1. Introduction
Numerous diet-related diseases stem from abnormal metabolic consequences. Examples include high glucose, non-esterified fatty acids, and acylethanolamide concentrations in blood of diabetes patients [
1]. Elevated amino acid levels are also associated with diabetes [
2]. Moreover, a recent metabolomic study showed altered levels of amino acids and increased oxylipin (OxL) inflammatory mediators in renal disease [
3]. Thus, identifying changes in metabolite levels as a means of associating metabolites to a particular disease underscores the value of metabolomics analyses. Metabolomics is a science of systems biology that can broadly profile endogenous metabolites within a biological system, thus providing a snapshot of physiologic state or cellular status [
4,
5]. As such, metabolomics is well suited as a discovery tool for biomarker metabolites to accurately characterize a disease state to individual variability such as age, sex, genetics, diet, environmental conditions, and time of day [
6,
7].
Health status often reflects dietary habits, and diet modification is an attractive approach to prevent or control disease [
8]. Several studies have demonstrated the beneficial effects of
n-3 polyunsaturated fatty acids (PUFA) consumption in the prevention or treatment of diseases [
9]. The endocannabinoid system (ECS) and its ligand endocannabinoids (eCB) are recognized as a homeostatic modulator of energy status specifically with respect to muscle [
10] and systemic inflammation [
11,
12]. The ECS actions on maintaining homeostatic balance are linked to regulating food intake and directing energy metabolism at both the central and peripheral levels [
13].
A review summarized findings on the effects of
n-3 PUFA and glucose and fat metabolism in mice and cell cultures [
14]. Some findings for glucose metabolism and physiology using molecular techniques in muscle and adipose tissues in the review are consistent with findings reported by Kim et. al. [
10]. However, the review, which cites some studies of the authors, lacks a rigorous approach for experimental diet formulations and methods as those reported by Kim et al. in mice [
10] and myoblasts [
15].
An overactive ECS is believed to be one of the underlying causes of obesity, hyperglycemia, dyslipidemia, type 2 diabetes, and insulin resistance [
5]. Accordingly, identifying ways to manage and normalize an overactive ECS, and the degree of stimulation or responsiveness of the system is dependent on the concentration of its ligands to activate its receptors involved in signaling [
11]. Since the type of PUFA in the phospholipids of cellular membranes can change based on the type of dietary fat consumed, the downstream metabolic effects from alternations in the PUFA composition of membranes have a direct impact on the ECS [
16,
17]. Arachidonic acid (AA) is the precursor fatty acid for the biosynthesis of the two most studied endogenous ligand eCB, N-arachidonoyl ethanolamine (anandamide; AEA) and 2-arachidonoyl glycerol (2-AG), that activate cannabinoid receptors of the ECS [
11,
18].
In myoblast cultures, glucose use was observed to improve with docosahexaenoic acid (DHA) and the DHA-derived eCB, docosahexaenoyl ethanolamide (DHEA) treatments [
15]. Moreover, when fed a DHA-rich diet, mice showed increased muscle mRNA and proteins involved in glucose uptake [
10]. Thus, the focus of the current investigation was to distinguish shifts in metabolite changes in mice given a DHA-rich diet compared to the control diet. The overall research hypothesis is that DHA in a semi-purified diet when fed to C57/blk6 mice will restore endocannabinoid tone (action of ligands) and signaling of this system to improve macronutrient metabolism associated with reduced risk to obesity and diabetes.
The aim of this study was to investigate the effects of DHA intake on macronutrient metabolism in C57/blk6 mice from weaning to 112 d. Herein, plasma, skeletal muscle, and liver from C57/blk6 mice given a DHA semi-purified diet were analyzed for changes in metabolites by metabolomics analysis. Identifying metabolites provides insight to better understand the physiology within a biological system and a metabolic glimpse into the overall biological status. Thus, our hypothesis is that replacing tissue AA with DHA will change lipid and glucose metabolism and adipose accretion in mice. Our experimental design and hypothesis are a logical step to characterize the metabolite levels of macronutrients based on previous findings for DHA effects on glucose and fatty acid metabolism.
4. Discussion
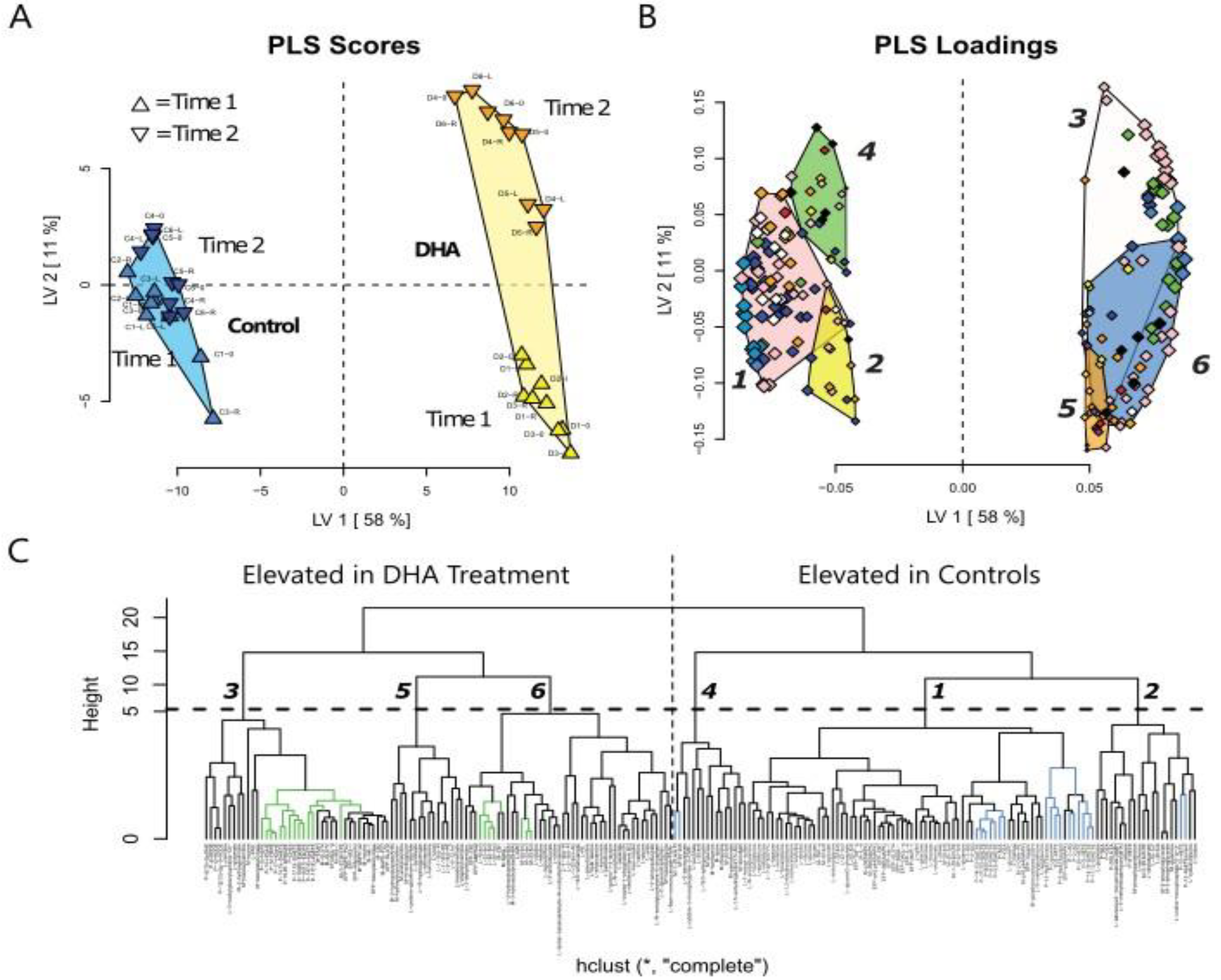
At the end of the study period, mice given the DHA semi-purified diet had less body fat, higher lean mass, and a lower body weight compared to the control diet group. However, one possible reason for no difference in body fat early on could be explained by the physiological state of growth versus maintenance at the second time point in control mice. Over the food intake collection period, no difference was found in food consumed between the DHA and control diet groups. Thus, DHA resulted in significant changes in macronutrient and systemic energy metabolism in mice leading to lower fat mass and higher lean mass at 112 d.
Not surprisingly, feeding mice a DHA semi-purified diet led to lower levels of
n-6 PUFA and related metabolites but higher levels of
n-3 PUFA and DHA-derived products. AA and arachidic acid were lower in plasma of mice given the DHA diet while DHA,
n-3 DPA, and EPA levels were higher in both liver and muscle tissue. Therefore, dietary DHA was able to modify the fatty acid composition of these tissues and various PUFA containing lipids. Our findings corroborate our previous results for mouse muscle and liver tissue fatty acid composition and plasma DHEA [
10], and blood DHA and DHEA in postmenopausal women [
12]. Lastly, the level of alpha-tocopherol was lower in the DHA group which may indicate that the DHA-fed mice remove more alpha-tocopherol from blood to support greater tissue accretion of
n-3 PUFA.
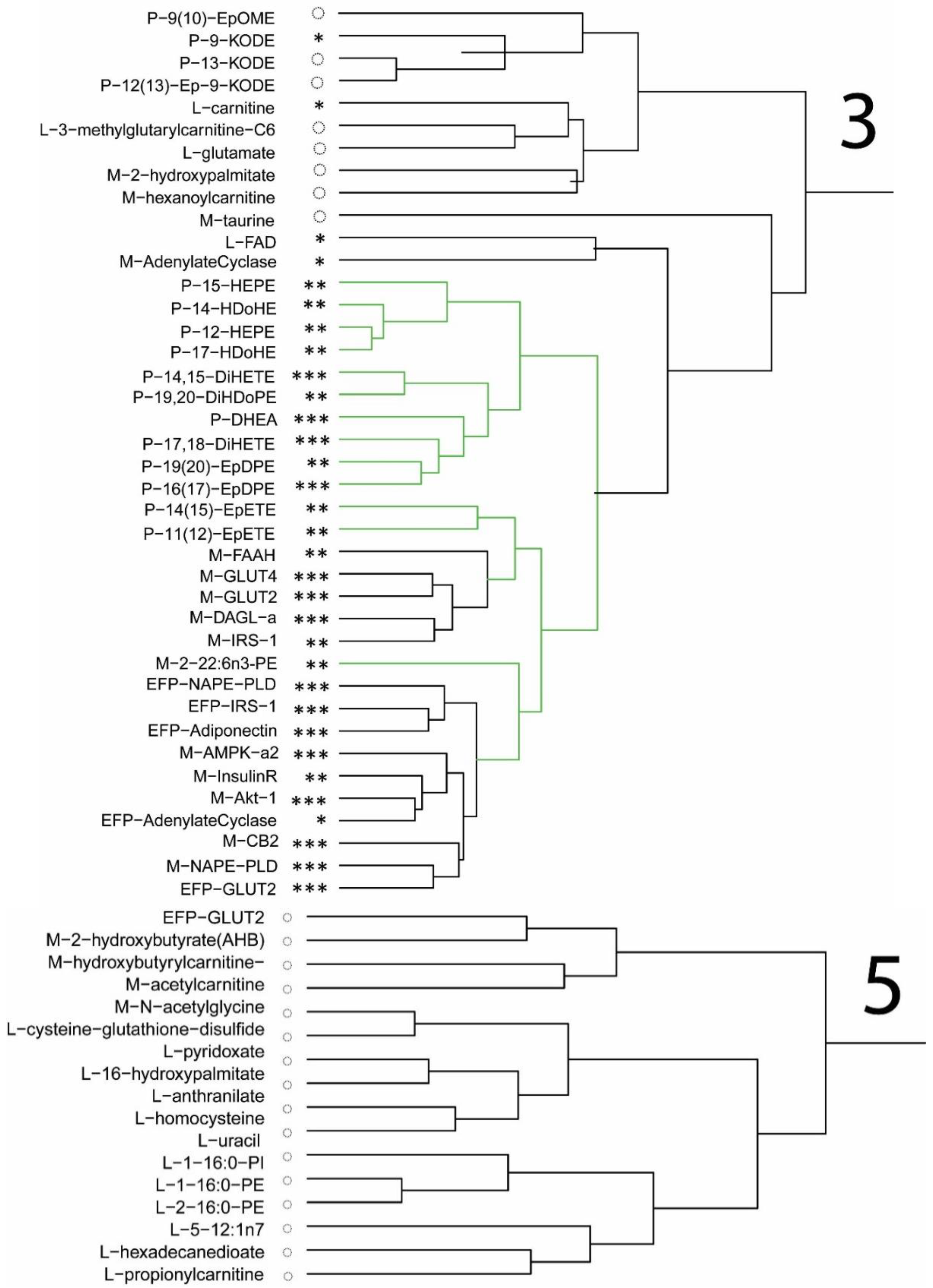
Relevant to the ECS is the lowering of the AA-derived endocannabinoids, 2-AG, and 1-AG in gastrocnemius of mice fed the DHA diet in this study. The reduction in the level of these AA-derived eCB confirms the finding that
n-3 PUFA (DHA) in myoblasts alters eCB levels [
10,
15]. A reduction in muscle AA-derived eCB supports the hypothesis that a DHA diet has the potential to improve ECS tone and ultimately restore control to a dysregulated modulator of systemic energy metabolism [
10,
11,
12].
The observed changes in tissue fatty acids to DHA or to fatty acids of the DHA lineage after consuming a diet enriched in DHA highlights the potential to affect the lipid-derived ECS ligands. This is evident in the higher levels of DHA-derived glycerol lipids, 2-DHA-GPE and 1-docosahexaenoylglycerol, in muscle and liver of mice after 56 d and 112 d of consuming the DHA diet. Levels of eCB derived from AA were lower in mice, and feeding DHA to mice lowered AA and increased DHA in tissues and increased DHEA in blood [
10]. Further, the treatment of DHEA in myoblast cultures was shown to increase mRNA levels of p38MAPK [
15].
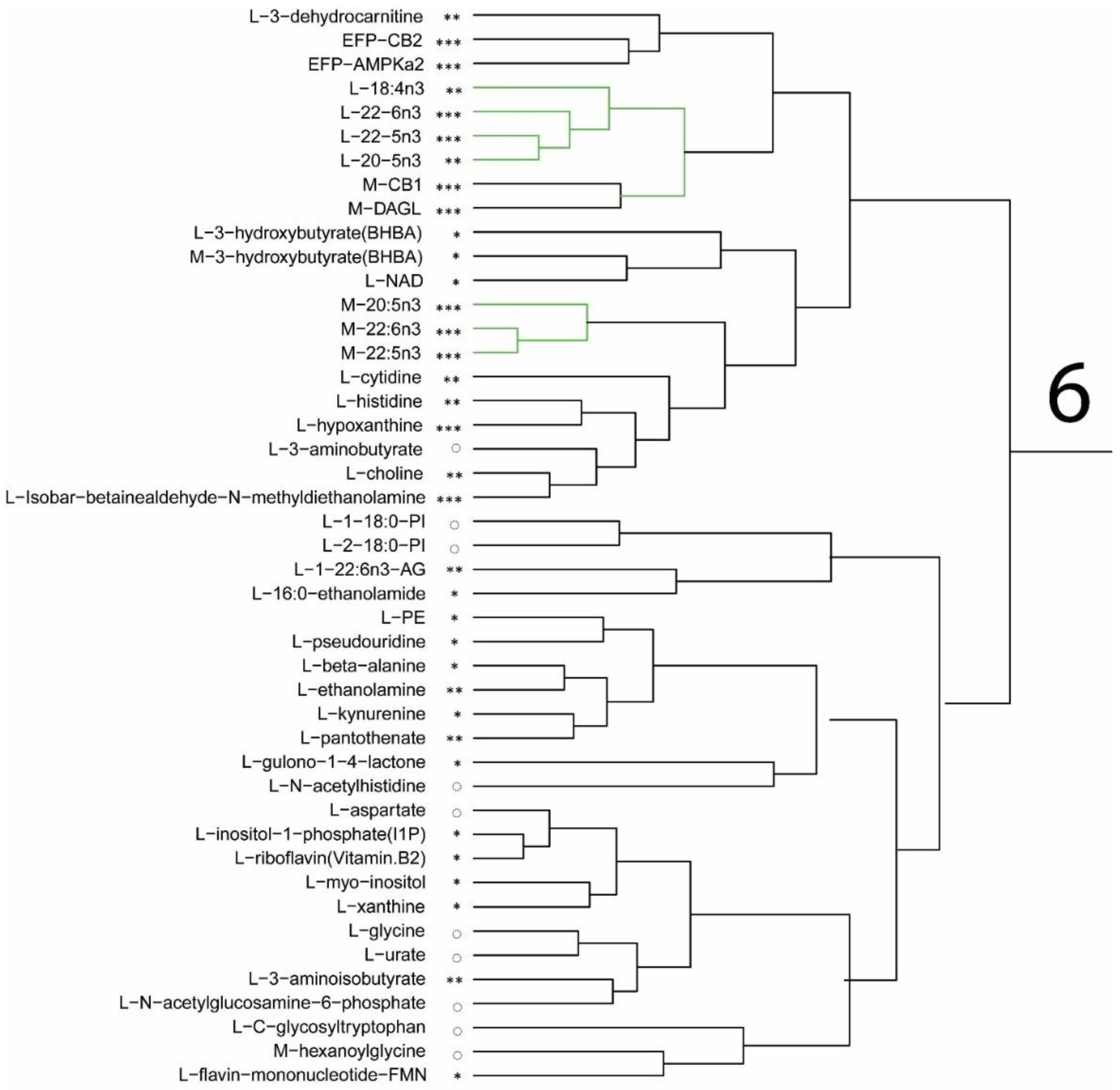
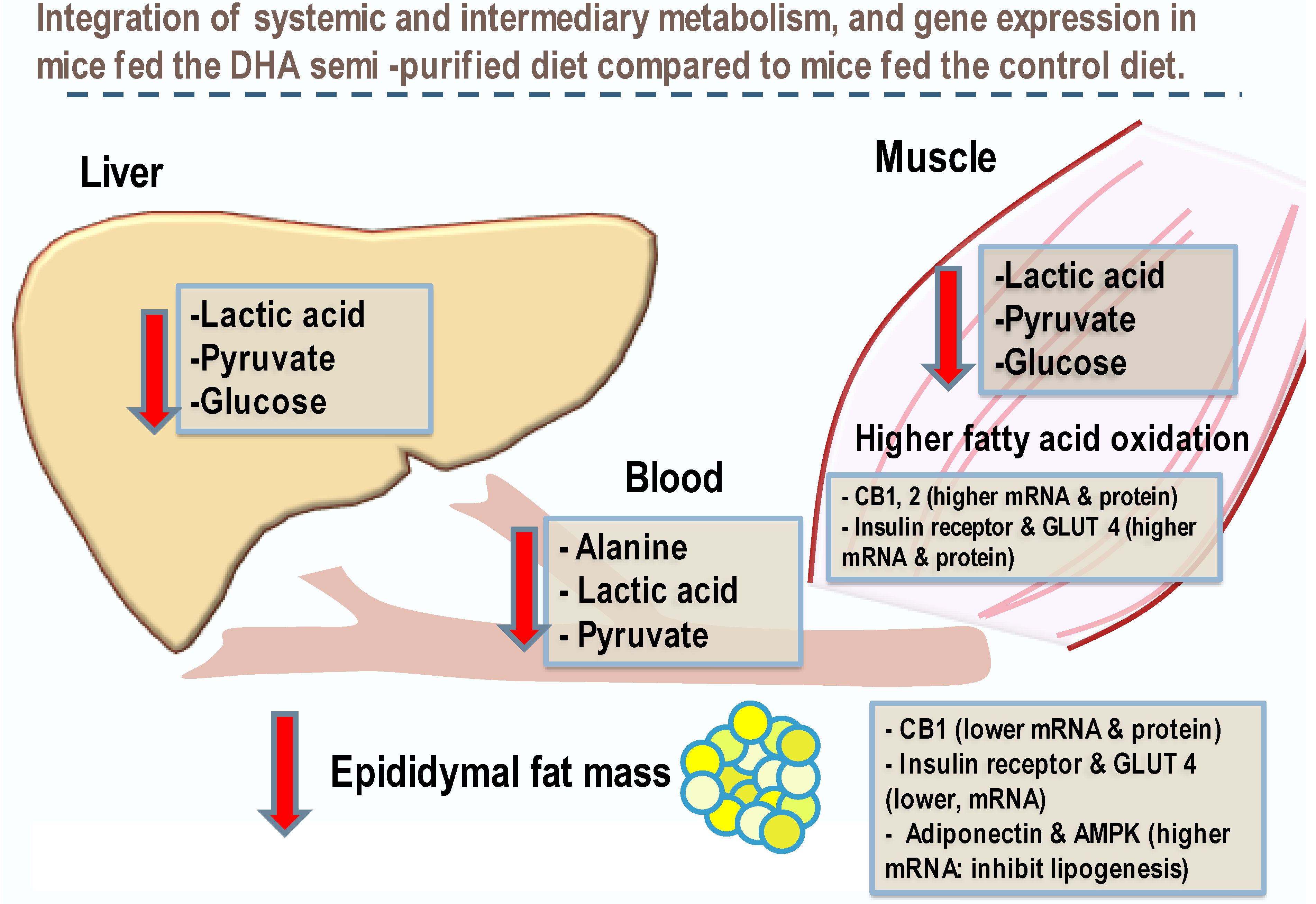
Free glucose levels in both liver and muscle were lower in the DHA diet fed mice compared to the control group. Concurrently, metabolites associated with glycolysis, such as pyruvate, 3-phosphoglycerate, and phosphoenolpyruvate, were lower in the DHA-fed mice, suggesting that the mice in the DHA group used glucose less as a fuel source, instead favoring fatty acids. The metabolite levels in the DHA group suggest reduced dependency on glucose catabolism and a shift in fuel sources to fatty acid oxidation. Lower levels of ribulose and ribose-5-phosphate in the liver, and lower flux via the pentose phosphate pathway, brings together the metabolic changes to support the observation of lower epididymal fat pad mass in the mice given the DHA diet compared to the control. Furthermore, the metabolic shift to using fat stores also corroborates the decrease in fat mass, as determined by DXA analysis [
10].
In a recent review, evidence suggests that DHA likely improves glucose use by the brain [
27]. The authors reported that DHA deficiency repressed GLUT-1 expression in association with low DHA levels in rat brain [
27]. These findings have implications for DHA actions in the nervous system and for general neurobiological health.
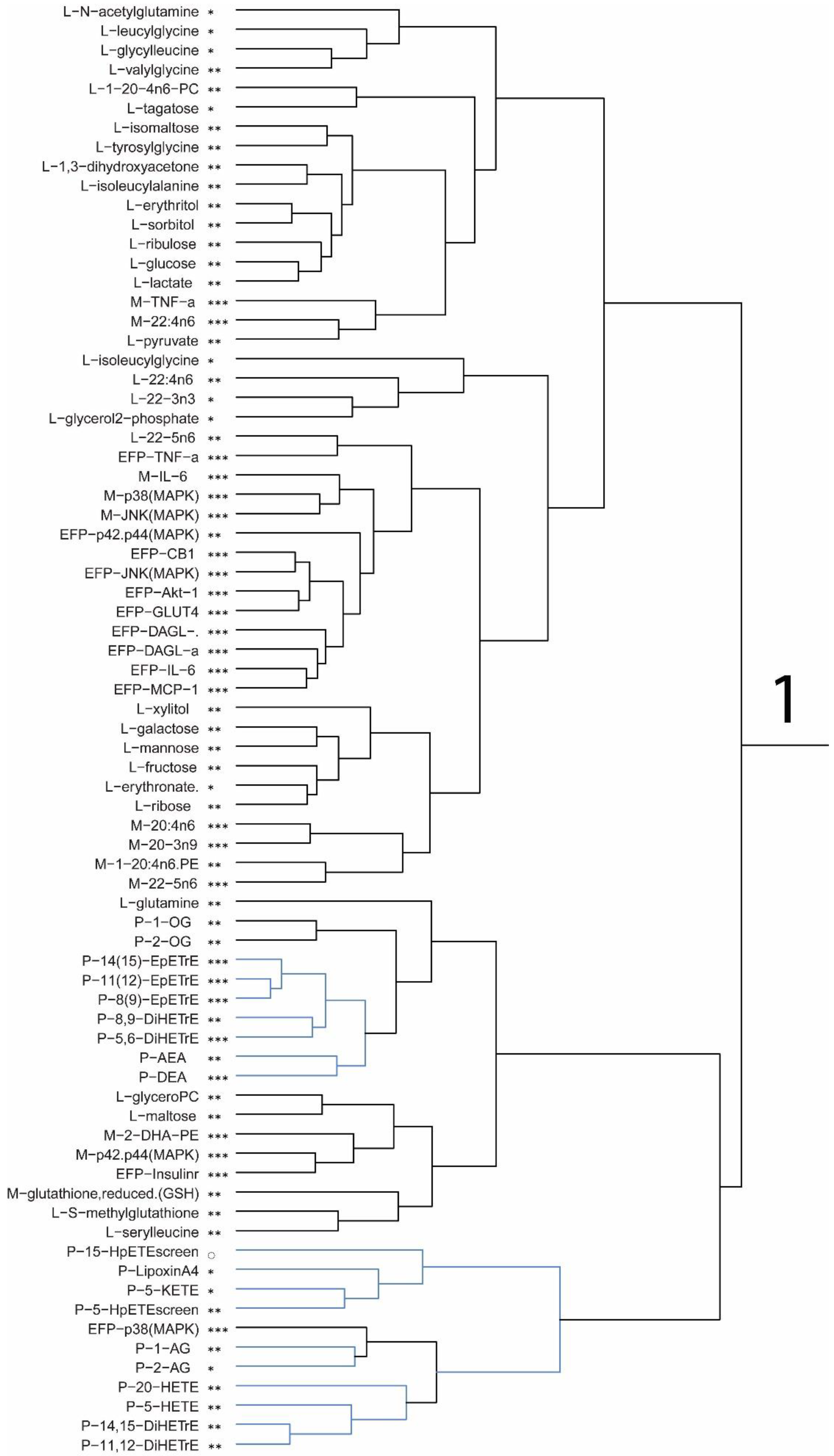
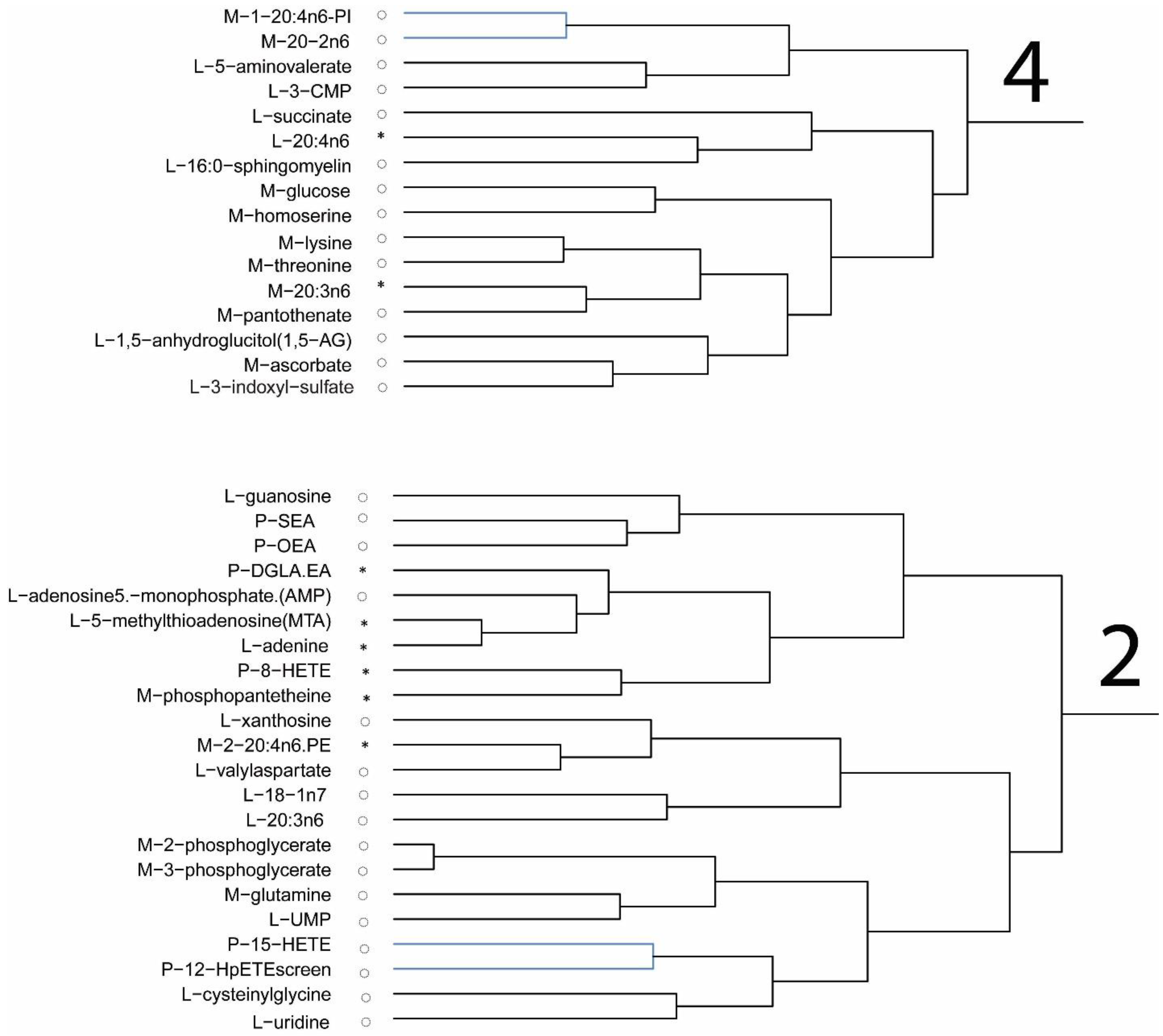
Plasma levels of macronutrients strongly suggest changes in glucose and fatty acid metabolism in mice fed the DHA diet. The differences in metabolic pathway flux would support less fat accretion, improved glucose handling, and higher lean mass in mice fed the DHA diet compared to controls. Metabolites of fatty acid catabolism and glucose use associated with the Krebs cycle and glycolytic pathways, respectively, were altered in a manner consistent with the differences in body mass and composition of mice. In support of the pathway changes and fat pad mass, a reduced flux through the glycolytic pathway would result in lower αGP as found, thus resulting in lower triglyceride synthesis to explain less fat accretion observed in mice fed the DHA diet. Higher levels of related metabolites suggest greater fatty acid oxidation. A higher level of fatty acid oxidation was observed in mouse muscle as acyl-carnitines (e.g., palmitoylcarnitine, oleoylcarnitine) and acyl-glycines (e.g., hexanoylglycine and 3-hydroxybutyrate) were higher in the gastrocnemius muscle of DHA-fed mice. Thus, higher levels of activation of fatty acids suggests greater muscle oxidation of fatty acids in mice fed the DHA diet. Glucuronic acid, ethanolamine, 3-hydroxy-3-methylglutaric acid, 2-hydroxybutanoic acid, and 3-hydroxybutanoic acid were higher in the DHA group. These metabolite changes suggest greater energy expenditure or higher flux in pathways of carbohydrate use and fatty acid oxidation. Further, higher adenosine found in the DHA group of mice may support a benefit to insulin sensitivity.
In this study, mice were given one of two high-fat diets, varying in the amount of DHA but containing the same level of protein and fat. Moreover, we report lower levels of 1-arachidonylglycerol (1-AG) and 2-arachidonylglycerol (2-AG) in muscle when the DHA diet was fed, which is consistent with lower plasma 1- and 2-AG in mice fed the same diet [
10].
Together these findings strengthen the relationship between the endocrine system and the ECS in characterizing obesity as a condition where the ECS is overactive and muscle glucose uptake is reduced, which in turn can lead to insulin resistance and the metabolic syndrome. These observations support our current findings of lower glucose, fat mass, and higher lean mass in mice fed the DHA diet. Further, our study is robust with dietary manipulation to alter the concentration of AA-derived and DHA-derived eCB [
10].
A recent investigation found that DHA elevated glucose and palmitic acid oxidation in L6 rat skeletal muscle [
28]. The authors suggest that DHA may increase glucose disposal and reduce lipid accumulation. These authors [
28] also reported higher AMPK phosphorylation and protein levels of carnitine-palmitoyl transferase-1b (CPT1b) with DHA treatment. The findings to a great extent are consistent with Kim et al. [
10,
15] for glucose use and metabolism. With respect to epididymal fat pad, a higher adiponectin gene expression was reported in similar mice fed the same DHA semi-purified diet compared to those fed the control diet [
10]. Adiponectin, secreted by adipocytes, increases insulin sensitivity and is associated with weight loss [
29].
Among the nine amino acids that were detected, only phenylalanine and glycine were found to be higher in mice fed the DHA diet. The finding suggests that dietary DHA may act to preserve or protect specific amino acids from being catabolized in tissue while liberating others into circulation for clearance. In this regard, the plasma levels point to an overall increased utilization of amino acids in the DHA group. Creatinine was found higher in the DHA group, which may indicate a difference in energy metabolism in these mice, thus supporting energy utilization in muscle consistent with the higher lean mass in these mice. Recently, investigators have noted significant differences in amino acid levels associated with elevated glucose and decreased insulin in men [
2].
To counter the overactivation of the ECS, dietary DHA appears to improve ECS tone and support better clearance of glucose from blood by increasing glucose transporters to the sarcolemma of the muscle [
10]. Incorporating DHA into the diet of an obese individual may have the potential of restoring balance and reverse the dysregulation of the ECS, resulting in an increase of glucose uptake into skeletal muscle and a reduction in insulin resistant-associated hyperglycemia. Our findings on systemic energy metabolism including glucose are summarized in
Figure 3.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}