, O.; Kudlińska-Chylak, A.; Czeczotko, M.; Rejman, K. Food products as sources of protein and amino acids—The case of Poland. Nutrients 2018, 10, 1977. [Google Scholar] [CrossRef]
- Choi, H.-G.; Choi, H.-S.; Choi, Y.-S.; Jung, M.-O.; Choi, J.-S.; Choi, Y.-I. Effects of mixed bone and brisket meat on physico-chemical characteristics of shank bone and rib extracts from Hanwoo. Korean J. Food Sci. Anim. Resour. 2016, 36, 61–67. [Google Scholar] [CrossRef]
- Hsu, D.-J.; Lee, C.-W.; Tsai, W.-C.; Chien, Y.-C. Essential and toxic metals in animal bone broths. Food Nutr. Res. 2017, 61, 1347478. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-J.; Kim, D.; Chae, H.-S.; Kim, N.-Y.; Jang, A. Nutritional composition in bone extracts from Jeju crossbred horses at different slaughter ages. Korean J. Food Sci. Anim. Resour. 2017, 37, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Young Seol, J.; Yoon, J.Y.; Jeong, H.S.; Joo, N.; Choi, S.Y. Anti-Aging Effects of the Hanwoo Leg Bone, Foot and Tail Infusions (HLI, HFI and HTI) on Skin Fibroblast. Korean J. Food Sci. Anim. Resour. 2016, 36, 237–243. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mar-Solís, L.M.; Soto-Domínguez, A.; Rodríguez-Tovar, L.E.; Rodríguez-Rocha, H.; García-García, A.; Aguirre-Arzola, V.E.; Zamora-Ávila, D.E.; Garza-Arredondo, A.J.; Castillo-Velázquez, U. Analysis of the Anti-Inflammatory Capacity of Bone Broth in a Murine Model of Ulcerative Colitis. Medicina 2021, 57, 1138. [Google Scholar] [CrossRef]
- Ahmed, W.M.; Ibrahim, M.A.; Helmy, N.A.; ElKashlan, A.M.; Elmaidomy, A.H.; Zaki, A.R. Amelioration of aluminum-induced hepatic and nephrotoxicity by Premna odorata extract is mediated by lowering MMP9 and TGF-β gene alterations in Wistar rat. Environ. Sci. Pollut. Res. 2022, 29, 72827–72838. [Google Scholar] [CrossRef]
- Bremner, J. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Alsenani, F.; Ashour, A.M.; Alzubaidi, M.A.; Azmy, A.F.; Hetta, M.H.; Abu-Baih, D.H.; Elrehany, M.A.; Zayed, A.; Sayed, A.M.; Abdelmohsen, U.R. Wound Healing Metabolites from Peters’ Elephant-Nose Fish Oil: An In Vivo Investigation Supported by In Vitro and In Silico Studies. Mar. Drugs 2021, 19, 605. [Google Scholar] [CrossRef]
- Al-Warhi, T.; Elmaidomy, A.H.; Selim, S.; Al-Sanea, M.M.; Albqmi, M.; Mostafa, E.M.; Ibrahim, S.; Ghoneim, M.M.; Sayed, A.M.; Abdelmohsen, U.R. Bioactive Phytochemicals of Citrus reticulata Seeds—An Example of Waste Product Rich in Healthy Skin Promoting Agents. Antioxidants 2022, 11, 984. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Mohamed, E.M.; Aly, H.F.; Younis, E.A.; Shams, S.G.E.; Altemani, F.H.; Alzubaidi, M.A.; Almaghrabi, M.; Harbi, A.A.; Alsenani, F. Anti-Inflammatory and Antioxidant Properties of Malapterurus electricus Skin Fish Methanolic Extract in Arthritic Rats: Therapeutic and Protective Effects. Mar. Drugs 2022, 20, 639. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Zahran, E.M.; Soltane, R.; Alasiri, A.; Saber, H.; Ngwa, C.J.; Pradel, G.; Alsenani, F.; Sayed, A.M.; Abdelmohsen, U.R. New Halogenated Compounds from Halimeda macroloba Seaweed with Potential Inhibitory Activity against Malaria. Molecules 2022, 27, 5617. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Mohammed, R.; Hassan, H.M.; Owis, A.I.; Rateb, M.E.; Khanfar, M.A.; Krischke, M.; Mueller, M.J.; Ramadan Abdelmohsen, U. Metabolomic profiling and cytotoxic tetrahydrofurofuran lignans investigations from Premna odorata Blanco. Metabolites 2019, 9, 223. [Google Scholar] [CrossRef]
- Bulska, E.; Ruszczyńska, A. Analytical techniques for trace element determination. Phys. Sci. Rev. 2017, 2, 20178002. [Google Scholar] [CrossRef]
- Andrade Korn, M.D.G.; da Boa Morte, E.S.; Batista dos Santos, D.C.M.; Castro, J.T.; Barbosa, J.T.P.; Teixeira, A.P.; Fernandes, A.P.; Welz, B.; dos Santos, W.P.C.; Nunes dos Santos, E.B.G. Sample preparation for the determination of metals in food samples using spectroanalytical methods—A review. Appl. Spectrosc. Rev. 2008, 43, 67–92. [Google Scholar] [CrossRef]
- Choo, C.; Sulong, N.; Man, F.; Wong, T. Vitexin and isovitexin from the leaves of Ficus deltoidea with in-vivo α-glucosidase inhibition. J. Ethnopharmacol. 2012, 142, 776–781. [Google Scholar] [CrossRef]
- Biswas, K.; Azad, A.; Sultana, T.; Khan, F.; Hossain, S.; Alam, S.; Chowdhary, R.; Khatun, Y. Assessment of in-vitro cholinesterase inhibitory and thrombolytic potential of bark and seed extracts of Tamarindus indica (L.) relevant to the treatment of Alzheimer’s disease and clotting disorders. J. Intercult. Ethnopharmacol. 2017, 6, 115–120. [Google Scholar] [CrossRef]
- Islam, M.S.; Choi, H. Effects of dietary onion (Allium cepa L.) in a high-fat diet streptozotocin-induced diabetes rodent model. Ann. Nutr. Metab. 2008, 53, 6–12. [Google Scholar] [CrossRef]
- Hayashi, K.; Kojima, R.; Ito, M. Strain differences in the diabetogenic activity of streptozotocin in mice. Biol. Pharm. Bull. 2006, 29, 1110–1119. [Google Scholar] [CrossRef]
- Mandour, M.; Al-Shami, S.; Al-Eknah, M.; Hussein, Y.; El-Ashmawy, I. The acute and long-term safety evaluation of aqueous, methanolic and ethanolic extracts of Achillea fragrantissima. Afr. J. Pharm. Pharmacol. 2013, 7, 2282–2290. [Google Scholar] [CrossRef]
- Richmond, W. Preparation and properties of a cholesterol oxidase from Nocardia sp. and its application to the enzymatic assay of total cholesterol in serum. Clin. Chem. 1973, 19, 1350–1356. [Google Scholar] [CrossRef]
- Wieland, H.; Seidel, D. A simple specific method for precipitation of low density lipoproteins. J. Lipid Res. 1983, 24, 904–909. [Google Scholar] [CrossRef]
- Burstein, M.; Scholnick, H.; Morfin, R. Rapid method for the isolation of lipoproteins from human serum by precipitation with polyanions. J. Lipid Res. 1970, 11, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Prencipe, L. Serum triglycerides determined colorimetrically with an enzyme that produces hydrogen peroxide. Clin. Chem. 1982, 28, 2077–2080. [Google Scholar] [CrossRef]
- Connerty, H.V.; Briggs, A.R.; Eaton Jr, E.H. Simplified determination of the lipid components of blood serum. Clin. Chem. 1961, 7, 37–53. [Google Scholar] [CrossRef]
- Elmaidomy, A.H.; Alhadrami, H.A.; Amin, E.; Aly, H.F.; Othman, A.M.; Rateb, M.E.; Hetta, M.H.; Abdelmohsen, U.R.; Hassan, H.M. Anti-inflammatory and antioxidant activities of terpene-and polyphenol-rich Premna odorata leaves on alcohol-inflamed female wistar albino rat liver. Molecules 2020, 25, 3116. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Gustafsson, U.; Scott, M.; Hubner, M.; Nygren, J.; Demartines, N.; Francis, N.; Rockall, T.; Young-Fadok, T.; Hill, A.; Soop, M. Guidelines for perioperative care in elective colorectal surgery: Enhanced Recovery After Surgery (ERAS®) Society recommendations: 2018. World J. Surg. 2019, 43, 659–695. [Google Scholar] [CrossRef]
- Bartels, H.; Böhmer, M.; Heierli, C. Serum kreatininbestimmung ohne enteiweissen. Clin. Chim. Acta 1972, 37, 193–197. [Google Scholar] [CrossRef]
- Fawcett, J.; Scott, J. A rapid and precise method for the determination of urea. J. Clin. Pathol. 1960, 13, 156–159. [Google Scholar] [CrossRef]
- Banchroft, J.; Stevens, A.; Turner, D. Theory and Practice of Histological Techniques; Churchil Livingstone: London, UK, 1996. [Google Scholar]
- Alnusaire, T.S.; Sayed, A.M.; Elmaidomy, A.H.; Al-Sanea, M.M.; Albogami, S.; Albqmi, M.; Alowaiesh, B.F.; Mostafa, E.M.; Musa, A.; Youssif, K.A. An In Vitro and In Silico Study of the Enhanced Antiproliferative and Pro-Oxidant Potential of Olea europaea L. cv. Arbosana Leaf Extract via Elastic Nanovesicles (Spanlastics). Antioxidants 2021, 10, 1860. [Google Scholar] [CrossRef] [PubMed]
- Korany, R.M.; Ahmed, K.S.; Halawany, H.; Ahmed, K.A. Effect of long-term arsenic exposure on female Albino rats with special reference to the protective role of Spirulina platensis. Explor. Anim. Med. Res. 2019, 9, 125–136. [Google Scholar]
- Briggs, M.A.; Petersen, K.S.; Kris-Etherton, P.M. Saturated fatty acids and cardiovascular disease: Replacements for saturated fat to reduce cardiovascular risk. Healthcare 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Shetty, K. Role of proline-linked pentose phosphate pathway in biosynthesis of plant phenolics for functional food and environmental applications: A review. Process Biochem. 2004, 39, 789–804. [Google Scholar] [CrossRef]
- Ahmadieh, H.; Azar, S.T. Liver disease and diabetes: Association, pathophysiology, and management. Diabetes Res. Clin. Pract. 2014, 104, 53–62. [Google Scholar] [CrossRef]
- Mohamed, J.; Nafizah, A.N.; Zariyantey, A.; Budin, S. Mechanisms of diabetes-induced liver damage: The role of oxidative stress and inflammation. Sultan Qaboos Univ. Med. J. 2016, 16, e132. [Google Scholar] [CrossRef]
- Stumvoll, M.; Goldstein, B.J.; Van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Weyer, C.; Bogardus, C.; Mott, D.M.; Pratley, R.E. The natural history of insulin secretory dysfunction and insulin resistance in the pathogenesis of type 2 diabetes mellitus. J. Clin. Investig. 1999, 104, 787–794. [Google Scholar] [CrossRef]
- Sudesna, C.; Khunti, K.; Davies, M.J. Type 2 diabetes. Lancet 2017, 389, 2239–2251. [Google Scholar] [CrossRef]
- Collaboration, N.R.F.; Zhou, B.; Lu, Y.; Hajifathalian, K.; Bentham, J.; Di Cesare, M. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4.4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef]
- Fronzo, R. From the triumvirate to the ominous octet: A new paradigm for the treatment of type 2 diabetes mellitus (banting lecture). Diabetes 2009, 58, 773–795. [Google Scholar] [CrossRef]
- Schwartz, S.S.; Epstein, S.; Corkey, B.E.; Grant, S.F.; Gavin III, J.R.; Aguilar, R.B. The time is right for a new classification system for diabetes: Rationale and implications of the β-cell–centric classification schema. Diabetes Care 2016, 39, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Itoh, Y.; Kawamata, Y.; Harada, M.; Kobayashi, M.; Fujii, R.; Fukusumi, S.; Ogi, K.; Hosoya, M.; Tanaka, Y.; Uejima, H. Free fatty acids regulate insulin secretion from pancreatic β cells through GPR40. Nature 2003, 422, 173–176. [Google Scholar] [CrossRef] [PubMed]
- Yaney, G.; Corkey, B. Fatty acid metabolism and insulin secretion in pancreatic beta cells. Diabetologia 2003, 46, 1297–1312. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Chi, Y.; Burkhardt, B.R.; Guan, Y.; Wolf, B.A. Leucine metabolism in regulation of insulin secretion from pancreatic beta cells. Nutr. Rev. 2010, 68, 270–279. [Google Scholar] [CrossRef]
- Wiederkehr, A.; Wollheim, C.B. Mitochondrial signals drive insulin secretion in the pancreatic β-cell. Mol. Cell. Endocrinol. 2012, 353, 128–137. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Liu, D. Regulation of insulin synthesis and secretion and pancreatic Beta-cell dysfunction in diabetes. Curr. Diabetes Rev. 2013, 9, 25–53. [Google Scholar] [CrossRef]
- Porzio, O.; Federici, M.; Hribal, M.L.; Lauro, D.; Accili, D.; Lauro, R.; Borboni, P.; Sesti, G. The Gly 972 → Arg amino acid polymorphism in IRS-1 impairs insulin secretion in pancreatic β cells. J. Clin. Investig. 1999, 104, 357–364. [Google Scholar] [CrossRef]
- Meloni, A.; DeYoung, M.; Lowe, C.; Parkes, D. GLP—1 receptor activated insulin secretion from pancreatic β—Cells: Mechanism and glucose dependence. Diabetes Obes. Metab. 2013, 15, 15–27. [Google Scholar] [CrossRef]
- Srinivasan, V.; Radhakrishnan, S.; Angayarkanni, N.; Sulochana, K. Antidiabetic effect of free amino acids supplementation in human visceral adipocytes through adiponectin-dependent mechanism. Indian J. Med. Res. 2019, 149, 41–46. [Google Scholar] [CrossRef]
- Gilon, P.; Chae, H.-Y.; Rutter, G.A.; Ravier, M.A. Calcium signaling in pancreatic β-cells in health and in Type 2 diabetes. Cell Calcium 2014, 56, 340–361. [Google Scholar] [CrossRef] [PubMed]
- Wiederkehr, A.; Wollheim, C.B. Impact of mitochondrial calcium on the coupling of metabolism to insulin secretion in the pancreatic β-cell. Cell Calcium 2008, 44, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Beck, A.; Launay, P.; Gross, S.A.; Stokes, A.J.; Kinet, J.-P.; Fleig, A.; Penner, R. TRPM4 controls insulin secretion in pancreatic β-cells. Cell Calcium 2007, 41, 51–61. [Google Scholar] [CrossRef]
- Palanivel, R.; Veluthakal, R.; McDonald, P.; Kowluru, A. Further evidence for the regulation of acetyl-CoA carboxylase activity by a glutamate-and magnesium-activated protein phosphatase in the pancreatic β cell. Endocrine 2005, 26, 71–77. [Google Scholar] [CrossRef]
- Kostov, K. Effects of magnesium deficiency on mechanisms of insulin resistance in type 2 diabetes: Focusing on the processes of insulin secretion and signaling. Int. J. Mol. Sci. 2019, 20, 1351. [Google Scholar] [CrossRef]
- Xu, H.; Du, X.; Xu, J.; Zhang, Y.; Tian, Y.; Liu, G.; Wang, X.; Ma, M.; Du, W.; Liu, Y. Pancreatic β cell microRNA-26a alleviates type 2 diabetes by improving peripheral insulin sensitivity and preserving β cell function. PLoS Biol. 2020, 18, e3000603. [Google Scholar] [CrossRef]
- Liu, Y.; Batchuluun, B.; Ho, L.; Zhu, D.; Prentice, K.J.; Bhattacharjee, A.; Zhang, M.; Pourasgari, F.; Hardy, A.B.; Taylor, K.M. Characterization of zinc influx transporters (ZIPs) in pancreatic β cells: Roles in regulating cytosolic zinc homeostasis and insulin secretion. J. Biol. Chem. 2015, 290, 18757–18769. [Google Scholar] [CrossRef]
- Brandão-Lima, P.N.; de Carvalho, G.B.; Santos, R.K.F.; da Cruz Santos, B.; Dias-Vasconcelos, N.L.; de Sousa Rocha, V.; Barbosa, K.B.F.; Pires, L.V. Intakes of zinc, potassium, calcium, and magnesium of individuals with type 2 diabetes mellitus and the relationship with glycemic control. Nutrients 2018, 10, 1948. [Google Scholar] [CrossRef]
- Bonfanti, D.H.; Alcazar, L.P.; Arakaki, P.A.; Martins, L.T.; Agustini, B.C.; de Moraes Rego, F.G.; Frigeri, H.R. ATP-dependent potassium channels and type 2 diabetes mellitus. Clin. Biochem. 2015, 48, 476–482. [Google Scholar] [CrossRef]
- Veronese, N.; Watutantrige-Fernando, S.; Luchini, C.; Solmi, M.; Sartore, G.; Sergi, G.; Manzato, E.; Barbagallo, M.; Maggi, S.; Stubbs, B. Effect of magnesium supplementation on glucose metabolism in people with or at risk of diabetes: A systematic review and meta-analysis of double-blind randomized controlled trials. Eur. J. Clin. Nutr. 2016, 70, 1354–1359. [Google Scholar] [CrossRef] [PubMed]
- Basyouni, W.M.; Abbas, S.Y.; El-Bayouki, K.A.; Younis, E.A.; Ali, S.A.; Aly, H.F. Synthesis and hyperglycemic, biochemical and histopathological evaluation of novel sulfonylbiguanide and sulfonylurea derivatives as potent anti-diabetic agents. Bioorg. Chem. 2021, 117, 105418. [Google Scholar] [CrossRef] [PubMed]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vasc. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef]
- Nair, H.B.; Sung, B.; Yadav, V.R.; Kannappan, R.; Chaturvedi, M.M.; Aggarwal, B.B. Delivery of antiinflammatory nutraceuticals by nanoparticles for the prevention and treatment of cancer. Biochem. Pharmacol. 2010, 80, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Quraishi, I.; Collins, S.; Pestaner, J.P.; Harris, T.; Bagasra, O. Role of zinc and zinc transporters in the molecular pathogenesis of diabetes mellitus. Med. Hypotheses 2005, 65, 887–892. [Google Scholar] [CrossRef]
- Spee, B.; Mandigers, P.J.; Arends, B.; Bode, P.; van den Ingh, T.S.; Hoffmann, G.; Rothuizen, J.; Penning, L.C. Differential expression of copper-associated and oxidative stress related proteins in a new variant of copper toxicosis in Doberman pinschers. Comp. Hepatol. 2005, 4, 3. [Google Scholar] [CrossRef][Green Version]
- Guo, C.-H.; Wang, C.-L.; Chen, P.-C.; Yang, T.-C. Linkage of some trace elements, peripheral blood lymphocytes, inflammation, and oxidative stress in patients undergoing either hemodialysis or peritoneal dialysis. Perit. Dial. Int. 2011, 31, 583–591. [Google Scholar] [CrossRef]
- Foresti, R.; Motterlini, R. Heme oxygenase-1 in diabetic vascular dysfunction. Vasc. Pharmacol. 2014, 62, 132–133. [Google Scholar] [CrossRef]
- Papadopoulou-Marketou, N.; Kanaka-Gantenbein, C.; Marketos, N.; Chrousos, G.P.; Papassotiriou, I. Biomarkers of diabetic nephropathy: A 2017 update. Crit. Rev. Clin. Lab. Sci. 2017, 54, 326–342. [Google Scholar] [CrossRef]
- Haak, E.; Usadel, K.; Kusterer, K.; Amini, P.; Frommeyer, R.; Tritschler, H.; Haak, T. Effects of alpha-lipoic acid on microcirculation in patients with peripheral diabetic neuropathy. Exp. Clin. Endocrinol. Diabetes 2000, 108, 168–174. [Google Scholar] [CrossRef]
- Abdali, D.; Samson, S.E.; Grover, A.K. How effective are antioxidant supplements in obesity and diabetes? Med. Princ. Pract. 2015, 24, 201–215. [Google Scholar] [CrossRef]
- de Almeida, D.A.T.; Braga, C.P.; Novelli, E.L.B.; Fernandes, A.A.H. Evaluation of lipid profile and oxidative stress in STZ-induced rats treated with antioxidant vitamin. Braz. Arch. Biol. Technol. 2012, 55, 527–536. [Google Scholar] [CrossRef]
- Bachmann, K.N.; Wang, T.J. Biomarkers of cardiovascular disease: Contributions to risk prediction in individuals with diabetes. Diabetologia 2018, 61, 987–995. [Google Scholar] [CrossRef]
- Varga, Z.V.; Giricz, Z.; Liaudet, L.; Haskó, G.; Ferdinandy, P.; Pacher, P. Interplay of oxidative, nitrosative/nitrative stress, inflammation, cell death and autophagy in diabetic cardiomyopathy. Biochim. Et Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 232–242. [Google Scholar] [CrossRef]
- Tappia, P.S.; Thliveris, J.; Xu, Y.-J.; Aroutiounova, N.; Dhalla, N.S. Effects of amino acid supplementation on myocardial cell damage and cardiac function in diabetes. Exp. Clin. Cardiol. 2011, 16, e17. [Google Scholar] [PubMed]
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The chemistry of reactive oxygen species (ROS) revisited: Outlining their role in biological macromolecules (DNA, lipids and proteins) and induced pathologies. Int. J. Mol. Sci. 2021, 22, 4642. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Buffer A | Buffer B | Column Regeneration Solution | Sample Dilution Buffer | |
|---|---|---|---|---|
| pH value | 3.45 | 10.85 | 2.20 | |
| Normality | 0.12 | 0.20 | 0.50 | 0.12 |
| Tri-sodium citrate dihydrate | 11.8 g | 19.6 g | 11.8 g | |
| NaOH | 3.1 g | 20.0 g | ||
| Citric acid | 6.0 g | 6.0 g | ||
| Boric acid | 5.0 g | |||
| Methanol | 65 mL | |||
| Thiodiglycol | 14 mL | |||
| Hydrochloric acid 32% | 6.5 mL | 12 mL | ||
| EDTA | 0.2 g | |||
| Phenol | 0.5 g | 2.0 g | ||
| Final volume | 1.0 L | 1.0 L | 1.0 L | 1.0 L |
| Parameter | Settings |
|---|---|
| RF power | 1.20 kW |
| Sampling depth | 3.0 mm |
| Plasma gas | Ar 8.0 L/min |
| Auxiliary gas | Ar 1.10 L/min |
| Carrier gas | Ar 0.70 L/min |
| Torch | Minitorch |
| Nebulizer | Nebulizer, 07UES |
| Chamber | Cyclone chamber |
| Chamber temp. | 5 °C |
| Cell gas (He) | 6.0 mL/min |
| Cell voltage | −21.0 V |
| Energy filter | 7 V |
| Amount (mg/100 mg) | ||||
|---|---|---|---|---|
| Bovine | Chicken | Sheep | Goat | |
| Moisture | 30,600 ± 0.001 d | 36,500 ± 0.008 b | 34,500 ± 0.003 c | 37,500 ± 0.009 a |
| Crude protein | 17,000 ± 0.008 a | 11,000 ± 0.002 c | 16,000 ± 0.003 b | 10,000 ± 0.001d |
| Crude fat | 0.170 ± 0.006 b | 0.120 ± 0.008 c | 0.160 ± 0.005 a | 0.100 ± 0.004 d |
| No. | Identified Compound | C:D | Type | Area % | RT | RI | |||
|---|---|---|---|---|---|---|---|---|---|
| Bovine | Chicken | Sheep | Goat | ||||||
| 1 | 13,16-Octadecadiynoic acid | C18:2 (13, 16) | PUFA | 0.80 | 4.04 | 707 | |||
| 2 | Caprylic acid | C8:0 | SFA | 0.15 | 0.21 | 5.82 | 905 | ||
| 3 | 9-Oxononanoic acid | C9:0 | SFA | 0.52 | 14.84 | 824 | |||
| 4 | Lauric acid | C12:0 | SFA | 0.41 | 16.99 | 846 | |||
| 5 | Tridecanoic acid | C13:0 | SFA | 0.83 | 21.51 | 807 | |||
| 6 | Myristic acid | C14:0 | SFA | 0.86 | 5.85 | 7.89 | 22.51 | 896 | |
| 7 | Pentadecanoic acid | C15:0 | SFA | 15.94 | 0.37 | 22.73 | 942 | ||
| 8 | Succinic acid | C15:0 | SFA | 2.25 | 0.07 | 24.39 | 941 | ||
| 9 | Palmitic acid | C16:0 | SFA | 19.30 | 33.89 | 23.62 | 12.07 | 26.66 | 969 |
| 10 | Palmitoleic acid | C16:1 (9) | MUFA | 4.26 | 0.82 | 0.35 | 27.10 | 911 | |
| 11 | Isomargaric acid | C17:0 | SFA | 7.48 | 29.43 | 832 | |||
| 12 | Margaric acid | C17:0 | SFA | 2.92 | 30.23 | 851 | |||
| 13 | Oleic acid | C18:1 (9) | MUFA | 22.99 | 54.70 * | 53.80 * | 60.81 * | 32.13 | 970 |
| 14 | Eicosanoic acid | C20:0 | SFA | 25.70 * | 1.01 | 0.84 | 32.74 | 937 | |
| 15 | Stearic acid | C18:0 | SFA | 5.04 | 8.64 | 9.03 | 34.10 | 924 | |
| 16 | 6,9-Linoleic acid | C18:2 (6, 9) | PUFA | 0.83 | 0.40 | 34.58 | 841 | ||
| 17 | 13,16-Octadecadiynoic acid | C18:2 (13, 16) | PUFA | 0.04 | 34.94 | 814 | |||
| 18 | 6,9,11-Octadecatrienoic acid | C18:3 (6, 9, 11) | PUFA | 1.00 | 35.26 | 851 | |||
| 19 | 8,11,14-Heptadecatrienoic acid | C17:3 (8, 11, 14) | PUFA | 0.14 | 35.71 | 831 | |||
| 20 | 9,12,15-Octadecatrienoic acid | C18:3 (9, 12, 15) | PUFA | 0.33 | 0.33 | 35.97 | 845 | ||
| 21 | 11-Eicosenoic acid | C20:1 (11) | MUFA | 0.35 | 1.28 | 36.17 | 884 | ||
| 22 | Cholestan-3-ol, 2-methylene- | C28:1 | Sterol | 0.16 | 0.89 | 37.20 | 778 | ||
| 23 | 6-Hexadecenoic acid, 7-methyl | C17:1 (6) | MUFA | 1.39 | 37.61 | 764 | |||
| 24 | 8,11,14-Eicosatrienoic acid | C20:3 (8, 11, 14) | PUFA | 0.11 | 37.72 | 804 | |||
| 25 | 5,8,11,14-Eicosatetraenoic acid | C20:4 (5, 8, 11, 14) | PUFA | 0.14 | 0.12 | 37.76 | 885 | ||
| 26 | 9,12-Linoleic acid | C18:2 (9, 12) | PUFA | 0.10 | 1.73 | 38.71 | 889 | ||
| 27 | Cholest-5-en-3-yl myristate | C41:1 | Sterol | 0.11 | 46.12 | 823 | |||
| 28 | Cholest-5-en-3-ol (3α), propanoate | C30:1 | Sterol | 0.10 | 46.14 | 775 | |||
| 29 | Cholesta-3,5-diene | C27:2 | Sterol | 0.37 | 46.36 | 886 | |||
| SFA | 71.91% | 41.47% | 42.31% | 29.2% | |||||
| USFA | 28.05% | 58.10% | 56.46% | 64.21% | |||||
| MUFA | 27.25% | 55.52% | 54.50% | 63.48% | |||||
| PUFA | 0.80% | 2.58% | 1.96% | 0.73% | |||||
| Sterol | 0.26% | 0.11% | 1.26% | ||||||
| Total | 99.96% | 99.83% | 98.88% | 94.67% | |||||
| No. | Identified Compound | RT | Amount (mg/100 mg) | |||
|---|---|---|---|---|---|---|
| Bovine | Chicken | Sheep | Goat | |||
| 1 | Aspartic acid | 7.699 | 0.080 ± 0.02 a | 0.254 ± 0.03 c | 0.093 ± 0.01 a | 0.122 ± 0.03 b |
| 2 | Threonine | 9.816 | 0.052 ± 0.01 a | 0.131 ± 0.03 b | 0.045 ± 0.03 a | 0.235 ± 0.03 c |
| 3 | Serine | 10.549 | 0.046 ± 0.01 a | 0.093 ± 0.01 a | 0.070 ± 0.03 a | 0.076 ± 0.03 a |
| 4 | Glutamic acid | 11.915 | 0.134 ± 0.03 a | 0.681 ± 0.03 b | 0.180 ± 0.01 a | 0.129 ± 0.02 a |
| 5 | Proline | 13.933 | 1.532 ± 0.01 b | 1.876 ± 0.04 b | 0.873 * ± 0.02 a | 3.570 ± 0.03 c |
| 6 | Glycine | 17.824 | 1.626 * ± 0.03 b | 2.688 * ± 0.03 c | 0.714 ± 0.03 a | 4.254 * ± 0.01 d |
| 7 | Alanine | 19.056 | 1.130 ± 0.01 b | 1.746 ± 0.03 b | 0.458 ± 0.03 a | 3.370 ± 0.03 c |
| 8 | Cystine | 21.264 | 0.761 ± 0.01 a | |||
| 9 | Valine | 21.915 | 0.327 ± 0.01 b | 0.417 ± 0.03 c | 0.189 ± 0.01 a | 0.701 ± 0.03 d |
| 10 | Methionine | 23.803 | 0.283 ± 0.03 a | 0.354 ± 0.05 b | 0.281 ± 0.01 a | 0.295 ± 0.03 a |
| 11 | Isoleucine | 25.915 | 0.158 ± 0.02 a | 0.357 ± 0.03 c | 0.280 ± 0.01 b | 0.366 ± 0.02 c |
| 12 | Leucine | 27.171 | 0.465 ± 0.03 b | 0.516 ± 0.03 c | 0.199 ± 0.03 a | 1.284 ± 0.01 d |
| 13 | Tyrosine | 30.315 | 0.076 ± 0.03 a | 0.067 ± 0.03 a | 0.043 ± 0.02 a | 0.202 ± 0.01 b |
| 14 | Phenylalanine | 31.389 | 0.241 ± 0.03 b | 0.379 ± 0.01 c | 0.083 ± 0.03 a | 0.623 ± 0.03 d |
| 15 | Histidine | 35.128 | 0.233 ± 0.01 c | 0.136 ± 0.03 b | 0.087 ± 0.03 a | 0.522 ± 0.04 d |
| 16 | Lysine | 39.381 | 0.258 ± 0.03 b | 0.377 ± 0.03 c | 0.060 ± 0.03 a | 0.978 ± 0.01 d |
| 17 | Arginine | 43.056 | 0.598 ± 0.02 b | 0.920 ± 0.01 c | 0.168 ± 0.01 a | 1.983 ± 0.05 d |
| Total AA | 7.237 | 11.753 | 3.823 | 18.708 | ||
| No. | Identified Element | Amount (mg/100 mg) | RDI | MDI | PMTDI | |||
|---|---|---|---|---|---|---|---|---|
| Bovine | Chicken | Sheep | Goat | |||||
| 1 | Al | 0.105 ± 0.02 a | 0.077 ± 0.01 b | 0.084 ± 0.02 b | 0.116 ± 0.02 a | 4.3 mg/kg/d | ||
| 2 | Cd | 0.051 ± 0.04 a | 0.038 ± 0.03 a | 0.046 ± 0.01 a | 0.067 ± 0.02 a | 1 mg/kg/d | ||
| 3 | Pb | 0.982 ± 0.02 b | 0.776 ± 0.07 c | 0.912 ± 0.04 b | 1.054 ± 0.02 a | 3.57 mg/kg/d | ||
| 4 | Cr | 6.010 ± 0.02 a | 5.600 ± 0.01 b | 5.600 ± 0.02 b | 6.700 ± 0.03 a | 25 µg (W), 35 µg (M) | 200 µg | 100 mg/kg/d |
| 5 | Ca | 2.722 ± 0.01 b | 2.181 ± 0.02 b | 2.475 ± 0.05 b | 3.000 ± 0.02 a | 700 mg | 2.500 mg | |
| 6 | Co | 0.195 ± 0.02 b | 0.093 ± 0.02 c | 0.126 ± 0.02 b | 0.288 ± 0.02 a | 5–8 µg | 8 µg/kg b.wt./d | |
| 7 | Cu | 0.016 ± 0.02 a | 0.082 ± 0.02 a | 0.012 ± 0.02 a | 0.019 ± 0.02 a | 900 µg | 1.400–1.700 µg | |
| 8 | Fe | 0.750 ± 0.01 b | 0.701 ± 0.04 b | 0.634 ± 0.02 c | 0.850 ± 0.07 a | 14.8 mg (W), 8.7 mg (M) | 45 mg | |
| 9 | Mg | 2.530 ± 0.01 a | 2.250 ± 0.02 a | 2.100 ± 0.02 a | 2.712 ± 0.02 a | 310–320 mg (W), 400–420 mg (M) | >420 mg | |
| 10 | P | 1.156 ± 0.03 a | 0.915 ± 0.06 b | 1.006 ± 0.02 a | 1.370 ± 0.02 a | 1.189 mg (W), 1.596 mg (M) | 4.000 mg | |
| 11 | Zn | 0.038 ± 0.02 a | 0.020 ± 0.05 a | 0.031 ± 0.02 a | 0.043 ± 0.09 a | 8 mg (W), 11 mg (M) | 40 mg | |
| Extract | Concentration | |
|---|---|---|
| 0.01 µg/mL | 0.05 µg/mL | |
| Bovine | 31.80 ± 1.11 a | 88.6 ± 7.00 b |
| Chicken | 6.80 ± 0.13 a | 16.00 ± 0.22 b |
| Sheep | 18.00 ± 0.44 a | 25.00 ± 1.90 b |
| Goat | 10.00 ± 0.03 a | 18.00 ± 0.04 b |
| Vitamin C | 82.00 ± 3.00 a | 88.00 ± 4.90 a |
| Extracts and Standard Drug | Control Rats | Diabetic Rats | Treated Rats |
|---|---|---|---|
| % Change | 110 ± 5.30 a | 350 ± 12.00 b 218.18 | |
| Bovine % Improvement | - | - | 155 ± 6.00 c 177.30 |
| Chicken % Improvement | - | - | 179 ± 10.12 d 155.50 |
| Sheep % Improvement | - | - | 160.00 ± 8.30 c 172.73 |
| Goat % Improvement | - | - | 175 ± 5.74 d 159.10 |
| Glibenclamide % Improvement | - | - | 140 ± 3.00 e 190.91 |
| Groups/Parameters | TL | TG | TC | HDL-C |
|---|---|---|---|---|
| Control | 600.00 ± 19.00 a | 102.00 ± 4.00 c | 110.00 ± 8.00 a | 50.00 ± 2.00 a |
| STZ-diabetic rats | 1250.00 ± 30.00 b | 250.00 ± 10.00 b | 279.00 ± 9.20 b | 19.10 ± 1.00 b |
| % Change | 108.33 | 145.10 | 153.64 | 61.80 |
| Bovine | 660.00 ± 20.00 c | 180.00 ± 3.00 a | 153.00 ± 8.00 a | 31.00 ± 2.00 b |
| % Improvement | 98.33 | 68.63 | 114.55 | 24.00 |
| Chicken | 950.00 ± 21.00 a | 200.00 ± 8.00 c | 191.00 ± 5.00 a | 23.00 ± 2.77 c |
| % Improvement | 50.00 | 49.01 | 80.00 | 8.00 |
| Sheep | 680.00 ± 15.00 d | 185.00 ± 5.00 d | 169.00 ± 5.20 d | 28.00 ± 1.00 c |
| % Improvement | 95.00 | 63.73 | 100.00 | 17.80 |
| Goat | 900.00 ± 22.00 c | 199.00 ± 4.00 a | 180.00 ± 5.00 c | 25.00 ± 1.10 b |
| % Improvement | 58.33 | 50.00 | 90.00 | 12.00 |
| Glibenclamide | 650.00 ± 28.70 ac | 111.20 ± 6.00 ac | 150.00 ± 6.00 a | 37.00 ± 3.20 a |
| % Improvement | 100.00 | 136.07 | 117.27 | 35.80 |
| Groups/Parameters | ALT (U/L) | AST(U/L) | ALP (U/L) | Bilirubin (mg/dL) | Total Protein (μg/mL) |
|---|---|---|---|---|---|
| Control | 85.00 ± 2.66 | 144.00 ± 8.20 a | 90.00 ± 5.00 a | 0.54 ± 0.05 a | 5.40 ± 0.43 a |
| STZ-Diabetic Rats | 230.00 ± 10.00 b | 265.00 ± 10.00 b | 190.00 ± 10.00 b | 1.40 ± 0.12 b | 2.00 ± 0.40 b |
| % Change | 170.56 | 84.03 | 111.11 | 159.30 | 62.96 |
| Bovine | 150.00 ± 8.10 c | 165.00 ± 6. 00 c | 140.00 ± 9.00 c | 0.90 ± 0.25 c | 3.98 ± 0.24 c |
| % Improvement | 94.12 | 69.44 | 55.56 | 92.59 | 36.67 |
| Chicken | 190.00 ± 9.66 a | 180.00 ± 9.00 a | 160.00 ± 8.12 a | 1.07 ± 0.30 a | 2.69 ± 0.41 a |
| % Improvement | 47.10 | 59.03 | 33.33 | 61.11 | 12.78 |
| Sheep | 182.55 ± 10.00 d | 169.00 ± 6.00 a | 144.00 ± 6.10 d | 1.00 ± 0.50 d | 2.34 ± 0.33 a |
| % Improvement | 55.83 | 66.67 | 51.11 | 74.10 | 6.30 |
| Goat | 189.00 ± 6.00 c | 178.00 ± 6.00 c | 151.00 ± 9.00 c | 1.02 ± 0.13 a | 2.89 ± 0.20 a |
| % Improvement | 48.34 | 60.42 | 43.33 | 70.37 | 16.48 |
| Glibenclamide | 89.00 ± 9.70 a | 159.00 ± 8.00 a | 133.00 ± 3.60 a | 0.59 ± 0.32 a | 4.90 ± 0.30 a |
| % Improvement | 165.88 | 73.61 | 63.33 | 150.00 | 53.70 |
| Groups/Parameters | Creatinine | Urea |
|---|---|---|
| Control | 0.34 ± 0.02 b | 34.00 ± 2.00 b |
| STZ-diabetic rats | 0.79 ± 0.04 a | 99.00 ± 4.00 a |
| % Change | 132.35 | 191.18 |
| Bovine | 0.50 ± 0.03 c | 50.00 ± 2.12 c |
| % Improvement | 85.29 | 144.12 |
| Chicken | 0.64 ± 0.04 b | 72.00 ± 3.22 b |
| % Improvement | 44.12 | 79.41 |
| Sheep | 0.56 ± 0.03 b | 52.00 ± 4.00 c |
| % Improvement | 67.65 | 138.24 |
| Goat | 0.60 ± 0.03 a | 69.00 ± 2.00 a |
| % Improvement | 55.88 | 88.23 |
| Glibenclamide | 0.39 ± 0.06 b | 43.00 ± 2.11 b |
| % Improvement | 117.65 | 164.71 |
| Groups/Parameters | IL-6 (pg/mL) | TNF-α (pg/mL) | sICAM-1 (pg/mL) | sVCAM-1 (pg/mL) |
|---|---|---|---|---|
| Control | 99.00 ± 5.00 a | 110.00 ± 9.00 a | 330.00 ± 11.00 a | 1200 ± 200.00 a |
| STZ-diabetic rats | 266.00 ± 11.00 b | 240.00 ± 10.00 b | 1200.00 ± 15.00 b | 21,900 ± 300.00 b |
| % Change | 168.69 | 118.18 | 63.64 | 1725.00 |
| Bovine | 190.00 ± 5.10 c | 194.00 ± 5.50 c | 600.00 ± 28.00 c | 10,500 ± 300.00 c |
| % Improvement | 76.77 | 41.82 | 181.82 | 950.00 |
| Chicken | 210.80 ± 5.10 a | 105.00 ± 5.34 a | 1000.00 ± 15.00 d | 12,000 ± 380.00 d |
| % Improvement | 55.76 | 122.73 | 60.61 | 825.00 |
| Sheep | 193.00 ± 6.00 d | 135.00 ± 6.30 d | 700.00 ± 10.00 d | 12,900 ± 230.00 d |
| % Improvement | 73.74 | 95.45 | 151.52 | 750.00 |
| Goat | 200.00 ± 8.00 c | 195.00 ± 5.98 c | 900.00 ± 22.00 b | 11,000 ± 350.00 b |
| % Improvement | 66.67 | 40.91 | 90.91 | 908.33 |
| Glibenclamide | 120.00 ± 9.00 d | 140.00 ± 10.00 d | 567.00 ± 18.90 d | 11,700 ± 160.00 a |
| % Improvement | 147.47 | 90.91 | 191.82 | 850.00 |
| Groups/Parameters | MDA (nmol/g Tissue) | GSH (nmol/g Tissue) |
|---|---|---|
| Control | 110.00 ± 5.00 a | 750.00 ± 12.00 a |
| STZ-Diabetic Rats | 450.00 ± 11.00 b | 200.00 ± 5.00 b |
| % Change | 309.10 | 73.33 |
| Bovine | 200.00 ± 15.10 c | 520.00 ± 10.00 c |
| % Improvement | 227.27 | 42.67 |
| Chicken | 310.00 ± 9.22 a | 313.00 ± 10.00 a |
| % Improvement | 127.27 | 15.10 |
| Sheep | 230.00 ± 6.00 d | 510.00 ± 6.00 d |
| % Improvement | 200.00 | 41.33 |
| Goat | 256.00 ± 7.00 c | 366.00 ± 10.00 c |
| % Improvement | 176.36 | 22.13 |
| Glibenclamide | 119.00 ± 6.10 a | 570.00 ± 10.00 d |
| % Improvement | 300.91 | 49.33 |
| Lesion | Control Group | Positive Control | Bovine | Chicken | Sheep | Goat | Standard Drug |
|---|---|---|---|---|---|---|---|
| Liver | |||||||
| Vacuolar degeneration of hepatocytes | 0 | 3 | 1 | 2 | 1 | 2 | 1 |
| Pancreas | |||||||
| Atrophy of islets of Langerhans | 0 | 3 | 1 | 2 | 1 | 2 | 1 |
| Periductal fibrosis | 0 | 2 | 1 | 1 | 1 | 1 | 1 |
| Vacuolation of exocrine pancreas | 0 | 2 | 0 | 1 | 1 | 1 | 1 |
| Necrosis of exocrine pancreas | 0 | 3 | 1 | 2 | 2 | 1 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Algehainy, N.A.; Mohamed, E.M.; Aly, H.F.; Younis, E.A.; Altemani, F.H.; Alanazi, M.A.; Bringmann, G.; Abdelmohsen, U.R.; Elmaidomy, A.H. Nutritional Composition and Anti-Type 2 Diabetes Mellitus Potential of Femur Bone Extracts from Bovine, Chicken, Sheep, and Goat: Phytochemical and In Vivo Studies. Nutrients 2023, 15, 4037. https://doi.org/10.3390/nu15184037
Algehainy NA, Mohamed EM, Aly HF, Younis EA, Altemani FH, Alanazi MA, Bringmann G, Abdelmohsen UR, Elmaidomy AH. Nutritional Composition and Anti-Type 2 Diabetes Mellitus Potential of Femur Bone Extracts from Bovine, Chicken, Sheep, and Goat: Phytochemical and In Vivo Studies. Nutrients. 2023; 15(18):4037. https://doi.org/10.3390/nu15184037
Chicago/Turabian StyleAlgehainy, Naseh A., Esraa M. Mohamed, Hanan F. Aly, Eman A. Younis, Faisal H. Altemani, Mohammad A. Alanazi, Gerhard Bringmann, Usama Ramadan Abdelmohsen, and Abeer H. Elmaidomy. 2023. "Nutritional Composition and Anti-Type 2 Diabetes Mellitus Potential of Femur Bone Extracts from Bovine, Chicken, Sheep, and Goat: Phytochemical and In Vivo Studies" Nutrients 15, no. 18: 4037. https://doi.org/10.3390/nu15184037
APA StyleAlgehainy, N. A., Mohamed, E. M., Aly, H. F., Younis, E. A., Altemani, F. H., Alanazi, M. A., Bringmann, G., Abdelmohsen, U. R., & Elmaidomy, A. H. (2023). Nutritional Composition and Anti-Type 2 Diabetes Mellitus Potential of Femur Bone Extracts from Bovine, Chicken, Sheep, and Goat: Phytochemical and In Vivo Studies. Nutrients, 15(18), 4037. https://doi.org/10.3390/nu15184037