
Toward Precision Weight-Loss Dietary Interventions: Findings from the POUNDS Lost Trial




Abstract
1. Introduction
2. The POUNDS Lost Trial
3. Gene–Diet Interactions
4. Epigenetic Modifications—DNA Methylation
5. Epigenetic Modifications—Thrifty microRNA
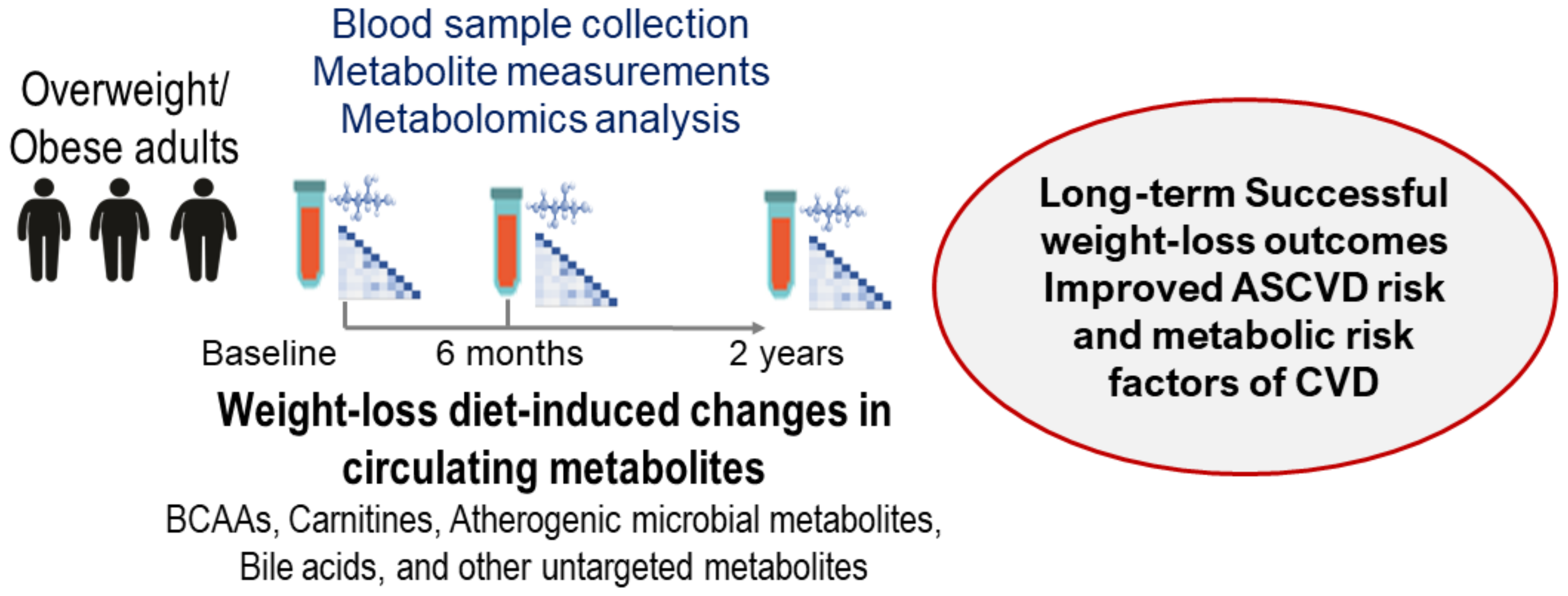
6. Metabolomics, Gut Microbiota Metabolites, Bile Acids, and Amino Acids
7. Biochemical, Behavioral and Psychological Factors
8. Summary and Future Directions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Alsalhe, T.A.; Chalghaf, N.; Riccò, M.; Bragazzi, N.L.; Wu, J. The global burden of disease attributable to high body mass index in 195 countries and territories, 1990–2017: An analysis of the Global Burden of Disease Study. PLoS Med. 2020, 17, e1003198. [Google Scholar] [CrossRef] [PubMed]
- Lingvay, I.; Sumithran, P.; Cohen, R.V.; le Roux, C.W. Obesity management as a primary treatment goal for type 2 diabetes: Time to reframe the conversation. Lancet 2022, 399, 394–405. [Google Scholar] [CrossRef]
- Bray, G.A.; Frühbeck, G.; Ryan, D.H.; Wilding, J. Management of obesity. Lancet 2016, 387, 1947–1956. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.E.; Kahali, B.; Berndt, S.I.; Justice, A.E.; Pers, T.H.; Day, F.R.; Powell, C.; Vedantam, S.; Buchkovich, M.L.; Yang, J.; et al. Genetic studies of body mass index yield new insights for obesity biology. Nature 2015, 518, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Chu, A.Y.; Kang, J.H.; Jensen, M.K.; Curhan, G.C.; Pasquale, L.R.; Qi, L. Sugar-sweetened beverages and genetic risk of obesity. N. Engl. J. Med. 2012, 367, 1387–1396. [Google Scholar] [CrossRef]
- Wang, T.; Heianza, Y.; Sun, D.; Huang, T.; Ma, W.; Rimm, E.B.; Manson, J.E.; Hu, F.B.; Willett, W.C.; Qi, L. Improving adherence to healthy dietary patterns, genetic risk, and long term weight gain: Gene-diet interaction analysis in two prospective cohort studies. BMJ 2018, 360, j5644. [Google Scholar] [CrossRef] [PubMed]
- Collins, F.S.; Varmus, H. A New Initiative on Precision Medicine. N. Engl. J. Med. 2015, 372, 793–795. [Google Scholar] [CrossRef]
- Qi, L. Nutrition for precision health: The time is now. Obesity 2022, 30, 1335–1344. [Google Scholar] [CrossRef]
- Qi, L.; Cho, Y.A. Gene-environment interaction and obesity. Nutr. Rev. 2008, 66, 684–694. [Google Scholar] [CrossRef]
- Wang, T.; Heianza, Y.; Sun, D.; Zheng, Y.; Huang, T.; Ma, W.; Rimm, E.B.; Manson, J.E.; Hu, F.B.; Willett, W.C.; et al. Improving fruit and vegetable intake attenuates the genetic association with long-term weight gain. Am. J. Clin. Nutr. 2019, 110, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Qi, L. Gene-Diet Interaction and Precision Nutrition in Obesity. Int. J. Mol. Sci. 2017, 18, 787. [Google Scholar] [CrossRef]
- Maki, K.C.; Slavin, J.L.; Rains, T.M.; Kris-Etherton, P.M. Limitations of observational evidence: Implications for evidence-based dietary recommendations. Adv. Nutr. Int. Rev. J. 2014, 5, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Murad, M.H.; Asi, N.; Alsawas, M.; Alahdab, F. New evidence pyramid. Evid. Based Med. 2016, 21, 125–127. [Google Scholar] [CrossRef]
- Qi, L. Gene–diet interaction and weight loss. Curr. Opin. Infect. Dis. 2014, 25, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, K.M.; Celis-Morales, C.; Papandonatos, G.D.; Erar, B.; Florez, J.C.; Jablonski, K.A.; Razquin, C.; Marti, A.; Heianza, Y.; Huang, T.; et al. FTO genotype and weight loss: Systematic review and meta-analysis of 9563 individual participant data from eight randomised controlled trials. BMJ 2016, 354, i4707. [Google Scholar] [CrossRef] [PubMed]
- Qi, L. Personalized nutrition and obesity. Ann. Med. 2014, 46, 247–252. [Google Scholar] [CrossRef]
- Meir, A.Y.; Keller, M.; Müller, L.; Bernhart, S.H.; Tsaban, G.; Zelicha, H.; Rinott, E.; Kaplan, A.; Gepner, Y.; Shelef, I.; et al. Effects of lifestyle interventions on epigenetic signatures of liver fat: Central randomized controlled trial. Liver Int. 2021, 41, 2101–2111. [Google Scholar] [CrossRef]
- Sacks, F.M.; Bray, G.A.; Carey, V.J.; Smith, S.R.; Ryan, D.H.; Anton, S.D.; McManus, K.; Champagne, C.M.; Bishop, L.M.; Laranjo, N.; et al. Comparison of weight-loss diets with different compositions of fat, protein, and carbohydrates. N. Engl. J. Med. 2009, 360, 859–873. [Google Scholar] [CrossRef]
- Dent, R.; McPherson, R.; Harper, M.-E. Factors affecting weight loss variability in obesity. Metabolism 2020, 113, 154388. [Google Scholar] [CrossRef]
- Bray, G.A.; Krauss, R.M.; Sacks, F.M.; Qi, L. Lessons Learned from the POUNDS Lost Study: Genetic, Metabolic, and Behavioral Factors Affecting Changes in Body Weight, Body Composition, and Cardiometabolic Risk. Curr. Obes. Rep. 2019, 8, 262–283. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.A.; Anton, S.D.; Han, H.; Champagne, C.M.; Allen, R.; LeBlanc, E.; Ryan, D.H.; McManus, K.; Laranjo, N.; Carey, V.J.; et al. Adherence is a multi-dimensional construct in the POUNDS LOST trial. J. Behav. Med. 2010, 33, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Qasim, A.; Turcotte, M.; de Souza, R.J.; Samaan, M.C.; Champredon, D.; Dushoff, J.; Speakman, J.R.; Meyre, D. On the origin of obesity: Identifying the biological, environmental and cultural drivers of genetic risk among human populations. Obes. Rev. 2018, 19, 121–149. [Google Scholar] [CrossRef] [PubMed]
- Shungin, D.; Winkler, T.W.; Croteau-Chonka, D.C.; Ferreira, T.; Locke, A.E.; Mägi, R.; Strawbridge, R.J.; Pers, T.H.; Fischer, K.; Justice, A.E.; et al. New genetic loci link adipose and insulin biology to body fat distribution. Nature 2015, 518, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Yengo, L.; Sidorenko, J.; Kemper, K.E.; Zheng, Z.; Wood, A.R.; Weedon, M.N.; Frayling, T.M.; Hirschhorn, J.; Yang, J.; Visscher, P.M.; et al. Meta-analysis of genome-wide association studies for height and body mass index in ∼700,000 individuals of European ancestry. Hum. Mol. Genet. 2018, 27, 3641–3649. [Google Scholar] [CrossRef] [PubMed]
- Claussnitzer, M.; Cho, J.H.; Collins, R.; Cox, N.J.; Dermitzakis, E.T.; Hurles, M.E.; Kathiresan, S.; Kenny, E.E.; Lindgren, C.M.; MacArthur, D.G.; et al. A brief history of human disease genetics. Nature 2020, 577, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Chu, A.Y.; Workalemahu, T.; Paynter, N.P.; Rose, L.M.; Giulianini, F.; Tanaka, T.; Ngwa, J.S.; Qi, Q.; Curhan, G.C.; Rimm, E.B.; et al. Novel locus including FGF21 is associated with dietary macronutrient intake. Hum. Mol. Genet. 2013, 22, 1895–1902. [Google Scholar] [CrossRef] [PubMed]
- Meddens, S.F.W.; de Vlaming, R.; Bowers, P.; Burik, C.A.P.; Linnér, R.K.; Lee, C.; Okbay, A.; Turley, P.; Rietveld, C.A.; Fontana, M.A.; et al. Genomic analysis of diet composition finds novel loci and associations with health and lifestyle. Mol. Psychiatry 2021, 26, 2056–2069. [Google Scholar] [CrossRef]
- May-Wilson, S.; Matoba, N.; Wade, K.H.; Hottenga, J.-J.; Concas, M.P.; Mangino, M.; Grzeszkowiak, E.J.; Menni, C.; Gasparini, P.; Timpson, N.J.; et al. Large-scale GWAS of food liking reveals genetic determinants and genetic correlations with distinct neurophysiological traits. Nat. Commun. 2022, 13, 2743. [Google Scholar] [CrossRef]
- Matoba, N.; Akiyama, M.; Ishigaki, K.; Kanai, M.; Takahashi, A.; Momozawa, Y.; Ikegawa, S.; Ikeda, M.; Iwata, N.; Hirata, M.; et al. GWAS of 165,084 Japanese individuals identified nine loci associated with dietary habits. Nat. Hum. Behav. 2020, 4, 308–316. [Google Scholar] [CrossRef]
- Satija, A.; Yu, E.; Willett, W.C.; Hu, F.B. Understanding Nutritional Epidemiology and Its Role in Policy. Adv. Nutr. Int. Rev. J. 2015, 6, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Bray, G.A.; Smith, S.R.; Hu, F.B.; Sacks, F.M.; Qi, L. Insulin receptor substrate 1 gene variation modifies insulin resistance response to weight-loss diets in a 2-year randomized trial: The preventing overweight using novel dietary strategies (pounds lost) trial. Circulation 2011, 124, 563–571. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, Q.; Zhang, C.; Smith, S.R.; Hu, F.B.; Sacks, F.M.; Bray, G.A.; Qi, L. FTO Genotype and 2-year change in body composition and fat distribution in response to weight-loss diets: The pounds lost trial. Diabetes 2012, 61, 3005–3011. [Google Scholar] [CrossRef]
- Huang, T.; Qi, Q.; Li, Y.; Hu, F.B.; Bray, G.A.; Sacks, F.M.; Williamson, D.A.; Qi, L. FTO genotype, dietary protein, and change in appetite: The Preventing Overweight Using Novel Dietary Strategies trial. Am. J. Clin. Nutr. 2014, 99, 1126–1130. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Zheng, Y.; Hruby, A.; A Williamson, D.; Bray, G.A.; Shen, Y.; Sacks, F.M.; Qi, L. Dietary Protein Modifies the Effect of the MC4R Genotype on 2-Year Changes in Appetite and Food Craving: The POUNDS Lost Trial. J. Nutr. 2017, 147, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Mattei, J.; Qi, Q.; Hu, F.B.; Sacks, F.M.; Qi, L. TCF7L2 genetic variants modulate the effect of dietary fat intake on changes in body composition during a weight-loss intervention. Am. J. Clin. Nutr. 2012, 96, 1129–1136. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.; Wang, T.; Heianza, Y.; Sun, D.; Ivey, K.; Durst, R.; Schwarzfuchs, D.; Stampfer, M.J.; Bray, G.A.; Sacks, F.M.; et al. HNF1A variant, energy-reduced diets and insulin resistance improvement during weight loss: The POUNDS Lost trial and DIRECT. Diabetes Obes. Metab. 2018, 20, 1445–1452. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. Weight-loss diets modify glucose-dependent insulinotropic polypeptide receptor rs2287019 genotype effects on changes in body weight, fasting glucose, and insulin resistance: The Preventing Overweight Using Novel Dietary Strategies trial. Am. J. Clin. Nutr. 2012, 95, 506–513. [Google Scholar] [CrossRef]
- Zhang, X.; Qi, Q.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. APOA5 genotype modulates 2-y changes in lipid profile in response to weight-loss diet intervention: The Pounds Lost Trial. Am. J. Clin. Nutr. 2012, 96, 917–922. [Google Scholar] [CrossRef]
- Xu, M.; Ng, S.S.; Bray, G.A.; Ryan, D.H.; Sacks, F.M.; Ning, G.; Qi, L. Dietary Fat Intake Modifies the Effect of a Common Variant in the LIPC Gene on Changes in Serum Lipid Concentrations during a Long-Term Weight-Loss Intervention Trial. J. Nutr. 2015, 145, 1289–1294. [Google Scholar] [CrossRef]
- Qi, Q.; Durst, R.; Schwarzfuchs, D.; Leitersdorf, E.; Shpitzen, S.; Li, Y.; Wu, H.; Champagne, C.M.; Shulman, G.I.; Stampfer, M.J.; et al. CETP genotype and changes in lipid levels in response to weight-loss diet intervention in the POUNDS LOST and DIRECT randomized trials. J. Lipid Res. 2015, 56, 713–721. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Qi, Q.; Liang, J.; Bray, G.A.; Hu, F.B.; Sacks, F.M.; Qi, L. Genetic determinant for amino acid metabolites and changes in body weight and insulin resistance in response to weight-loss diets: The preventing overweight using novel dietary strategies (POUNDS LOST) trial. Circulation 2013, 127, 1283–1289. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Sun, D.; Ma, W.; Zheng, Y.; Champagne, C.M.; Bray, G.A.; Sacks, F.M.; Qi, L. Gut-microbiome-related LCT genotype and 2-year changes in body composition and fat distribution: The POUNDS Lost Trial. Int. J. Obes. 2018, 42, 1565–1573. [Google Scholar] [CrossRef]
- Goni, L.; Sun, D.; Heianza, Y.; Wang, T.; Huang, T.; Cuervo, M.; Martínez, J.A.; Shang, X.; Bray, G.A.; Sacks, F.M.; et al. Macronutrient-specific effect of the MTNR1B genotype on lipid levels in response to 2 year weight-loss diets. J. Lipid Res. 2018, 59, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, K.; Xu, M.; Qi, Q.; de Jonge, L.; Bray, G.A.; Sacks, F.; Qi, L. Variants in glucose- and circadian rhythm–related genes affect the response of energy expenditure to weight-loss diets: The POUNDS LOST Trial. Am. J. Clin. Nutr. 2014, 99, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Ma, W.; Huang, T.; Wang, T.; Zheng, Y.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Macronutrient Intake–Associated FGF21 Genotype Modifies Effects of Weight-Loss Diets on 2-Year Changes of Central Adiposity and Body Composition: The POUNDS Lost Trial. Diabetes Care 2016, 39, 1909–1914. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Zhou, T.; Yuhang, C.; Huang, T.; Willett, W.C.; Hu, F.B.; Bray, G.A.; Sacks, F.M.; Qi, L. Starch Digestion–Related Amylase Genetic Variants, Diet, and Changes in Adiposity: Analyses in Prospective Cohort Studies and a Randomized Dietary Intervention. Diabetes 2020, 69, 1917–1926. [Google Scholar] [CrossRef] [PubMed]
- Groenendyk, J.W.; Greenland, P.; Khan, S.S. Incremental Value of Polygenic Risk Scores in Primary Prevention of Coronary Heart Disease: A Review. JAMA Intern. Med. 2022, 182, 1082–1088. [Google Scholar] [CrossRef]
- Zhou, T.; Sun, D.; Heianza, Y.; Li, X.; Champagne, C.M.; LeBoff, M.S.; Shang, X.; Pei, X.; Bray, G.A.; Sacks, F.M.; et al. Genetically determined vitamin D levels and change in bone density during a weight-loss diet intervention: The Preventing Overweight Using Novel Dietary Strategies (POUNDS Lost) Trial. Am. J. Clin. Nutr. 2018, 108, 1129–1134. [Google Scholar] [CrossRef]
- Huang, T.; Ley, S.H.; Zheng, Y.; Wang, T.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic susceptibility to diabetes and long-term improvement of insulin resistance and β cell function during weight loss: The Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Am. J. Clin. Nutr. 2016, 104, 198–204. [Google Scholar] [CrossRef]
- Kurilshikov, A.; Medina-Gomez, C.; Bacigalupe, R.; Radjabzadeh, D.; Wang, J.; Demirkan, A.; Le Roy, C.I.; Garay, J.A.R.; Finnicum, C.T.; Liu, X.; et al. Large-scale association analyses identify host factors influencing human gut microbiome composition. Nat. Genet. 2021, 53, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xue, Q.; Ma, H.; Champagne, C.M.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetically determined gut microbial abundance and 2-year changes in central adiposity and body composition: The POUNDS lost trial. Clin. Nutr. 2022, 41, 2817–2824. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhou, T.; Li, X.; Heianza, Y.; Liang, Z.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic Susceptibility, Dietary Protein Intake, and Changes of Blood Pressure: The POUNDS Lost Trial. Hypertension 2019, 74, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhou, T.; Ma, H.; Heianza, Y.; Champagne, C.M.; Williamson, D.A.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic variation in lean body mass, changes of appetite and weight loss in response to diet interventions: The POUNDS Lost trial. Diabetes Obes. Metab. 2020, 22, 2305–2315. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhou, T.; Sun, D.; Li, X.; Ma, H.; Liang, Z.; Heianza, Y.; Pei, X.; Bray, G.A.; Sacks, F.M.; et al. Distinct genetic subtypes of adiposity and glycemic changes in response to weight-loss diet intervention: The POUNDS Lost trial. Eur. J. Nutr. 2020, 60, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Sun, D.; Li, X.; Heianza, Y.; LeBoff, M.S.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetically determined SCFA concentration modifies the association of dietary fiber intake with changes in bone mineral density during weight loss: The Preventing Overweight Using Novel Dietary Strategies (POUNDS LOST) trial. Am. J. Clin. Nutr. 2021, 114, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Smith, Z.D.; Meissner, A. DNA methylation: Roles in mammalian development. Nat. Rev. Genet. 2013, 14, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Qi, L. Epigenetics in Precision Nutrition. J. Pers. Med. 2022, 12, 533. [Google Scholar] [CrossRef]
- Dick, K.J.; Nelson, C.P.; Tsaprouni, L.; Sandling, J.K.; Aïssi, D.; Wahl, S.; Meduri, E.; Morange, P.-E.; Gagnon, F.; Grallert, H.; et al. DNA methylation and body-mass index: A genome-wide analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.-C.; Ried, J.S.; Zhang, W.; Yang, Y.; et al. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2016, 541, 81–86. [Google Scholar] [CrossRef]
- Mandaviya, P.R.; Joehanes, R.; Brody, J.; Castillo-Fernandez, J.E.; Dekkers, K.F.; Do, A.N.; Graff, M.; Hänninen, I.K.; Tanaka, T.; de Jonge, E.A.L.; et al. Association of dietary folate and vitamin B-12 intake with genome-wide DNA methylation in blood: A large-scale epigenome-wide association analysis in 5841 individuals. Am. J. Clin. Nutr. 2019, 110, 437–450. [Google Scholar] [CrossRef]
- Do, W.L.; Whitsel, E.A.; Costeira, R.; Masachs, O.M.; Le Roy, C.I.; Bell, J.T.; Staimez, L.R.; Stein, A.D.; Smith, A.K.; Horvath, S.; et al. Epigenome-wide association study of diet quality in the Women’s Health Initiative and TwinsUK cohort. Int. J. Epidemiol. 2021, 50, 675–684. [Google Scholar] [CrossRef]
- Li, X.; Shao, X.; Bazzano, L.A.; Xue, Q.; Koseva, B.S.; Grundberg, E.; Shai, I.; Bray, G.A.; Sacks, F.M.; Qi, L. Blood DNA methylation at TXNIP and glycemic changes in response to weight-loss diet interventions: The POUNDS lost trial. Int. J. Obes. 2022, 46, 1122–1127. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shao, X.; Xue, Q.; Kou, M.; Champagne, C.M.; Koseva, B.S.; Heianza, Y.; Grundberg, E.; Bazzano, L.A.; Bray, G.A.; et al. DNA methylation near cpt1a and changes in triglyceride-rich lipoproteins in response to weight-loss diet interventions. J. Clin. Endocrinol. Metab. 2023, 108, e542–e549. [Google Scholar] [CrossRef]
- Kou, M.; Li, X.; Shao, X.; Grundberg, E.; Wang, X.; Ma, H.; Heianza, Y.; Martinez, J.A.; Bray, G.A.; Sacks, F.M.; et al. DNA Methylation of Birthweight–Blood Pressure Genes and Changes of Blood Pressure in Response to Weight-Loss Diets in the POUNDS Lost Trial. Hypertension 2023, 80, 1223–1230. [Google Scholar] [CrossRef]
- Sun, D.; Heianza, Y.; Li, X.; Shang, X.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Genetic, epigenetic and transcriptional variations at NFATC2IP locus with weight loss in response to diet interventions: The POUNDS Lost Trial. Diabetes Obes. Metab. 2018, 20, 2298–2303. [Google Scholar] [CrossRef] [PubMed]
- Yoshihara, E. TXNIP/TBP-2: A Master Regulator for Glucose Homeostasis. Antioxidants 2020, 9, 765. [Google Scholar] [CrossRef] [PubMed]
- Parikh, H.; Carlsson, E.; Chutkow, W.A.; Johansson, L.E.; Storgaard, H.; Poulsen, P.; Saxena, R.; Ladd, C.; Schulze, P.C.; Mazzini, M.J.; et al. TXNIP regulates peripheral glucose metabolism in humans. PLOS Med. 2007, 4, e158. [Google Scholar] [CrossRef]
- Florath, I.; Butterbach, K.; Heiss, J.; Bewerunge-Hudler, M.; Zhang, Y.; Schöttker, B.; Brenner, H. Type 2 diabetes and leucocyte DNA methylation: An epigenome-wide association study in over 1500 older adults. Diabetologia 2016, 59, 130–138. [Google Scholar] [CrossRef]
- Meeks, K.A.C.; Henneman, P.; Venema, A.; Addo, J.; Bahendeka, S.; Burr, T.; Danquah, I.; Galbete, C.; Mannens, M.M.A.M.; Mockenhaupt, F.P.; et al. Epigenome-wide association study in whole blood on type 2 diabetes among sub-Saharan African individuals: Findings from the RODAM study. Leuk. Res. 2018, 48, 58–70. [Google Scholar] [CrossRef]
- Jones, A.C.; Irvin, M.R.; Claas, S.A.; Arnett, D.K. Lipid Phenotypes and DNA Methylation: A Review of the Literature. Curr. Atheroscler. Rep. 2021, 23, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Küpers, L.K.; Monnereau, C.; Sharp, G.C.; Yousefi, P.; Salas, L.A.; Ghantous, A.; Page, C.M.; Reese, S.E.; Wilcox, A.J.; Czamara, D.; et al. Meta-analysis of epigenome-wide association studies in neonates reveals widespread differential DNA methylation associated with birthweight. Nat. Commun. 2019, 10, 1893. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Ollikainen, M.; Muniandy, M.; Zhang, T.; van Dongen, J.; Hao, G.; van der Most, P.J.; Pan, Y.; Pervjakova, N.; Sun, Y.V.; et al. Identification, Heritability, and Relation with Gene Expression of Novel DNA Methylation Loci for Blood Pressure. Hypertension 2020, 76, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Miao, F.; Braffett, B.H.; Lachin, J.M.; Zhang, L.; Wu, X.; Roshandel, D.; Carless, M.; Li, X.A.; Tompkins, J.D.; et al. DNA methylation mediates development of HbA1c-associated complications in type 1 diabetes. Nat. Metab. 2020, 2, 744–762. [Google Scholar] [CrossRef] [PubMed]
- Tobi, E.W.; Slieker, R.C.; Luijk, R.; Dekkers, K.F.; Stein, A.D.; Xu, K.M.; Slagboom, P.E.; van Zwet, E.W.; Lumey, L.H.; Heijmans, B.T.; et al. DNA methylation as a mediator of the association between prenatal adversity and risk factors for metabolic disease in adulthood. Sci. Adv. 2018, 4, eaao4364. [Google Scholar] [CrossRef] [PubMed]
- Neel, J.V. Diabetes mellitus: A “thrifty” genotype rendered detrimental by “progress”? Am. J. Hum. Genet. Dec. 1962, 14, 353–362. [Google Scholar]
- Gibson, G. Human Evolution: Thrifty Genes and the Dairy Queen. Curr. Biol. 2007, 17, R295–R296. [Google Scholar] [CrossRef]
- Heianza, Y.; Xue, Q.; Rood, J.; Bray, G.A.; Sacks, F.M.; Qi, L. Circulating thrifty microRNA is related to insulin sensitivity, adiposity, and energy metabolism in adults with overweight and obesity: The POUNDS Lost trial. Am. J. Clin. Nutr. 2023, 117, 121–129. [Google Scholar] [CrossRef]
- de Jonge, L.; Bray, G.A.; Smith, S.R.; Ryan, D.H.; de Souza, R.J.; Loria, C.M.; Champagne, C.M.; Williamson, D.A.; Sacks, F.M. Effect of diet composition and weight loss on resting energy expenditure in the pounds lost study. Obesity 2012, 20, 2384–2389. [Google Scholar] [CrossRef]
- Bray, G.A.; Smith, S.R.; DeJonge, L.; de Souza, R.; Rood, J.; Champagne, C.M.; Laranjo, N.; Carey, V.; Obarzanek, E.; Loria, C.M.; et al. Effect of diet composition on energy expenditure during weight loss: The POUNDS LOST Study. Int. J. Obes. 2012, 36, 448–455. [Google Scholar] [CrossRef][Green Version]
- Wang, L.; Sinnott-Armstrong, N.; Wagschal, A.; Wark, A.R.; Camporez, J.-P.; Perry, R.J.; Ji, F.; Sohn, Y.; Oh, J.; Wu, S.; et al. A MicroRNA Linking Human Positive Selection and Metabolic Disorders. Cell 2020, 183, 684–701.e14. [Google Scholar] [CrossRef] [PubMed]
- Isaac, R.; Reis, F.C.G.; Ying, W.; Olefsky, J.M. Exosomes as mediators of intercellular crosstalk in metabolism. Cell Metab. 2021, 33, 1744–1762. [Google Scholar] [CrossRef] [PubMed]
- Huang-Doran, I.; Zhang, C.-Y.; Vidal-Puig, A. Extracellular Vesicles: Novel Mediators of Cell Communication In Metabolic Disease. Trends Endocrinol. Metab. 2017, 28, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Thomou, T.; Mori, M.A.; Dreyfuss, J.M.; Konishi, M.; Sakaguchi, M.; Wolfrum, C.; Rao, T.N.; Winnay, J.N.; Garcia-Martin, R.; Grinspoon, S.K.; et al. Adipose-derived circulating miRNAs regulate gene expression in other tissues. Nature 2017, 542, 450–455. [Google Scholar] [CrossRef] [PubMed]
- Valles-Colomer, M.; Menni, C.; Berry, S.E.; Valdes, A.M.; Spector, T.D.; Segata, N. Cardiometabolic health, diet and the gut microbiome: A meta-omics perspective. Nat. Med. 2023, 29, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, H.K.; Gudmundsdottir, V.; Nielsen, H.B.; Hyotylainen, T.; Nielsen, T.; Jensen, B.A.H.; Forslund, K.; Hildebrand, F.; Prifti, E.; Falony, G.; et al. Human gut microbes impact host serum metabolome and insulin sensitivity. Nature 2016, 535, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Visconti, A.; Le Roy, C.I.; Rosa, F.; Rossi, N.; Martin, T.C.; Mohney, R.P.; Li, W.; de Rinaldis, E.; Bell, J.T.; Venter, J.C.; et al. Interplay between the human gut microbiome and host metabolism. Nat. Commun. 2019, 10, 4505. [Google Scholar] [CrossRef]
- Heianza, Y.; Sun, D.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Changes in Gut Microbiota–Related Metabolites and Long-term Successful Weight Loss in Response to Weight-Loss Diets: The POUNDS Lost Trial. Diabetes Care 2018, 41, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Ceglarek, U.; Huang, T.; Li, L.; Rood, J.; Ryan, D.H.; Bray, G.A.; Sacks, F.M.; Schwarzfuchs, D.; Thiery, J.; et al. Weight-loss diets and 2-y changes in circulating amino acids in 2 randomized intervention trials1–3. Am. J. Clin. Nutr. 2016, 103, 505–511. [Google Scholar] [CrossRef]
- Li, X.; Sun, D.; Zhou, T.; Ma, H.; Heianza, Y.; Liang, Z.; Bray, G.A.; Sacks, F.M.; Qi, L. Changes of Branched-Chain Amino Acids and Ectopic Fat in Response to Weight-loss Diets: The POUNDS Lost Trial. J. Clin. Endocrinol. Metab. 2020, 105, e3747–e3756. [Google Scholar] [CrossRef]
- Heianza, Y.; Sun, D.; Li, X.; DiDonato, J.A.; Bray, G.A.; Sacks, F.M.; Qi, L. Gut microbiota metabolites, amino acid metabolites and improvements in insulin sensitivity and glucose metabolism: The POUNDS Lost trial. Gut 2019, 68, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, B.; Marchianò, S.; Moraca, F.; Sepe, V.; Carino, A.; Rapacciuolo, P.; Biagioli, M.; Limongelli, V.; Zampella, A.; Catalanotti, B.; et al. Discovery of Bile Acid Derivatives as Potent ACE2 Activators by Virtual Screening and Essential Dynamics. J. Chem. Inf. Model. 2022, 62, 196–209. [Google Scholar] [CrossRef]
- Porez, G.; Prawitt, J.; Gross, B.; Staels, B. Bile acid receptors as targets for the treatment of dyslipidemia and cardiovascular disease. J. Lipid Res. 2012, 53, 1723–1737. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, T.R.; Haeusler, R.A. Bile acids in glucose metabolism and insulin signalling—Mechanisms and research needs. Nat. Rev. Endocrinol. 2019, 15, 701–712. [Google Scholar] [CrossRef]
- Molinaro, A.; Wahlström, A.; Marschall, H.-U. Role of Bile Acids in Metabolic Control. Trends Endocrinol. Metab. 2018, 29, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Wang, X.; Rood, J.; Clish, C.B.; Bray, G.A.; Sacks, F.M.; Qi, L. Changes in circulating bile acid subtypes in response to weight-loss diets are associated with improvements in glycemic status and insulin resistance: The POUNDS Lost trial. Metabolism 2022, 136, 155312. [Google Scholar] [CrossRef] [PubMed]
- Heianza, Y.; Zhou, T.; He, H.; Rood, J.; Clish, C.B.; Bray, G.A.; Sacks, F.M.; Qi, L. Changes in bile acid subtypes and long-term successful weight-loss in response to weight-loss diets: The POUNDS lost trial. Liver Int. 2022, 42, 363–373. [Google Scholar] [CrossRef]
- Achari, A.E.; Jain, S.K. Adiponectin, A Therapeutic Target for Obesity, Diabetes, and Endothelial Dysfunction. Int. J. Mol. Sci. 2017, 18, 1321. [Google Scholar] [CrossRef]
- Ma, W.; Huang, T.; Zheng, Y.; Wang, M.; Bray, G.A.; Sacks, F.M.; Qi, L. Weight-Loss Diets, Adiponectin, and Changes in Cardiometabolic Risk in the 2-Year POUNDS Lost Trial. J. Clin. Endocrinol. Metab. 2016, 101, 2415–2422. [Google Scholar] [CrossRef]
- Liu, G.; Liang, L.; Bray, G.A.; Qi, L.; Hu, F.B.; Rood, J.; Sacks, F.M.; Sun, Q. Thyroid hormones and changes in body weight and metabolic parameters in response to weight loss diets: The POUNDS LOST trial. Int. J. Obes. 2017, 41, 878–886. [Google Scholar] [CrossRef]
- Xue, Q.; Li, X.; Ma, H.; Tao, Z.; Heianza, Y.; Rood, J.C.; Bray, G.A.; Sacks, F.M.; Qi, L. Changes in pedometer-measured physical activity are associated with weight loss and changes in body composition and fat distribution in response to reduced-energy diet interventions: The POUNDS Lost trial. Diabetes Obes. Metab. 2022, 24, 1000–1009. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Li, X.; Zhou, T.; Ma, H.; Heianza, Y.; Williamson, D.A.; Smith, S.R.; Bray, G.A.; Sacks, F.M.; Qi, L. Sleep Disturbance and Changes in Energy Intake and Body Composition During Weight Loss in the POUNDS Lost Trial. Diabetes 2022, 71, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hanseman, D.J.; Champagne, C.M.; Bray, G.A.; Qi, L.; Williamson, D.A.; Anton, S.D.; Sacks, F.M.; Tong, J. Predicting Weight Loss Using Psychological and Behavioral Factors: The POUNDS LOST Trial. J. Clin. Endocrinol. Metab. 2020, 105, 1274–1283. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Snyder, M.P. Integrative omics for health and disease. Nat. Rev. Genet. 2018, 19, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Li, Y.; Stoll, M.L.; Tollefsbol, T.O. The Epigenetic Connection Between the Gut Microbiome in Obesity and Diabetes. Front. Genet. 2020, 10, 1329. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Asmis, R.; Hawkins, G.A.; Howard, T.D.; Cox, L.A. The Need for Multi-Omics Biomarker Signatures in Precision Medicine. Int. J. Mol. Sci. 2019, 20, 4781. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Rienks, M.; Theofilatos, K.; Mayr, M. Systems biology in cardiovascular disease: A multiomics approach. Nat. Rev. Cardiol. 2021, 18, 313–330. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.M. Precision Medicine in Type 2 Diabetes: Using Individualized Prediction Models to Optimize Selection of Treatment. Diabetes 2020, 69, 2075–2085. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Fat (%) | Proteins (%) | Carbohydrates (%) |
|---|---|---|---|
| low-fat, average protein | 20 | 15 | 65 |
| low-fat, high protein | 20 | 25 | 55 |
| high-fat, average protein | 40 | 15 | 45 |
| high-fat, high protein | 40 | 25 | 35 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qi, L.; Heianza, Y.; Li, X.; Sacks, F.M.; Bray, G.A. Toward Precision Weight-Loss Dietary Interventions: Findings from the POUNDS Lost Trial. Nutrients 2023, 15, 3665. https://doi.org/10.3390/nu15163665
Qi L, Heianza Y, Li X, Sacks FM, Bray GA. Toward Precision Weight-Loss Dietary Interventions: Findings from the POUNDS Lost Trial. Nutrients. 2023; 15(16):3665. https://doi.org/10.3390/nu15163665
Chicago/Turabian StyleQi, Lu, Yoriko Heianza, Xiang Li, Frank M. Sacks, and George A. Bray. 2023. "Toward Precision Weight-Loss Dietary Interventions: Findings from the POUNDS Lost Trial" Nutrients 15, no. 16: 3665. https://doi.org/10.3390/nu15163665
APA StyleQi, L., Heianza, Y., Li, X., Sacks, F. M., & Bray, G. A. (2023). Toward Precision Weight-Loss Dietary Interventions: Findings from the POUNDS Lost Trial. Nutrients, 15(16), 3665. https://doi.org/10.3390/nu15163665