
Nutritional Supplementation and Exercise as Essential Allies in the Treatment of Chronic Heart Failure: The Metabolic and Molecular Bases




{kind=link}
{kind=link}
Highlights
- Both Physical Exercise (PE) and Nutritional Supplementation (NS), combined with traditional therapy, are promising integrated therapeutic strategies to treat patients with Chronic Heart Failure (CHF);
- Both PE and NS stimulate anabolic metabolic pathways which counteract the hypercatabolic syndrome responsible for CHF worsening.
- Personalised PE and NS with specific nutritional mixtures based on human metabolic needs are allied with the concomitant therapies in the treatment of CHF;
- This article presents and discusses the metabolic and molecular evidence that support the use of combined personalised PE and NS in CHF patients.
Abstract
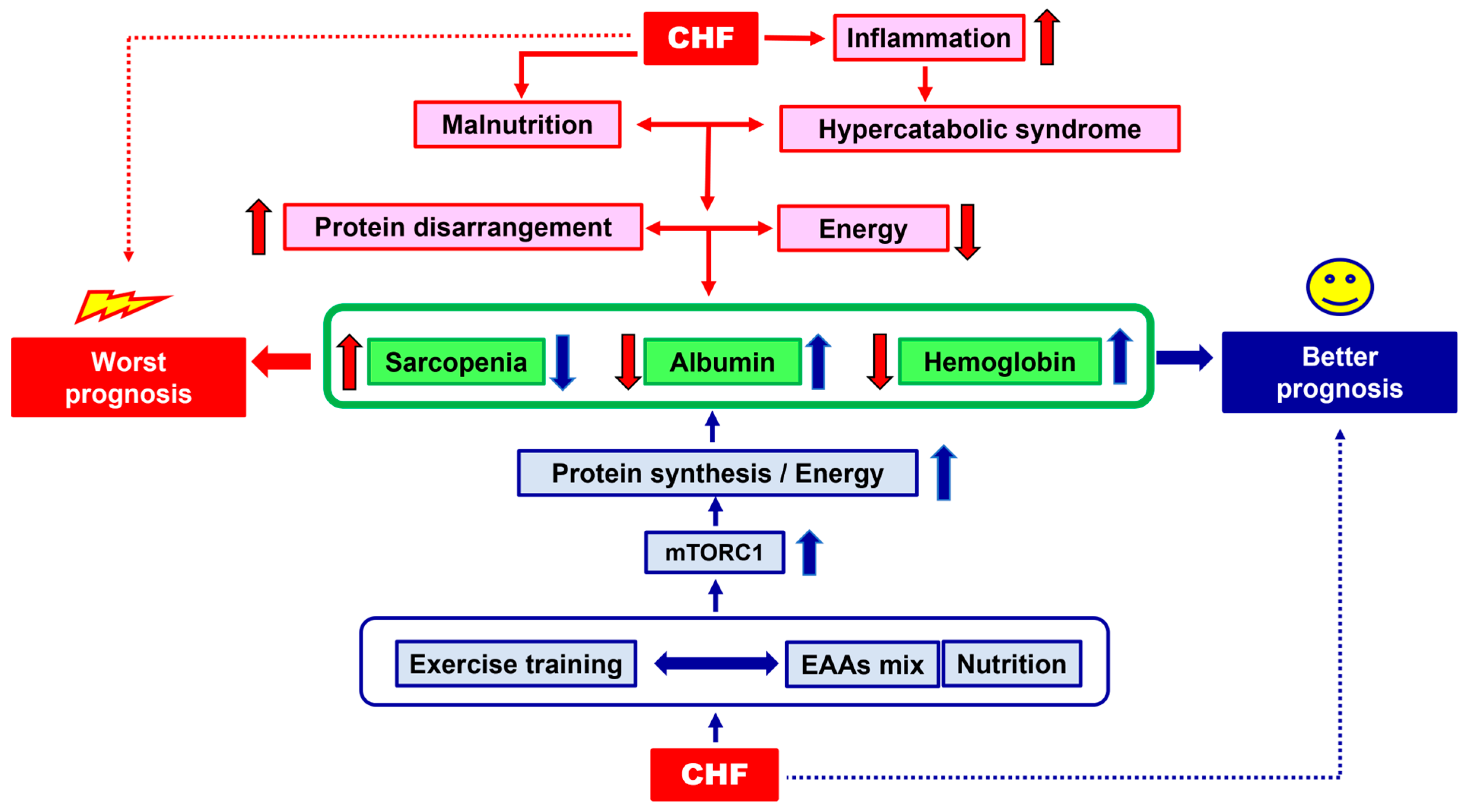
1. The Clinical Problem
2. Malnutrition: Quantification and Causes
3. Integrated Therapeutical Strategies: Exercise Training and Nutritional Supplementation
3.1. Exercise Training
3.2. Nutritional Supplementation
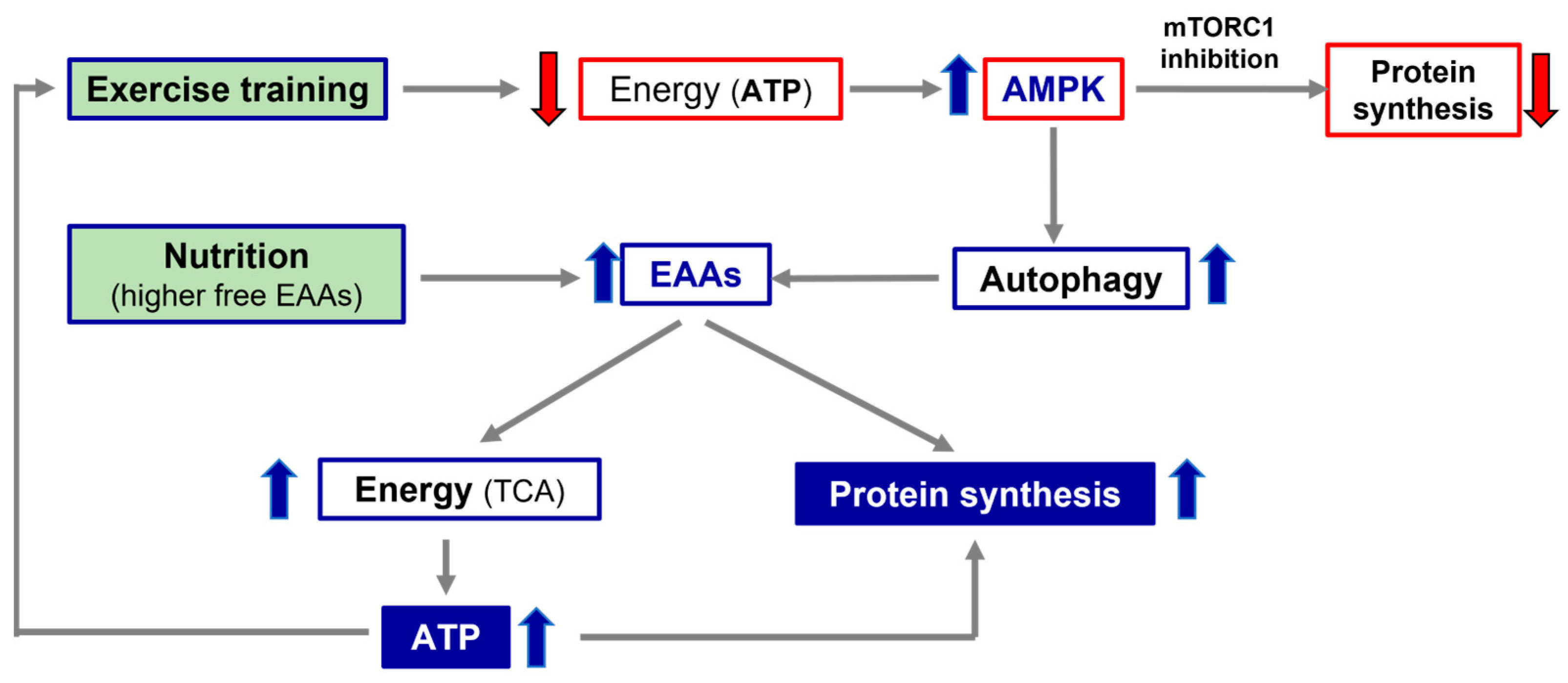
4. Molecular Hypothesis of Exercise Training and Nutritional Supplementation Alliance: The Roles of AMPK and mTOR
5. Conclusions and Clinical Suggestions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sandek, A.; Doehner, W.; Anker, S.D.; von Haehling, S. Nutrition in heart failure: An update. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 384–391. [Google Scholar] [CrossRef]
- Butler, J.; Kalogeropoulos, A. Worsening heart failure hospitalization epidemic we do not know how to prevent and we do not know how to treat! J. Am. Coll. Cardiol. 2008, 52, 435–437. [Google Scholar] [CrossRef][Green Version]
- Aquilani, R.; Opasich, C.; Dossena, M.; Iadarola, P.; Gualco, A.; Arcidiaco, P.; Viglio, S.; Boschi, F.; Verri, M.; Pasini, E. Increased skeletal muscle amino acid release with light exercise in deconditioned patients with heart failure. J. Am. Coll. Cardiol. 2005, 45, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Corsetti, G.; Romano, C.; Aquilani, R.; Scarabelli, T.; Chen-Scarabelli, C.; Dioguardi, F.S. Management of Anaemia of Chronic Disease: Beyond Iron-Only Supplementation. Nutrients 2021, 13, 237. [Google Scholar] [CrossRef]
- von Haehling, S.; Ebner, N.; Dos Santos, M.R.; Springer, J.; Anker, S.D. Muscle wasting and cachexia in heart failure: Mechanisms and therapies. Nat. Rev. Cardiol. 2017, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Anker, S.D.; Coats, A.J. Cardiac cachexia: A syndrome with impaired survival and immune and neuroendocrine activation. Chest 1999, 115, 836–847. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Aquilani, R.; Corsetti, G.; Dioguardi, F.S. Malnutrition and Gut Flora Dysbiosis: Specific Therapies for Emerging Comorbidities in Heart Failure. Biomed. Res. Int. 2015, 2015, 382585. [Google Scholar] [CrossRef] [PubMed]
- Aquilani, R.; Opasich, C.; Verri, M.; Boschi, F.; Febo, O.; Pasini, E.; Pastoris, O. Is nutritional intake adequate in chronic heart failure patients? J. Am. Coll. Cardiol. 2003, 42, 1218–1223. [Google Scholar] [CrossRef]
- Pasini, E.; Aquilani, R.; Dioguardi, F.S. The enemy within. How to identify chronic diseases induced-protein metabolism impairment and its possible pharmacological treatment. Pharmacol. Res. 2013, 76, 28–33. [Google Scholar] [CrossRef]
- Akirov, A.; Masri-Iraqi, H.; Atamna, A.; Shimon, I. Low Albumin Levels Are Associated with Mortality Risk in Hospitalized Patients. Am. J. Med. 2017, 130, 1465.e11–1465.e19. [Google Scholar] [CrossRef]
- Aquilani, R.; Zuccarelli, G.C.; Condino, A.M.; Catani, M.; Rutili, C.; Del Vecchio, C.; Pisano, P.; Verri, M.; Iadarola, P.; Viglio, S.; et al. Despite Inflammation, Supplemented Essential Amino Acids May Improve Circulating Levels of Albumin and Haemoglobin in Patients after Hip Fractures. Nutrients 2017, 9, 637. [Google Scholar] [CrossRef] [PubMed]
- Dioguardi, F.S. Where the Pendulum of Knowledge Stands Now: Is Circulating Albumin a Marker of Inflammation or of Malnutrition? How to Manage Hypoalbuminemia by Nutrition? J. Nutr. Biol. 2018, 4, 217. [Google Scholar] [CrossRef][Green Version]
- Soukoulis, V.; Dihu, J.B.; Sole, M.; Anker, S.D.; Cleland, J.; Fonarow, G.C.; Metra, M.; Pasini, E.; Strzelczyk, T.; Taegtmeyer, H.; et al. Micronutrient deficiencies an unmet need in heart failure. J. Am. Coll. Cardiol. 2009, 54, 1660–1673. [Google Scholar] [CrossRef]
- Lehninger, A.L.; Nelson, D.L.; Cox, M.M. Principles of Biochemistry, 2nd ed.; Worth Publishers Inc.: New York, NY, USA, 2000. [Google Scholar]
- O’Keeffe, M.; Kelly, M.; O’Herlihy, E.; O’Toole, P.W.; Kearney, P.M.; Timmons, S.; O’Shea, E.; Stanton, C.; Hickson, M.; Rolland, Y.; et al. Potentially modifiable determinants of malnutrition in older adults: A systematic review. Clin. Nutr. 2019, 38, 2477–2498. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Aquilani, R.; Testa, C.; Baiardi, P.; Angioletti, S.; Boschi, F.; Verri, M.; Dioguardi, F.S. Pathogenic Gut Flora in Patients with Chronic Heart Failure. J. Am. Coll. Cardiol. Heart Fail. 2014, 4, 220–227. [Google Scholar] [CrossRef]
- Pasini, E.; Flati, V.; Paiardi, S.; Rizzoni, D.; Porteri, E.; Aquilani, R.; Assanelli, D.; Corsetti, G.; Speca, S.; Rezzani, R.; et al. Intracellular molecular effects of insulin resistance in patients with metabolic syndrome. Cardiovasc. Diabetol. 2010, 9, 46–50. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.; Aquilani, R.; Dioguardi, F.S. Amino acids: Chemistry and metabolism in normal and hypercatabolic states. Am. J. Cardiol. 2004, 93, 3A–5A. [Google Scholar] [CrossRef]
- Pasini, E.; Aquilani, R.; Dioguardi, F.S.; D’Antona, G.; Gheorghiade, M.; Taegtmeyer, H. Hypercatabolic syndrome: Molecular basis and effects of nutritional supplements with amino acids. Am. J. Cardiol. 2008, 101, 11E–15E. [Google Scholar] [CrossRef]
- Pasini, E.; Corsetti, G.; Aquilani, R.; Romano, C.; Picca, A.; Calvani, R.; Dioguardi, F.S. Protein-Amino Acid Metabolism Disarrangements: The Hidden Enemy of Chronic Age-Related Conditions. Nutrients 2018, 10, 391. [Google Scholar] [CrossRef]
- Haupt, S.; Niedrist, T.; Sourij, S.; Schwarzinger, S.; Moser, O. The impact of exercise on telomere length, DNA methylation and metabolic footprints. Cell 2022, 11, 153. [Google Scholar] [CrossRef]
- Hasten, D.L.; Pak-Loduca, J.; Obert, K.A.; Yarasheski, K.E. Resistance exercise acutely increases MHC and mixed muscle protein synthesis rates in 78–84 and 23–32 yr olds. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E620–E626. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.S.; Dalal, H.M.; McDonagh, S.T.J. The role of cardiac rehabilitation in improving cardiovascular outcomes. Nat. Rev. Cardiol. 2022, 19, 180–194. [Google Scholar] [CrossRef] [PubMed]
- Bull, F.C.; Al-Ansari, S.S.; Biddle, S.; Borodulin, K.; Bumanat, M.P.; Cardonal, G.; Carty, C.; Chaput, J.P.; Chastin, S.; Chou, R.; et al. World Health Organization 2020 guidelines on physical activity and sedentary behaviour. Br. J. Sports Med. 2020, 54, 1451. [Google Scholar] [CrossRef] [PubMed]
- Aquilani, R.; Opasich, C.; Gualco, A.; Verri, M.; Testa, A.; Pasini, E.; Viglio, S.; Iadarola, P.; Pastoris, O.; Dossena, M.; et al. Adequate energy-protein intake is not enough to improve nutritional and metabolic status in muscle-depleted patients with chronic heart failure. Eur. J. Heart Fail. 2008, 10, 1127–1135. [Google Scholar] [CrossRef] [PubMed]
- Gwin, J.A.; Church, D.D.; Hatch-McChesney, A.; Allen, J.T.; Wilson, M.A.; Varanoske, A.N.; Carrigan, C.T.; Murphy, N.E.; Margolis, L.M.; Carbone, J.W.; et al. Essential amino acid-enriched whey enhances post-exercise whole-body protein balance during energy deficit more than iso-nitrogenous whey or a mixed-macronutrient meal: A randomized, crossover study. J. Int. Soc. Sports Nutr. 2021, 18, 4. [Google Scholar] [CrossRef]
- Azhar, G.; Verma, A.; Zhang, X.; Pangle, A.; Patyal, P.; Zhang, W.; Che, Y.; Coker, K.; Wolfe, R.R.; Wei, J.Y. Differential plasma protein expression after ingestion of essential amino acid-based dietary supplement verses whey protein in low physical functioning older adults. Geroscience 2023. [Google Scholar] [CrossRef] [PubMed]
- Church, D.D.; Hirsch, K.R.; Park, S.; Kim, I.Y.; Gwin, J.A.; Pasiakos, S.M.; Wolfe, R.R.; Ferrando, A.A. Essential Amino Acids and Protein Synthesis: Insights into Maximizing the Muscle and Whole-Body Response to Feeding. Nutrients 2020, 12, 3717. [Google Scholar] [CrossRef]
- Romano, C.; Corsetti, G.; Flati, V.; Pasini, E.; Picca, A.; Calvani, R.; Marzetti, E.; Dioguardi, F.S. Influence of Diets with Varying Essential/Nonessential Amino Acid Ratios on Mouse Lifespan. Nutrients 2019, 1811, 1367. [Google Scholar] [CrossRef] [PubMed]
- D’Antona, G.; Ragni, M.; Cardile, A.; Tedesco, L.; Dossena, M.; Bruttini, F.; Caliaro, F.; Corsetti, G.; Bottinelli, R.; Carruba, M.O.; et al. Branched-chain amino acid supplementation promotes survival and supports cardiac and skeletal muscle mitochondrial biogenesis in middle-aged mice. Cell. Metab. 2010, 12, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Romano, C.; Corsetti, G.; Pasini, E.; Flati, V.; Dioguardi, F.S. Dietary Modifications of Nitrogen Intake Decreases Inflammation and Promotes Rejuvenation of Spleen in Aged Mice. J. Food Nutr. Res. 2018, 6, 419–432. [Google Scholar] [CrossRef][Green Version]
- Wolfson, R.L.; Chantranupong, L.; Saxton, R.A.; Shen, K.; Scaria, S.M.; Cantor, J.R.; Sabatini, D.M. Sestrin2 is a leucine sensor for mTORC1 pathway. Science 2016, 351, 43. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Liu, R.; Xiong, Y.; Li, X.; Wang, X.; Ma, Y.; Guo, H.; Hao, L.; Yao, P.; Liu, L.; et al. Leucine facilitates the insulin-stimulated glucose uptake and insulin signaling in skeletal muscle cells: Involving mTORC1 and mTORC2. Amino Acids 2014, 46, 1971–1979. [Google Scholar] [CrossRef] [PubMed]
- Molfino, A.; Gioia, G.; Rossi Fanelli, F.; Muscaritoli, M. Beta-hydroxy-beta-methylbutyrate supplementation in health and disease: A systematic review of randomized trials. Amino Acids 2013, 45, 1273–1292. [Google Scholar] [CrossRef] [PubMed]
- Valerio, A.; D’Antona, G.; Nisoli, E. Branched-chain amino acids, mitochondrial biogenesis, and healthspan: An evolutionary perspective. Aging 2011, 3, 464–478. [Google Scholar] [CrossRef] [PubMed]
- MacConnell, A.; Robets, L.D. Metbokines in the regulation of systemic energy metaboslim. Curr. Opin. Pharmacol. 2022, 67, 102286. [Google Scholar] [CrossRef] [PubMed]
- Nedaeinia, R. Lifestyl Genomic Interaction in Health and Diseased; Kelishadi, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2022; Volume 3. [Google Scholar]
- Yan, Z. Exercise, PGC1-α and metabolic adaptation in skeletal muscle. Appl. Physiol. Nutr. Metabol. 2009, 34, 424. [Google Scholar] [CrossRef]
- Voulgaridou, G.; Papadopoulou, S.D.; Spanoudaki, M.; Kondyli, F.S.; Alexandropoulou, I.; Michailidou, S.; Zarogoulidis, P.; Matthaios, D.; Giannakidis, D.; Romanidou, M.; et al. Increasing Muscle Mass in Elders through Diet and Exercise: A Literature Review of Recent RCTs. Foods 2023, 12, 1218. [Google Scholar] [CrossRef]
- DeBerardinis, R.J.; Thompson, C.B. Cellular metabolism and disease: What do metabolic outliers teach us? Cell 2012, 148, 1132–1144. [Google Scholar] [CrossRef] [PubMed]
- An, P.; Wan, S.; Luo, Y.; Luo, J.; Zhang, X.; Zhou, S.; Xu, T.; He, J.; Mechanick, J.I.; Wu, W.C.; et al. Micronutrient Supplementation to Reduce Cardiovascular Risk. J. Am. Coll. Cardiol. 2022, 80, 2269–2285. [Google Scholar] [CrossRef] [PubMed]
- Yoon, K.J.; Zhang, D.; Kim, S.J.; Lee, M.C.; Moon, H.Y. Exercise-induced AMPK activation is involved in delay of skeletal muscle senescence. Biochem. Biophys. Res. Commun. 2019, 512, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Lo Verso, F.; Carnio, S.; Vainshtein, A.; Sandri, M. Autophagy is not reguired to sustain exercise and PRKAA1/AMPK activity damage during physical activity. Autophagy 2014, 10, 1883. [Google Scholar] [CrossRef] [PubMed]
- Shimobayashi, M.; Hall, M.N. Making new contacts: The mTOR network in metabolism and signalling crosstalk. Nat. Rev. Mol. Cell Biol. 2014, 15, 155–162. [Google Scholar] [CrossRef]
- Wälchli, M.; Berneiser, K.; Mangia, F.; Imseng, S.; Craigie, L.M.; Stuttfeld, E.; Hall, M.N.; Maier, T. Regulation of human mTOR complexes by DEPTOR. eLife 2021, 10, e70871. [Google Scholar] [CrossRef] [PubMed]
- Shimkus, K.L.; Jefferson, L.S.; Gordon, B.S.; Kimball, S.R. Repressors of mTORC1 act to blunt the anabolic response to feeding in the soleus muscle of a cast-immobilized mouse hindlimb. Physiol. Rep. 2018, 6, e13891. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasini, E.; Corsetti, G.; Dioguardi, F.S. Nutritional Supplementation and Exercise as Essential Allies in the Treatment of Chronic Heart Failure: The Metabolic and Molecular Bases. Nutrients 2023, 15, 2337. https://doi.org/10.3390/nu15102337
Pasini E, Corsetti G, Dioguardi FS. Nutritional Supplementation and Exercise as Essential Allies in the Treatment of Chronic Heart Failure: The Metabolic and Molecular Bases. Nutrients. 2023; 15(10):2337. https://doi.org/10.3390/nu15102337
Chicago/Turabian StylePasini, Evasio, Giovanni Corsetti, and Francesco Saverio Dioguardi. 2023. "Nutritional Supplementation and Exercise as Essential Allies in the Treatment of Chronic Heart Failure: The Metabolic and Molecular Bases" Nutrients 15, no. 10: 2337. https://doi.org/10.3390/nu15102337
APA StylePasini, E., Corsetti, G., & Dioguardi, F. S. (2023). Nutritional Supplementation and Exercise as Essential Allies in the Treatment of Chronic Heart Failure: The Metabolic and Molecular Bases. Nutrients, 15(10), 2337. https://doi.org/10.3390/nu15102337