
The Effect of Delivery Matrix on Bifidobacterium animalis subsp. lactis HN019 Survival through In Vitro Human Digestion

Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Food Matrices
2.3. Upper Gastro-Intestinal (GI) Digestion and Colon Simulation
2.4. DNA Extraction
2.5. qPCR: Bifidobacterium animalis subsp. lactis HN019 Primer Design
2.6. qPCR: Assay Conditions
2.7. Short-Chain, Branched-Chain Fatty Acid and Lactic Acid Quantification
2.8. Statistical Analysis
3. Results
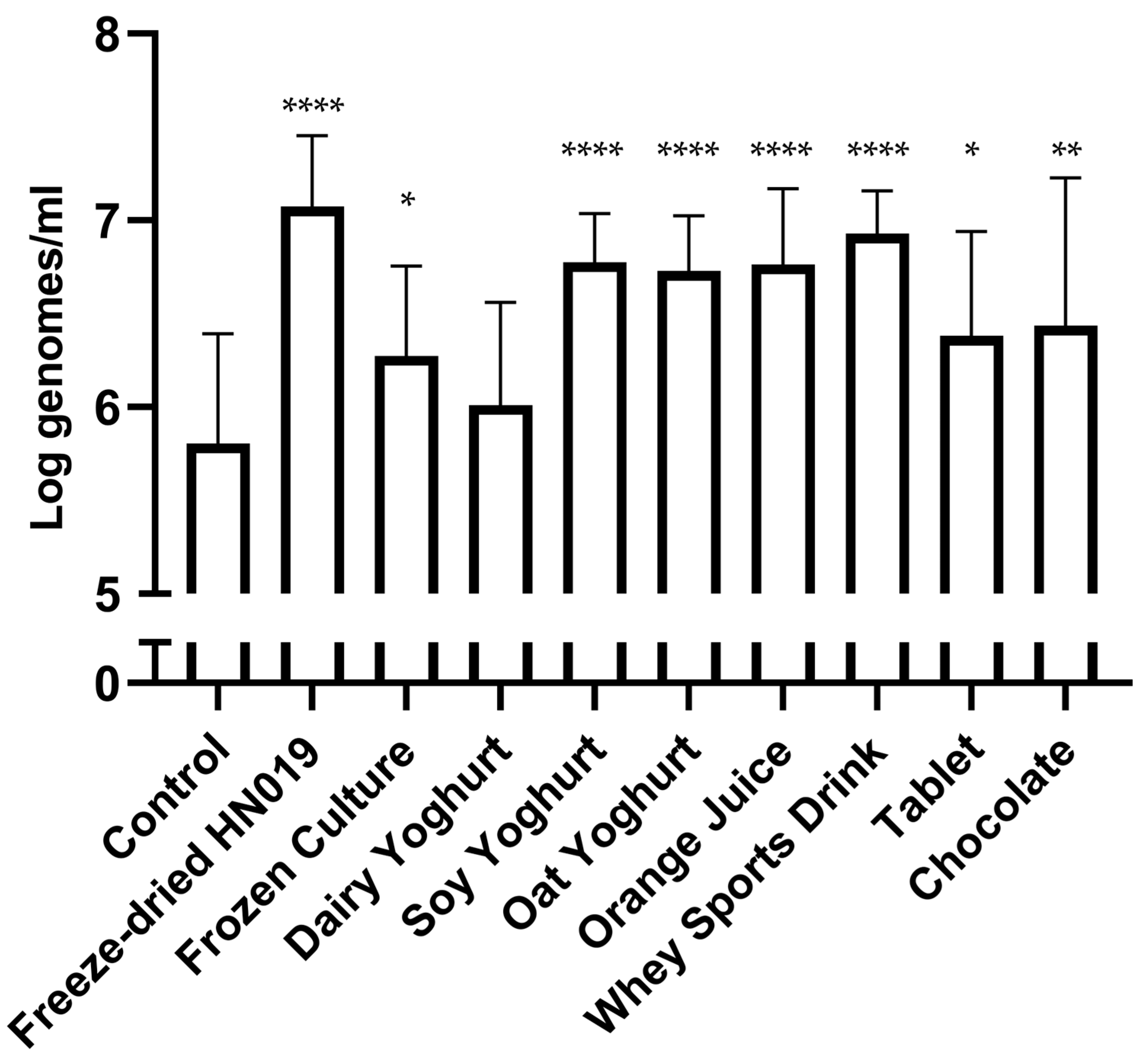
3.1. HN019 DNA Recovery
3.2. Short-Chain Fatty Acids
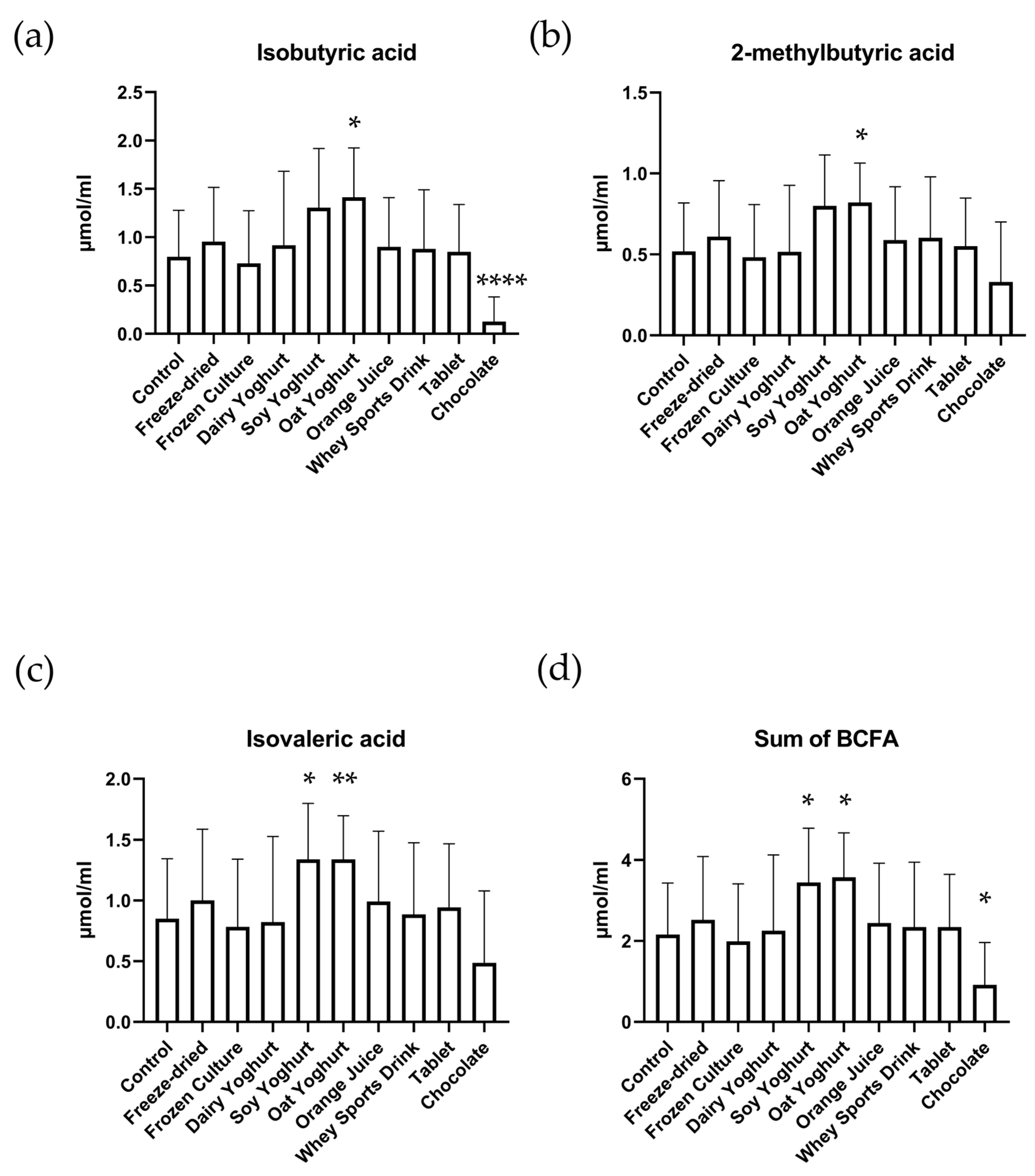
3.3. Branched-Chain Fatty Acids
3.4. Lactic Acid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miller, L.E.; Lehtoranta, L.; Lehtinen, M.J. The Effect of Bifidobacterium animalis ssp. lactis HN019 on Cellular Immune Function in Healthy Elderly Subjects: Systematic Review and Meta-Analysis. Nutrients 2017, 9, 191. [Google Scholar] [CrossRef]
- Cheng, J.; Laitila, A.; Ouwehand, A.C. Bifidobacterium animalis subsp. lactis HN019 Effects on Gut Health: A Review. Front. Nutr. 2021, 8, 790561. [Google Scholar] [CrossRef] [PubMed]
- Hemalatha, R.; Ouwehand, C.A.; Forssten, D.S.; Geddan, J.J.B.; Mamidi, R.; Bhaskar, V.; Radhakrishna, K.V. A Community-based Randomized Double Blind Controlled Trial of Lactobacillus paracasei and Bifidobacterium lactis on Reducing Risk for Diarrhea and Fever in Preschool Children in an Urban Slum in India. Eur. J. Nutr. Food Saf. 2014, 4, 325–341. [Google Scholar] [CrossRef]
- Sazawal, S.; Dhingra, U.; Hiremath, G.; Sarkar, A.; Dhingra, P.; Dutta, A.; Verma, P.; Menon, V.P.; Black, R.E. Prebiotic and probiotic fortified milk in prevention of morbidities among children: Community-based, randomized, double-blind, controlled trial. PLoS ONE 2010, 5, e12164. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Champagne, C.P.; Gardner, N.J.; Roy, D. Challenges in the addition of probiotic cultures to foods. Crit. Rev. Food Sci. Nutr. 2005, 45, 61–84. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Marco, M.L. Food formats for effective delivery of probiotics. Annu. Rev. Food Sci. Technol. 2010, 1, 65–85. [Google Scholar] [CrossRef]
- Mäkivuokko, H.; Kettunen, H.; Saarinen, M.; Kamiwaki, T.; Yokoyama, Y.; Stowell, J.; Rautonen, N. The effect of cocoa and polydextrose on bacterial fermentation in gastrointestinal tract simulations. Biosci. Biotechnol. Biochem. 2007, 71, 1834–1843. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mäkeläinen, H.; Forssten, S.; Olli, K.; Granlund, L.; Rautonen, N.; Ouwehand, A.C. Probiotic lactobacilli in a semi-soft cheese survive in the simulated human gastrointestinal tract. Int. Dairy J. 2009, 19, 675–683. [Google Scholar] [CrossRef]
- Airaksinen, K.; Yeung, N.; Lyra, A.; Lahtinen, S.J.; Huttunen, T.; Shanahan, F.; Ouwehand, A.C. The effect of a probiotic blend on gastrointestinal symptoms in constipated patients: A double blind, randomised, placebo controlled 2-week trial. Benef. Microbes 2019, 10, 617–627. [Google Scholar] [CrossRef]
- Invernici, M.M.; Salvador, S.L.; Silva, P.H.F.; Soares, M.S.M.; Casarin, R.; Palioto, D.B.; Souza, S.L.S.; Taba, M., Jr.; Novaes, A.B., Jr.; Furlaneto, F.A.C.; et al. Effects of Bifidobacterium probiotic on the treatment of chronic periodontitis: A randomized clinical trial. J. Clin. Periodontol. 2018, 45, 1198–1210. [Google Scholar] [CrossRef] [PubMed]
- Arunachalam, K.; Gill, H.S.; Chandra, R.K. Enhancement of natural immune function by dietary consumption of Bifidobacterium lactis (HN019). Eur. J. Clin. Nutr. 2000, 54, 263–267. [Google Scholar] [PubMed]
- Chiang, B.L.; Sheih, Y.H.; Wang, L.H.; Liao, C.K.; Gill, H.S. Enhancing immunity by dietary consumption of a probiotic lactic acid bacterium (Bifidobacterium lactis HN019): Optimization and definition of cellular immune responses. Eur. J. Clin. Nutr. 2000, 54, 849–855. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.S.; Rutherfurd, K.J.; Prasad, J.; Gopal, P.K. Enhancement of natural and acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus acidophilus (HN017) and Bifidobacterium lactis (HN019). Br. J. Nutr. 2000, 83, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, A.; Latreille-Barbier, M.; Donazzolo, Y.; Pelletier, X.; Ouwehand, A.C. Effects of 28-day Bifidobacterium animalis subsp. lactis HN019 supplementation on colonic transit time and gastrointestinal symptoms in adults with functional constipation: A double-blind, randomized, placebo-controlled, and dose-ranging trial. Gut Microbes 2017, 9, 236–251. [Google Scholar] [CrossRef]
- Magro, D.O.; de Oliveira, L.M.; Bernasconi, I.; Ruela Mde, S.; Credidio, L.; Barcelos, I.K.; Leal, R.F.; Ayrizono Mde, L.; Fagundes, J.J.; Teixeira Lde, B.; et al. Effect of yogurt containing polydextrose, Lactobacillus acidophilus NCFM and Bifidobacterium lactis HN019: A randomized, double-blind, controlled study in chronic constipation. Nutr. J. 2014, 13, 75. [Google Scholar] [CrossRef]
- Waller, P.A.; Gopal, P.K.; Leyer, G.J.; Ouwehand, A.C.; Reifer, C.; Stewart, M.E.; Miller, L.E. Dose-response effect of Bifidobacterium lactis HN019 on whole gut transit time and functional gastrointestinal symptoms in adults. Scand. J. Gastroenterol. 2011, 46, 1057–1064. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a Three-Stage Compound Continuous Culture System for Investigating the Effect of Retention Time on the Ecology and Metabolism of Bacteria in the Human Colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Mäkivuokko, H.; Nurmi, J.; Nurminen, P.; Stowell, J.; Rautonen, N. In vitro effects on polydextrose by colonic bacteria and caco-2 cell cyclooxygenase gene expression. Nutr. Cancer 2005, 52, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Forssten, S.D.; Ouwehand, A.C. Simulating colonic survival of probiotics in single-strain products compared to multi-strain products. Microb. Ecol. Health Dis. 2017, 28, 1378061. [Google Scholar] [CrossRef]
- Milani, C.; Duranti, S.; Lugli, G.A.; Bottacini, F.; Strati, F.; Arioli, S.; Foroni, E.; Turroni, F.; van Sinderen, D.; Ventura, M. Comparative genomics of Bifidobacterium animalis subsp. lactis reveals a strict monophyletic bifidobacterial taxon. Appl. Environ. Microbiol. 2013, 79, 4304–4315. [Google Scholar] [CrossRef]
- Morovic, W.; Roos, P.; Zabel, B.; Hidalgo-Cantabrana, C.; Kiefer, A.; Barrangou, R. Transcriptional and Functional Analysis of Bifidobacterium animalis subsp. lactis Exposure to Tetracycline. Appl. Environ. Microbiol. 2018, 84, e01999-18. [Google Scholar] [CrossRef]
- Lee, J.H.; O’Sullivan, D.J. Genomic insights into bifidobacteria. Microbiol. Mol. Biol. Rev. 2010, 74, 378–416. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Tiihonen, K.; Saarinen, M.; Putaala, H.; Rautonen, N. Influence of a combination of Lactobacillus acidophilus NCFM and lactitol on healthy elderly: Intestinal and immune parameters. Br. J. Nutr. 2009, 101, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, Y.; Dong, P.; An, R.; Xue, C.; Ge, Y.; Wei, L.; Liang, X. Digestion of Nucleic Acids Starts in the Stomach. Sci. Rep. 2015, 5, 11936. [Google Scholar] [CrossRef]
- Stenman, L.K.; Lehtinen, M.J.; Meland, N.; Christensen, J.E.; Yeung, N.; Saarinen, M.T.; Courtney, M.; Burcelin, R.; Lahdeaho, M.L.; Linros, J.; et al. Probiotic With or Without Fiber Controls Body Fat Mass, Associated With Serum Zonulin, in Overweight and Obese Adults-Randomized Controlled Trial. EBioMedicine 2016, 13, 190–200. [Google Scholar] [CrossRef]
- Savard, P.; Lamarche, B.; Paradis, M.E.; Thiboutot, H.; Laurin, E.; Roy, D. Impact of Bifidobacterium animalis subsp. lactis BB-12 and, Lactobacillus acidophilus LA-5-containing yoghurt, on fecal bacterial counts of healthy adults. Int. J. Food. Microbiol. 2011, 149, 50–57. [Google Scholar] [CrossRef]
- Kundi, Z.M.; Lee, J.C.; Pihlajamaki, J.; Chan, C.B.; Leung, K.S.; So, S.S.Y.; Nordlund, E.; Kolehmainen, M.; El-Nezami, H. Dietary Fiber from Oat and Rye Brans Ameliorate Western Diet-Induced Body Weight Gain and Hepatic Inflammation by the Modulation of Short-Chain Fatty Acids, Bile Acids, and Tryptophan Metabolism. Mol. Nutr. Food Res. 2021, 65, e1900580. [Google Scholar] [CrossRef]
- Lecumberri, E.; Goya, L.; Mateos, R.; Alia, M.; Ramos, S.; Izquierdo-Pulido, M.; Bravo, L. A diet rich in dietary fiber from cocoa improves lipid profile and reduces malondialdehyde in hypercholesterolemic rats. Nutrition 2007, 23, 332–341. [Google Scholar] [CrossRef]
- Hossain, M.N.; Senaka Ranadheera, C.; Fang, Z.; Ajlouni, S. Production of short chain fatty acids and vitamin B12 during the in-vitro digestion and fermentation of probiotic chocolate. Food Biosci. 2022, 47, 101682. [Google Scholar] [CrossRef]
- Duque, A.L.R.F.; Monteiro, M.; Adorno, M.A.T.; Sakamoto, I.K.; Sivieri, K. An exploratory study on the influence of orange juice on gut microbiota using a dynamic colonic model. Food Res. Int. 2016, 84, 160–169. [Google Scholar] [CrossRef]
- Smith, N.W.; Dave, A.C.; Hill, J.P.; McNabb, W.C. Nutritional assessment of plant-based beverages in comparison to bovine milk. Front. Nutr. 2022, 9, 957486. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Yao, G.; Sun, Y.; Guo, S.; Wang, J.; Mu, X.; Sun, Z.; Zhang, H. Comparative effects of the single and binary probiotics of Lacticaseibacillus casei Zhang and Bifidobacterium lactis V9 on the growth and metabolomic profiles in yogurts. Food Res. Int. 2022, 152, 110603. [Google Scholar] [CrossRef] [PubMed]
- Bourriaud, C.; Robins, R.J.; Martin, L.; Kozlowski, F.; Tenailleau, E.; Cherbut, C.; Michel, C. Lactate is mainly fermented to butyrate by human intestinal microfloras but inter-individual variation is evident. J. Appl. Microbiol. 2005, 99, 201–212. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Aryal, U.; Cooper, B.; Biswas, D. Metabolites produced during the growth of probiotics in cocoa supplementation and the limited role of cocoa in host-enteric bacterial pathogen interactions. Food Control 2015, 53, 124–133. [Google Scholar] [CrossRef]
- Kiefer, A.; Tang, P.; Arndt, S.; Fallico, V.; Wong, C. Optimization of Viability Treatment Essential for Accurate Droplet Digital PCR Enumeration of Probiotics. Front. Microbiol. 2020, 11, 1811. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food Matrix | Total CFU | Matrix Manufacturer | Other Major Ingredients |
|---|---|---|---|
| Frozen culture | 2.025 × 109 | IFF (Madison, WI, USA) | none |
| Freeze-dried powder | 2.025 × 109 | IFF (Madison, WI, USA) | none |
| Dairy yogurt | 2.025 × 109 | Valio (Helsinki, Finland) | pasteurized milk, starter culture, vitamin D |
| Non-dairy soy yogurt | 2.025 × 109 | Alpro (Wevelgem, Belgium) | soya base (water and hulled soya beans), sugar, tri-calcium citrate, pectins |
| Non-dairy oat yogurt | 2.025 × 109 | Fazer (Helsinki, Finland) | water, oat, sugar, corn starch, locust bean gum |
| Chocolate | 7.842 × 109 | IFF (Madison, WI, USA) | cocoa mass, sugar, cocoa butter, palm oil, vanilla (min. cocoa solids 70%) |
| Tablet | 1.32 × 109 | Wasa Medicals (Halmstad, Sweden) | inulin, xanthan gum, magnesium stearate |
| Orange juice (pulpless) | 2.025 × 109 | Eckes-Granini (Turku, Finland) | none |
| Whey sports drink | 2.025 × 109 | Midsona (Malmö, Sweden) | UHT-treated milk, maltodextrin, milk protein, sugar, cocoa |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yeung, N.; Forssten, S.D.; Saarinen, M.T.; Anjum, M.; Ouwehand, A.C. The Effect of Delivery Matrix on Bifidobacterium animalis subsp. lactis HN019 Survival through In Vitro Human Digestion. Nutrients 2023, 15, 3541. https://doi.org/10.3390/nu15163541
Yeung N, Forssten SD, Saarinen MT, Anjum M, Ouwehand AC. The Effect of Delivery Matrix on Bifidobacterium animalis subsp. lactis HN019 Survival through In Vitro Human Digestion. Nutrients. 2023; 15(16):3541. https://doi.org/10.3390/nu15163541
Chicago/Turabian StyleYeung, Nicolas, Sofia D. Forssten, Markku T. Saarinen, Mehreen Anjum, and Arthur C. Ouwehand. 2023. "The Effect of Delivery Matrix on Bifidobacterium animalis subsp. lactis HN019 Survival through In Vitro Human Digestion" Nutrients 15, no. 16: 3541. https://doi.org/10.3390/nu15163541
APA StyleYeung, N., Forssten, S. D., Saarinen, M. T., Anjum, M., & Ouwehand, A. C. (2023). The Effect of Delivery Matrix on Bifidobacterium animalis subsp. lactis HN019 Survival through In Vitro Human Digestion. Nutrients, 15(16), 3541. https://doi.org/10.3390/nu15163541