
Understanding the Roles of Very-Long-Chain Polyunsaturated Fatty Acids (VLC-PUFAs) in Eye Health


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Importance of VLC-PUFAs to Eye Health
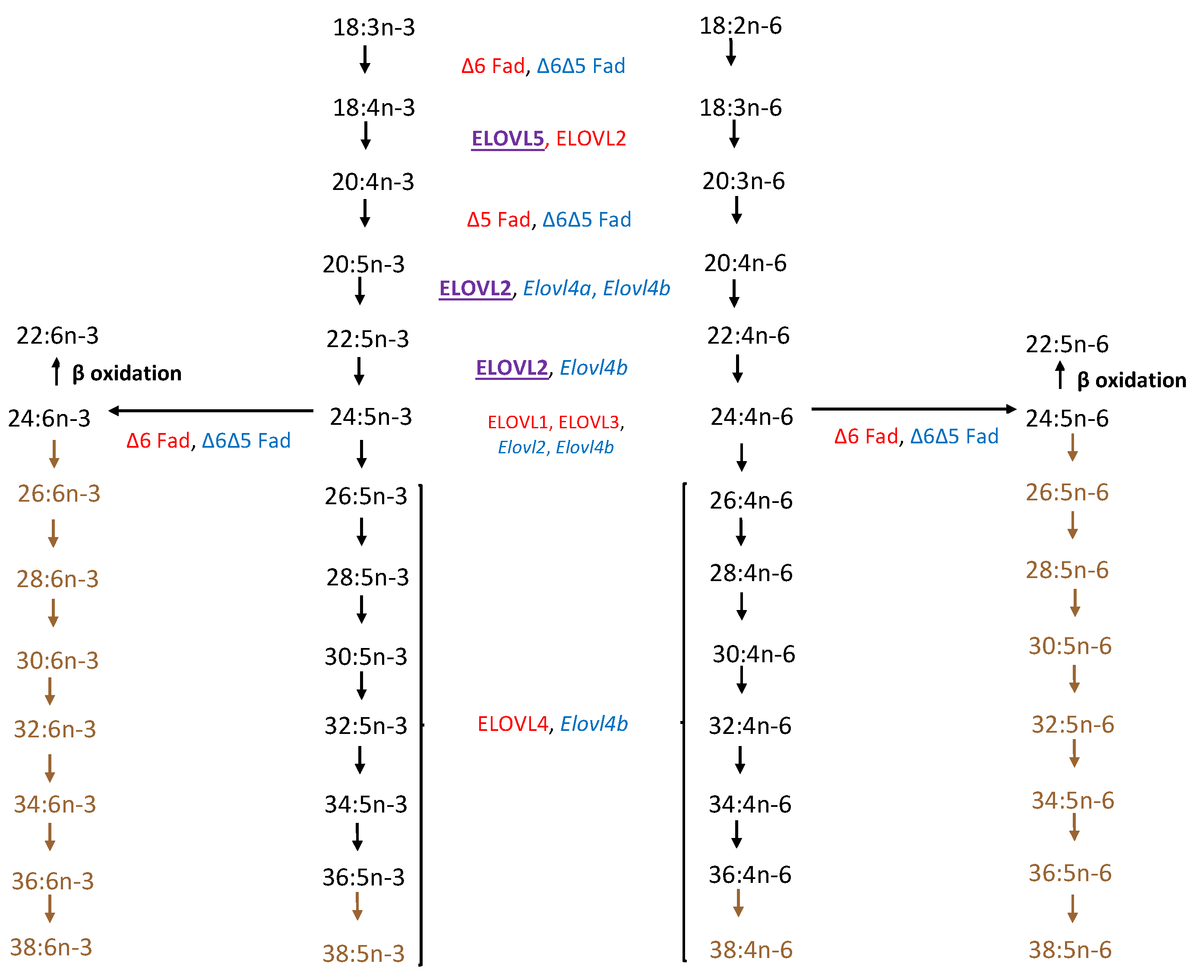
3. ELOVL4 Structure and Biosynthesis of VLC-PUFA
4. VLC-PUFAs
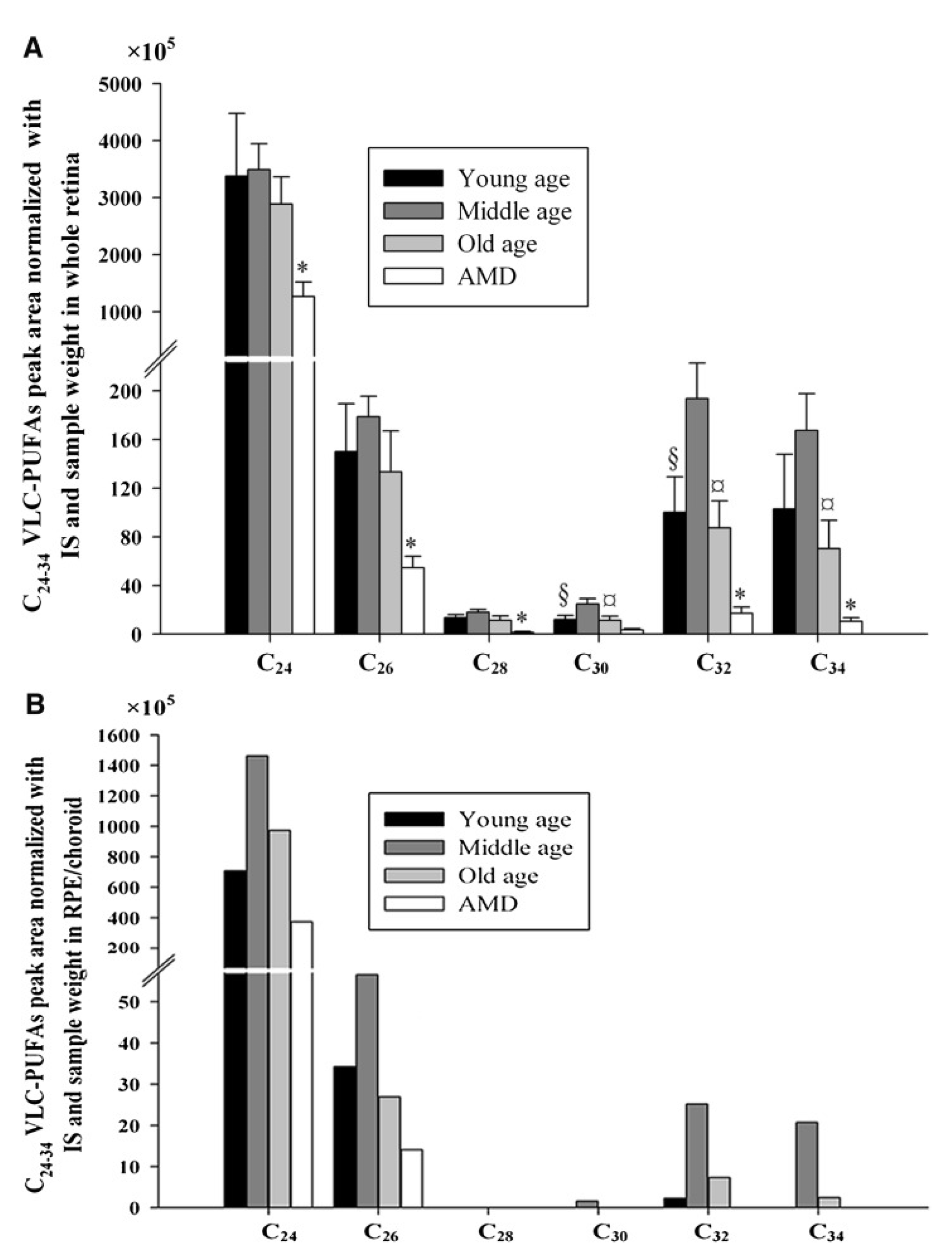
4.1. Localization of VLC-PUFAs in the Eye
4.2. Physiological Roles of VLC-PUFA
4.3. VLC-PUFAs in the RPE
4.4. VLC-PUFAs in Neurotransmission
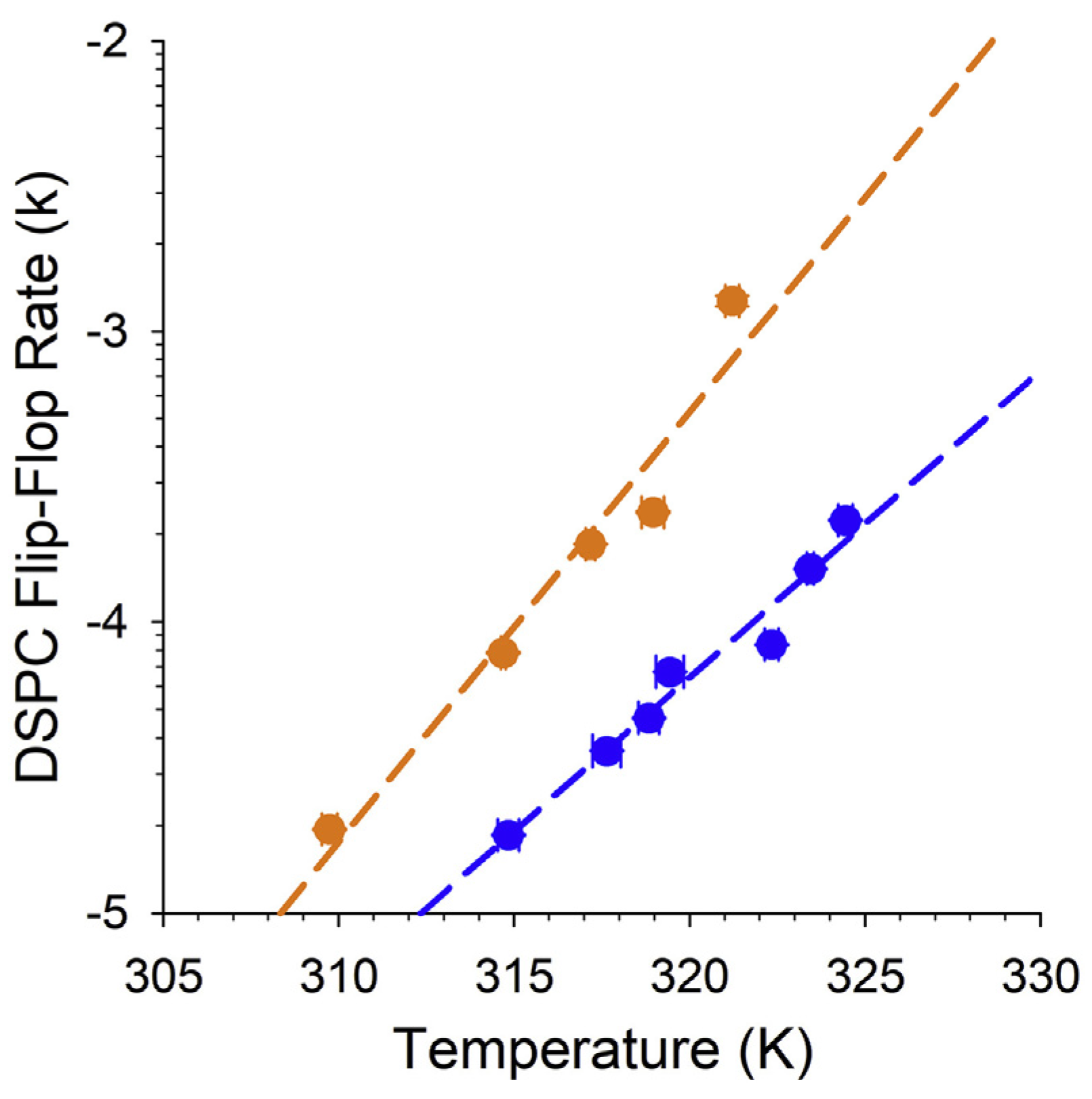
4.5. Biophysical Chemistry
4.6. ELOVL2 and VLC-PUFAs
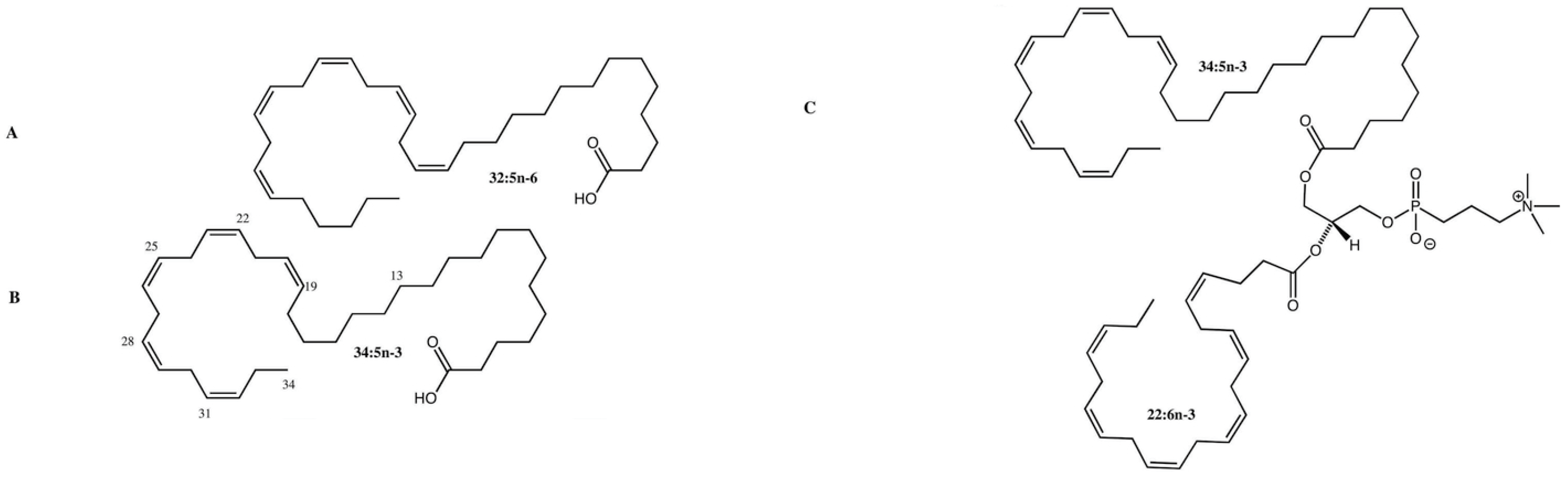
4.7. VLC-PUFA Derivatives
5. Dietary Studies
6. Cell Culture Studies and Substrate Specificity
7. Animal Models
8. VLC-PUFA Analysis Tools
8.1. GC-MS
8.2. LC-MS/MS
8.3. Other Analytical Methods
9. VLC-PUFAs in Relation to Cholesteryl Esters and Acylcarnitines in Eye Health
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aveldaño, M.I. A Novel Group of Very Long Chain Polyenoic Fatty Acids in Dipolyunsaturated Phosphatidylcholines from Vertebrate Retina. J. Biol. Chem. 1987, 262, 1172–1179. [Google Scholar] [CrossRef] [PubMed]
- Agbaga, M.-P.; Mandal, N.A.; Anderson, R.E. Retinal very long-chain PUFAs: New insights from studies on ELOVL4 protein. J. Lipid Res. 2010, 51, 1624–1642. [Google Scholar] [CrossRef] [PubMed]
- Aveldaño, M.I.; Sprecher, H. Very long chain (C24 to C36) polyenoic fatty acids of the n-3 and n-6 series in dipolyunsaturated phosphatidylcholines from bovine retina. J. Biol. Chem. 1987, 262, 1180–1186. [Google Scholar] [CrossRef] [PubMed]
- Berdeaux, O.; Juaneda, P.; Martine, L.; Cabaret, S.; Bretillon, L.; Acar, N. Identification and quantification of phosphatidylcholines containing very-long-chain polyunsaturated fatty acid in bovine and human retina using liquid chromatography/tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 7738–7748. [Google Scholar] [CrossRef]
- Takashima, S.; Takemoto, S.; Toyoshi, K.; Ohba, A.; Shimozawa, N. Zebrafish model of human Zellweger syndrome reveals organ-specific accumulation of distinct fatty acid species and widespread gene expression changes. Mol. Genet. Metab. 2021, 133, 307–323. [Google Scholar] [CrossRef]
- He, Y.; Phan, K.; Bhatia, S.; Pickford, R.; Fu, Y.; Yang, Y.; Hodges, J.R.; Piguet, O.; Halliday, G.M.; Kim, W.S. Increased VLCFA-lipids and ELOVL4 underlie neurodegeneration in frontotemporal dementia. Sci. Rep. 2021, 11, 21348. [Google Scholar] [CrossRef]
- Wade, A.; Rallabandi, R.; Lucas, S.; Oberg, C.; Gorusupudi, A.; Bernstein, P.S.; Rainier, J.D. The synthesis of the very long chain polyunsaturated fatty acid (VLC-PUFA) 32:6 n-3. Org. Biomol. Chem. 2021, 19, 5563–5566. [Google Scholar] [CrossRef]
- Gorusupudi, A.; Chang, F.; Nelson, K.; Hageman, G.S.; Bernstein, P.S. n-3 PUFA Supplementation Alters Retinal Very-Long-Chain-PUFA Levels and Ratios in Diabetic Animal Models. Mol. Nutr. Food Res. 2019, 63, e1801058. [Google Scholar] [CrossRef]
- Monroig, O.; Rotllant, J.; Cerdá-Reverter, J.M.; Dick, J.R.; Figueras, A.; Tocher, D.R. Expression and role of Elovl4 elongases in biosynthesis of very long-chain fatty acids during zebrafish Danio rerio early embryonic development. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2010, 1801, 1145–1154. [Google Scholar] [CrossRef]
- Hopiavuori, B.R.; Anderson, R.E.; Agbaga, M.-P. ELOVL4: Very long-chain fatty acids serve an eclectic role in mammalian health and function. Prog. Retin. Eye Res. 2019, 69, 137–158. [Google Scholar] [CrossRef]
- Lawrenson, J.G.; Evans, J.R. Omega 3 fatty acids for preventing or slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2015, 2015, CD010015. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, A.F.; Askew, E.W.; Singh, N.; Leppert, M.; Bernstein, P.S. Association of Adipose and Red Blood Cell Lipids With Severity of Dominant Stargardt Macular Dystrophy (STGD3) Secondary to an ELOVL4 Mutation. Arch. Ophthalmol. 2006, 124, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Sala-Vila, A.; Díaz-López, A.; Valls-Pedret, C.; Cofán, M.; García-Layana, A.; Lamuela-Raventós, R.M.; Castañer, O.; Zanon-Moreno, V.; Martinez-Gonzalez, M.A.; Toledo, E.; et al. Dietary Marine ω-3 Fatty Acids and Incident Sight-Threatening Retinopathy in Middle-Aged and Older Individuals with Type 2 Diabetes: Prospective Investigation From the PREDIMED Trial. JAMA Ophthalmol. 2016, 134, 1142–1149. [Google Scholar] [CrossRef]
- Berson, E.L.; Rosner, B.; Sandberg, M.A.; Weigel-DiFranco, C.; Willett, W.C. ω-3 Intake and Visual Acuity in Patients With Retinitis Pigmentosa Receiving Vitamin A. JAMA Ophthalmol 2012, 130, 707–711. [Google Scholar] [CrossRef]
- Arunkumar, R.; Gorusupudi, A.; Li, B.; Blount, J.D.; Nwagbo, U.; Kim, H.J.; Sparrow, J.R.; Bernstein, P.S. Lutein and zeaxanthin reduce A2E and iso-A2E levels and improve visual performance in Abca4/Bco2 double knockout mice. Exp. Eye Res. 2021, 209, 108680. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Chang, J.; Lin, Y.; Shen, Z.; Bernstein, P.S. Long-chain and very long-chain polyunsaturated fatty acids in ocular aging and age-related macular degeneration. J. Lipid Res. 2010, 51, 3217–3229. [Google Scholar] [CrossRef] [PubMed]
- Bazan, H.E.; Bazan, N.G.; Feeney-Burns, L.; Berman, E.R. Lipids in human lipofuscin-enriched subcellular fractions of two age populations. Comparison with rod outer segments and neural retina. Investig. Opthalmol. Vis. Sci. 1990, 31, 1433–1443. [Google Scholar]
- Kihara, A. Very long-chain fatty acids: Elongation, physiology and related disorders. J. Biochem. 2012, 152, 387–395. [Google Scholar] [CrossRef]
- Agbaga, M.-P.; Brush, R.S.; Mandal, N.A.; Henry, K.; Elliott, M.H.; Anderson, R.E. Role of Stargardt-3 macular dystrophy protein (ELOVL4) in the biosynthesis of very long chain fatty acids. Proc. Natl. Acad. Sci. USA 2008, 105, 12843–12848. [Google Scholar] [CrossRef]
- Martin, F.J.; Amode, M.R.; Aneja, A.; Austine-Orimoloye, O.; Azov, A.G.; Barnes, I.; Becker, A.; Bennett, R.; Berry, A.; Bhai, J.; et al. Ensembl 2023. Nucleic Acids Res. 2023, 51, D933–D941. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Farrell, C.M.; Feldgarden, M.; Fine, A.M.; Funk, K.; et al. Database Resources of the National Center for Biotechnology Information in 2023. Nucleic Acids Res. 2023, 51, D29–D38. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; Scimone, C.; Rinaldi, C.; Aragona, P.; Briuglia, S.; D’Ascola, A.; D’Angelo, R.; Sidoti, A. Stargardt Phenotype Associated With Two ELOVL4 Promoter Variants and ELOVL4 Downregulation: New Possible Perspective to Etiopathogenesis? Investig. Opthalmol. Vis. Sci. 2018, 59, 843–857. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Gao, F.; Li, J.; Xu, P.; Wang, D.; Chen, F.; Zhang, S.; Wu, J. Novel variants associated with Stargardt disease in Chinese patients. Gene 2020, 754, 144890. [Google Scholar] [CrossRef]
- Sassa, T.; Kihara, A. Metabolism of Very Long-Chain Fatty Acids: Genes and Pathophysiology. Biomol. Ther. 2014, 22, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Sherry, D.M.; Hopiavuori, B.R.; Stiles, M.A.; Rahman, N.S.; Ozan, K.G.; Deak, F.; Agbaga, M.-P.; Anderson, R.E. Distribution of ELOVL4 in the Developing and Adult Mouse Brain. Front. Neuroanat. 2017, 11, 38. [Google Scholar] [CrossRef]
- Mukherjee, S.; Roy, M.; Ghosh, S.; Guha, G.; Saha, S.P.; Dalal, A. Rare mutation in ELOVL4 gene in SCA34 and cognitive affection: Expounding the role of cerebellum. Clin. Neurol. Neurosurg. 2021, 210, 106983. [Google Scholar] [CrossRef]
- Diociaiuti, A.; Martinelli, D.; Nicita, F.; Cesario, C.; Pisaneschi, E.; Macchiaiolo, M.; Rossi, S.; Condorelli, A.G.; Zambruno, G.; El Hachem, M. Two Italian Patients with ELOVL4-Related Neuro-Ichthyosis: Expanding the Genotypic and Phenotypic Spectrum and Ultrastructural Characterization. Genes 2021, 12, 343. [Google Scholar] [CrossRef]
- Agbaga, M.-P.; Stiles, M.A.; Brush, R.S.; Sullivan, M.T.; Machalinski, A.; Jones, K.L.; Anderson, R.E.; Sherry, D.M. The Elovl4 Spinocerebellar Ataxia-34 Mutation 736T>G (p.W246G) Impairs Retinal Function in the Absence of Photoreceptor Degeneration. Mol. Neurobiol. 2020, 57, 4735–4753. [Google Scholar] [CrossRef]
- Xiao, C.; Binkley, E.M.; Rexach, J.; Knight-Johnson, A.; Khemani, P.; Fogel, B.L.; Das, S.; Stone, E.M.; Gomez, C.M. A family with spinocerebellar ataxia and retinitis pigmentosa attributed to an ELOVL4 mutation. Neurol. Genet. 2019, 5, e357. [Google Scholar] [CrossRef]
- McMahon, A.; Lu, H.; Butovich, I.A. A Role for ELOVL4 in the Mouse Meibomian Gland and Sebocyte Cell Biology. Investig. Opthalmol. Vis. Sci. 2014, 55, 2832–2840. [Google Scholar] [CrossRef]
- Nagaraja, R.Y.; Sherry, D.M.; Fessler, J.L.; Stiles, M.A.; Li, F.; Multani, K.; Orock, A.; Ahmad, M.; Brush, R.S.; Anderson, R.E.; et al. W246G Mutant ELOVL4 Impairs Synaptic Plasticity in Parallel and Climbing Fibers and Causes Motor Defects in a Rat Model of SCA34. Mol. Neurobiol. 2021, 58, 4921–4943. [Google Scholar] [CrossRef] [PubMed]
- McMahon, A.; Butovich, I.A.; Mata, N.L.; Klein, M.; Ritter, R.; Richardson, J.; Birch, D.G.; Edwards, A.O.; Kedzierski, W. Retinal pathology and skin barrier defect in mice carrying a Stargardt disease-3 mutation in elongase of very long chain fatty acids-4. Mol. Vis. 2007, 13, 258–272. [Google Scholar]
- Nie, L.; Pascoa, T.C.; Pike, A.C.W.; Bushell, S.R.; Quigley, A.; Ruda, G.F.; Chu, A.; Cole, V.; Speedman, D.; Moreira, T.; et al. The structural basis of fatty acid elongation by the ELOVL elongases. Nat. Struct. Mol. Biol. 2021, 28, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Torrissen, M.; Svensen, H.; Stoknes, I.; Nilsson, A.; Østbye, T.-K.; Berge, G.M.; Bou, M.; Ruyter, B. Deposition and metabolism of dietary n-3 very-long-chain PUFA in different organs of rat, mouse and Atlantic salmon. Br. J. Nutr. 2022, 127, 35–54. [Google Scholar] [CrossRef] [PubMed]
- Serrano, R.; Navarro, J.C.; Portolés, T.; Sales, C.; Beltrán, J.; Monroig; Hernández, F. Identification of new, very long-chain polyunsaturated fatty acids in fish by gas chromatography coupled to quadrupole/time-of-flight mass spectrometry with atmospheric pressure chemical ionization. Anal. Bioanal. Chem. 2021, 413, 1039–1046. [Google Scholar] [CrossRef]
- Sandhoff, R. Very long chain sphingolipids: Tissue expression, function and synthesis. FEBS Lett. 2009, 584, 1907–1913. [Google Scholar] [CrossRef]
- Li, W.; Sandhoff, R.; Kono, M.; Zerfas, P.; Hoffmann, V.; Ding, B.C.-H.; Proia, R.L.; Deng, C.-X. Depletion of ceramides with very long chain fatty acids causes defective skin permeability barrier function, and neonatal lethality in ELOVL4 deficient mice. Int. J. Biol. Sci. 2007, 3, 120–128. [Google Scholar] [CrossRef]
- Verra, D.M.; Spinnhirny, P.; Sandu, C.; Grégoire, S.; Acar, N.; Berdeaux, O.; Brétillon, L.; Sparrow, J.R.; Hicks, D. Intrinsic differences in rod and cone membrane composition: Implications for cone degeneration. Graefe’s Arch. Clin. Exp. Ophthalmol. 2022, 260, 3131–3148. [Google Scholar] [CrossRef]
- Agbaga, M.-P.; Merriman, D.K.; Brush, R.S.; Lydic, T.A.; Conley, S.M.; Naash, M.I.; Jackson, S.; Woods, A.S.; Reid, G.E.; Busik, J.V.; et al. Differential composition of DHA and very-long-chain PUFAs in rod and cone photoreceptors. J. Lipid Res. 2018, 59, 1586–1596. [Google Scholar] [CrossRef]
- Berry, K.A.Z.; Gordon, W.C.; Murphy, R.C.; Bazan, N.G. Spatial organization of lipids in the human retina and optic nerve by MALDI imaging mass spectrometry. J. Lipid Res. 2014, 55, 504–515. [Google Scholar] [CrossRef]
- Moser, A.E.; Singh, I.; Brown, F.R.; Solish, G.I.; Kelley, R.I.; Benke, P.J.; Moser, H.W. The Cerebrohepatorenal (Zellweger) Syndrome. N. Engl. J. Med. 1984, 310, 1141–1146. [Google Scholar] [CrossRef] [PubMed]
- Shen, O.; Michaelson-Cohen, R.; Gross-Tsur, V.; Eilat, A.; Yanai, N.; Green, T.; Rabinowitz, R.; Meiner, V. Prenatal observation of nystagmus, cataracts, and brain abnormalities in a case of Zellweger spectrum disorder syndrome. Prenat. Diagn. 2016, 36, 894–895. [Google Scholar] [CrossRef] [PubMed]
- Braverman, N.E.; Raymond, G.V.; Rizzo, W.B.; Moser, A.B.; Wilkinson, M.E.; Stone, E.M.; Steinberg, S.J.; Wangler, M.F.; Rush, E.T.; Hacia, J.G.; et al. Peroxisome biogenesis disorders in the Zellweger spectrum: An overview of current diagnosis, clinical manifestations, and treatment guidelines. Mol. Genet. Metab. 2016, 117, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Swinkels, D.; Das, Y.; Kocherlakota, S.; Vinckier, S.; Wever, E.; van Kampen, A.H.; Vaz, F.M.; Baes, M. Cell Type-Selective Loss of Peroxisomal β-Oxidation Impairs Bipolar Cell but Not Photoreceptor Survival in the Retina. Cells 2022, 11, 161. [Google Scholar] [CrossRef] [PubMed]
- Dejos, C.; Kuny, S.; Han, W.H.; Capel, H.; Lemieux, H.; Sauvé, Y. Photoreceptor-induced RPE phagolysosomal maturation defects in Stargardt-like Maculopathy (STGD3). Sci. Rep. 2018, 8, 5944. [Google Scholar] [CrossRef] [PubMed]
- Esteve-Rudd, J.; Hazim, R.A.; Diemer, T.; Paniagua, A.E.; Volland, S.; Umapathy, A.; Williams, D.S. Defective phagosome motility and degradation in cell nonautonomous RPE pathogenesis of a dominant macular degeneration. Proc. Natl. Acad. Sci. USA 2018, 115, 5468–5473. [Google Scholar] [CrossRef]
- Chen, D.; Chao, D.L.; Rocha, L.; Kolar, M.; Huu, V.A.N.; Krawczyk, M.; Dasyani, M.; Wang, T.; Jafari, M.; Jabari, M.; et al. The lipid elongation enzyme ELOVL2 is a molecular regulator of aging in the retina. Aging Cell 2020, 19, e13100. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Wang, L.; Gao, Y.; Feng, G.; Li, G.; Zou, J.; Yu, M.; Li, Y.F.; Liu, C.; et al. Lipid metabolism dysfunction induced by age-dependent DNA methylation accelerates aging. Signal Transduct. Target. Ther. 2022, 7, 162. [Google Scholar] [CrossRef]
- Cheng, S.-Y.; Cipi, J.; Ma, S.; Hafler, B.P.; Kanadia, R.N.; Brush, R.S.; Agbaga, M.-P.; Punzo, C. Altered photoreceptor metabolism in mouse causes late stage age-related macular degeneration-like pathologies. Proc. Natl. Acad. Sci. USA 2020, 117, 13094–13104. [Google Scholar] [CrossRef]
- Bennett, L.D.; Brush, R.S.; Chan, M.; Lydic, T.A.; Reese, K.; Reid, G.E.; Busik, J.V.; Elliott, M.H.; Anderson, R.E. Effect of Reduced Retinal VLC-PUFA on Rod and Cone Photoreceptors. Investig. Opthalmol. Vis. Sci. 2014, 55, 3150–3157. [Google Scholar] [CrossRef]
- Aveldano, M.I. Phospholipid species containing long and very long polyenoic fatty acids remain with rhodopsin after hexane extraction of photoreceptor membranes. Biochemistry 1988, 27, 1229–1239. [Google Scholar] [CrossRef] [PubMed]
- Cheng, V.; Rallabandi, R.; Gorusupudi, A.; Lucas, S.; Rognon, G.; Bernstein, P.S.; Rainier, J.D.; Conboy, J.C. Influence of very-long-chain polyunsaturated fatty acids on membrane structure and dynamics. Biophys. J. 2022, 121, 2730–2741. [Google Scholar] [CrossRef]
- McMahon, A.; Kedzierski, W. Polyunsaturated very-long-chain C28-C36 fatty acids and retinal physiology. Br. J. Ophthalmol. 2010, 94, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Quazi, F.; Molday, R.S. Differential Phospholipid Substrates and Directional Transport by ATP-binding Cassette Proteins ABCA1, ABCA7, and ABCA4 and Disease-causing Mutants. J. Biol. Chem. 2013, 288, 34414–34426. [Google Scholar] [CrossRef] [PubMed]
- Osada, H.; Toda, E.; Homma, K.; Guzman, N.A.; Nagai, N.; Ogawa, M.; Negishi, K.; Arita, M.; Tsubota, K.; Ozawa, Y. ADIPOR1 deficiency-induced suppression of retinal ELOVL2 and docosahexaenoic acid levels during photoreceptor degeneration and visual loss. Cell Death Dis. 2021, 12, 458. [Google Scholar] [CrossRef]
- Dasyani, M.; Gao, F.; Xu, Q.; Van Fossan, D.; Zhang, E.; Pinto, A.F.M.; Saghatelian, A.; Skowronska-Krawczyk, D.; Chao, D.L. Elovl2 Is Required for Robust Visual Function in Zebrafish. Cells 2020, 9, 2583. [Google Scholar] [CrossRef]
- Bazan, N.G. Overview of how N32 and N34 elovanoids sustain sight by protecting retinal pigment epithelial cells and photoreceptors. J. Lipid Res. 2021, 62, 100058. [Google Scholar] [CrossRef]
- Bazan, N.G.; Molina, M.F.; Gordon, W.C. Docosahexaenoic Acid Signalolipidomics in Nutrition: Significance in Aging, Neuroinflammation, Macular Degeneration, Alzheimer’s, and Other Neurodegenerative Diseases. Annu. Rev. Nutr. 2011, 31, 321–351. [Google Scholar] [CrossRef]
- Magny, R.; Regazzetti, A.; Kessal, K.; Christin, O.; Baudouin, C.; Roulland, E.; Brignole-Baudouin, F.; Laprévote, O.; Auzeil, N. Identification of new Omega-3 very long chain poly-unsaturated fatty acids in meibomian gland secretions. Biochimie 2022, 203, 3–10. [Google Scholar] [CrossRef]
- Gorusupudi, A.; Rallabandi, R.; Li, B.; Arunkumar, R.; Blount, J.D.; Rognon, G.T.; Chang, F.-Y.; Wade, A.; Lucas, S.; Conboy, J.C.; et al. Retinal bioavailability and functional effects of a synthetic very-long-chain polyunsaturated fatty acid in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2017739118. [Google Scholar] [CrossRef]
- Kady, N.M.; Liu, X.; Lydic, T.A.; Syed, M.H.; Navitskaya, S.; Wang, Q.; Hammer, S.S.; O’reilly, S.; Huang, C.; Seregin, S.S.; et al. ELOVL4-Mediated Production of Very Long-Chain Ceramides Stabilizes Tight Junctions and Prevents Diabetes-Induced Retinal Vascular Permeability. Diabetes 2018, 67, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Rugolo, F.; Bazan, N.G.; Calandria, J.; Jun, B.; Raschellà, G.; Melino, G.; Agostini, M. The expression of ELOVL4, repressed by MYCN, defines neuroblastoma patients with good outcome. Oncogene 2021, 40, 5741–5751. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Bell, B.A.; Song, Y.; Zhang, K.; Anderson, B.; Axelsen, P.H.; Bohannan, W.; Agbaga, M.; Park, H.G.; James, G.; et al. Deuterated docosahexaenoic acid protects against oxidative stress and geographic atrophy-like retinal degeneration in a mouse model with iron overload. Aging Cell 2022, 21, e13579. [Google Scholar] [CrossRef] [PubMed]
- James, G.; Bohannan, W.; Adewunmi, E.; Schmidt, K.; Park, H.G.; Shchepinov, M.S.; Agbaga, M.-P.; Brenna, J.T. Pharmacokinetics and metabolism in mouse retina of bis-allylic deuterated docosahexaenoic acid (D-DHA), a new dry AMD drug candidate. Exp. Eye Res. 2022, 222, 109193. [Google Scholar] [CrossRef]
- Mazzoli, R.; Pessione, E. The Neuro-endocrinological Role of Microbial Glutamate and GABA Signaling. Front. Microbiol. 2016, 7, 1934. [Google Scholar] [CrossRef]
- Orešič, M.; Seppänen-Laakso, T.; Yetukuri, L.; Bäckhed, F.; Hänninen, V. Gut microbiota affects lens and retinal lipid composition. Exp. Eye Res. 2009, 89, 604–607. [Google Scholar] [CrossRef]
- Albouery, M.; Buteau, B.; Grégoire, S.; Cherbuy, C.; De Barros, J.-P.P.; Martine, L.; Chain, F.; Cabaret, S.; Berdeaux, O.; Bron, A.M.; et al. Age-Related Changes in the Gut Microbiota Modify Brain Lipid Composition. Front. Cell. Infect. Microbiol. 2020, 9, 444. [Google Scholar] [CrossRef]
- Richardson, R.; Tracey-White, D.; Webster, A.; Moosajee, M. The zebrafish eye—A paradigm for investigating human ocular genetics. Eye 2017, 31, 68–86. [Google Scholar] [CrossRef]
- Hamano, F.; Kuribayashi, H.; Iwagawa, T.; Tsuhako, A.; Nagata, K.; Sagara, H.; Shimizu, T.; Shindou, H.; Watanabe, S. Correction: Mapping membrane lipids in the developing and adult mouse retina under physiological and pathological conditions using mass spectrometry. J. Biol. Chem. 2022, 298, 100303. [Google Scholar] [CrossRef]
- Yu, M.; Benham, A.; Logan, S.; Brush, R.S.; Mandal, N.A.; Anderson, R.E.; Agbaga, M.-P. ELOVL4 protein preferentially elongates 20:5n3 to very long chain PUFAs over 20:4n6 and 22:6n3. J. Lipid Res. 2012, 53, 494–504. [Google Scholar] [CrossRef]
- Suh, M.; Clandinin, M.T. 20:5n-3 but not 22:6n-3 is a Preferred Substrate for Synthesis of n-3 Very-Long- Chain Fatty Acids (C24–C36) in Retina. Curr. Eye Res. 2005, 30, 959–968. [Google Scholar] [CrossRef] [PubMed]
- Yeboah, G.K.; Lobanova, E.S.; Brush, R.S.; Agbaga, M.-P. Very long chain fatty acid-containing lipids: A decade of novel insights from the study of ELOVL4. J. Lipid Res. 2021, 62, 100030. [Google Scholar] [CrossRef] [PubMed]
- Karan, G.; Lillo, C.; Yang, Z.; Cameron, D.J.; Locke, K.G.; Zhao, Y.; Thirumalaichary, S.; Li, C.; Birch, D.G.; Vollmer-Snarr, H.R.; et al. Lipofuscin accumulation, abnormal electrophysiology, and photoreceptor degeneration in mutant ELOVL4 transgenic mice: A model for macular degeneration. Proc. Natl. Acad. Sci. USA 2005, 102, 4164–4169. [Google Scholar] [CrossRef] [PubMed]
- Vasireddy, V.; Jablonski, M.M.; Mandal, N.A.; Raz-Prag, D.; Wang, X.F.; Nizol, L.; Iannaccone, A.; Musch, D.C.; Bush, R.A.; Salem, N., Jr.; et al. Elovl4 5-bp–Deletion Knock-in Mice Develop Progressive Photoreceptor Degeneration. Investig. Opthalmol. Vis. Sci. 2006, 47, 4558–4568. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.; Cameron, D.J.; Wang, C.; Karan, G.; Yang, Z.; Zhao, Y.; Pearson, E.; Chen, H.; Deng, C.; et al. Elovl4 haploinsufficiency does not induce early onset retinal degeneration in mice. Vis. Res. 2007, 47, 714–722. [Google Scholar] [CrossRef]
- Sommer, J.R.; Estrada, J.L.; Collins, E.B.; Bedell, M.; Alexander, C.A.; Yang, Z.; Hughes, G.; Mir, B.; Gilger, B.C.; Grob, S.; et al. Production of ELOVL4 transgenic pigs: A large animal model for Stargardt-like macular degeneration. Br. J. Ophthalmol. 2011, 95, 1749–1754. [Google Scholar] [CrossRef]
- Barabas, P.; Liu, A.; Xing, W.; Chen, C.-K.; Tong, Z.; Watt, C.B.; Jones, B.W.; Bernstein, P.S.; Križaj, D. Role of ELOVL4 and very long-chain polyunsaturated fatty acids in mouse models of Stargardt type 3 retinal degeneration. Proc. Natl. Acad. Sci. USA 2013, 110, 5181–5186. [Google Scholar] [CrossRef]
- Zampaglione, E.; Kinde, B.; Place, E.M.; Navarro-Gomez, D.; Maher, M.; Jamshidi, F.; Nassiri, S.; Mazzone, J.A.; Finn, C.; Schlegel, D.; et al. Copy-number variation contributes 9% of pathogenicity in the inherited retinal degenerations. Anesthesia Analg. 2020, 22, 1079–1087. [Google Scholar] [CrossRef]
- McMahon, A.; Jackson, S.N.; Woods, A.S.; Kedzierski, W. A Stargardt disease-3 mutation in the mouse Elovl4 gene causes retinal deficiency of C32-C36 acyl phosphatidylcholines. FEBS Lett. 2007, 581, 5459–5463. [Google Scholar] [CrossRef]
- Berdeaux, O.; Acar, N. Very-long-chain polyunsaturated fatty acids in the retina: Analysis and clinical relevance in physiological and pathological conditions. Oléagineux Corps Gras Lipides 2011, 18, 284–290. [Google Scholar] [CrossRef]
- Vidal, E.; Jun, B.; Gordon, W.C.; Maire, M.-A.; Martine, L.; Grégoire, S.; Khoury, S.; Cabaret, S.; Berdeaux, O.; Acar, N.; et al. Bioavailability and spatial distribution of fatty acids in the rat retina after dietary omega-3 supplementation. J. Lipid Res. 2020, 61, 1733–1746. [Google Scholar] [CrossRef] [PubMed]
- Suh, M.; Wierzbicki, A.A.; Clandinin, M. Dietary fat alters membrane composition in rod outer segments in normal and diabetic rats: Impact on content of very-long-chain (C ≥ 24) polyenoic fatty acids. Biochim. Biophys. Acta (BBA)—Lipids Lipid Metab. 1994, 1214, 54–62. [Google Scholar] [CrossRef]
- Gorusupudi, A.; Liu, A.; Hageman, G.S.; Bernstein, P.S. Associations of human retinal very long-chain polyunsaturated fatty acids with dietary lipid biomarkers. J. Lipid Res. 2016, 57, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Terry, R.; Lin, Y.; Nelson, K.; Bernstein, P.S. Comprehensive and sensitive quantification of long-chain and very long-chain polyunsaturated fatty acids in small samples of human and mouse retina. J. Chromatogr. A 2013, 1307, 191–200. [Google Scholar] [CrossRef]
- Serrano, R.; Navarro, J.C.; Sales, C.; Portolés, T.; Monroig; Beltran, J.; Hernández, F. Determination of very long-chain polyunsaturated fatty acids from 24 to 44 carbons in eye, brain and gonads of wild and cultured gilthead sea bream (Sparus aurata). Sci. Rep. 2022, 12, 10112. [Google Scholar] [CrossRef]
- Harkewicz, R.; Du, H.; Tong, Z.; Alkuraya, H.; Bedell, M.; Sun, W.; Wang, X.; Hsu, Y.-H.; Esteve-Rudd, J.; Hughes, G.; et al. Essential Role of ELOVL4 Protein in Very Long Chain Fatty Acid Synthesis and Retinal Function. J. Biol. Chem. 2012, 287, 11469–11480. [Google Scholar] [CrossRef]
- Jun, B.; Mukherjee, P.K.; Asatryan, A.; Kautzmann, M.-A.; Heap, J.; Gordon, W.C.; Bhattacharjee, S.; Yang, R.; Petasis, N.A.; Bazan, N.G. Elovanoids are novel cell-specific lipid mediators necessary for neuroprotective signaling for photoreceptor cell integrity. Sci. Rep. 2017, 7, 5279. [Google Scholar] [CrossRef]
- Busik, J.V.; Decot, H.; Lin, A.B.; Lydic, T.A. Untargeted Analysis of Lipids Containing Very Long Chain Fatty Acids in Retina and Retinal Tight Junctions. In Lipidomics: Methods and Protocols; Bhattacharya, S.K., Ed.; Springer: New York, NY, USA, 2023; pp. 269–290. [Google Scholar]
- Landowski, M.; Rickman, C.B. Targeting Lipid Metabolism for the Treatment of Age-Related Macular Degeneration: Insights from Preclinical Mouse Models. J. Ocul. Pharmacol. Ther. 2022, 38, 3–32. [Google Scholar] [CrossRef]
- Lewandowski, D.; Sander, C.L.; Tworak, A.; Gao, F.; Xu, Q.; Skowronska-Krawczyk, D. Dynamic lipid turnover in photoreceptors and retinal pigment epithelium throughout life. Prog. Retin. Eye Res. 2022, 89, 101037. [Google Scholar] [CrossRef]
- Phillips, G.R.; Hancock, S.E.; Brown, S.H.J.; Jenner, A.M.; Kreilaus, F.; Newell, K.A.; Mitchell, T.W. Cholesteryl ester levels are elevated in the caudate and putamen of Huntington’s disease patients. Sci. Rep. 2020, 10, 20314. [Google Scholar] [CrossRef]
- Pikuleva, I.A.; Curcio, C.A. Cholesterol in the retina: The best is yet to come. Prog. Retin. Eye Res. 2014, 41, 64–89. [Google Scholar] [CrossRef]
- Fliesler, S.J.; Bretillon, L. The ins and outs of cholesterol in the vertebrate retina. J. Lipid Res. 2010, 51, 3399–3413. [Google Scholar] [CrossRef] [PubMed]
- Puskas, L.G.; Bereczki, E.; Santha, M.; Vigh, L.; Csanadi, G.; Spener, F.; Ferdinandy, P.; Onody, A.; Kitajka, K. Erratum to: Cholesterol and cholesterol plus DHA diet-induced gene expression and fatty acid changes in mouse eye and brain. Biochimie 2005, 87, 239. [Google Scholar] [CrossRef]
- Furland, N.E.; Maldonado, E.N.; Aveldaño, M.I. Very long chain PUFA in murine testicular triglycerides and cholesterol esters. Lipids 2003, 38, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Makrecka-Kuka, M.; Sevostjanovs, E.; Vilks, K.; Volska, K.; Antone, U.; Kuka, J.; Makarova, E.; Pugovics, O.; Dambrova, M.; Liepinsh, E. Plasma acylcarnitine concentrations reflect the acylcarnitine profile in cardiac tissues. Sci. Rep. 2017, 7, 17528. [Google Scholar] [CrossRef]
- Zhao, S.; Feng, X.-F.; Huang, T.; Luo, H.-H.; Chen, J.-X.; Zeng, J.; Gu, M.; Li, J.; Sun, X.-Y.; Sun, D.; et al. The Association Between Acylcarnitine Metabolites and Cardiovascular Disease in Chinese Patients With Type 2 Diabetes Mellitus. Front. Endocrinol. 2020, 11, 212. [Google Scholar] [CrossRef]
- Ikeda, T.; Obayashi, H.; Hasegawa, G.; Nakamura, N.; Yoshikawa, T.; Imamura, Y.; Koizumi, K.; Kinoshita, S. Paraoxonase gene polymorphisms and plasma oxidized low-density lipoprotein level as possible risk factors for exudative age-related macular degeneration. Am. J. Ophthalmol. 2001, 132, 191–195. [Google Scholar] [CrossRef]
- Li, S.; Gao, D.; Jiang, Y. Function, Detection and Alteration of Acylcarnitine Metabolism in Hepatocellular Carcinoma. Metabolites 2019, 9, 36. [Google Scholar] [CrossRef]
- Dambrova, M.; Makrecka-Kuka, M.; Kuka, J.; Vilskersts, R.; Nordberg, D.; Attwood, M.M.; Smesny, S.; Sen, Z.D.; Guo, A.C.; Oler, E.; et al. Acylcarnitines: Nomenclature, Biomarkers, Therapeutic Potential, Drug Targets, and Clinical Trials. Pharmacol. Rev. 2022, 74, 506–551. [Google Scholar] [CrossRef]
- Liew, G.; Tse, B.; Ho, I.-V.; Joachim, N.; White, A.; Pickford, R.; Maltby, D.; Gopinath, B.; Mitchell, P.; Crossett, B. Acylcarnitine Abnormalities Implicate Mitochondrial Dysfunction in Patients With Neovascular Age-Related Macular Degeneration. Investig. Opthalmol. Vis. Sci. 2020, 61, 32. [Google Scholar] [CrossRef]
- Theodoridis, K.; Gika, H.; Kotali, A. Acylcarnitines in Ophthalmology: Promising Emerging Biomarkers. Int. J. Mol. Sci. 2022, 23, 16183. [Google Scholar] [CrossRef] [PubMed]
- Ferdinandusse, S.; Denis, S.; van Roermund, C.W.; Wanders, R.J.; Dacremont, G. Identification of the peroxisomal β-oxidation enzymes involved in the degradation of long-chain dicarboxylic acids. J. Lipid Res. 2004, 45, 1104–1111. [Google Scholar] [CrossRef]
- Jarrell, Z.R.; Smith, M.R.; Hu, X.; Orr, M.; Liu, K.H.; Quyyumi, A.A.; Jones, D.P.; Go, Y.-M. Plasma acylcarnitine levels increase with healthy aging. Aging 2020, 12, 13555–13570. [Google Scholar] [CrossRef]
- Tanno, H.; Sassa, T.; Sawai, M.; Kihara, A. Production of branched-chain very-long-chain fatty acids by fatty acid elongases and their tissue distribution in mammals. Biochim. Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2021, 1866, 158842. [Google Scholar] [CrossRef] [PubMed]
- Chew, E.Y.; Clemons, T.E.; SanGiovanni, J.P.; Danis, R.; Ferris, F.L.; Elman, M.; Antoszyk, A.; Ruby, A.; Orth, D.; Bressler, S.; et al. Lutein plus Zeaxanthin and Omega-3 Fatty Acids for Age-Related Macular Degeneration The Age-Related Eye Disease Study 2 (AREDS2) Randomized Clinical Trial. J. Am. Med. Assoc. 2013, 309, 2005–2015. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nwagbo, U.; Bernstein, P.S. Understanding the Roles of Very-Long-Chain Polyunsaturated Fatty Acids (VLC-PUFAs) in Eye Health. Nutrients 2023, 15, 3096. https://doi.org/10.3390/nu15143096
Nwagbo U, Bernstein PS. Understanding the Roles of Very-Long-Chain Polyunsaturated Fatty Acids (VLC-PUFAs) in Eye Health. Nutrients. 2023; 15(14):3096. https://doi.org/10.3390/nu15143096
Chicago/Turabian StyleNwagbo, Uzoamaka, and Paul S. Bernstein. 2023. "Understanding the Roles of Very-Long-Chain Polyunsaturated Fatty Acids (VLC-PUFAs) in Eye Health" Nutrients 15, no. 14: 3096. https://doi.org/10.3390/nu15143096
APA StyleNwagbo, U., & Bernstein, P. S. (2023). Understanding the Roles of Very-Long-Chain Polyunsaturated Fatty Acids (VLC-PUFAs) in Eye Health. Nutrients, 15(14), 3096. https://doi.org/10.3390/nu15143096