
Influence of Gender on Plasma Leptin Levels, Fat Oxidation, and Insulin Sensitivity in Young Adults: The Mediating Role of Fitness and Fatness

 , ,
, ,  , , ,
, , ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Design
2.2. Subjects
2.3. Procedure
2.3.1. Blood Extraction and Biochemical Parameters
2.3.2. Anthropometry and Body Composition
2.3.3. Resting Metabolism
2.3.4. Maximal Fat Oxidation (MFO) and Cardiorespiratory Fitness (VO2peak)
2.4. Statistical Analyses
3. Results
3.1. Descriptive Statistics and Differences by Sex
3.2. Associations of Leptin Concentration with Fat Oxidation and Insulin Sensitivity in Men and Women
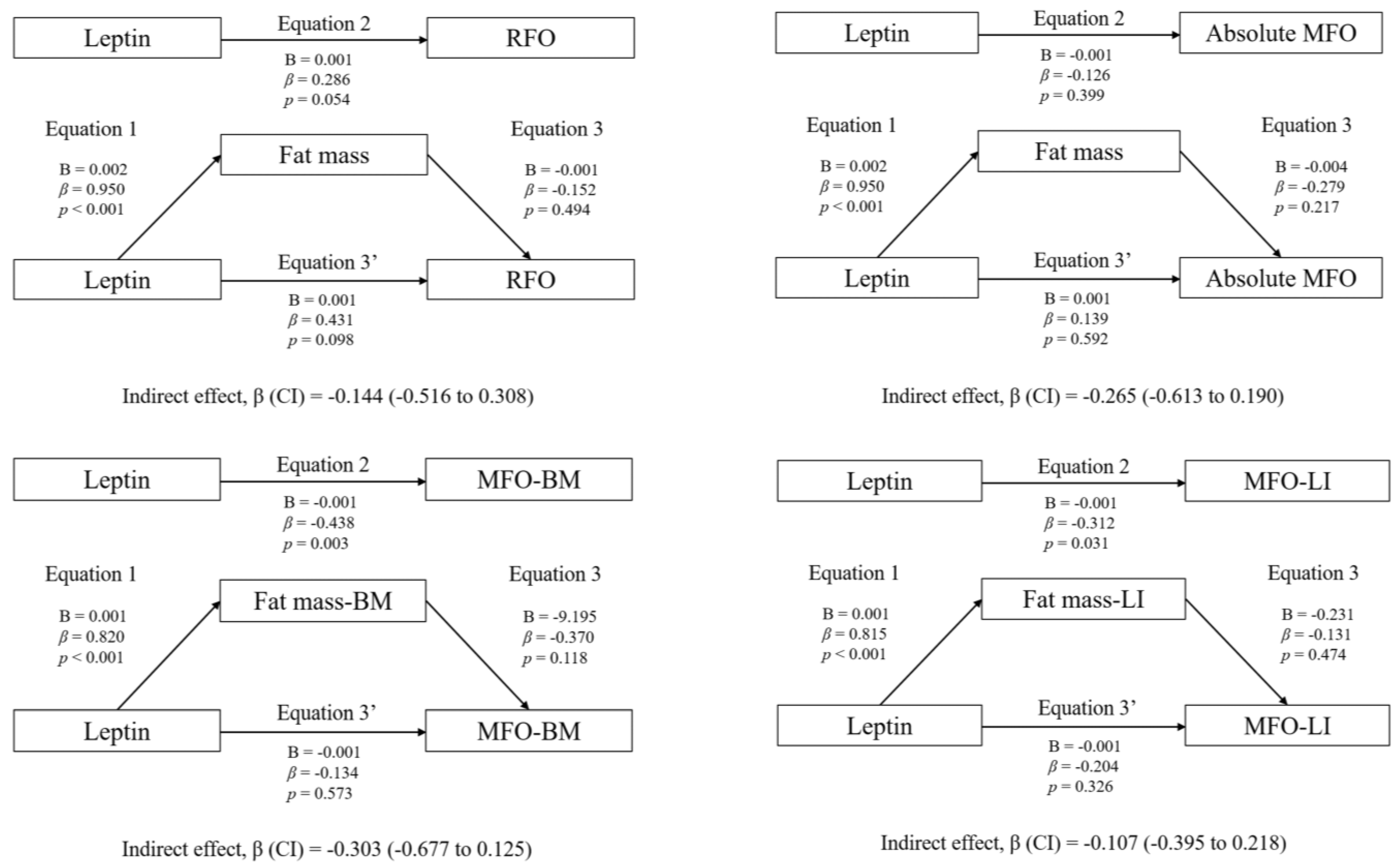
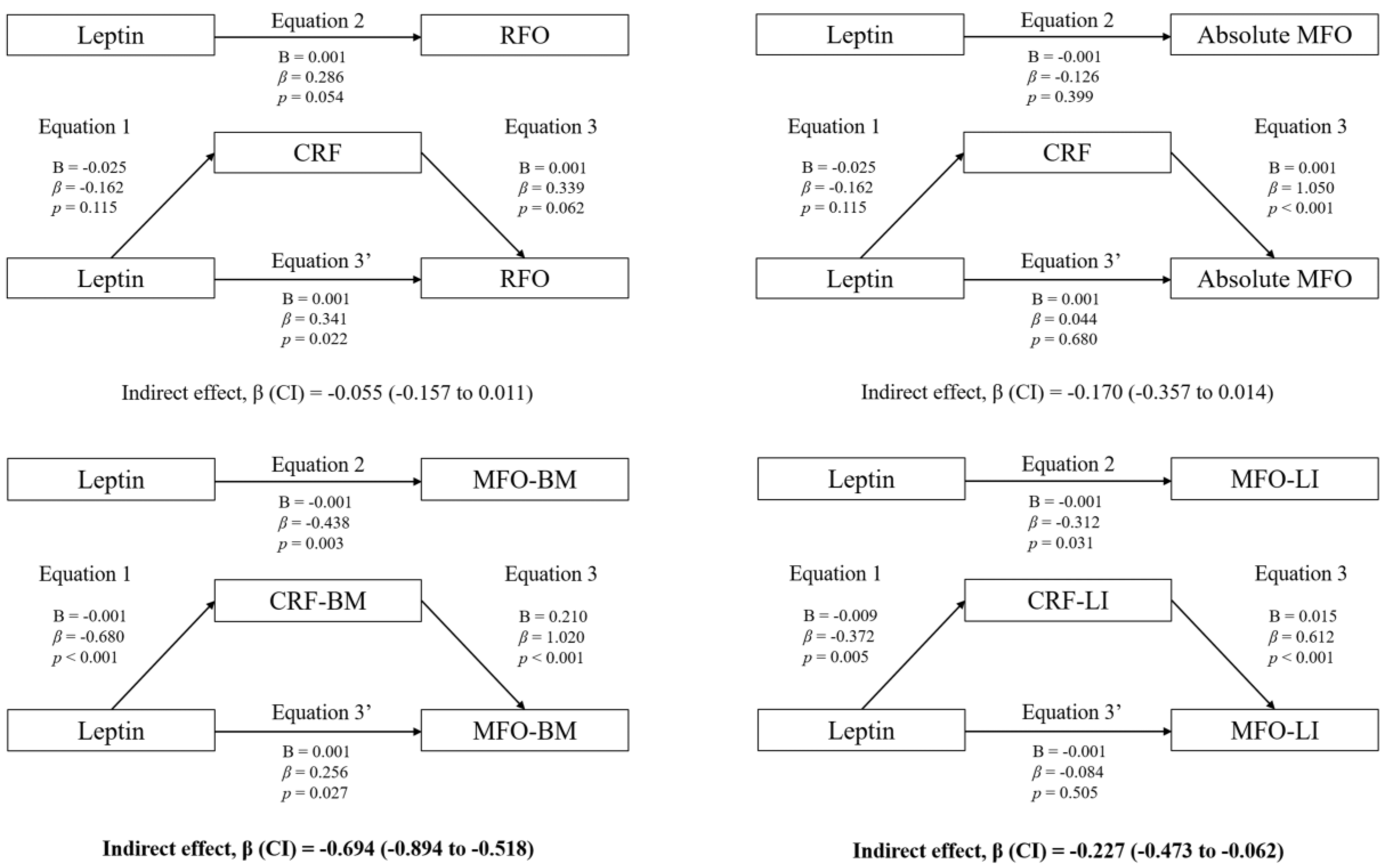
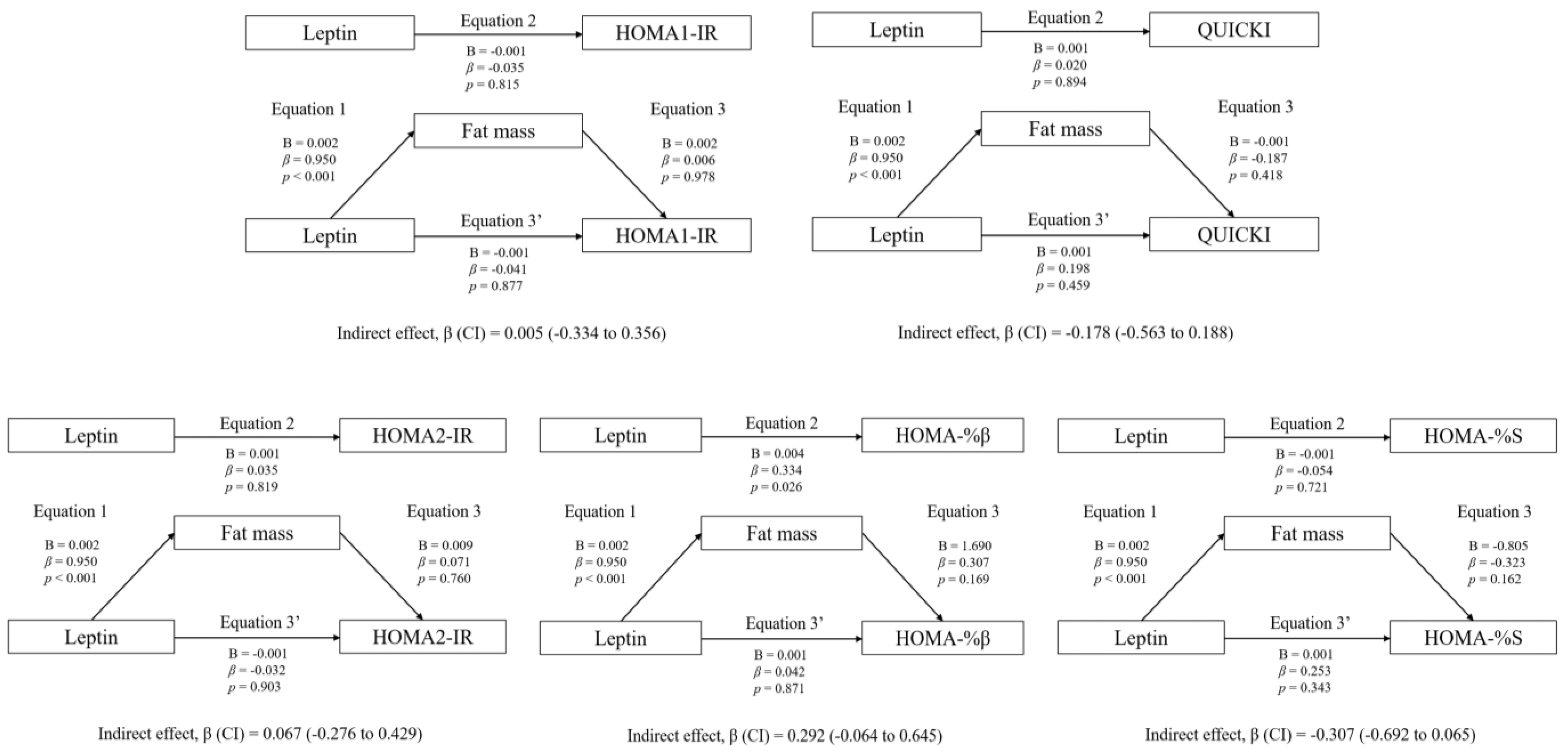
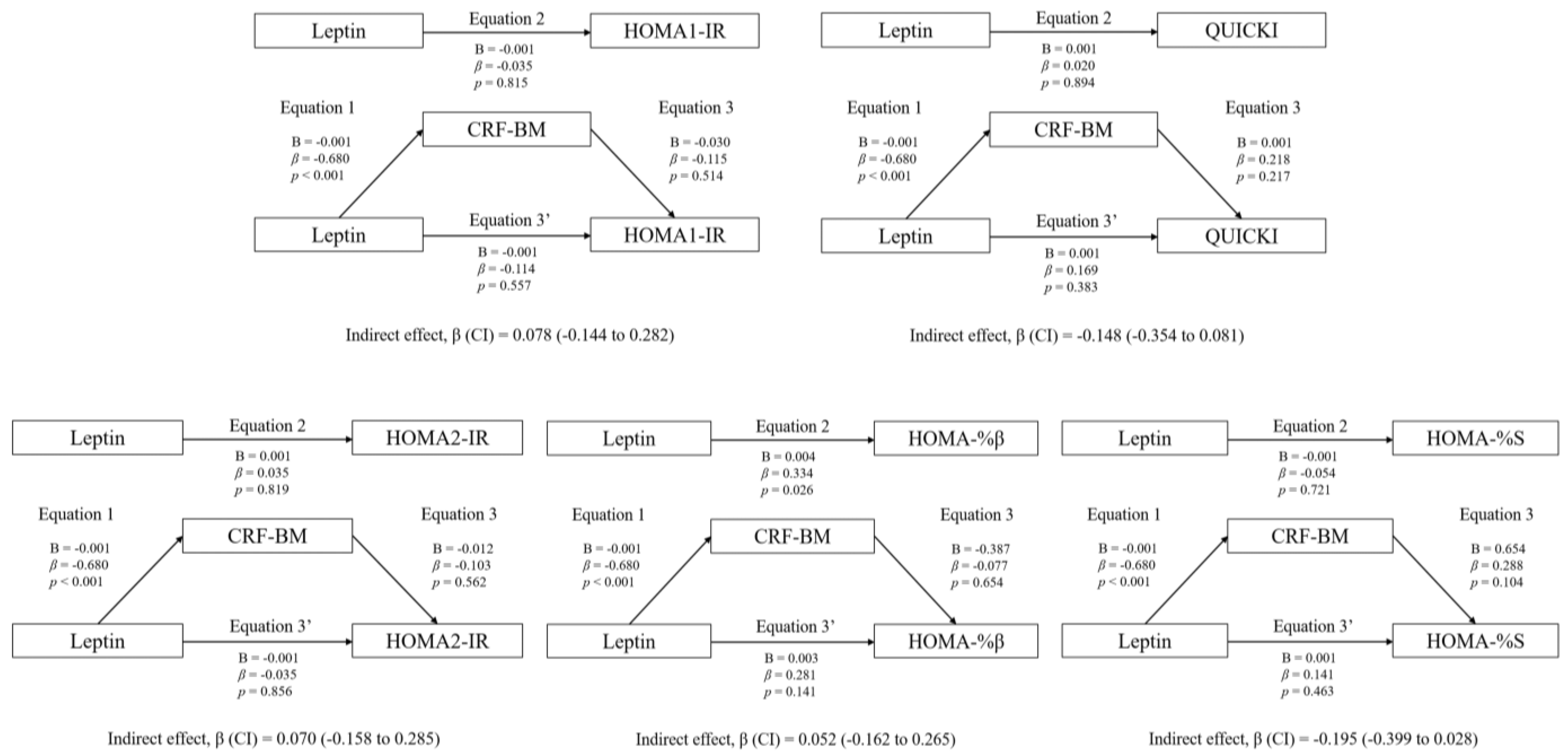
3.3. Mediation Analyses: Adiposity and CRF
4. Discussion
4.1. Main Research Findings
4.2. Association between Plasma Leptin and RFO
4.3. Association between Plasma Leptin and MFO: The Mediating Role of CRF and Adiposity
4.4. Association between Plasma Leptin and Insulin
4.5. Limitations, Strengths, and Future Lines of Research
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montes-de-Oca-García, A.; Perez-Bey, A.; Corral-Pérez, J.; Velázquez-Díaz, D.; Opazo-Díaz, E.; Fernandez-Santos, J.R.; Rebollo-Ramos, M.; Amaro-Gahete, F.J.; Cuenca-García, M.; Ponce-González, J.G. Maximal Fat Oxidation Capacity Is Associated with Cardiometabolic Risk Factors in Healthy Young Adults. Eur. J. Sport Sci. 2020, 21, 907–917. [Google Scholar] [CrossRef]
- Tikkanen, E.; Gustafsson, S.; Ingelsson, E. Associations of Fitness, Physical Activity, Strength, and Genetic Risk with Cardiovascular Disease: Longitudinal Analyses in the UK Biobank Study. Circulation 2018, 137, 2583–2591. [Google Scholar] [CrossRef] [PubMed]
- Amaro-Gahete, F.J.; Sanchez-Delgado, G.; Ara, I.; Ruiz, J.R. Cardiorespiratory Fitness May Influence Metabolic Inflexibility during Exercise in Obese Persons. J. Clin. Endocrinol. Metab. 2019, 104, 5780–5790. [Google Scholar] [CrossRef] [PubMed]
- Montes-De-Oca-García, A.; Perez-Bey, A.; Velázquez-Díaz, D.; Corral-Pérez, J.; Opazo-Díaz, E.; Rebollo-Ramos, M.; Gómez-Gallego, F.; Cuenca-García, M.; Casals, C.; Ponce-González, J.G. Influence of Ace Gene I/D Polymorphism on Cardiometabolic Risk, Maximal Fat Oxidation, Cardiorespiratory Fitness, Diet and Physical Activity in Young Adults. Int. J. Environ. Res. Public Health 2021, 18, 3443. [Google Scholar] [CrossRef] [PubMed]
- Ormazabal, V.; Nair, S.; Elfeky, O.; Aguayo, C.; Salomon, C.; Zuñiga, F.A. Association between Insulin Resistance and the Development of Cardiovascular Disease. Cardiovasc. Diabetol. 2018, 17, 122. [Google Scholar] [CrossRef]
- Landecho, M.F.; Tuero, C.; Valentí, V.; Bilbao, I.; de la Higuera, M.; Frühbeck, G. Relevance of Leptin and Other Adipokines in Obesity-Associated Cardiovascular Risk. Nutrients 2019, 11, 2664. [Google Scholar] [CrossRef]
- Chen, W.; Balland, E.; Cowley, M.A. Hypothalamic Insulin Resistance in Obesity: Effects on Glucose Homeostasis. Neuroendocrinology 2017, 104, 364–381. [Google Scholar] [CrossRef]
- Katsiki, N.; Mikhailidis, D.P.; Banach, M. Leptin, Cardiovascular Diseases and Type 2 Diabetes Mellitus Review-Article. Acta Pharm. Sin. 2018, 39, 1176–1188. [Google Scholar] [CrossRef]
- Negrea, M.O.; Neamtu, B.; Dobrotă, I.; Sofariu, C.R.; Crisan, R.M.; Ciprian, B.I.; Domnariu, C.D.; Teodoru, M. Causative Mechanisms of Childhood and Adolescent Obesity Leading to Adult Cardiometabolic Disease: A Literature Review. Appl. Sci. 2021, 11, 11565. [Google Scholar] [CrossRef]
- Tsiroukidou, K.; Hatziagorou, E.; Grammatikopoulou, M.G.; Vamvakis, A.; Kontouli, K.; Tzimos, C.; Tsanakas, J.; Spiliotis, B.E. Cardiorespiratory Fitness Predicted by Fibrinogen and Leptin Concentrations in Children with Obesity and Risk for Diabetes: A Cross-Sectional Study and a Roc Curve Analysis. Nutrients 2021, 13, 674. [Google Scholar] [CrossRef]
- Antunes, B.M.; Rossi, F.E.; Oyama, L.M.; Rosa-Neto, J.C.; Lira, F.S. Exercise Intensity and Physical Fitness Modulate Lipoproteins Profile during Acute Aerobic Exercise Session. Sci. Rep. 2020, 10, 4160. [Google Scholar] [CrossRef]
- Brand, C.; Leite, N.; Lopes, W.A.; Dias, A.F.; Da Silva, L.R.; Araujo Gaya, A.C.; Mota, J.; Cavaglieri, C.R.; Gaya, A.R. Adiposity and Adipocytokines: The Moderator Role of Cardiorespiratory Fitness and Pubertal Stage in Girls. J. Pediatr. Endocrinol. Metab. 2019, 32, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Carbone, S.; Del Buono, M.G.; Ozemek, C.; Lavie, C.J. Obesity, Risk of Diabetes and Role of Physical Activity, Exercise Training and Cardiorespiratory Fitness. Prog. Cardiovasc. Dis. 2019, 62, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Fuentes, T.; Delgado-Guerra, S.; Guadalupe-Grau, A.; Olmedillas, H.; Santana, A.; Ponce-Gonzalez, J.G.; Dorado, C.; Calbet, J.A.L. Gender Dimorphism in Skeletal Muscle Leptin Receptors, Serum Leptin and Insulin Sensitivity. PLoS ONE 2008, 3, e3466. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Amaro-Gahete, F.J.; Landgrebe, A.; Dela, F.; Ruiz, J.R.; Helge, J.W.; Larsen, S. The Influence of Age, Sex and Cardiorespiratory Fitness on Maximal Fat Oxidation Rate. Appl. Physiol. Nutr. Metab. 2021, 46, 1241–1247. [Google Scholar] [CrossRef]
- Rebollo-Ramos, M.; Velázquez-Díaz, D.; Corral-Pérez, J.; Barany-Ruiz, A.; Pérez-Bey, A.; Fernández-Ponce, C.; García-Cózar, F.J.; Ponce-González, J.G.; Cuenca-García, M. Aerobic Fitness, Mediterranean Diet and Cardiometabolic Risk Factors in Adults. Endocrinol. Diabetes Nutr. 2019, 67, 113–121. [Google Scholar] [CrossRef]
- Corral-Pérez, J.; Velázquez-Díaz, D.; Perez-Bey, A.; Montes-de-Oca-García, A.; Fernandez-Santos, J.R.; Amaro-Gahete, F.J.; Jiménez-Pavón, D.; Casals, C.; Ponce-González, J.G. Accelerometer-Measured Physical Activity and Sedentary Time Are Associated with Maximal Fat Oxidation in Young Adults. Eur. J. Sport Sci. 2021, 22, 1595–1604. [Google Scholar] [CrossRef]
- Besnier, F.; Lenclume, V.; Gérardin, P.; Fianu, A.; Martinez, J.; Naty, N.; Porcherat, S.; Boussaid, K.; Schneebeli, S.; Jarlet, E.; et al. Individualized Exercise Training at Maximal Fat Oxidation Combined with Fruit and Vegetable-Rich Diet in Overweight or Obese Women: The LIPOXmax-Réunion Randomized Controlled Trial. PLoS ONE 2015, 10, e0139246. [Google Scholar] [CrossRef]
- Knopp, J.L.; Holder-Pearson, L.; Chase, J.G. Insulin Units and Conversion Factors: A Story of Truth, Boots, and Faster Half-Truths. J. Diabetes Sci. Technol. 2019, 13, 597–600. [Google Scholar] [CrossRef]
- Montes-de-Oca-García, A.; Corral-Pérez, J.; Velázquez-Díaz, D.; Perez-Bey, A.; Rebollo-Ramos, M.; Marín-Galindo, A.; Gómez-Gallego, F.; Calderon-Dominguez, M.; Casals, C.; Ponce-González, J.G. Influence of Peroxisome Proliferator-Activated Receptor (PPAR)-Gamma Coactivator (PGC)-1 Alpha Gene Rs8192678 Polymorphism by Gender on Different Health-Related Parameters in Healthy Young Adults. Front. Physiol. 2022, 13, 885185. [Google Scholar] [CrossRef]
- Frayn, K.N. Calculation of Substrate Oxidation Rates in Vivo from Gaseous Exchange. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the Exercise Intensity That Elicits Maximal Fat Oxidation Shifts in Energy Substrate Mobilization and Utilization. Med. Sci. Sport. Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef] [PubMed]
- CHENEVIÈRE, X.; MALATESTA, D.; PETERS, E.M.; BORRANI, F. A Mathematical Model to Describe Fat Oxidation Kinetics during Graded Exercise. Med. Sci. Sport. Exerc. 2009, 41, 1615–1625. [Google Scholar] [CrossRef] [PubMed]
- Bolin, J.H. Andrew F. Hayes. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach. New York, NY: The Guilford Press. J. Educ. Meas. 2014, 51, 335–337. [Google Scholar] [CrossRef]
- Preacher, K.J.; Hayes, A.F. SPSS and SAS Procedures for Estimating Indirect Effects in Simple Mediation Models. Behav. Res. Methods Instrum. Comput. 2004, 36, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Goossens, G.H.; Jocken, J.W.E.; Blaak, E.E. Sexual Dimorphism in Cardiometabolic Health: The Role of Adipose Tissue, Muscle and Liver. Nat. Rev. Endocrinol. 2021, 17, 47–66. [Google Scholar] [CrossRef]
- Randell, R.K.; Rollo, I.; Roberts, T.J.; Dalrymple, K.J.; Jeukendrup, A.E.; Carter, J.M. Maximal Fat Oxidation Rates in an Athletic Population. Med. Sci. Sport. Exerc. 2017, 49, 133–140. [Google Scholar] [CrossRef]
- Kovarik, M.; Najpaverova, S.; Koblizek, V.; Zadak, Z.; Hronek, M. Association of Resting Energy Expenditure and Nutritional Substrate Oxidation with COPD Stage and Prediction Indexes. Respir. Med. 2020, 174, 106174. [Google Scholar] [CrossRef]
- Karina Zapata, J.; Catalán, V.; Rodríguez, A.; Ramírez, B.; Silva, C.; Escalada, J.; Salvador, J.; Calamita, G.; Cristina Azcona-Sanjulian, M.; Frühbeck, G.; et al. Resting Energy Expenditure Is Not Altered in Children and Adolescents with Obesity. Effect of Age and Gender and Association with Serum Leptin Levels. Nutrients 2021, 13, 1216. [Google Scholar] [CrossRef]
- Sanches, R.B.; Poli, V.F.S.; Fidalgo, J.P.N.; Andrade-Silva, S.G.; Cerrone, L.A.; Oyama, L.M.; Dâmaso, A.R.; dos Santos, R.T.; Caranti, D.A. The Hyperleptinemia State Can Downregulate Cardiorespiratory Fitness and Energy Expenditure in Obese Women. Physiol. Behav. 2018, 184, 34–38. [Google Scholar] [CrossRef]
- Zampino, M.; Semba, R.D.; Adelnia, F.; Spencer, R.G.; Fishbein, K.W.; Schrack, J.A.; Simonsick, E.M.; Ferrucci, L. Greater Skeletal Muscle Oxidative Capacity Is Associated With Higher Resting Metabolic Rate: Results From the Baltimore Longitudinal Study of Aging. J. Gerontol. A Biol. Sci. Med. Sci. 2020, 75, 2262–2268. [Google Scholar] [CrossRef] [PubMed]
- Adelantado-Renau, M.; Jiménez-Pavón, D.; Beltran-Valls, M.R.; Ponce-González, J.G.; Chiva-Bartoll, Ó.; Moliner-Urdiales, D. Fitness and Academic Performance in Adolescents. The Mediating Role of Leptin: DADOS Study. Eur. J. Pediatr. 2018, 177, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Ara, I.; Larsen, S.; Stallknecht, B.; Guerra, B.; Morales-Alamo, D.; Andersen, J.L.; Ponce-González, J.G.; Guadalupe-Grau, A.; Galbo, H.; Calbet, J.A.L.; et al. Normal Mitochondrial Function and Increased Fat Oxidation Capacity in Leg and Arm Muscles in Obese Humans. Int. J. Obes. 2011, 35, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Olmedillas, H.; Guerra, B.; Guadalupe-Grau, A.; Santana, A.; Fuentes, T.; Dorado, C.; Serrano-Sanchez, J.A.; Calbet, J.A.L. Training, Leptin Receptors and SOCS3 in Human Muscle. Int. J. Sport. Med. 2011, 32, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Frandsen, J.; Hansen, I.M.D.; Wismann, J.F.; Olsen, M.H.; Brage-Andersen, M.R.; Sahl, R.E.; Hansen, M.; Ingersen, A.; Modvig, J.L.; Schmücker, M.; et al. Maximal Fat Oxidation Rate Is Higher in Fit Women and Unfit Women With Obesity, Compared to Normal-Weight Unfit Women. J. Clin. Endocrinol. Metab. 2021, 106, e4389–e4399. [Google Scholar] [CrossRef]
- Dragović, G.; Sumarac-Dumanovic, M.; Khawla, A.M.; Soldatović, I.; Andjić, M.; Jevtović, D.; Nair, D. Correlation between PAI-1, Leptin and Ferritin with HOMA in HIV/AIDS Patients. Exp. Mol. Pathol. 2018, 105, 115–119. [Google Scholar] [CrossRef]
- Frithioff-Bøjsøe, C.; Lund, M.A.V.; Lausten-Thomsen, U.; Hedley, P.L.; Pedersen, O.; Christiansen, M.; Baker, J.L.; Hansen, T.; Holm, J.C. Leptin, Adiponectin, and Their Ratio as Markers of Insulin Resistance and Cardiometabolic Risk in Childhood Obesity. Pediatr. Diabetes 2020, 21, 194–202. [Google Scholar] [CrossRef]
- Blüher, M. Metabolically Healthy Obesity. Endocr. Rev. 2020, 41, 405–420. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total (n = 65) | Men (n = 42) | Women (n = 23) | p | d | |
|---|---|---|---|---|---|
| Age (years) | 22.55 ± 4.30 | 22.29 ± 3.79 | 23.04 ± 5.17 | 0.502 | −0.17 |
| Height (cm) | 172.58 ± 8.34 | 176.61 ± 6.30 | 165.21 ± 6.38 | <0.001 | 1.80 |
| Body Mass (kg) | 75.28 ± 15.30 | 78.16 ± 14.91 | 70.01 ± 14.88 | 0.039 | 0.55 |
| BMI (kg·m−2) | 25.26 ± 4.74 | 25.01 ± 4.17 | 25.70 ± 5.70 | 0.574 | −0.14 |
| Total lean mass (%) | 73.35 ± 8.48 | 76.95 ± 6.58 | 66.76 ± 7.62 | <0.001 | 1.46 |
| Body fat (%) | 22.17 ± 9.27 | 18.49 ± 7.40 | 28.86 ± 8.69 | <0.001 | −1.32 |
| VO2peak (mL·kg−1·min−1) | 41.79 ± 11.18 | 45.19 ± 10.56 | 35.57 ± 9.66 | 0.001 | 0.94 |
| Leptin (pg·mL−1) | 3881.94 ± 4593.64 | 1993.93 ± 2957.42 | 7329.59 ± 5084.73 | <0.001 | −1.39 |
| Insulin (pg·mL−1) | 644.25 ± 382.70 | 678.51 ± 417.50 | 581.70 ± 308.04 | 0.333 | 0.25 |
| Glucose (mg·dL−1) | 101.62 ± 9.59 | 103.45 ± 9.21 | 98.26 ± 9.56 | 0.036 | 0.56 |
| HOMA1-IR | 4.67 ± 2.92 | 5.01 ± 3.17 | 4.07 ± 2.35 | 0.225 | 0.32 |
| HOMA2-IR | 2.40 ± 1.36 | 2.53 ± 1.48 | 2.16 ± 1.11 | 0.301 | 0.27 |
| HOMA-%β | 135.28 ± 56.07 | 134.86 ± 58.12 | 136.03 ± 53.37 | 0.937 | −0.02 |
| HOMA-%S | 53.47 ± 25.40 | 51.87 ± 25.82 | 56.41 ± 24.92 | 0.495 | −0.18 |
| QUICKI | 0.313 ± 0.022 | 0.310 ± 0.023 | 0.317 ± 0.020 | 0.263 | −0.32 |
| RFO (mg·min−1) | 98.39 ± 27.29 | 102.30 ± 29.99 | 91.23 ± 20.17 | 0.119 | 0.41 |
| MFO (mg·min−1) | 378.35 ± 156.20 | 403.25 ± 171.04 | 332.86 ± 114.51 | 0.053 | 0.46 |
| MFO-BM (mg·kg−1·min−1) | 5.21 ± 2.30 | 5.33 ± 2.48 | 4.98 ± 1.96 | 0.532 | 0.15 |
| MFO-LI (mg·(kg·m−2)−1·min−1) | 6.99 ± 2.93 | 6.38 ± 2.68 | 8.09 ± 3.09 | 0.023 | −0.60 |
| Model 1 | Model 2 | Model 3 | Model 4 | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| β | R2 | p | β | R2 | p | β | R2 | p | β | R2 | p | |
| Men (n = 42) | ||||||||||||
| Fatty acid oxidation | ||||||||||||
| RFO (mg·min−1) | 0.122 | 0.015 | 0.441 | 0.140 | 0.020 | 0.396 | 0.144 | 0.015 | 0.641 | 0.171 | 0.078 | 0.283 |
| MFO (mg·min−1) | −0.128 | 0.016 | 0.418 | −0.047 | 0.126 | 0.763 | 0.007 | 0.023 | 0.981 | 0.008 | 0.511 | 0.946 |
| MFO-BM (mg·kg−1·min−1) | −0.359 | 0.129 | 0.020 | −0.270 | 0.258 | 0.065 | −0.239 | 0.136 | 0.366 | 0.202 | 0.675 | 0.086 |
| MFO-LI (mg·(kg·m−2)−1·min−1) | −0.234 | 0.055 | 0.135 | −0.138 | 0.208 | 0.353 | −0.184 | 0.057 | 0.441 | −0.125 | 0.329 | 0.359 |
| Insulin resistance | ||||||||||||
| HOMA1-IR | 0.143 | 0.021 | 0.365 | 0.156 | 0.023 | 0.344 | 0.328 | 0.033 | 0.285 | 0.149 | 0.021 | 0.457 |
| HOMA2-IR | 0.199 | 0.040 | 0.206 | 0.216 | 0.044 | 0.189 | 0.352 | 0.048 | 0.249 | 0.225 | 0.041 | 0.259 |
| HOMA-%β | 0.398 | 0.158 | 0.009 | 0.430 | 0.175 | 0.007 | 0.414 | 0.158 | 0.151 | 0.469 | 0.167 | 0.015 |
| Insulin sensitivity | ||||||||||||
| QUICKI | −0.186 | 0.035 | 0.238 | −0.188 | 0.035 | 0.254 | −0.160 | 0.035 | 0.601 | −0.157 | 0.036 | 0.431 |
| HOMA-%S | −0.256 | 0.066 | 0.102 | −0.254 | 0.066 | 0.119 | −0.099 | 0.075 | 0.740 | −0.196 | 0.072 | 0.317 |
| Women (n = 23) | ||||||||||||
| Fatty acid oxidation | ||||||||||||
| RFO (mg·min−1) | 0.501 | 0.251 | 0.015 | 0.584 | 0.311 | 0.007 | 0.807 | 0.296 | 0.023 | 0.538 | 0.271 | 0.013 |
| MFO (mg·min−1) | −0.103 | 0.011 | 0.639 | −0.008 | 0.088 | 0.972 | 0.331 | 0.102 | 0.384 | 0.090 | 0.566 | 0.559 |
| MFO-BM (mg·kg−1·min−1) | −0.436 | 0.190 | 0.037 | −0.309 | 0.330 | 0.126 | 0.039 | 0.300 | 0.906 | 0.239 | 0.650 | 0.215 |
| MFO-LI (mg·(kg·m−2)−1·min−1) | −0.313 | 0.098 | 0.146 | −0.217 | 0.176 | 0.323 | −0.121 | 0.125 | 0.710 | 0.025 | 0.443 | 0.896 |
| Insulin resistance | ||||||||||||
| HOMA1-IR | −0.289 | 0.083 | 0.182 | −0.282 | 0.084 | 0.227 | −0.354 | 0.085 | 0.356 | −0.515 | 0.135 | 0.094 |
| HOMA2-IR | −0.210 | 0.044 | 0.335 | −0.216 | 0.045 | 0.360 | −0.363 | 0.056 | 0.352 | −0.466 | 0.110 | 0.133 |
| HOMA-%β | 0.164 | 0.027 | 0.454 | 0.122 | 0.042 | 0.603 | −0.278 | 0.122 | 0.457 | −0.130 | 0.114 | 0.667 |
| Insulin sensitivity | ||||||||||||
| QUICKI | 0.268 | 0.072 | 0.216 | 0.267 | 0.072 | 0.254 | 0.380 | 0.078 | 0.324 | 0.597 | 0.181 | 0.049 |
| HOMA-%S | 0.187 | 0.035 | 0.393 | 0.191 | 0.035 | 0.421 | 0.384 | 0.054 | 0.326 | 0.559 | 0.175 | 0.065 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montes-de-Oca-García, A.; Perez-Bey, A.; Corral-Pérez, J.; Marín-Galindo, A.; Calderon-Dominguez, M.; Velázquez-Díaz, D.; Casals, C.; Ponce-Gonzalez, J.G. Influence of Gender on Plasma Leptin Levels, Fat Oxidation, and Insulin Sensitivity in Young Adults: The Mediating Role of Fitness and Fatness. Nutrients 2023, 15, 2628. https://doi.org/10.3390/nu15112628
Montes-de-Oca-García A, Perez-Bey A, Corral-Pérez J, Marín-Galindo A, Calderon-Dominguez M, Velázquez-Díaz D, Casals C, Ponce-Gonzalez JG. Influence of Gender on Plasma Leptin Levels, Fat Oxidation, and Insulin Sensitivity in Young Adults: The Mediating Role of Fitness and Fatness. Nutrients. 2023; 15(11):2628. https://doi.org/10.3390/nu15112628
Chicago/Turabian StyleMontes-de-Oca-García, Adrián, Alejandro Perez-Bey, Juan Corral-Pérez, Alberto Marín-Galindo, Maria Calderon-Dominguez, Daniel Velázquez-Díaz, Cristina Casals, and Jesus G. Ponce-Gonzalez. 2023. "Influence of Gender on Plasma Leptin Levels, Fat Oxidation, and Insulin Sensitivity in Young Adults: The Mediating Role of Fitness and Fatness" Nutrients 15, no. 11: 2628. https://doi.org/10.3390/nu15112628
APA StyleMontes-de-Oca-García, A., Perez-Bey, A., Corral-Pérez, J., Marín-Galindo, A., Calderon-Dominguez, M., Velázquez-Díaz, D., Casals, C., & Ponce-Gonzalez, J. G. (2023). Influence of Gender on Plasma Leptin Levels, Fat Oxidation, and Insulin Sensitivity in Young Adults: The Mediating Role of Fitness and Fatness. Nutrients, 15(11), 2628. https://doi.org/10.3390/nu15112628