

Folate Deficiency Enhanced Inflammation and Exacerbated Renal Fibrosis in High-Fat High-Fructose Diet-Fed Mice
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Animals and Composition of Diets
2.2. Measurement of Serum Folate Level and Biomarkers
2.3. Measurement of Fasting Blood Glucose and Serum Cholesterol
2.4. Measurement of Blood Pressure and Heart Rate
2.5. Determination of Renal Function Biomarkers
2.6. Assay of Cytokine Secretions from Primary Splenocytes
2.7. Histopathological Analysis
2.8. Determination of Cytokines and Collagen Contents in Renal Tissue Homogenates
2.9. Western Blot Analysis
2.10. Statistical Analysis
3. Results
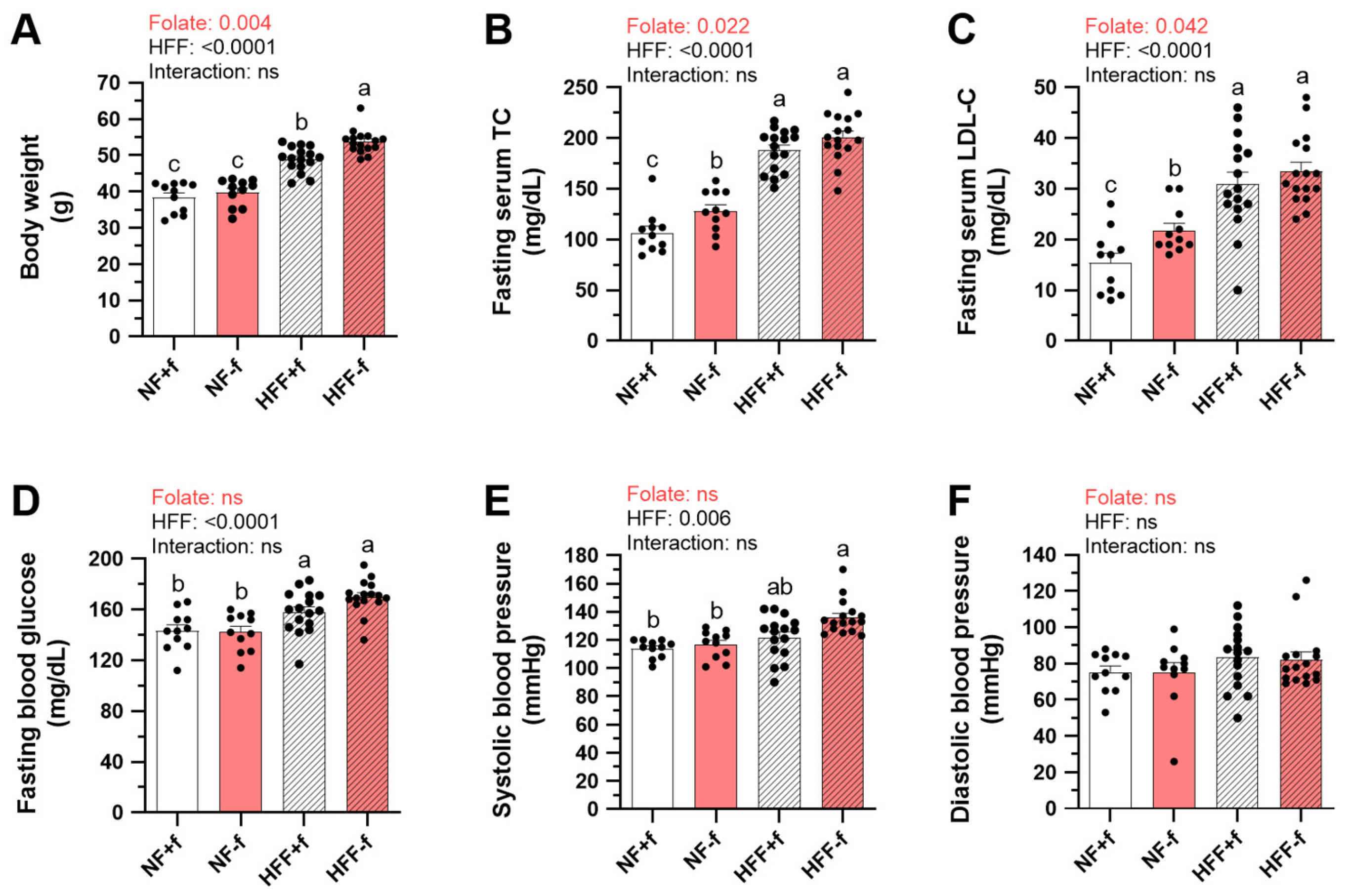
3.1. The Highest Body Weight, Serum Cholesterol, and Systolic Blood Pressure in Mice Fed High-Fat High-Fructose Diet without Folate
3.2. Higher Inflammatory Cytokine Productions by Splenocytes from Mice with Folate Deficiency
3.3. Higher Serum and Urinary Biomarkers of Kidney Dysfunction in Mice with Folate Deficiency
3.4. Higher Perirenal White Adipose Tissue, Renal Leptin, and Renal TGF-β1 Contents in Mice with Folate Deficiency
3.5. Higher Renal Pro-Inflammatory but Lower Anti-Inflammatory Cytokines in Mice with Folate Deficiency
3.6. Exacerbated Renal Extracellular Matrix Depositions and Fibrosis in Mice with Folate Deficiency
3.7. Folate Deficiency Promoted Renal Pro-Fibrosis Signaling Activation in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Choe, S.S.; Huh, J.Y.; Hwang, I.J.; Kim, J.I.; Kim, J.B. Adipose tissue remodeling: Its role in energy metabolism and metabolic disorders. Front. Endocrinol. 2016, 7, 30. [Google Scholar] [CrossRef] [PubMed]
- Câmara, N.O.; Iseki, K.; Kramer, H.; Liu, Z.H.; Sharma, K. Kidney disease and obesity: Epidemiology, mechanisms and treatment. Nat. Rev. Nephrol. 2017, 13, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Carney, E.F. The impact of chronic kidney disease on global health. Nat. Rev. Nephrol. 2020, 16, 251. [Google Scholar] [CrossRef]
- D’Agati, V.D.; Chagnac, A.; de Vries, A.P.; Levi, M.; Porrini, E.; Herman-Edelstein, M.; Praga, M. Obesity-related glomerulopathy: Clinical and pathologic characteristics and pathogenesis. Nat. Rev. Nephrol. 2016, 12, 453–471. [Google Scholar] [CrossRef]
- Kambham, N.; Markowitz, G.S.; Valeri, A.M.; Lin, J.; D’Agati, V.D. Obesity-related glomerulopathy: An emerging epidemic. Kidney Int. 2001, 59, 1498–1509. [Google Scholar] [CrossRef]
- Wuerzner, G.; Pruijm, M.; Maillard, M.; Bovet, P.; Renaud, C.; Burnier, M.; Bochud, M. Marked association between obesity and glomerular hyperfiltration: A cross-sectional study in an African population. Am. J. Kidney Dis. 2010, 56, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Coppolino, G.; Comi, N.; Bolignano, D.; Patella, G.; Comi, A.; Provenzano, M.; Rivoli, L.; Andreucci, M.; Fuiano, G. Urinary neutrophil gelatinase-associated lipocalin (NGAL) predicts renal function decline in patients with glomerular diseases. Front. Cell Dev. Biol. 2020, 8, 336. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Oh, K.H.; Park, S.K. Dietary micronutrients and risk of chronic kidney disease: A cohort study with 12 year follow-up. Nutrients 2021, 13, 1517. [Google Scholar] [CrossRef]
- Thuesen, B.H.; Husemoen, L.L.; Ovesen, L.; Jørgensen, T.; Fenger, M.; Linneberg, A. Lifestyle and genetic determinants of folate and vitamin B12 levels in a general adult population. Br. J. Nutr. 2010, 103, 1195–1204. [Google Scholar] [CrossRef]
- Chan, C.W.; Chan, P.H.; Lin, B.F. Folate deficiency increased lipid accumulation and leptin production of adipocytes. Front. Nutr. 2022, 9, 852451. [Google Scholar] [CrossRef]
- Mendoza-Herrera, K.; Florio, A.A.; Moore, M.; Marrero, A.; Tamez, M.; Bhupathiraju, S.N.; Mattei, J. The leptin system and diet: A mini review of the current evidence. Front. Endocrinol. 2021, 12, 749050. [Google Scholar] [CrossRef] [PubMed]
- Alix, P.M.; Guebre-Egziabher, F.; Soulage, C.O. Leptin as an uremic toxin: Deleterious role of leptin in chronic kidney disease. Biochimie 2014, 105, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.C.; Teo, B.W.; Tai, E.S.; Lim, S.C.; Chan, C.M.; Sethi, S.; Wong, T.Y.; Sabanayagam, C. Elevated serum leptin, adiponectin and leptin to adiponectin ratio is associated with chronic kidney disease in Asian adults. PLoS ONE 2015, 10, e0122009. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Riejos, P.; Najib, S.; Santos-Alvarez, J.; Martín-Romero, C.; Pérez-Pérez, A.; González-Yanes, C.; Sánchez-Margalet, V. Role of leptin in the activation of immune cells. Mediat. Inflamm. 2010, 2010, 568343. [Google Scholar] [CrossRef]
- Kolb, A.F.; Petrie, L. Folate deficiency enhances the inflammatory response of macrophages. Mol. Immunol. 2013, 54, 164–172. [Google Scholar] [CrossRef]
- Kinoshita, M.; Kayama, H.; Kusu, T.; Yamaguchi, T.; Kunisawa, J.; Kiyono, H.; Sakaguchi, S.; Takeda, K. Dietary folic acid promotes survival of Foxp3+ regulatory T cells in the colon. J. Immunol. 2012, 189, 2869–2878. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Huang, T.C.; Lin, B.F. Folate deficiency affects dendritic cell function and subsequent T helper cell differentiation. J. Nutr. Biochem. 2017, 41, 65–72. [Google Scholar] [CrossRef]
- Zhu, Q.; Scherer, P.E. Immunologic and endocrine functions of adipose tissue: Implications for kidney disease. Nat. Rev. Nephrol. 2018, 14, 105–120. [Google Scholar] [CrossRef]
- Martin-Taboada, M.; Vila-Bedmar, R.; Medina-Gómez, G. From obesity to chronic kidney disease: How can adipose tissue affect renal function? Nephron 2021, 145, 609–613. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, Z.; Proctor, G.; Moskowitz, S.; Liebman, S.E.; Rogers, T.; Lucia, M.S.; Li, J.; Levi, M. Diet-induced obesity in C57BL/6J mice causes increased renal lipid accumulation and glomerulosclerosis via a sterol regulatory element-binding protein-1c-dependent pathway. J. Biol. Chem. 2005, 280, 32317–32325. [Google Scholar] [CrossRef]
- Proctor, G.; Jiang, T.; Iwahashi, M.; Wang, Z.; Li, J.; Levi, M. Regulation of renal fatty acid and cholesterol metabolism, inflammation, and fibrosis in Akita and OVE26 mice with type 1 diabetes. Diabetes 2006, 55, 2502–2509. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ge, X.; Li, X.; He, J.; Wei, X.; Du, J.; Sun, J.; Li, X.; Xun, Z.; Liu, W.; et al. High-fat diet promotes renal injury by inducing oxidative stress and mitochondrial dysfunction. Cell Death Dis. 2020, 11, 914. [Google Scholar] [CrossRef]
- Haller, H.; Bertram, A.; Nadrowitz, F.; Menne, J. Monocyte chemoattractant protein-1 and the kidney. Curr. Opin. Nephrol. Hypertens. 2016, 25, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Lei, C.T.; Zhang, C. Interleukin-6 signaling pathway and its role in kidney disease: An update. Front. Immunol. 2017, 8, 405. [Google Scholar] [CrossRef]
- Meng, X.M.; Tang, P.M.; Li, J.; Lan, H.Y. TGF-β/Smad signaling in renal fibrosis. Front. Physiol. 2015, 6, 82. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Booz, G.W.; Wang, Y.; Fan, F.; Roman, R.J. Inflammation and renal fibrosis: Recent developments on key signaling molecules as potential therapeutic targets. Eur. J. Pharmacol. 2018, 820, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Gao, C.; Wang, M.; Fei, X.; Zhao, N. TRIM18-regulated STAT3 signaling pathway via PTP1B promotes renal epithelial-mesenchymal transition, inflammation, and fibrosis in diabetic kidney disease. Front. Physiol. 2021, 12, 709506. [Google Scholar] [CrossRef]
- Zheng, C.; Huang, L.; Luo, W.; Yu, W.; Hu, X.; Guan, X.; Cai, Y.; Zou, C.; Yin, H.; Xu, Z.; et al. Inhibition of STAT3 in tubular epithelial cells prevents kidney fibrosis and nephropathy in STZ-induced diabetic mice. Cell Death Dis. 2019, 10, 848. [Google Scholar] [CrossRef]
- Stemmer, K.; Perez-Tilve, D.; Ananthakrishnan, G.; Bort, A.; Seeley, R.J.; Tschöp, M.H.; Dietrich, D.R.; Pfluger, P.T. High-fat-diet-induced obesity causes an inflammatory and tumor-promoting microenvironment in the rat kidney. Dis. Models Mech. 2012, 5, 627–635. [Google Scholar] [CrossRef]
- Hung, L.H.; Wu, C.H.; Lin, B.F.; Hwang, L.S. Hyperimmune colostrum alleviates rheumatoid arthritis in a collagen-induced arthritis murine model. J. Dairy Sci. 2018, 101, 3778–3787. [Google Scholar] [CrossRef]
- Ojo, B.A.; O’Hara, C.; Wu, L.; El-Rassi, G.D.; Ritchey, J.W.; Chowanadisai, W.; Lin, D.; Smith, B.J.; Lucas, E.A. Wheat germ supplementation increases Lactobacillaceae and promotes an anti-inflammatory gut milieu in C57BL/6 mice fed a high-fat, high-sucrose diet. J. Nutr. 2019, 149, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K. Association of low potassium diet and folic acid deficiency in patients with CKD. Ther. Clin. Risk Manag. 2015, 11, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, T.; Kiyohara, Y.; Kubo, M.; Tanizaki, Y.; Tanaka, K.; Okubo, K.; Nakamura, H.; Hata, J.; Oishi, Y.; Kato, I.; et al. Hyperhomocysteinemia and the development of chronic kidney disease in a general population: The Hisayama study. Am. J. Kidney Dis. 2004, 44, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Y.; Tsai, Y.W.; Chen, S.Y.; Ho, C.I.; Weng, Y.M.; Hsiao, C.T.; Li, W.C. The association of leptin and homocysteine with renal function impairment in a population of Taiwanese adults. Clin. Nutr. 2015, 34, 943–950. [Google Scholar] [CrossRef]
- Cianciolo, G.; De Pascalis, A.; Di Lullo, L.; Ronco, C.; Zannini, C.; LaManna, G. Folic acid and homocysteine in chronic kidney disease and cardiovascular disease progression: Which comes first? Cardiorenal Med. 2017, 7, 255–266. [Google Scholar] [CrossRef]
- Ejerblad, E.; Fored, C.M.; Lindblad, P.; Fryzek, J.; McLaughlin, J.K.; Nyrén, O. Obesity and risk for chronic renal failure. J. Am. Soc. Nephrol. 2006, 17, 1695–1702. [Google Scholar] [CrossRef]
- Sun, X.; Han, F.; Miao, W.; Hou, N.; Cao, Z.; Zhang, G. Sonographic evaluation of para- and perirenal fat thickness is an independent predictor of early kidney damage in obese patients. Int. Urol. Nephrol. 2013, 45, 1589–1595. [Google Scholar] [CrossRef]
- Zhao, M.; Yuan, M.M.; Yuan, L.; Huang, L.L.; Liao, J.H.; Yu, X.L.; Su, C.; Chen, Y.H.; Yang, Y.Y.; Yu, H.; et al. Chronic folate deficiency induces glucose and lipid metabolism disorders and subsequent cognitive dysfunction in mice. PLoS ONE 2018, 13, e0202910. [Google Scholar] [CrossRef]
- Najib, S.; Sánchez-Margalet, V. Homocysteine thiolactone inhibits insulin-stimulated DNA and protein synthesis: Possible role of mitogen-activated protein kinase (MAPK), glycogen synthase kinase-3 (GSK-3) and p70 S6K phosphorylation. J. Mol. Endocrinol. 2005, 34, 119–126. [Google Scholar] [CrossRef]
- Li, H.; Lewis, A.; Brodsky, S.; Rieger, R.; Iden, C.; Goligorsky, M.S. Homocysteine induces 3-hydroxy-3-methylglutaryl coenzyme a reductase in vascular endothelial cells: A mechanism for development of atherosclerosis? Circulation 2002, 105, 1037–1043. [Google Scholar] [CrossRef]
- Buchan, L.; St Aubin, C.R.; Fisher, A.L.; Hellings, A.; Castro, M.; Al-Nakkash, L.; Broderick, T.L.; Plochocki, J.H. High-fat, high-sugar diet induces splenomegaly that is ameliorated with exercise and genistein treatment. BMC Res. Notes 2018, 11, 752. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Zeng, L.; Zheng, C.; Song, B.; Li, F.; Kong, X.; Xu, K. Inflammatory links between high fat diets and diseases. Front. Immunol. 2018, 9, 2649. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, K.; Inoue, M.; Masaki, T.; Chiba, S.; Shiraishi, K.; Shimasaki, T.; Matsuoka, K.; Ando, H.; Fujiwara, K.; Fukunaga, N.; et al. Obesity-related chronic kidney disease is associated with spleen-derived IL-10. Nephrol. Dial. Transplant. 2013, 28, 1120–1130. [Google Scholar] [CrossRef] [PubMed]
- Spranger, J.; Kroke, A.; Möhlig, M.; Hoffmann, K.; Bergmann, M.M.; Ristow, M.; Boeing, H.; Pfeiffer, A.F. Inflammatory cytokines and the risk to develop type 2 diabetes: Results of the prospective population-based European Prospective Investigation into Cancer and Nutrition (EPIC)-Potsdam Study. Diabetes 2003, 52, 812–817. [Google Scholar] [CrossRef]
- Stasi, A.; Cosola, C.; Caggiano, G.; Cimmarusti, M.T.; Palieri, R.; Acquaviva, P.M.; Rana, G.; Gesualdo, L. Obesity-related chronic kidney disease: Principal mechanisms and new approaches in nutritional management. Front. Nutr. 2022, 9, 925619. [Google Scholar] [CrossRef]
- Shankar, A.; Syamala, S.; Xiao, J.; Muntner, P. Relationship between plasma leptin level and chronic kidney disease. Int. J. Nephrol. 2012, 2012, 269532. [Google Scholar] [CrossRef]
- Xu, T.; Sheng, Z.; Yao, L. Obesity-related glomerulopathy: Pathogenesis, pathologic, clinical characteristics and treatment. Front. Med. 2017, 11, 340–348. [Google Scholar] [CrossRef]
- van der Heijden, R.A.; Bijzet, J.; Meijers, W.C.; Yakala, G.K.; Kleemann, R.; Nguyen, T.Q.; de Boer, R.A.; Schalkwijk, C.G.; Hazenberg, B.P.; Tietge, U.J.; et al. Obesity-induced chronic inflammation in high fat diet challenged C57BL/6J mice is associated with acceleration of age-dependent renal amyloidosis. Sci. Rep. 2015, 5, 16474. [Google Scholar] [CrossRef]
- Gu, Y.Y.; Liu, X.S.; Huang, X.R.; Yu, X.Q.; Lan, H.Y. Diverse role of TGF-β in kidney disease. Front. Cell Dev. Biol. 2020, 8, 123. [Google Scholar] [CrossRef]
- Mou, X.; Zhou, D.Y.; Zhou, D.Y.; Ma, J.R.; Liu, Y.H.; Chen, H.P.; Hu, Y.B.; Shou, C.M.; Chen, J.W.; Liu, W.H.; et al. Serum TGF-β1 as a biomarker for type 2 diabetic nephropathy: A meta-analysis of randomized controlled trials. PLoS ONE 2016, 11, e0149513. [Google Scholar] [CrossRef]
- Muñoz-Félix, J.M.; Martínez-Salgado, C. Dissecting the involvement of Ras GTPases in kidney fibrosis. Genes 2021, 12, 800. [Google Scholar] [CrossRef] [PubMed]
- Rozen-Zvi, B.; Hayashida, T.; Hubchak, S.C.; Hanna, C.; Platanias, L.C.; Schnaper, H.W. TGF-β/Smad3 activates mammalian target of rapamycin complex-1 to promote collagen production by increasing HIF-1α expression. Am. J. Physiol. Ren. Physiol. 2013, 305, F485–F494. [Google Scholar] [CrossRef] [PubMed]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, C.-W.; Lin, B.-F. Folate Deficiency Enhanced Inflammation and Exacerbated Renal Fibrosis in High-Fat High-Fructose Diet-Fed Mice. Nutrients 2023, 15, 3616. https://doi.org/10.3390/nu15163616
Chan C-W, Lin B-F. Folate Deficiency Enhanced Inflammation and Exacerbated Renal Fibrosis in High-Fat High-Fructose Diet-Fed Mice. Nutrients. 2023; 15(16):3616. https://doi.org/10.3390/nu15163616
Chicago/Turabian StyleChan, Chun-Wai, and Bi-Fong Lin. 2023. "Folate Deficiency Enhanced Inflammation and Exacerbated Renal Fibrosis in High-Fat High-Fructose Diet-Fed Mice" Nutrients 15, no. 16: 3616. https://doi.org/10.3390/nu15163616
APA StyleChan, C.-W., & Lin, B.-F. (2023). Folate Deficiency Enhanced Inflammation and Exacerbated Renal Fibrosis in High-Fat High-Fructose Diet-Fed Mice. Nutrients, 15(16), 3616. https://doi.org/10.3390/nu15163616