
Alpha-Lipoic Acid and Glucose Metabolism: A Comprehensive Update on Biochemical and Therapeutic Features
 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Chemistry and Metabolism of ALA
3. ALA: Cellular Energy and Oxidative Stress
4. ALA and Glucose Metabolism
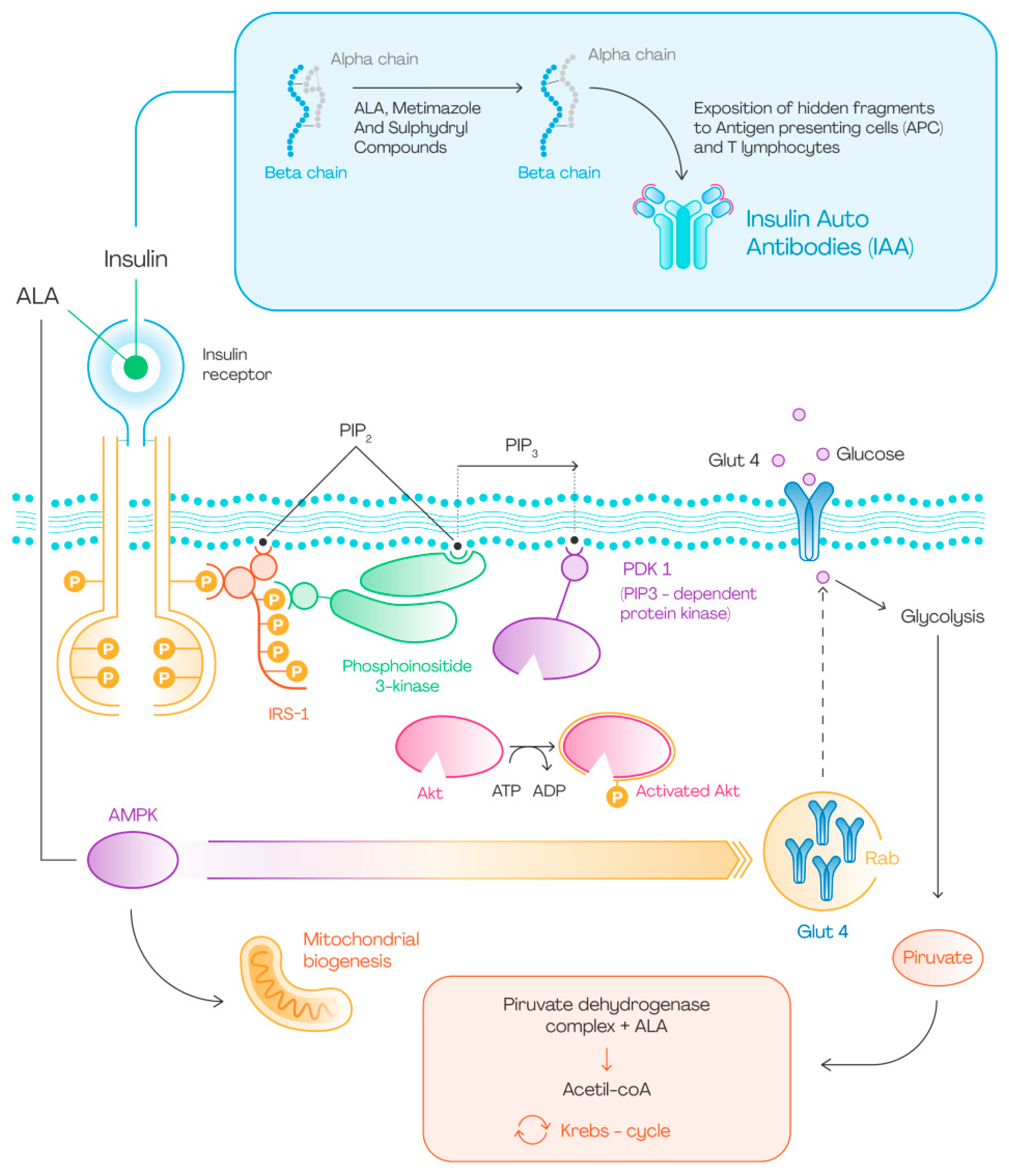
4.1. Effect of ALA on Glucose Uptake
4.2. Effect of ALA on Pancreatic Beta-Cells
4.3. ALA for the Treatment of Diabetes

{kind=link}
| Country/Population | Intervention | Duration | Other Antidiabetic Drugs | Results | |
|---|---|---|---|---|---|
| [6] Ziegler et al., 1997 | Germany/T2D | ALA 800 mg or placebo (oral administration) per day | 4 months | 14 patients on oral antidiabetics while 47 patients on insulin therapy | No HbA1c differences between ALA and placebo |
| [79] Heinisch et al., 2010 | Austria/T2D | ALA 600 mg or placebo (oral administration) per day | 3 weeks | 23 patients on oral antidiabetics (metformin, glitazones and sulphonylureas), 5 patients on insulin and one patient no therapy | No HbA1c differences between ALA and placebo |
| [80] Hegazy et al., 2013 | Egypt/T1D | ALA 600 mg or placebo (oral administration) per day | 4 months | Insulin therapy | No HbA1c differences between ALA and placebo |
| [71] Porasuphatana et al., 2011 | Thailand/T2D | ALA 300 mg, 600 mg, 900 mg, 1200 mg or placebo (oral administration) per day | 6 months | Metformin and/or sulphonylureas | No differences between placebo and other groups, HbA1c significantly decreased in all treatment groups compared to placebo |
| [81] Udupa et al., 2012 | India/T2D | ALA 300 mg or Vitamin E or Omega-3 or placebo (oral administration) per day | 3 months | Metformin plus Glimepiride | No HbA1c differences between ALA and placebo |
| [82] Huang et al., 2013 | China/T2D | ALA 600 mg or placebo (intravenous infusion) per day | 3 months | Short term continuous subcutaneous insulin infusion | No HbA1c differences between ALA and placebo |
| [68] Zhao et al., 2014 | China/T2D complicated by acute cerebral infarction | ALA 600 mg or placebo (intravenous infusion) per day | 1 month | Insulin therapy | ALA significantly decreased HbA1c compared to placebo |
| [70] Mendoza-Núñez et al., 2019 | Mexico/T2D | ALA 600 mg or placebo (oral administration) per day | 6 months | Metformin/glibenclamide | No HbA1c differences between ALA and placebo |
4.4. ALA for Treatment of PCOS and Conditions of Insulin Resistance
5. ALA and Diabetic Neuropathy
6. ALA and Insulin Autoimmune Syndrome
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ALA | alpha-lipoic acid |
| DHLA | dihydro-lipoic acid |
| T1D | type 1 diabetes |
| T2D | type 2 diabetes |
| MS | metabolic syndrome |
| PCOS | polycystic ovary syndrome |
| IAS | insulin autoimmune syndrome |
| IAA | insulin autoantibodies |
| IRS-1 | insulin receptor substrate—1 |
| IR | insulin receptor |
| HSP | heat shock proteins |
| PTDM | post-transplant diabetes mellitus |
| DPN | diabetic polyneuropathy |
| DAN | diabetic autonomic neuropathy |
| NIS | neuropathy impairment score |
| TSS | total symptoms score |
| CAN | cardiovascular autonomic neuropathy |
| FGM | flash glucose monitoring |
References
- Rochette, L.; Ghibu, S.; Richard, C.; Zeller, M.; Cottin, Y.; Vergely, C. Direct and indirect antioxidant properties of alpha-lipoic acid and therapeutic potential. Mol. Nutr. Food Res. 2013, 57, 114–125. [Google Scholar] [CrossRef]
- Konrad, D.; Somwar, R.; Sweeney, G.; Yaworsky, K.; Hayashi, M.; Ramlal, T.; Klip, A. The antihyperglycemic drug alpha-lipoic acid stimulates glucose uptake via both GLUT4 translocation and GLUT4 activation: Potential role of p38 mitogen-activated protein kinase in GLUT4 activation. Diabetes 2001, 50, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Targonsky, E.D.; Dai, F.; Koshkin, V.; Karaman, G.T.; Gyulkhandanyan, A.V.; Zhang, Y.; Chan, C.B.; Wheeler, M.B. α-lipoic acid regulates AMP-activated protein kinase and inhibits insulin secretion from beta cells. Diabetologia 2006, 49, 1587–1598. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghelani, H.; Razmovski-Naumovski, V.; Nammi, S. Chronic treatment of (R)-α-lipoic acid reduces blood glucose and lipid levels in high-fat diet and low-dose streptozotocin-induced metabolic syndrome and type 2 diabetes in Sprague-Dawley rats. Pharmacol. Res. Perspect. 2017, 5, e00306. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.; Li, X.; Qu, Z.C.; Perriott, L.; Whitesell, R.R.; May, J.M. Uptake, recycling, and antioxidant actions of α-lipoic acid in endothelial cells. Free Radic. Biol. Med. 2002, 33, 83–93. [Google Scholar] [CrossRef]
- Ziegler, D.; Hanefeld, M.; Ruhnau, K.J.; Hasche, H.; Lobisch, M.; Schütte, K.; Kerum, G.; Malessa, R. Treatment of symptomatic diabetic polyneuropathy with the antioxidant alpha-lipoic acid: A 7-month multicenter randomized controlled trial (ALADIN III Study). ALADIN III Study Group. Alpha-Lipoic Acid in Diabetic Neuropathy. Diabetes Care 1999, 22, 1296–1301. [Google Scholar] [CrossRef]
- Mezza, T.; Cinti, F.; Cefalo, C.M.A.; Pontecorvi, A.; Kulkarni, R.N.; Giaccari, A. β-Cell Fate in Human Insulin Resistance and Type 2 Diabetes: A Perspective on Islet Plasticity. Diabetes 2019, 68, 1121–1129. [Google Scholar] [CrossRef]
- Mezza, T.; Cefalo, C.M.A.; Cinti, F.; Quero, G.; Pontecorvi, A.; Alfieri, S.; Holst, J.J.; Giaccari, A. Endocrine and Metabolic Insights from Pancreatic Surgery. Trends Endocrinol. Metab. 2020, 31, 760–772. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Shefer, K.; Della Casa, D.; Prati, A.; Napolitano, A.; Manzo, A.; Despini, G.; Simoncini, T. Modulatory effects of alpha-lipoic acid (ALA) administration on insulin sensitivity in obese PCOS patients. J. Endocrinol. Investig. 2018, 41, 583–590. [Google Scholar] [CrossRef]
- Muscogiuri, G.; Policola, C.; Prioletta, A.; Sorice, G.; Mezza, T.; Lassandro, A.; Della Casa, S.; Pontecorvi, A.; Giaccari, A. Low levels of 25(OH)D and insulin-resistance: 2 unrelated features or a cause-effect in PCOS? Clin. Nutr. 2012, 31, 476–480. [Google Scholar] [CrossRef]
- Moffa, S.; Mezza, T.; Cefalo, C.M.A.; Cinti, F.; Impronta, F.; Sorice, G.P.; Santoro, A.; Di Giuseppe, G.; Pontecorvi, A.; Giaccari, A. The Interplay between Immune System and Microbiota in Diabetes. Mediat. Inflamm. 2019, 2019, 9367404. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, Y.; Miyamoto, T.; Kakizawa, T.; Shigematsu, S.; Hashizume, K. Insulin Autoimmune Syndrome possibly caused by alpha lipoic acid. Intern. Med. 2007, 46, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Gullo, D.; Evans, J.L.; Sortino, G.; Goldfine, I.D.; Vigneri, R. Insulin autoimmune syndrome (Hirata Disease) in European Caucasians taking α-lipoic acid. Clin. Endocrinol. 2014, 81, 204–209. [Google Scholar] [CrossRef]
- Bresciani, E.; Bussi, A.; Bazzigaluppi, E.; Balestrieri, G. Insulin autoimmune syndrome induced by α-lipoic acid in a Caucasian woman: Case report. Diabetes Care 2011, 34, e146. [Google Scholar] [CrossRef] [PubMed]
- Moffa, S.; Improta, I.; Rocchetti, S.; Mezza, T.; Giaccari, A. Potential cause-effect relationship between insulin autoimmune syndrome and alpha lipoic acid: Two case reports. Nutrition 2019, 57, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Berkson, B.M. Thioctic acid in treatment of hepatotoxic mushroom (Phalloides) poisoning. N. Engl. J. Med. 1979, 300, 371. [Google Scholar] [CrossRef] [PubMed]
- Amenta, F.; Traini, E.; Tomassoni, D.; Mignini, F. Pharmacokinetics of different formulations of tioctic (alpha-lipoic) acid in healthy volunteers. Clin. Exp. Hypertens. 2008, 30, 767–775. [Google Scholar] [CrossRef]
- Shay, K.P.; Moreau, R.F.; Smith, E.J.; Smith, A.R.; Hagen, T.M. Alpha-lipoic acid as a dietary supplement: Molecular mechanisms and therapeutic potential. Biochim. Biophys. Acta 2009, 1790, 1149–1160. [Google Scholar] [CrossRef]
- Kagan, V.E.; Shvedova, A.; Serbinova, E.; Khan, S.; Swanson, C.; Powell, R.; Packer, L. Dihydrolipoic acid—A universal antioxidant both in the membrane and in the aqueous phase. Reduction of peroxyl, ascorbyl and chromanoxyl radicals. Biochem. Pharmacol. 1992, 44, 1637–1649. [Google Scholar] [CrossRef]
- Morikawa, T.; Yasuno, R.; Wada, H. Do mammalian cells synthesize lipoic acid? Identification of a mouse cDNA encoding a lipoic acid synthase located in mitochondria. FEBS Lett. 2001, 498, 16–21. [Google Scholar] [CrossRef]
- Bilska, A.; Wlodek, L. Lipoic acid—The drug of the future? Pharmacol. Rep. 2005, 57, 570–577. [Google Scholar] [PubMed]
- Biewenga, G.P.; Haenen, G.R.; Bast, A. The pharmacology of the antioxidant lipoic acid. Gen. Pharmacol. 1997, 29, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Mattulat, A.; Baltes, W. Determination of lipoic acid in meat of commercial quality. Z. Lebensm. Unters. Forsch. 1992, 194, 326–329. [Google Scholar] [CrossRef]
- Bast, A.; Haenen, G.R. Lipoic acid: A multifunctional antioxidant. Biofactors 2003, 17, 207–213. [Google Scholar] [CrossRef]
- Palaniappan, A.R.; Dai, A. Mitochondrial ageing and the beneficial role of α-lipoic acid. Neurochem. Res. 2007, 32, 1552–1558. [Google Scholar] [CrossRef]
- Behal, R.H.; Buxton, D.B.; Robertson, J.G.; Olson, M.S. Regulation of the pyruvate dehydrogenase multienzyme complex. Annu. Rev. Nutr. 1993, 13, 497–520. [Google Scholar] [CrossRef]
- Tibullo, D.; Li Volti, G.; Giallongo, C.; Grasso, S.; Tomassoni, D.; Anfuso, C.D.; Lupo, G.; Amenta, F.; Avola, R.; Bramanti, V. Biochemical and clinical relevance of alpha lipoic acid: Antioxidant and anti-inflammatory activity, molecular pathways and therapeutic potential. Inflamm. Res. 2017, 66, 947–959. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.P.; Wells, W.W. α-Lipoic acid dependent regeneration of ascorbic acid from dehydroascorbic acid in rat liver mitochondria. J. Bioenerg. Biomembr. 1996, 28, 77–85. [Google Scholar] [CrossRef]
- Mousavi, S.M.; Shab-Bidar, S.; Kord-Varkaneh, H.; Khorshidi, M.; Djafarian, K. Effect of alpha-lipoic acid supplementation on lipid profile: A systematic review and meta-analysis of controlled clinical trials. Nutrition 2019, 59, 121–130. [Google Scholar] [CrossRef]
- Eason, R.C.; Archer, H.E.; Akhtar, S.; Bailey, C.J. Lipoic acid increases glucose uptake by skeletal muscles of obese-diabetic ob/ob mice. Diabetes Obes. Metab. 2002, 4, 29–35. [Google Scholar] [CrossRef]
- Yaworsky, K.; Somwar, R.; Ramlal, T.; Tritschler, H.J.; Klip, A. Engagement of the insulin-sensitive pathway in the stimulation of glucose transport by alpha-lipoic acid in 3T3-L1 adipocytes. Diabetologia 2000, 43, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Diesel, B.; Kulhanek-Heinze, S.; Holtje, M.; Brandt, B.; Höltje, H.-D.; Vollmar, A.M.; Kiemer, A.K. α-lipoic acid as a directly binding activator of the insulin receptor: Protection from hepatocyte apoptosis. Biochemistry 2007, 46, 2146–2155. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.J.; Moon, H.E.; Moini, H.; Packer, L.; Yoon, D.Y.; Chung, A.S. α-lipoic acid inhibits adipocyte differentiation by regulating pro-adipogenic transcription factors via mitogen-activated protein kinase pathways. J. Biol. Chem. 2003, 278, 34823–34833. [Google Scholar] [CrossRef]
- Moini, H.; Tirosh, O.; Park, Y.C.; Cho, K.J.; Packer, L. R-α-lipoic acid action on cell redox status, the insulin receptor, and glucose uptake in 3T3-L1 adipocytes. Arch. Biochem. Biophys. 2002, 397, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.J.; Song, K.-H.; Koh, E.H.; Won, J.C.; Kim, H.S.; Park, H.-S.; Kim, M.-S.; Kim, S.-W.; Lee, K.-U.; Park, J.-Y. α-lipoic acid increases insulin sensitivity by activating AMPK in skeletal muscle. Biochem. Biophys. Res. Commun. 2005, 332, 885–891. [Google Scholar] [CrossRef]
- Kim, M.-S.; Park, J.-Y.; Namkoong, C.; Jang, P.-G.; Ryu, J.-W.; Song, H.-S.; Yun, J.-Y.; Nam-Goong, I.S.; Ha, J.; Park, I.-S.; et al. Anti-obesity effects of α-lipoic acid mediated by suppression of hypothalamic AMP-activated protein kinase. Nat. Med. 2004, 10, 727–733. [Google Scholar] [CrossRef]
- Bergeron, R.; Ren, J.M.; Cadman, K.S.; Moore, I.K.; Perret, P.; Pypaert, M.; Young, L.H.; Semenkovich, C.F.; Shulman, G.I. Chronic activation of AMP kinase results in NRF-1 activation and mitochondrial biogenesis. Am. J. Physiol. Endocrinol. Metab. 2001, 281, E1340–E1346. [Google Scholar] [CrossRef]
- Zong, H.; Ren, J.M.; Young, L.H.; Pypaert, M.; Mu, J.; Birnbaum, M.J.; Shulman, G.I. AMP kinase is required for mitochondrial biogenesis in skeletal muscle in response to chronic energy deprivation. Proc. Natl. Acad. Sci. USA 2002, 99, 15983–15987. [Google Scholar] [CrossRef]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef]
- Diane, A.; Mahmoud, N.; Bensmail, I.; Khattab, N.; Abunada, H.A.; Dehbi, M. Alpha lipoic acid attenuates ER stress and improves glucose uptake through DNAJB3 cochaperone. Sci. Rep. 2020, 10, 20482. [Google Scholar] [CrossRef]
- El Midaoui, A.; Wu, L.; Wang, R.; de Champlain, J. Modulation of cardiac and aortic peroxisome proliferator-activated receptor-gamma expression by oxidative stress in chronically glucose-fed rats. Am. J. Hypertens. 2006, 19, 407–412. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gupte, A.A.; Bomhoff, G.L.; Morris, J.K.; Gorres, B.K.; Geiger, P.C. Lipoic acid increases heat shock protein expression and inhibits stress kinase activation to improve insulin signaling in skeletal muscle from high-fat-fed rats. J. Appl. Physiol. 2009, 106, 1425–1434. [Google Scholar] [CrossRef] [PubMed]
- Abu-Farha, M.; Cherian, P.; Al-Khairi, I.; Tiss, A.; Khadir, A.; Kavalakatt, S.; Warsame, S.; Dehbi, M.; Behbehani, K.; Abubaker, J. DNAJB3/HSP-40 cochaperone improves insulin signaling and enhances glucose uptake in vitro through JNK repression. Sci. Rep. 2015, 5, 14448. [Google Scholar] [CrossRef] [PubMed]
- Arredouani, A.; Diane, A.; Khattab, N.; Bensmail, I.; Aoude, I.; Chikri, M.; Mohammad, R.; Abou-Samra, A.B.; Dehbi, M. DNAJB3 attenuates metabolic stress and promotes glucose uptake by eliciting Glut4 translocation. Sci. Rep. 2019, 9, 4772. [Google Scholar] [CrossRef] [PubMed]
- Hooper, P.L.; Hooper, P.L. Inflammation, heat shock proteins, and type 2 diabetes. Cell Stress Chaperones 2009, 14, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Koshkin, V.; Dai, F.F.; Robson-Doucette, C.A.; Chan, C.B.; Wheeler, M.B. Limited mitochondrial permeabilization is an early manifestation of palmitate-induced lipotoxicity in pancreatic β-cells. J. Biol. Chem. 2008, 283, 7936–7948. [Google Scholar] [CrossRef]
- Koh, G.; Lee, D.H.; Woo, J.T. 2-Deoxy-D-ribose induces cellular damage by increasing oxidative stress and protein glycation in a pancreatic β-cell line. Metabolism 2010, 59, 325–332. [Google Scholar] [CrossRef]
- Shen, W.; Liu, K.; Tian, C.; Yang, L.; Li, X.; Ren, J.; Packer, L.; Head, E.; Sharman, E.; Liu, J. Protective effects of R-alpha-lipoic acid and acetyl-L-carnitine in MIN6 and isolated rat islet cells chronically exposed to oleic acid. J. Cell. Biochem. 2008, 104, 1232–1243. [Google Scholar] [CrossRef]
- Koh, G.; Yang, E.J.; Kim, M.K.; Lee, S.A.; Lee, D.H. Alpha-lipoic acid treatment reverses 2-deoxy-D-ribose-induced oxidative damage and suppression of insulin expression in pancreatic beta-cells. Biol. Pharm. Bull. 2013, 36, 1570–1576. [Google Scholar] [CrossRef]
- Cummings, B.P.; Stanhope, K.L.; Graham, J.L.; Evans, J.L.; Baskin, D.G.; Griffen, S.C.; Havel, P.J. Dietary fructose accelerates the development of diabetes in UCD-T2DM rats: Amelioration by the antioxidant, α-lipoic acid. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R1343–R1350. [Google Scholar] [CrossRef]
- Hsieh, P.S.; Chan, J.Y.; Shyu, J.F.; Chen, Y.T.; Loh, C.H. Mild portal endotoxaemia induces subacute hepatic inflammation and pancreatic β-cell dysfunction in rats. Eur. J. Clin. Investig. 2008, 38, 640–648. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.F.; Hsieh, C.H.; Hsieh, Y.J.; Chen, Y.T.; Peng, Y.J.; Hsieh, P.S. α-Lipoic acid prevents mild portal endotoxaemia-induced hepatic inflammation and beta cell dysfunction. Eur. J. Clin. Investig. 2012, 42, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Mezza, T.; Muscogiuri, G.; Sorice, G.P.; Clemente, G.; Hu, J.; Pontecorvi, A.; Holst, J.J.; Giaccari, A.; Kulkarni, R.N. Insulin resistance alters islet morphology in nondiabetic humans. Diabetes 2014, 63, 994–1007. [Google Scholar] [CrossRef] [PubMed]
- Mezza, T.; Ferraro, P.M.; Di Giuseppe, G.; Moffa, S.; Cefalo, C.M.; Cinti, F.; Impronta, F.; Capece, U.; Quero, G.; Pontecorvi, A.; et al. Pancreaticoduodenectomy model demonstrates a fundamental role of dysfunctional β cells in predicting diabetes. J. Clin. Investig. 2021, 131, e146788. [Google Scholar] [CrossRef] [PubMed]
- Nobakht-Haghighi, N.; Rahimifard, M.; Baeeri, M.; Rezvanfar, M.A.; Nodeh, S.M.; Haghi-Aminjan, H.; Hamurtekin, E.; Abdollahi, M. Regulation of aging and oxidative stress pathways in aged pancreatic islets using alpha-lipoic acid. Mol. Cell. Biochem. 2018, 449, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Abd-Ellah, H.F.; Abou-Zeid, N.R. Role of alpha-lipoic acid in ameliorating Cyclosporine A-induced pancreatic injury in albino rats: A structural, ultrastructural, and morphometric study. Ultrastruct. Pathol. 2017, 41, 196–208. [Google Scholar] [CrossRef]
- Ghoneim, F.M.; Alrefai, H.; Elsamanoudy, A.Z.; Abo El-Khair, S.M.; Khalaf, H.A. The Protective Role of Prenatal Alpha Lipoic Acid Supplementation against Pancreatic Oxidative Damage in Offspring of Valproic Acid-Treated Rats: Histological and Molecular Study. Biology 2020, 9, 239. [Google Scholar] [CrossRef]
- Schroeder, M.M.; Belloto, R.J., Jr.; Hudson, R.A.; McInerney, M.F. Effects of antioxidants coenzyme Q10 and lipoic acid on interleukin-1β-mediated inhibition of glucose-stimulated insulin release from cultured mouse pancreatic islets. Immunopharmacol. Immunotoxicol. 2005, 27, 109–122. [Google Scholar] [CrossRef] [PubMed]
- Brusco, N.; Sebastiani, G.; Di Giuseppe, G.; Licata, G.; Grieco, G.E.; Fignani, D.; Nigi, L.; Formichi, C.; Aiello, E.; Auddino, S.; et al. Intra-islet insulin synthesis defects are associated with endoplasmic reticulum stress and loss of beta cell identity in human diabetes. Diabetologia 2022. [Google Scholar] [CrossRef]
- Yang, Y.; Wang, W.; Liu, Y.; Guo, T.; Chen, P.; Ma, K.; Zhou, C. α-lipoic acid inhibits high glucose-induced apoptosis in HIT-T15 cells. Dev. Growth Differ. 2012, 54, 557–565. [Google Scholar] [CrossRef]
- Cinti, F.; Mezza, T.; Severi, I.; Suleiman, M.; Cefalo, C.; Sorice, G.; Moffa, S.; Impronta, F.; Quero, G.; Alfieri, S.; et al. Noradrenergic fibers are associated with beta-cell dedifferentiation and impaired beta-cell function in humans. Metabolism 2021, 114, 154414. [Google Scholar] [CrossRef] [PubMed]
- Mezza, T.; Sorice, G.P.; Conte, C.; Sun, V.A.; Cefalo, C.M.A.; Moffa, S.; Pontecorvi, A.; Mari, A.; Kulkarni, R.N.; Giaccari, A. β-Cell Glucose Sensitivity Is Linked to Insulin/Glucagon Bihormonal Cells in Nondiabetic Humans. J. Clin. Endocrinol. Metab. 2016, 101, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Azzam, S.K.; Jang, H.; Choi, M.C.; Alsafar, H.; Lukman, S.; Lee, S. Inhibition of Human Amylin Aggregation and Cellular Toxicity by Lipoic Acid and Ascorbic Acid. Mol. Pharm. 2018, 15, 2098–2106. [Google Scholar] [CrossRef] [PubMed]
- Marzban, L.; Park, K.; Verchere, C.B. Islet amyloid polypeptide and type 2 diabetes. Exp. Gerontol. 2003, 38, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Hirohata, M.; Yamada, M. α-lipoic acid exhibits anti-amyloidogenicity for beta-amyloid fibrils in vitro. Biochem. Biophys. Res. Commun. 2006, 341, 1046–1052. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Ferrannini, E.; Groop, L.; Henry, R.R.; Herman, W.H.; Holst, J.J.; Hu, F.B.; Kahn, C.R.; Raz, I.; Shulman, G.; et al. Type 2 diabetes mellitus. Nat. Rev. Dis. Prim. 2015, 1, 15019. [Google Scholar] [CrossRef]
- Ebada, M.A.; Fayed, N.; Fayed, L.; Alkanj, S.; Abdelkarim, A.; Farwati, H.; Hanafy, A.; Negida, A.; Ebada, M.; Noser, Y. Efficacy of Alpha-lipoic Acid in The Management of Diabetes Mellitus: A Systematic Review and Meta-analysis. Iran. J. Pharm. Res. 2019, 18, 2144–2156. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Hu, F.X. alpha-Lipoic acid treatment of aged type 2 diabetes mellitus complicated with acute cerebral infarction. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 3715–3719. [Google Scholar] [PubMed]
- Akbari, M.; Ostadmohammadi, V.; Lankarani, K.B.; Tabrizi, R.; Kolahdooz, F.; Khatibi, S.R.; Asemi, Z. The effects of alpha-lipoic acid supplementation on glucose control and lipid profiles among patients with metabolic diseases: A systematic review and meta-analysis of randomized controlled trials. Metabolism 2018, 87, 56–69. [Google Scholar] [CrossRef]
- Mendoza-Nunez, V.M.; Garcia-Martinez, B.I.; Rosado-Perez, J.; Santiago-Osorio, E.; Pedraza-Chaverri, J.; Hernandez-Abad, V.J. The Effect of 600 mg Alpha-lipoic Acid Supplementation on Oxidative Stress, Inflammation, and RAGE in Older Adults with Type 2 Diabetes Mellitus. Oxidative Med. Cell. Longev. 2019, 2019, 3276958. [Google Scholar] [CrossRef]
- Porasuphatana, S.; Suddee, S.; Nartnampong, A.; Konsil, J.; Harnwong, B.; Santaweesuk, A. Glycemic and oxidative status of patients with type 2 diabetes mellitus following oral administration of alpha-lipoic acid: A randomized double-blinded placebo-controlled study. Asia Pac. J. Clin. Nutr. 2012, 21, 12–21. [Google Scholar] [PubMed]
- Jacob, S.; Henriksen, E.J.; Tritschler, H.J.; Augustin, H.J.; Dietze, G.J. Improvement of insulin-stimulated glucose-disposal in type 2 diabetes after repeated parenteral administration of thioctic acid. Exp. Clin. Endocrinol. Diabetes 1996, 104, 284–288. [Google Scholar] [CrossRef] [PubMed]
- Jacob, S.; Ruus, P.; Hermann, R.; Tritschler, H.; Maerker, E.; Renn, W.; Augustin, H.; Dietze, G.; Rett, K. Oral administration of RAC-α-lipoic acid modulates insulin sensitivity in patients with type-2 diabetes mellitus: A placebo-controlled pilot trial. Free Radic. Biol. Med. 1999, 27, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kamenova, P. Improvement of insulin sensitivity in patients with type 2 diabetes mellitus after oral administration of alpha-lipoic acid. Hormones 2006, 5, 251–258. [Google Scholar] [CrossRef]
- Bacha, F.; Lee, S.; Gungor, N.; Arslanian, S.A. From pre-diabetes to type 2 diabetes in obese youth: Pathophysiological characteristics along the spectrum of glucose dysregulation. Diabetes Care 2010, 33, 2225–2231. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, L.E.; Chrapowitzky, L.; Rideout, T.C. Metabolic effects of α-lipoic acid supplementation in pre-diabetics: A randomized, placebo-controlled pilot study. Food Funct. 2019, 10, 5732–5738. [Google Scholar] [CrossRef]
- Mandani, M.; Badehnoosh, B.; Jalali-Mashayekhi, F.; Tavakoli-Far, B.; Khosrowbeygi, A. Alpha-lipoic acid supplementation effects on serum values of some oxidative stress biomarkers in women with gestational diabetes. Gynecol. Endocrinol. 2021, 37, 1111–1115. [Google Scholar] [CrossRef]
- Aslfalah, H.; Jamilian, M.; Rafiei, F.; Khosrowbeygi, A. Reduction in maternal serum values of glucose and gamma-glutamyltransferase after supplementation with alpha-lipoic acid in women with gestational diabetes mellitus. J. Obstet. Gynaecol. Res. 2019, 45, 313–317. [Google Scholar] [CrossRef]
- Heinisch, B.B.; Francesconi, M.; Mittermayer, F.; Schaller, G.; Gouya, G.; Wolzt, M.; Pleiner, J. Alpha-lipoic acid improves vascular endothelial function in patients with type 2 diabetes: A placebo-controlled randomized trial. Eur. J. Clin. Investig. 2010, 40, 148–154. [Google Scholar] [CrossRef]
- Hegazy, S.K.; Tolba, O.A.; Mostafa, T.M.; Eid, M.A.; El-Afify, D.R. Alpha-lipoic acid improves subclinical left ventricular dysfunction in asymptomatic patients with type 1 diabetes. Rev. Diabet. Stud. 2013, 10, 58–67. [Google Scholar] [CrossRef]
- Udupa, A.S.; Nahar, P.S.; Shah, S.H.; Kshirsagar, M.J.; Ghongane, B.B. Study of comparative effects of antioxidants on insulin sensitivity in type 2 diabetes mellitus. J. Clin. Diagn. Res. 2012, 6, 1469–1473. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wan, X.; Liu, J.; Deng, W.; Chen, A.; Liu, L.; Liu, J.; Wei, G.; Li, H.; Fang, D.; et al. Short-term continuous subcutaneous insulin infusion combined with insulin sensitizers rosiglitazone, metformin, or antioxidant α-lipoic acid in patients with newly diagnosed type 2 diabetes mellitus. Diabetes Technol. Ther. 2013, 15, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Masharani, U.; Gjerde, C.; Evans, J.L.; Youngren, J.F.; Goldfine, I.D. Effects of controlled-release alpha lipoic acid in lean, nondiabetic patients with polycystic ovary syndrome. J. Diabetes Sci. Technol. 2010, 4, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Giacca, A.; Lewis, G.F. Short-term oral α-lipoic acid does not prevent lipid-induced dysregulation of glucose homeostasis in obese and overweight nondiabetic men. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E736–E741. [Google Scholar] [CrossRef]
- Azziz, R.; Carmina, E.; Dewailly, D.; Diamanti-Kandarakis, E.; Escobar-Morreale, H.F.; Futterweit, W.; Janssen, O.E.; Legro, R.S.; Norman, R.; Taylor, A.E.; et al. The Androgen Excess and PCOS Society criteria for the polycystic ovary syndrome: The complete task force report. Fertil. Steril. 2009, 91, 456–488. [Google Scholar] [CrossRef]
- Azziz, R.; Woods, K.S.; Reyna, R.; Key, T.J.; Knochenhauer, E.S.; Yildiz, B.O. The prevalence and features of the polycystic ovary syndrome in an unselected population. J. Clin. Endocrinl. Metab. 2004, 89, 2745–2749. [Google Scholar] [CrossRef]
- Moghetti, P.; Tosi, F. Insulin resistance and PCOS: Chicken or egg? J. Endocrinol. Investig. 2021, 44, 233–244. [Google Scholar] [CrossRef]
- Solis-Herrera, C.; Triplitt, C.; Cersosimo, E.; DeFronzo, R.A.; Feingold, K.R.; Anawalt, B.; Boyce, A.; Chrousos, G.; de Herder, W.W.; Dhatariya, K.; et al. Pathogenesis of Type 2 Diabetes Mellitus. In Endotext; MDText.com, Inc.: Portland, OR, USA, 2000. [Google Scholar]
- Kakoly, N.S.; Khomami, M.B.; Joham, A.E.; Cooray, S.; Misso, M.L.; Norman, R.; Harrison, C.L.; Ranasinha, S.; Teede, H.; Moran, L.J. Ethnicity, obesity and the prevalence of impaired glucose tolerance and type 2 diabetes in PCOS: A systematic review and meta-regression. Hum. Reprod. Update 2018, 24, 455–467. [Google Scholar] [CrossRef]
- Lewy, V.D.; Danadian, K.; Witchel, S.F.; Arslanian, S. Early metabolic abnormalities in adolescent girls with polycystic ovarian syndrome. J. Pediatr. 2001, 138, 38–44. [Google Scholar] [CrossRef]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef]
- Genazzani, A.D.; Prati, A.; Santagni, S.; Ricchieri, F.; Chierchia, E.; Rattighieri, E.; Campedelli, A.; Simoncini, T.; Artini, P.G. Differential insulin response to myo-inositol administration in obese polycystic ovary syndrome patients. Gynecol. Endocrinol. 2012, 28, 969–973. [Google Scholar] [CrossRef] [PubMed]
- Fruzzetti, F.; Capozzi, A.; Canu, A.; Lello, S. Treatment with d-chiro-inositol and alpha lipoic acid in the management of polycystic ovary syndrome. Gynecol. Endocrinol. 2019, 35, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Fruzzetti, F.; Fidecicchi, T.; Palla, G.; Gambacciani, M. Long-term treatment with α-lipoic acid and myo-inositol positively affects clinical and metabolic features of polycystic ovary syndrome. Gynecol. Endocrinol. 2020, 36, 152–155. [Google Scholar] [CrossRef]
- Cianci, A.; Panella, M.; Fichera, M.; Falduzzi, C.; Bartolo, M.; Caruso, S. d-chiro-Inositol and alpha lipoic acid treatment of metabolic and menses disorders in women with PCOS. Gynecol. Endocrinol. 2015, 31, 483–486. [Google Scholar] [CrossRef]
- De Cicco, S.; Immediata, V.; Romualdi, D.; Policola, C.; Tropea, A.; Di Florio, C.; Tagliaferri, V.; Scarinci, E.; Della Casa, S.; Lanzone, A.; et al. Myoinositol combined with alpha-lipoic acid may improve the clinical and endocrine features of polycystic ovary syndrome through an insulin-independent action. Gynecol. Endocrinol. 2017, 33, 698–701. [Google Scholar] [CrossRef] [PubMed]
- Rago, R.; Marcucci, I.; Leto, G.; Caponecchia, L.; Salacone, P.; Bonanni, P.; Fiori, C.; Sorrenti, G.; Sebastianelli, A. Effect of myo-inositol and alpha-lipoic acid on oocyte quality in polycystic ovary syndrome non-obese women undergoing in vitro fertilization: A pilot study. J. Biol. Regul. Homeost. Agents 2015, 29, 913–923. [Google Scholar] [PubMed]
- Artini, P.G.; Obino, M.E.R.; Micelli, E.; Malacarne, E.; Vacca, C.; Papini, F.; Cela, V. Effect of d-chiro-inositol and alpha-lipoic acid combination on COH outcomes in overweight/obese PCOS women. Gynecol. Endocrinol. 2020, 36, 755–759. [Google Scholar] [CrossRef]
- Cappelli, V.; Di Sabatino, A.; Musacchio, M.C.; De Leo, V. Valutazione di una nuova associazione tra insulino-sensibilizzanti e acido alpha-lipoico in donne obese affette da PCOS [Evaluation of a new association between insulin-sensitizers and alpha-lipoic acid in obese women affected by PCOS]. Minerva Ginecol. 2013, 65, 425–433. [Google Scholar]
- Li, N.; Yan, W.; Hu, X.; Huang, Y.; Wang, F.; Zhang, W.; Wang, Q.; Wang, X.; Sun, K. Effects of oral α-lipoic acid administration on body weight in overweight or obese subjects: A crossover randomized, double-blind, placebo-controlled trial. Clin. Endocrinol. 2017, 86, 680–687. [Google Scholar] [CrossRef]
- Kucukgoncu, S.; Zhou, E.; Lucas, K.B.; Tek, C. Alpha-lipoic acid (ALA) as a supplementation for weight loss: Results from a meta-analysis of randomized controlled trials. Obes. Rev. 2017, 18, 594–601. [Google Scholar] [CrossRef]
- Namazi, N.; Larijani, B.; Azadbakht, L. Alpha-lipoic acid supplement in obesity treatment: A systematic review and meta-analysis of clinical trials. Clin. Nutr. 2018, 37, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Gries, F.A. α-lipoic acid in the treatment of diabetic peripheral and cardiac autonomic neuropathy. Diabetes 1997, 46 (Suppl. 2), S62–S66. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, D.; Nowak, H.; Kempler, P.; Vargha, P.; Low, P.A. Treatment of symptomatic diabetic polyneuropathy with the antioxidant alpha-lipoic acid: A meta-analysis. Diabet. Med. 2004, 21, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, S.; Boulton, A.J.; Dyck, P.J.; Freeman, R.; Horowitz, M.; Kempler, P.; Lauria, G.; Malik, R.A.; Spallone, V.; Vinik, A.; et al. Diabetic neuropathies: Update on definitions, diagnostic criteria, estimation of severity, and treatments. Diabetes Care 2010, 33, 2285–2293. [Google Scholar] [CrossRef]
- D’Amato, C.; Morganti, R.; Greco, C.; Di Gennaro, F.; Cacciotti, L.; Longo, S.; Mataluni, G.; Lauro, D.; Marfia, G.A.; Spallone, V. Diabetic peripheral neuropathic pain is a stronger predictor of depression than other diabetic complications and comorbidities. Diabetes Vasc. Dis. Res. 2016, 13, 418–428. [Google Scholar] [CrossRef]
- Zelman, D.C.; Brandenburg, N.A.; Gore, M. Sleep impairment in patients with painful diabetic peripheral neuropathy. Clin. J. Pain 2006, 22, 681–685. [Google Scholar] [CrossRef]
- Mijnhout, G.S.; Alkhalaf, A.; Kleefstra, N.; Bilo, H.J. Alpha lipoic acid: A new treatment for neuropathic pain in patients with diabetes? Neth. J. Med. 2010, 68, 158–162. [Google Scholar]
- Nguyen, N.; Takemoto, J.K. A Case for Alpha-Lipoic Acid as an Alternative Treatment for Diabetic Polyneuropathy. J. Pharm. Pharm. Sci. 2018, 21, 177s–191s. [Google Scholar] [CrossRef]
- Ibrahimpasic, K. Alpha lipoic acid and glycaemic control in diabetic neuropathies at type 2 diabetes treatment. Med. Arch. 2013, 67, 7–9. [Google Scholar] [CrossRef]
- Nagamatsu, M.; Nickander, K.K.; Schmelzer, J.D.; Raya, A.; Wittrock, D.A.; Tritschler, H.; Low, P.A. Lipoic acid improves nerve blood flow, reduces oxidative stress, and improves distal nerve conduction in experimental diabetic neuropathy. Diabetes Care 1995, 18, 1160–1167. [Google Scholar] [CrossRef]
- Mitsui, Y.; Schmelzer, J.D.; Zollman, P.J.; Mitsui, M.; Tritschler, H.J.; Low, P.A. Alpha-lipoic acid provides neuroprotection from ischemia-reperfusion injury of peripheral nerve. J. Neurol. Sci. 1999, 163, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Han, T.; Bai, J.; Liu, W.; Hu, Y. A systematic review and meta-analysis of alpha-lipoic acid in the treatment of diabetic peripheral neuropathy. Eur. J. Endocrinol. 2012, 167, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Pop-Busui, R.; Boulton, A.J.; Feldman, E.L.; Bril, V.; Freeman, R.; Malik, R.A.; Sosenko, J.M.; Ziegler, D. Diabetic Neuropathy: A Position Statement by the American Diabetes Association. Diabetes Care 2017, 40, 136–154. [Google Scholar] [CrossRef]
- Bril, V.; England, J.; Franklin, G.M.; Backonja, M.; Cohen, J.; Del Toro, D.; Feldman, E.; Iverson, D.J.; Perkins, B.; Russell, J.W.; et al. Evidence-based guideline: Treatment of painful diabetic neuropathy: Report of the American Academy of Neurology, the American Association of Neuromuscular and Electrodiagnostic Medicine, and the American Academy of Physical Medicine and Rehabilitation. PM R 2011, 3, 345–352.e21. [Google Scholar] [CrossRef] [PubMed]
- Spallone, V.; Ziegler, D.; Freeman, R.; Bernardi, L.; Frontoni, S.; Pop-Busui, R.; Stevens, M.; Kempler, P.; Hilsted, J.; Tesfaye, S.; et al. Cardiovascular autonomic neuropathy in diabetes: Clinical impact, assessment, diagnosis, and management. Diabetes Metab. Res. Rev. 2011, 27, 639–653. [Google Scholar] [CrossRef]
- Ziegler, D.; Schatz, H.; Conrad, F.; Gries, F.A.; Ulrich, H.; Reichel, G. Effects of treatment with the antioxidant α-lipoic acid on cardiac autonomic neuropathy in NIDDM patients. A 4-month randomized controlled multicenter trial (DEKAN Study). Deutsche KardialeAutonomeNeuropathie. Diabetes Care 1997, 20, 369–373. [Google Scholar] [CrossRef]
- Haritoglou, C.; Gerss, J.; Hammes, H.P.; Kampik, A.; Ulbig, M.W.; Group, R.S. Alpha-lipoic acid for the prevention of diabetic macular edema. Ophthalmologica 2011, 226, 127–137. [Google Scholar] [CrossRef]
- Vakali, E.; Rigopoulos, D.; Carrillo, A.E.; Flouris, A.D.; Dinas, P.C. Effects of Alpha-lipoic Acid Supplementation on Human Diabetic Nephropathy: A Systematic Review and Meta-analysis. Curr. Diabetes Rev. 2022, 18, e140921196457. [Google Scholar] [CrossRef]
- Uchigata, Y.; Hirata, Y. Insulin autoimmune syndrome (IAS, Hirata disease). Ann. Med. Interne 1999, 150, 245–253. [Google Scholar]
- Yamada, Y.; Kitayama, K.; Oyachi, M.; Higuchi, S.; Kawakita, R.; Kanamori, Y.; Yorifuji, T. Nationwide survey of endogenous hyperinsulinemic hypoglycemia in Japan (2017–2018): Congenital hyperinsulinism, insulinoma, non-insulinoma pancreatogenous hypoglycemia syndrome and insulin autoimmune syndrome (Hirata’s disease). J. Diabetes Investig. 2020, 11, 554–563. [Google Scholar] [CrossRef]
- Ismail, A.A. The insulin autoimmune syndrome (IAS) as a cause of hypoglycaemia: An update on the pathophysiology, biochemical investigations and diagnosis. Clin. Chem. Lab. Med. 2016, 54, 1715–1724. [Google Scholar] [CrossRef] [PubMed]
- Uchigata, Y. The novel agent, alpha lipoic acid, can cause the development of insulin autoimmune syndrome. Intern. Med. 2007, 46, 1321–1322. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Nutrition, Novel Foods and Food Allergens (NDA); Turck, D.; Castenmiller, J.; de Henauw, S.; Hirsch-Ernst, K.I.; Kearney, J.; Knutsen, H.K.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Scientific opinion on the relationship between intake of alpha-lipoic acid (thioctic acid) and the risk of insulin autoimmune syndrome. EFSA J. 2021, 19, e06577. [Google Scholar] [CrossRef] [PubMed]
- Uchigata, Y.; Kuwata, S.; Tokunaga, K.; Omon, Y.; Hirata, Y.; Kuwata, S.; Tokunaga, K.; Miyamoto, M.; Juji, T. Strong association of insulin autoimmune syndrome with HLA-DR4. Lancet 1992, 339, 393–394. [Google Scholar] [CrossRef]
- Gonzalez-Galarza, F.F.; McCabe, A.; Santos, E.; Jones, J.; Takeshita, L.; Ortega-Rivera, N.D.; Del Cid-Pavon, G.M.; Ramsbottom, K.; Ghattaoraya, G.; Alfirevic, A.; et al. Allele frequency net database (AFND) 2020 update: Gold-standard data classification, open access genotype data and new query tools. Nucleic Acids Res. 2020, 48, D783–D788. [Google Scholar] [CrossRef]
- Burden, A.C.; Rosenthal, F.D. Methimazole and insulin autoimmune syndrome. Lancet 1983, 2, 1311. [Google Scholar] [CrossRef]
- Matsushita, S.; Takahashi, K.; Motoki, M.; Komoriya, K.; Ikagawa, S.; Nishimura, Y. Allele specificity of structural requirement for peptides bound to HLA-DRB1*0405 and -DRB1*0406 complexes: Implication for the HLA-associated susceptibility to methimazole-induced insulin autoimmune syndrome. J. Exp. Med. 1994, 180, 873–883. [Google Scholar] [CrossRef]
- Uchigata, Y.; Hirata, Y.; Omori, Y.; Iwamoto, Y.; Tokunaga, K. Worldwide differences in the incidence of insulin autoimmune syndrome (Hirata disease) with respect to the evolution of HLA-DR4 alleles. Hum. Immunol. 2000, 61, 154–157. [Google Scholar] [CrossRef]
- Roep, B.O.; Schipper, R.; Verduyn, W.; Bruining, G.J.; Schreuder, G.M.; de Vries, R.R. HLA-DRB1*0403 is associated with dominant protection against IDDM in the general Dutch population and subjects with high-risk DQA1*0301-DQB1*0302/DQA1*0501-DQB1*0201 genotype. Tissue Antigens 1999, 54, 88–90. [Google Scholar] [CrossRef][Green Version]
- Falorni, A.; Brozzetti, A.; Perniola, R. From Genetic Predisposition to Molecular Mechanisms of Autoimmune Primary Adrenal Insufficiency. Cortisol Excess Insufficiency 2016, 46, 115–132. [Google Scholar] [CrossRef]
- Miyagawa, S.; Higashimine, I.; Iida, T.; Yamashina, Y.; Fukumoto, T.; Shirai, T. HLA-DRB1*04 and DRB1*14 alleles are associated with susceptibility to pemphigus among Japanese. J. Investig. Dermatol. 1997, 109, 615–618. [Google Scholar] [CrossRef] [PubMed]
- Redmon, J.B.; Nuttall, F.Q. Autoimmune hypoglycemia. Endocrinol. Metab. Clin. N. Am. 1999, 28, 603–618. [Google Scholar] [CrossRef]
- Hattori, N.; Duhita, M.R.; Mukai, A.; Matsueda, M.; Shimatsu, A. Development of insulin antibodies and changes in titers over a long-term period in patients with type 2 diabetes. Clin. Chim. Acta 2014, 433, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Cappellani, D.; Sardella, C.; Campopiano, M.C.; Falorni, A.; Marchetti, P.; Macchia, E. Spontaneously remitting insulin autoimmune syndrome ina patient taking alpha-lipoic acid. Endocrinol. Diabetes Metab. Case Rep. 2018, 2018, 1. [Google Scholar] [CrossRef]
- Bae, S.M.; Bae, M.N.; Kim, E.Y.; Kim, I.K.; Seo, M.W.; Shin, J.K.; Cho, S.R.; Jeong, G.H. Recurrent Insulin Autoimmune Syndrome Caused by alpha-Lipoic Acid in Type 2 Diabetes. Endocrinol. Metab. 2013, 28, 326–330. [Google Scholar] [CrossRef]
- Di Giuseppe, G.; Ciccarelli, G.; Cefalo, C.M.; Cinti, F.; Moffa, S.; Impronta, F.; Capece, U.; Pontecorvi, A.; Giaccari, A.; Mezza, T. Prediabetes: How pathophysiology drives potential intervention on a subclinical disease with feared clinical consequences. Minerva Endocrinol. 2021, 46, 272–292. [Google Scholar] [CrossRef]

| [72] Jacob et al., 1996 | Germany/T2D | ALA 100 mg/500 NaCl (intravenous infusion) or placebo | During glucose clamp study | Oral antihyperglycemic therapy | Mean insulin-mediated glucose disposal increased by about 50% in ALA group (from 3.76 mL/kg/min to 5.82 mL/kg/min, p < 0.05), no significant change in control group (from 3.57 mL/kg/min to 3.91 mL/kg/min) |
| [73] Jacob et al., 1999 | Germany/T2D (About 20% of the patients exhibited diabetic complications) | ALA 600 mg, 1200 mg, 1800 mg or placebo (oral administration) per day | 1 month | Oral antihyperglycemic therapy | No insulin-mediated glucose disposal differences between placebo and other groups. Insulin sensitivity significantly increased in all the active groups pooled together (+27%, p < 0.01) compared to placebo |
| [74] Kamenova et al., 2006 | Greece/T2D | ALA 1200 mg (oral administration) per day. A group of normo-tolerant subjects served as control | One month | Metformin (850 mg once daily to three times daily) | Mean insulin-mediated glucose disposal significantly increased in ALA group (from 3.202 to 5.951 mg/kg/min, p < 0.01). and after treatment was not statistically different from control group. |
| [83] Masharani et al., 2010 | USA/PCOS with oligomenorrhoea without hyperandrogenism | Controlled-release ALA 1200 mg per day (No control groups) | 4 months | Oral contraceptives and/or spironolactone | Mean insulin-mediated glucose disposal significantly increased (from 9.7 ± 1.3 mg/min/kg/mU to 11.1 ± 1.7 mg/min/kg/mU after treatment). Two subjects not on oral contraceptives experienced a doubling of periods respect to four months prior to study entry. |
| [84] Xiao et al., 2011 | Canada/Overweight and obese patients without history of diabetes | Patients were treated in random order with oral placebo followed by NaCl infusion (SAL), oral placebo followed by intralipid infusion (IH), oral ALA 1800 mg per day followed by NaCl infusion (ALA) and oral ALA 1800 mg per day followed by intralipid infusion (ALA + IH) | 2 weeks each phase | No medications | Insulin sensitivity index and disposition were 19 and 25% lower in IH and IH + ALA, respectively, vs. SAL (p < 0.05), indicating lipid-inducedinsulin resistance. Insulin sensitivity in ALA was similar to SAL. |
| [84] Xiao et al., 2011 | Canada/Overweight and obese patients without history of diabetes | Patients were treated in random order with: oral placebo followed by NaCl infusion (SAL), oral placebo followed by intralipid infusion (IH), oral ALA 1800 mg per day followed by NaCl infusion (ALA) and oral ALA 1800 mg per day followed by intralipid infusion (ALA + IH) | 2 weeks each phase | No medications | Insulin secretion rate was similar between treatment groups |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Capece, U.; Moffa, S.; Improta, I.; Di Giuseppe, G.; Nista, E.C.; Cefalo, C.M.A.; Cinti, F.; Pontecorvi, A.; Gasbarrini, A.; Giaccari, A.; et al. Alpha-Lipoic Acid and Glucose Metabolism: A Comprehensive Update on Biochemical and Therapeutic Features. Nutrients 2023, 15, 18. https://doi.org/10.3390/nu15010018
Capece U, Moffa S, Improta I, Di Giuseppe G, Nista EC, Cefalo CMA, Cinti F, Pontecorvi A, Gasbarrini A, Giaccari A, et al. Alpha-Lipoic Acid and Glucose Metabolism: A Comprehensive Update on Biochemical and Therapeutic Features. Nutrients. 2023; 15(1):18. https://doi.org/10.3390/nu15010018
Chicago/Turabian StyleCapece, Umberto, Simona Moffa, Ilaria Improta, Gianfranco Di Giuseppe, Enrico Celestino Nista, Chiara M. A. Cefalo, Francesca Cinti, Alfredo Pontecorvi, Antonio Gasbarrini, Andrea Giaccari, and et al. 2023. "Alpha-Lipoic Acid and Glucose Metabolism: A Comprehensive Update on Biochemical and Therapeutic Features" Nutrients 15, no. 1: 18. https://doi.org/10.3390/nu15010018
APA StyleCapece, U., Moffa, S., Improta, I., Di Giuseppe, G., Nista, E. C., Cefalo, C. M. A., Cinti, F., Pontecorvi, A., Gasbarrini, A., Giaccari, A., & Mezza, T. (2023). Alpha-Lipoic Acid and Glucose Metabolism: A Comprehensive Update on Biochemical and Therapeutic Features. Nutrients, 15(1), 18. https://doi.org/10.3390/nu15010018