The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study)

 , , and
, , and 
Abstract
:1. Introduction
2. Methods
2.1. Ethical Considerations and Registration
2.2. Participants, Randomization, and Blinding
2.3. Intervention Products
2.4. Clinical Examination and Sample Collection
2.5. SCFA in Plasma
2.6. Microbiota Analysis
2.7. Statistical Analysis
3. Results
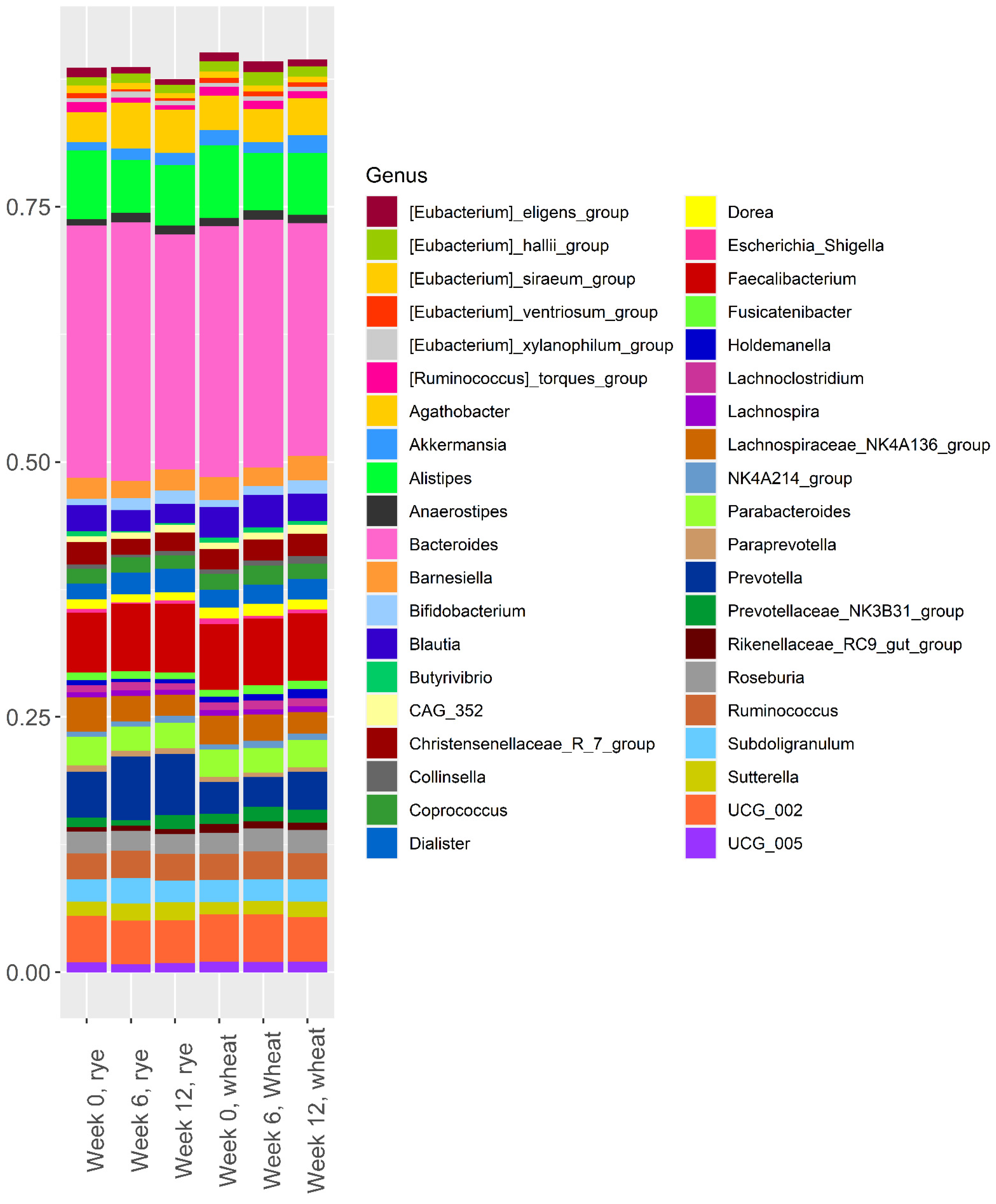
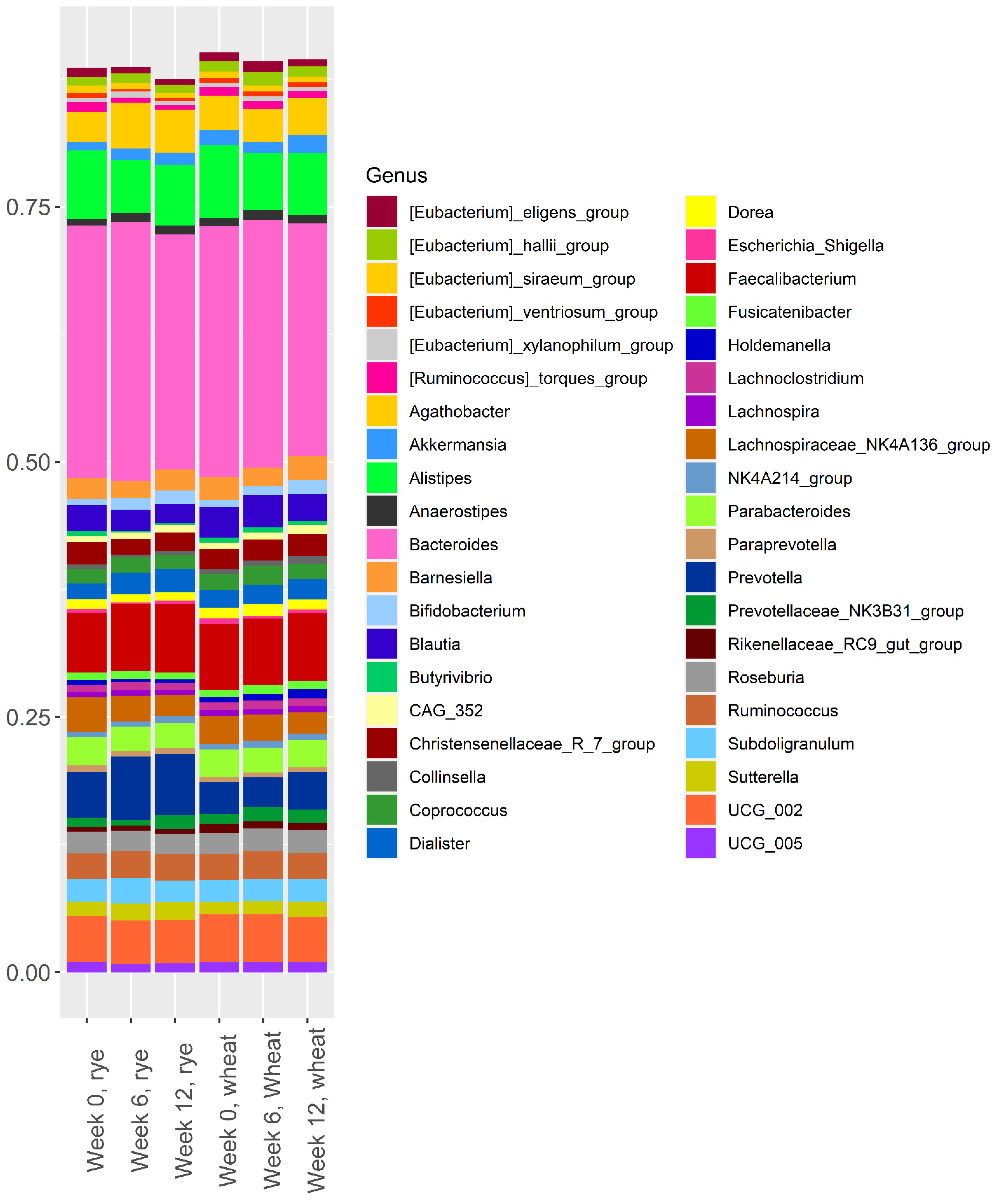
3.1. Effect of the Intervention of Microbiota Composition
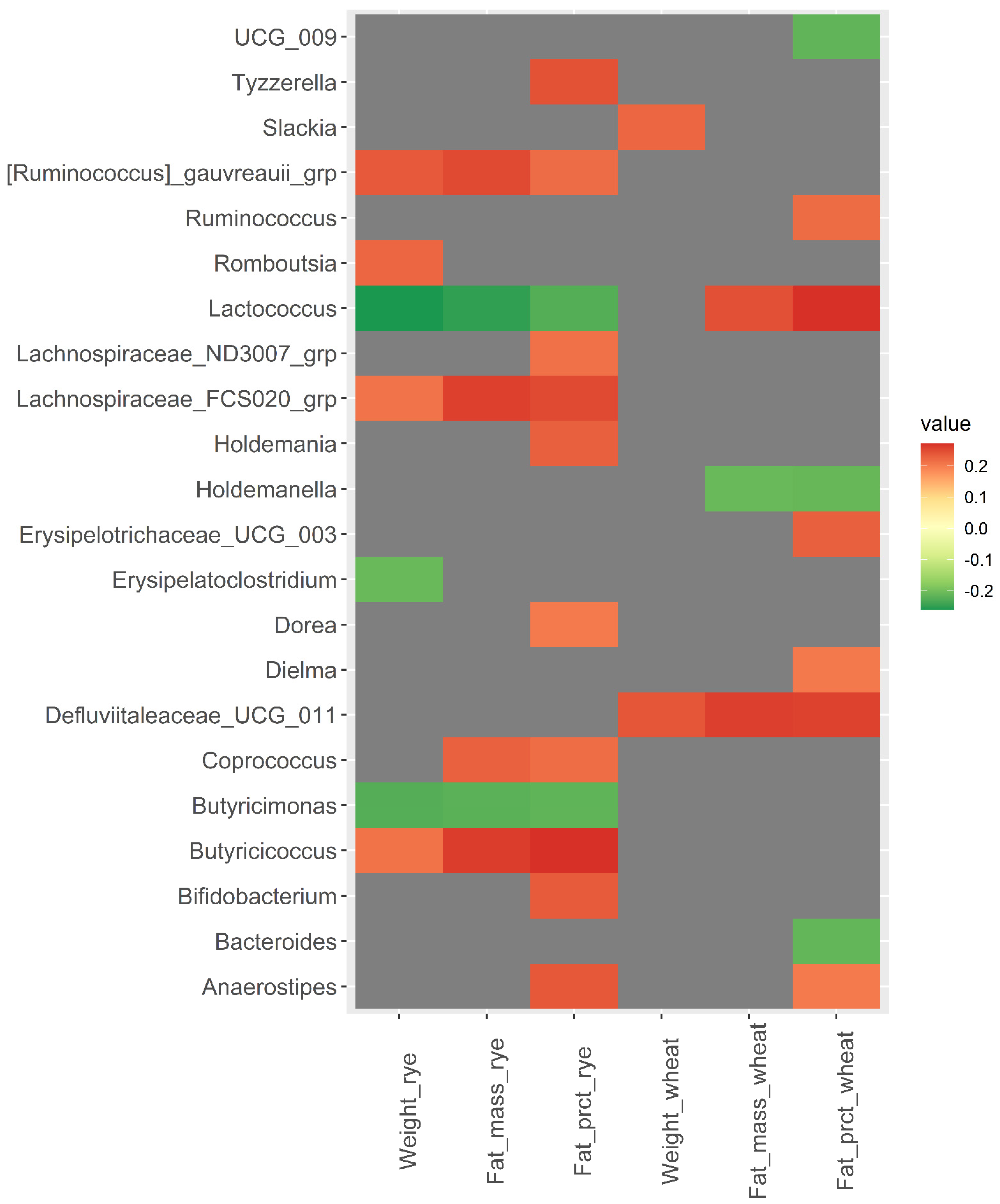
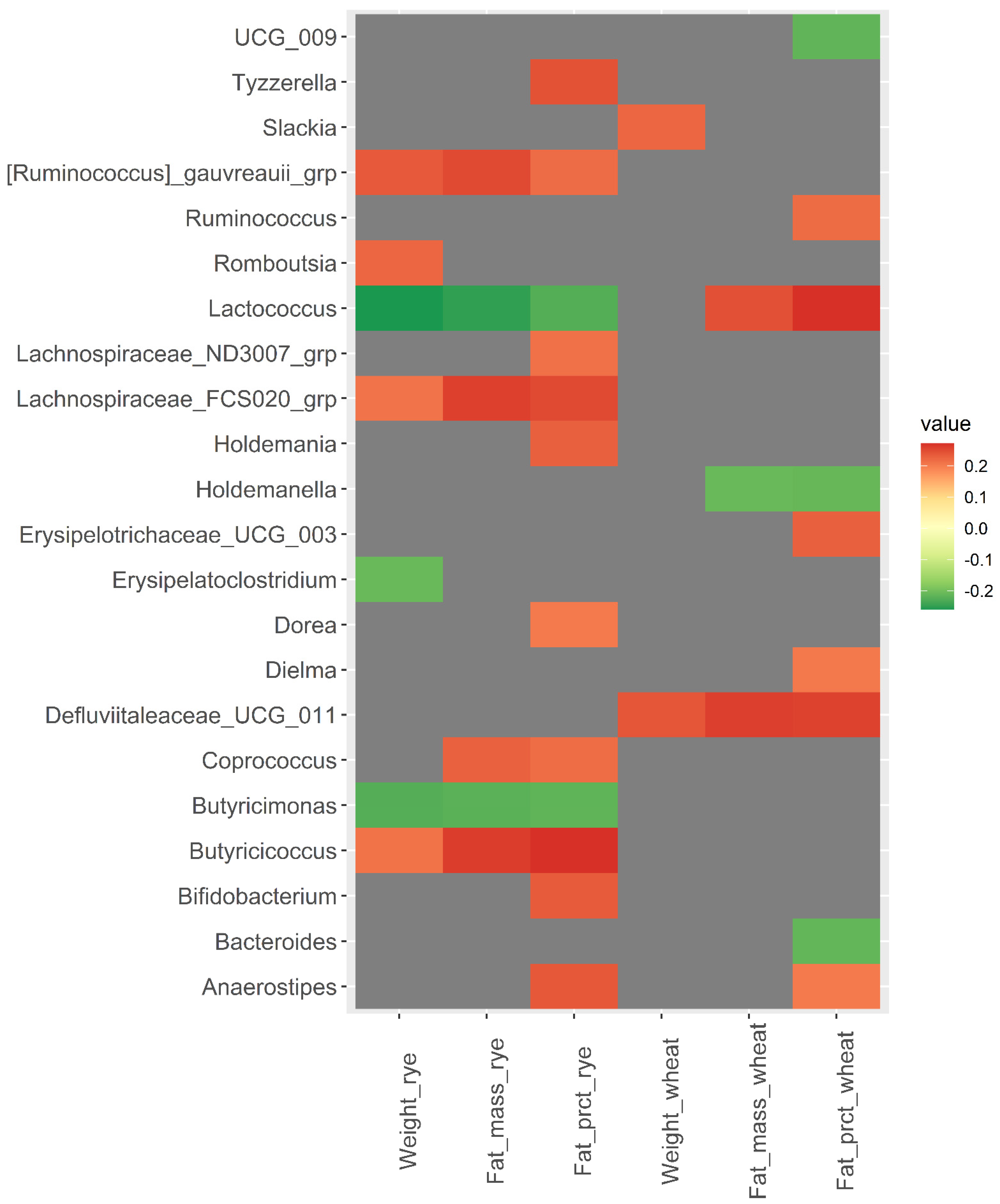
3.2. Baseline Microbiota and Change in Body Weight and Body Fat over 12 Weeks
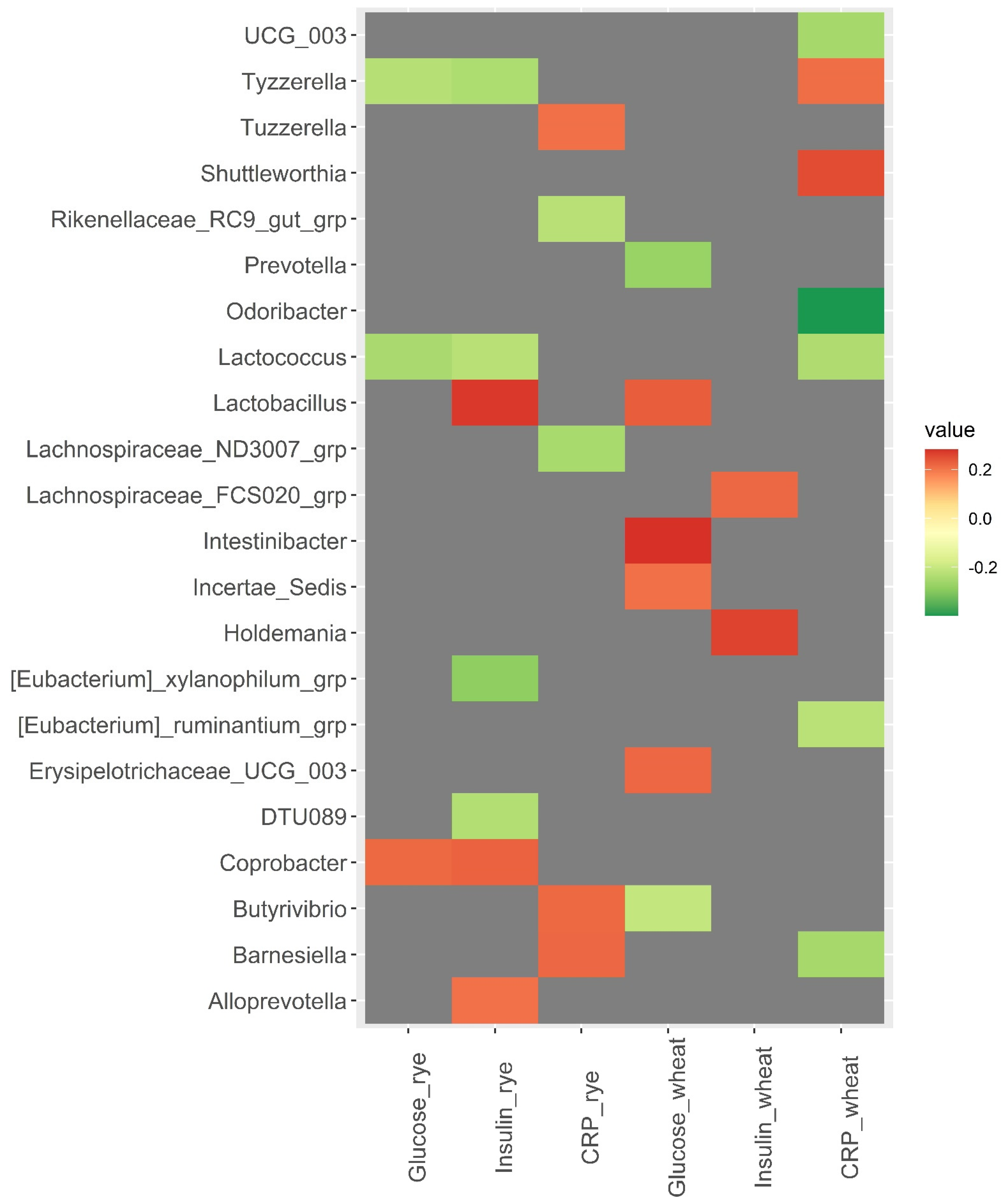
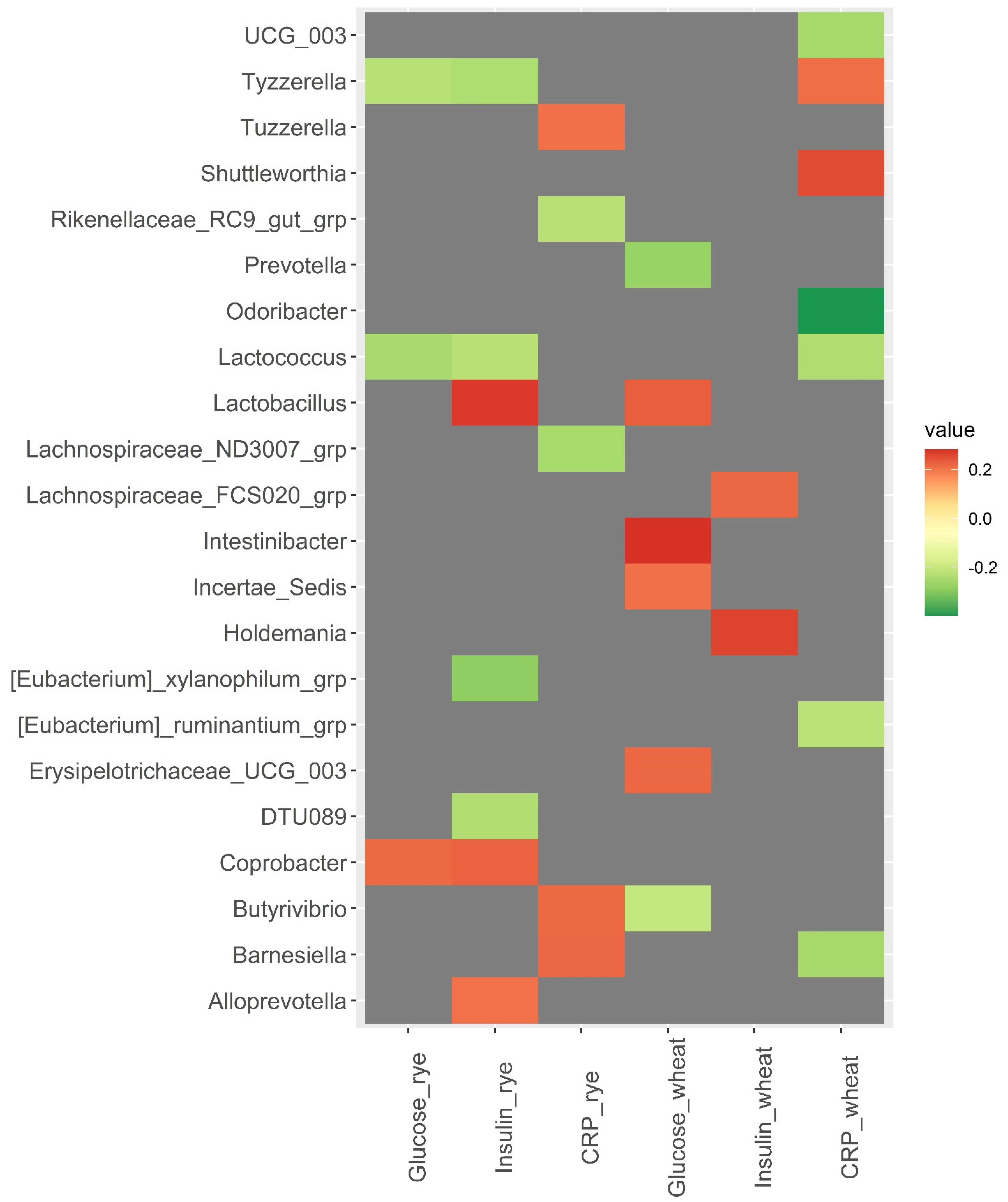
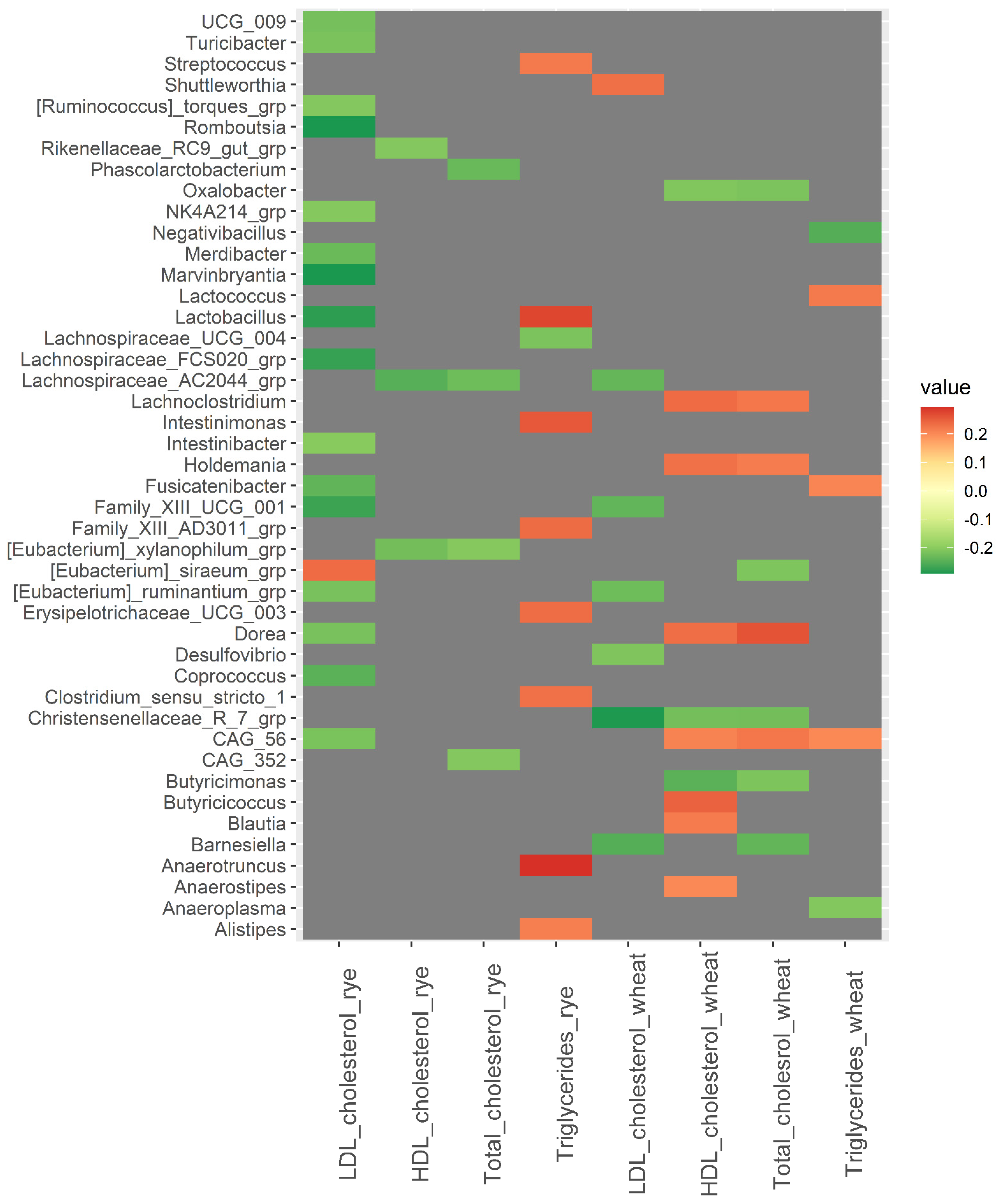
3.3. Baseline Microbiota and Changes in Metabolic Risk Markers over 12 Weeks
3.4. SCFAs in Plasma
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hollænder, P.L.B.; Ross, A.B.; Kristensen, M. Whole-Grain and Blood Lipid Changes in Apparently Healthy Adults: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Am. J. Clin. Nutr. 2015, 102, 556–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wan, Q.; Feng, J.; Du, L.; Li, K.; Zhou, Y. Whole grain diet reduces systemic inflammation: A Meta-Analysis of 9 Randomized Trials. Medicine 2018, 97, e12995. [Google Scholar] [CrossRef] [PubMed]
- Pol, K.; Christensen, R.; Bartels, E.M.; Raben, A.; Tetens, I.; Kristensen, M. Whole grain and body weight changes in apparently healthy adults: A systematic review and meta-analysis of randomized controlled studies. Am. J. Clin. Nutr. 2013, 98, 872–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, E.Q.; Chacko, S.A.; Chou, E.L.; Kugizaki, M.; Liu, S. Greater Whole-Grain Intake Is Associated with Lower Risk of Type 2 Diabetes, Cardiovascular Disease, and Weight Gain. J. Nutr. 2012, 142, 1304–1313. [Google Scholar] [CrossRef]
- Seal, C.J.; Brownlee, I.A. Whole Grains and Health, Evidence from Observational and Intervention Studies. Cereal Chem. 2010, 87, 167–174. [Google Scholar] [CrossRef]
- Gong, L.; Cao, W.; Chi, H.; Wang, J.; Zhang, H.; Liu, J.; Sun, B. Whole Cereal Grains and Potential Health Effects: Involvement of the Gut Microbiota; Elsevier Ltd.: Amsterdam, The Netherlands, 2018; Volume 103, pp. 84–102. [Google Scholar]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [Green Version]
- Sonnenburg, J.L.; Bäckhed, F. Diet–microbiota interactions as moderators of human metabolism. Nature 2016, 535, 56–64. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Liang, Q.; Balakrishnan, B.; Belobrajdic, D.P.; Feng, Q.-J.; Zhang, W. Role of Dietary Nutrients in the Modulation of Gut Microbiota: A Narrative Review. Nutrients 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Poutanen, K.S.; Kårlund, A.O.; Gómez-Gallego, C.; Johansson, D.P.; Scheers, N.M.; Marklinder, I.M.; Eriksen, A.K.; Silventoinen, P.C.; Nordlund, E.; Sozer, N.; et al. Grains—A major source of sustainable protein for health. Nutr. Rev. 2021. [Google Scholar] [CrossRef]
- Vuholm, S.; Nielsen, D.S.; Iversen, K.N.; Suhr, J.; Westermann, P.; Krych, L.; Andersen, J.R.; Kristensen, M. Whole-Grain Rye and Wheat Affect Some Markers of Gut Health without Altering the Fecal Microbiota in Healthy Overweight Adults: A 6-Week Randomized Trial. J. Nutr. 2017, 147, 2067–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Trijp, M.P.H.; Schutte, S.; Esser, D.; Wopereis, S.; Hoevenaars, F.P.M.; Hooiveld, G.J.E.J.; Afman, L.A. Minor Changes in the Composition and Function of the Gut Microbiota During a 12-Week Whole Grain Wheat or Refined Wheat Intervention Correlate with Liver Fat in Overweight and Obese Adults. J. Nutr. 2021, 151, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Vogt, J.K.; Kristensen, M.; Hansen, L.B.S.; Ibrügger, S.; Mærkedahl, R.B.; Bahl, M.I.; Lind, M.V.; Nielsen, R.L.; Frøkiær, H.; et al. Whole grain-rich diet reduces body weight and systemic low-grade inflammation without inducing major changes of the gut microbiome: A randomised cross-over trial. Gut 2019, 68, 83–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faith, J.J.; Guruge, J.L.; Charbonneau, M.; Subramanian, S.; Seedorf, H.; Goodman, A.L.; Clemente, J.C.; Knight, R.; Heath, A.C.; Leibel, R.L.; et al. The Long-Term Stability of the Human Gut Microbiota. Science 2013, 341, 1237439. [Google Scholar] [CrossRef] [Green Version]
- Fragiadakis, G.K.; Wastyk, H.C.; Robinson, J.L.; Sonnenburg, E.D.; Sonnenburg, J.L.; Gardner, C.D. Long-term dietary intervention reveals resilience of the gut microbiota despite changes in diet and weight. Am. J. Clin. Nutr. 2020, 111, 1127–1136. [Google Scholar] [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [Green Version]
- Jefferson, A.; Adolphus, K. The Effects of Intact Cereal Grain Fibers, Including Wheat Bran on the Gut Microbiota Composition of Healthy Adults: A Systematic Review. Front. Nutr. 2019, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Tangestani, H.; Emamat, H.; Ghalandari, H.; Shab-Bidar, S. Whole Grains, Dietary Fibers and the Human Gut Microbiota: A Systematic Review of Existing Literature. Recent Pat. Food Nutr. Agric. 2020, 11, 235–248. [Google Scholar] [CrossRef]
- Hughes, R.L.; Marco, M.L.; Hughes, J.P.; Keim, N.L.; Kable, M.E. The Role of the Gut Microbiome in Predicting Response to Diet and the Development of Precision Nutrition Models—Part I: Overview of Current Methods; Oxford University Press: Oxford, UK, 2019; Volume 10, pp. 953–978. [Google Scholar]
- Hughes, R.L.; Kable, M.E.; Marco, M.; Keim, N.L. The Role of the Gut Microbiome in Predicting Response to Diet and the Development of Precision Nutrition Models. Part II: Results; Oxford University Press: Oxford, UK, 2019; Volume 10, pp. 979–998. [Google Scholar]
- Christensen, L.; Vuholm, S.; Roager, H.M.; Nielsen, D.S.; Krych, L.; Kristensen, M.; Astrup, A.; Hjorth, M. Prevotella Abundance Predicts Weight Loss Success in Healthy, Overweight Adults Consuming a Whole-Grain Diet Ad Libitum: A Post Hoc Analysis of a 6-Wk Randomized Controlled Trial. J. Nutr. 2019, 149, 2174–2181. [Google Scholar] [CrossRef]
- Hjorth, M.F.; Roager, H.M.; Larsen, T.M.; Poulsen, S.K.; Licht, T.R.; Bahl, M.I.; Zohar, Y.; Astrup, A. Pre-treatment microbial Prevotella-to-Bacteroides ratio, determines body fat loss success during a 6-month randomized controlled diet intervention. Int. J. Obes. 2018, 42, 580–583, Erratum in Int. J. Obes. 2018, 42, 284. [Google Scholar] [CrossRef] [Green Version]
- Hjorth, M.F.; Blædel, T.; Bendtsen, L.Q.; Lorenzen, J.K.; Holm, J.B.; Kiilerich, P.; Roager, H.M.; Kristiansen, K.; Larsen, L.H.; Astrup, A. Prevotella-to-Bacteroides ratio predicts body weight and fat loss success on 24-week diets varying in macronutrient composition and dietary fiber: Results from a post-hoc analysis. Int. J. Obes. 2019, 43, 149–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hjorth, M.F.; Christensen, L.; Larsen, T.M.; Roager, H.M.; Krych, L.; Kot, W.; Nielsen, D.S.; Ritz, C.; Astrup, A. Pretreatment Prevotella-to-Bacteroides ratio and salivary amylase gene copy number as prognostic markers for dietary weight loss. Am. J. Clin. Nutr. 2020, 111, 1079–1086. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, A.K.; Brunius, C.; Mazidi, M.; Hellström, P.M.; Risérus, U.; Iversen, K.N.; Fristedt, R.; Sun, L.; Huang, Y.; Nørskov, N.P.; et al. Effects of whole-grain wheat, rye, and lignan supplementation on cardiometabolic risk factors in men with metabolic syndrome: A randomized crossover trial. Am. J. Clin. Nutr. 2020, 111, 864–876. [Google Scholar] [CrossRef] [PubMed]
- Mills, S.; Stanton, C.; Lane, J.A.; Smith, G.J.; Ross, R.P. Precision Nutrition and the Microbiome, Part I: Current State of the Science. Nutrients 2019, 11, 923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mills, S.; Lane, J.A.; Smith, G.J.; Grimaldi, K.A.; Ross, R.P.; Stanton, C. Precision Nutrition and the Microbiome Part II: Potential Opportunities and Pathways to Commercialisation. Nutrients 2019, 11, 923. [Google Scholar] [CrossRef] [Green Version]
- Iversen, K.N.; Landberg, R. Whole Grains, Gut Microbiota, and Health—Time to Get Personal? J. Nutr. 2021, 151, 459–461. [Google Scholar] [CrossRef]
- Iversen, K.N.; Carlsson, F.; Andersson, A.; Michaëlsson, K.; Langton, M.; Risérus, U.; Hellström, P.M.; Landberg, R. A hypocaloric diet rich in high fiber rye foods causes greater reduction in body weight and body fat than a diet rich in refined wheat: A parallel randomized controlled trial in adults with overweight and obesity (the RyeWeight study). Clin. Nutr. ESPEN 2021, 45, 155–169. [Google Scholar] [CrossRef]
- Ross, A.B.; Svelander, C.; Savolainen, O.I.; Lind, M.V.; Kirwan, J.P.; Breton, I.; Godin, J.-P.; Sandberg, A.-S. A high-throughput method for liquid chromatography–tandem mass spectrometry determination of plasma alkylresorcinols, biomarkers of whole grain wheat and rye intake. Anal. Biochem. 2016, 499, 1–7. [Google Scholar] [CrossRef]
- Landberg, R.; Kamal-Eldin, A.; Andersson, A.; Vessby, B.; Åman, P. Alkylresorcinols as biomarkers of whole-grain wheat and rye intake: Plasma concentration and intake estimated from dietary records. Am. J. Clin. Nutr. 2008, 87, 832–838. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Lin, K.; Sequeira, C.; Borchers, C.H. An isotope-labeled chemical derivatization method for the quantitation of short-chain fatty acids in human feces by liquid chromatography–tandem mass spectrometry. Anal. Chim. Acta 2015, 854, 86–94. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Velikonja, A.; Lipoglavšek, L.; Zorec, M.; Orel, R.; Avguštin, G. Alterations in gut microbiota composition and metabolic parameters after dietary intervention with barley beta glucans in patients with high risk for metabolic syndrome development. Anaerobe 2019, 55, 67–77. [Google Scholar] [CrossRef]
- Che, L.; Hu, Q.; Wang, R.; Zhang, D.; Liu, C.; Zhang, Y.; Xin, G.; Fang, Z.; Lin, Y.; Xu, S.; et al. Inter-correlated gut microbiota and SCFAs changes upon antibiotics exposure links with rapid body-mass gain in weaned piglet model. J. Nutr. Biochem. 2019, 74, 108246. [Google Scholar] [CrossRef]
- Rosero, J.A.; Killer, J.; Sechovcová, H.; Mrazek, J.; Benada, O.; Fliegerová, K.; Havlik, J.; Kopečný, J. Reclassification of Eubacterium rectale (Hauduroy et al. 1937) Prévot 1938 in a new genus Agathobacter gen. nov. as Agathobacter rectalis comb. nov., and description of Agathobacter ruminis sp. nov., isolated from the rumen contents of sheep and cows. Int. J. Syst. Evol. Microbiol. 2016, 66, 768–773. [Google Scholar] [CrossRef]
- Lin, H.V.; Frassetto, A.; Kowalik, E.J., Jr.; Nawrocki, A.R.; Lu, M.M.; Kosinski, J.R.; Hubert, J.A.; Szeto, D.; Yao, X.; Forrest, G.; et al. Butyrate and Propionate Protect against Diet-Induced Obesity and Regulate Gut Hormones via Free Fatty Acid Receptor 3-Independent Mechanisms. PLoS ONE 2012, 7, e35240. [Google Scholar] [CrossRef]
- Li, Z.; Yi, C.-X.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; van den Heuvel, J.K.; Meijer, O.C.; Berbée, J.F.P.; et al. Butyrate reduces appetite and activates brown adipose tissue via the gut-brain neural circuit. Gut 2018, 67, 1269–1279. [Google Scholar] [CrossRef] [Green Version]
- Yadav, H.; Lee, J.-H.; Lloyd, J.; Walter, P.; Rane, S.G. Beneficial Metabolic Effects of a Probiotic via Butyrate-induced GLP-1 Hormone Secretion. J. Biol. Chem. 2013, 288, 25088–25097. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Liu, C.; Jiang, Q.; Yin, Y. Butyrate in Energy Metabolism: There Is Still More to Learn. Trends Endocrinol. Metab. 2021, 32, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, J.J.D.R.; Su, W.; Rahat-Rozenbloom, S.; Wolever, T.M.S.; Comelli, E. Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 2014, 4, e121. [Google Scholar] [CrossRef] [PubMed]
- Schwiertz, A.; Taras, D.; Schäfer, K.; Beijer, S.; Bos, N.A.; Donus, C.; Hardt, P.D. Microbiota and SCFA in Lean and Overweight Healthy Subjects. Obesity 2010, 18, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Hernández, M.A.G.; Goossens, G.H.; Reijnders, D.; Holst, J.J.; Jocken, J.W.E.; Van Eijk, H.; Canfora, E.E.; Blaak, E.E. Circulating but not faecal short-chain fatty acids are related to insulin sensitivity, lipolysis and GLP-1 concentrations in humans. Sci. Rep. 2019, 9, 12515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajilić-Stojanović, M.; De Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Hoffman, K.L.; Chen, J.-S.; Shivappa, N.; Sood, A.; Browman, G.J.; Dirba, D.D.; Hanash, S.; Wei, P.; Hebert, J.R.; et al. Dietary inflammatory potential in relation to the gut microbiome: Results from a cross-sectional study. Br. J. Nutr. 2020, 124, 931–942. [Google Scholar] [CrossRef]
- Cani, P. The gut microbiota manages host metabolism. Nat. Rev. Endocrinol. 2014, 10, 74–76. [Google Scholar] [CrossRef]
- Fukui, H. Increased Intestinal Permeability and Decreased Barrier Function: Does It Really Influence the Risk of Inflammation? Inflamm. Intest. Dis. 2016, 1, 135–145. [Google Scholar] [CrossRef]
- Cani, P.D.; Osto, M.; Geurts, L.; Everard, A. Involvement of gut microbiota in the development of low-grade inflammation and type 2 diabetes associated with obesity. Gut Microbes 2012, 3, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Van den Munckhof, I.C.L.; Kurilshikov, A.; Ter Horst, R.; Riksen, N.P.; Joosten, L.A.B.; Zhernakova, A.; Fu, J.; Keating, S.T.; Netea, M.G.; De Graaf, J.; et al. Role of gut microbiota in chronic low-grade inflammation as potential driver for atherosclerotic cardiovascular disease: A systematic review of human studies. Obes. Rev. 2018, 19, 1719–1734. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, K.E.B.; Lærke, H.N.; Hedemann, M.S.; Nielsen, T.S.; Ingerslev, A.K.; Nielsen, D.S.G.; Theil, P.K.; Purup, S.; Hald, S.; Schioldan, A.G.; et al. Impact of Diet-Modulated Butyrate Production on Intestinal Barrier Function and Inflammation. Nutrients 2018, 10, 1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, G.L.; Rifai, N.; Tracy, R.P.; Roberts, W.L.; Alexander, R.W.; Biasucci, L.M.; Catravas, J.D.; Cole, T.G.; Cooper, G.R.; Khan, B.V.; et al. CDC/AHA Workshop on Markers of Inflammation and Cardiovascular Disease: Application to Clinical and Public Health Practice: Report from the Laboratory Science Discussion Group. Circulation 2004, 110, e545–e549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pearson, T.A.; Mensah, G.A.; Alexander, R.W.; Anderson, J.L.; Cannon, R.O., III; Criqui, M.; Fadl, Y.Y.; Fortmann, S.P.; Hong, Y.; Myers, G.L.; et al. Markers of Inflammation and Cardiovascular Disease: Application to Clinical and Public Health Practice: A Statement for Healthcare Professionals from the Centers for Disease Control and Prevention and the American Heart Association. Circulation 2003, 107, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Yang, J.; Martínez, I.; Walter, J.; Keshavarzian, A.; Rose, D.J. In vitro characterization of the impact of selected dietary fibers on fecal microbiota composition and short chain fatty acid production. Anaerobe 2013, 23, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Kedenko, L.; Antonielli, L.; Gemeier, C.; Leitner, M.; Kautzky-Willer, A.; Paulweber, B.; Hackl, E. Gut microbiota dysbiosis associated with glucose metabolism disorders and the metabolic syndrome in older adults. Benef. Microbes 2017, 8, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Laatikainen, R.; Jalanka, J.; Loponen, J.; Hongisto, S.-M.; Hillilä, M.; Koskenpato, J.; Korpela, R.; Salonen, A. Randomised clinical trial: Effect of low-FODMAP rye bread versus regular rye bread on the intestinal microbiota of irritable bowel syndrome patients: Association with individual symptom variation. BMC Nutr. 2019, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Collins, S.M. A role for the gut microbiota in IBS. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 497–505. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rye Group (n = 108) | Wheat Group (n = 99) | |
|---|---|---|
| Sex (n, females/males) | 61/47 | 62/37 |
| Age (years) | 56.8 ± 9.4 | 57.3 ± 9.6 |
| Body weight (kg) | 88.8 ± 12.8 | 89.1 ± 12.3 |
| BMI (kg/m2) | 29.8 ± 2.5 | 30.3 ± 2.5 |
| Body fat mass (kg) | 34.0 ± 6.5 | 35.8 ± 7.2 |
| Body fat percentage | 38.9 ± 6.8 | 40.7 ± 7.0 |
| CRP (mg/L) | 1.5 (0.7; 3.0) | 1.5 (0.9; 2.4) |
| Total cholesterol (mmol/L) | 4.7 ± 0.9 | 4.8 ± 1.0 |
| LDL cholesterol (mmol/L) | 3.1 ± 0.8 | 3.1 ± 0.8 |
| HDL cholesterol (mmol/L) | 1.4 ± 0.3 | 1.4 ± 0.3 |
| Triglycerides (mmol/L) | 1.0 (0.9; 1.3) | 1.1 (0.9; 1.3) |
| Glucose (mmol/L) | 5.5 ± 0.5 | 5.6 ± 0.6 |
| Insulin (mIU/L) | 8.7 (6.6; 12.3) | 10.4 (7.3; 13.2) |
| Raw p-Value/FDR Corrected p-Value ** | ||||||
|---|---|---|---|---|---|---|
| Week 0 * | Week 6 * | Week 12 * | Week 6 | Week 12 | ||
| Agathobacter | Wheat | 2.70 (2.36;3.10) | 2.79 (2.49;3.14) | 3.07 (2.72;3.46) | <0.001/0.010 | 0.033/0.985 |
| Rye | 2.32 (2.04;2.65) | 3.66 (3.28;4.10) | 3.46 (3.08;3.88) | |||
| [Ruminococcus] torques group | Wheat | 0.69 (0.61;0.79) | 0.62 (0.55;0.71) | 0.60 (0.54;0.68) | <0.001/0.010 | <0.001/0.010 |
| Rye | 0.73 (0.65;0.83) | 0.42 (0.38;0.48) | 0.38 (0.34;0.43) | |||
| [Eubacterium] ventriosum group | Wheat | 0.37 (0.32;0.43) | 0.30 (0.26;0.35) | 0.27 (0.24;0.31) | <0.001/0.010 | <0.001/0.010 |
| Rye | 0.34 (0.30;0.40) | 0.20 (0.17;0.23) | 0.18 (0.16;0.20) | |||
| Anaerotruncus | Wheat | 0.009 (0.007;0.012) | 0.008 (0.006;0.011) | 0.006 (0.005;0.008) | <0.001/0.010 | 0.004/0.367 |
| Rye | 0.006 (0.005;0.008) | 0.003 (0.003;0.004) | 0.003 (0.003;0.004) | |||
| Anaerofilum | Wheat | 0.008 (0.006;0.011) | 0.006 (0.005;0.008) | 0.007 (0.005;0.008) | 0.006/0.586 | <0.001/0.032 |
| Rye | 0.008 (0.006;0.010) | 0.004 (0.003;0.005) | 0.004 (0.003;0.005) | |||
| UCG-003 | Wheat | 0.27 (0.24;0.32) | 0.29 (0.25;0.33) | 0.37 (0.32;0.42) | <0.001/0.010 | 0.010/0.874 |
| Rye | 0.27 (0.23;0.31) | 0.40 (0.35;0.46) | 0.45 (0.40;0.51) | |||
| Holdemania | Wheat | 0.006 (0.005;0.008) | 0.004 (0.003;0.005) | 0.006 (0.005;0.008) | 0.0129/0.996 | <0.001/0.010 |
| Rye | 0.006 (0.005;0.008) | 0.002 (0.002;0.003) | 0.003 (0.002;0.004) | |||
| Haemophilus | Wheat | 0.03 (0.02;0.04) | 0.05 (0.04;0.06) | 0.05 (0.04;0.07) | <0.001/0.011 | <0.001/0.032 |
| Rye | 0.04 (0.03;0.06) | 0.11 (0.08;0.14) | 0.11 (0.08;0.14) | |||
| ΔWeight | ΔFat Mass | ΔFat% | ΔCRP | ΔLDL Cholesterol | ΔHDL Cholesterol | ΔTriglyceride | ΔTotal Cholesterol | ΔGlucose | ΔInsulin | |
|---|---|---|---|---|---|---|---|---|---|---|
| ΔAgathobacter | ||||||||||
| - Rye group | 0.104 (0.284) | 0.082 (0.401) | 0.058 (0.552) | 0.115 (0.236) | 0.048 (0.622) | −0.051 (0.788) | −0.136 (0.160) | −0.080 (0.410) | 0.205 (0.034) | 0.1630 (0.092) |
| - Wheat group | −0.069 (0.498) | 0.010 (0.921) | 0.028 (0.784) | 0.0352 (0.730) | −0.038 (0.709) | −0.032 (0.757) | 0.017 (0.867) | −0.022 (0.828) | 0.170 (0.093) | 0.159 (0.116) |
| - Pooled groups | −0.033 (0.640) | −0.005 (0.942) | −0.005 (0.939) | 0.021 (0.760) | −0.025 (0.717) | −0.063 (0.371) | −0.055 (0.429) | −0.066 (0.348) | 0.178 (0.011) | 0.155 (0.025) |
| Δ[Ruminococcus] torques group | ||||||||||
| - Rye group | 0.045 (0.642) | −0.052 (0.595) | −0.116 (0.233) | 0.102 (0.292) | 0.135 (0.163) | 0.105 (0.742) | 0.057 (0.561) | 0.027 (0.785) | −0.018 (0.851) | −0.015 (0.882) |
| - Wheat group | 0.013 (0.901) | 0.065 (0.523) | 0.032 (0.751) | −0.242 (0.016) | −0.031 (0.760) | −0.096 (0.346) | −0.243 (0.016) | −0.177 (0.080) | −0.243 (0.015) | 0.064 (0.530) |
| - Pooled groups | 0.056 (0.420) | 0.034 (0.626) | −0.013 (0.856) | −0.002 (0.974) | 0.081 (0.248) | −0.026 (0.714) | −0.124 (0.074) | −0.065 (0.350) | −0.126 (0.070) | 0.016 (0.824) |
| Δ[Eubacterium] ventriosum group | ||||||||||
| - Rye group | −0.042 (0.665) | −0.097 (0.319) | −0.120 (0.216) | −0.119 (0.219) | −0.055 (0.570) | 0.122 (0.732) | −0.066 (0.497) | 0.019 (0.848) | 0.074 (0.445) | 0.133 (0.171) |
| - Wheat group | −0.069 (0.498) | −0.045 (0.659) | −0.079 (0.438) | −0.286 (0.004) | 0.046 (0.650) | 0.032 (0.754) | −0.117 (0.249) | 0.027 (0.788) | 0.036 (0.726) | 0.084 (0.408) |
| - Pooled groups | −0.021 (0.769) | −0.043 (0.534) | −0.070 (0.316) | −0.121 (0.081) | 0.022 (0.750) | 0.059 (0.399) | −0.104 (0.137) | 0.029 (0.680) | 0.064 (0.360) | 0.119 (0.088) |
| Δ Anaerotruncus | ||||||||||
| - Rye group | −0.120 (0.217) | −0.107 (0.272) | −0.093 (0.337) | 0.008 (0.931) | −0.154 (0.112) | −0.030 (0.405) | −0.254 (0.008) | −0.108 (0.267) | −0.035 (0.719) | −0.145 (0.133) |
| - Wheat group | −0.142 (0.161) | −0.160 (0.113) | −0.155 (0.124) | −0.097 (0.339) | −0.009 (0.933) | 0.170 (0.093) | −0.131 (0.196) | 0.099 (0.328) | −0.116 (0.251) | 0.123 (0.224) |
| - Pooled groups | −0.113 (0.104) | −0.113 (0.104) | −0.105 (0.132) | −0.021 (0.769) | −0.069 (0.321) | 0.052 (0.453) | −0.192 (0.006) | −0.005 (0.949) | −0.065 (0.349) | −0.016 (0.819) |
| Δ Anaerofilum | ||||||||||
| - Rye group | −0.002 (0.987) | −0.045 (0.643) | −0.056 (0.567) | 0.050 (0.607) | 0.023 (0.817) | 0.037 (0.917) | −0.105 (0.279) | 0.018 (0.850) | −0.039 (0.689) | 0.036 (0.708) |
| - Wheat group | 0.005 (0.961) | −0.022 (0.827) | 0.003 (0.973) | 0.093 (0.360) | 0.004 (0.966) | 0.114 (0.263) | 0.058 (0.571) | 0.093 (0.360) | −0.072 (0.477) | 0.014 (0.894) |
| - Pooled groups | 0.036 (0.611) | −0.005 (0.944) | 0.003 (0.961) | 0.113 (0.105) | 0.050 (0.477) | 0.090 (0.195) | −0.037 (0.600) | 0.068 (0.333) | −0.045 (0.523) | 0.028 (0.689) |
| Δ UCG−003 | ||||||||||
| - Rye group | −0.139 (0.153) | −0.124 (0.201) | −0.088 (0.367) | 0.052 (0.595) | 0.123 (0.204) | 0.037 (0.706) | 0.089 (0.357) | 0.109 (0.263) | −0.112 (0.248) | −0.136 (0.161) |
| - Wheat group | 0.154 (0.129) | 0.208 (0.039) | 0.180 (0.075) | −0.045 (0.656) | −0.006 (0.954) | −0.178 (0.078) | 0.145 (0.153) | −0.115 (0.259) | −0.041 (0.689) | −0.009 (0.931) |
| - Pooled groups | −0.030 (0.666) | −0.001 (0.985) | 0.005 (0.943) | −0.040 (0.566) | 0.053 (0.451) | −0.079 (0.257) | 0.123 (0.077) | 0.003 (0.964) | −0.091 (0.194) | −0.088 (0.205) |
| Δ Holdemania | ||||||||||
| - Rye group | −0.157 (0.106) | −0.215 (0.025) | −0.267 (0.005) | 0.012 (0.899) | 0.035 (0.717) | −0.054 (0.658) | −0.011 (0.914) | 0.049 (0.615) | 0.063 (0.519) | 0.081 (0.405) |
| - Wheat group | 0.173 (0.087) | 0.173 (0.086) | 0.138 (0.173) | 0.131 (0.196) | 0.096 (0.344) | −0.003 (0.976) | −0.134 (0.186) | 0.008 (0.940) | −0.093 (0.360) | −0.063 (0.538) |
| - Pooled groups | 0.049 (0.480) | 0.012 (0.865) | −0.037 (0.599) | 0.128 (0.065) | 0.088 (0.206) | 0.044 (0.526) | −0.095 (0.172) | 0.038 (0.590) | −0.007 (0.915) | 0.0206 (0.768) |
| Δ Haemophilus | ||||||||||
| - Rye group | −0.180 (0.062) | −0.124 (0.203) | −0.083 (0.393) | 0.068 (0.484) | 0.007 (0.939) | −0.019 (0.343) | −0.009 (0.930) | −0.018 (0.856) | −0.059 (0.543) | −0.064 (0.514) |
| - Wheat group | 0.101 (0.322) | 0.167 (0.099) | 0.179 (0.077) | −0.055 (0.591) | −0.031 (0.762) | −0.107 (0.294) | 0.019 (0.851) | −0.079 (0.436) | −0.175 (0.083) | −0.061 (0.552) |
| - Pooled groups | −0.082 (0.240) | −0.028 (0.685) | 0.003 (0.962) | −0.028 (0.687) | −0.023 (0.743) | −0.121 (0.082) | 0.019 (0.791) | −0.052 (0.460) | −0.105 (0.132) | −0.073 (0.295) |
| Week 0 * | Week 6 * | Week 12 * | p-Value Week 6 ** | p-Value Week 12 ** | ||
|---|---|---|---|---|---|---|
| Formic acid | Wheat | 87.5 (81.6; 93.8) | 84.5 (79.0; 90.3) | 88.2 (81.6; 95.2) | 0.5218 | 0.7776 |
| Rye | 84.3 (78.8; 90.1) | 84.94 (79.7; 90.6) | 87.6 (81.4; 94.3) | |||
| Acetic acid | Wheat | 114.5 (100.6; 130.4) | 95.3 (82.7; 109.8) | 95.2 (83.0; 109.3) | 0.0259 | 0.1989 |
| Rye | 111.8 (98.7; 126.6) | 114.9 (100.4; 131.5) | 105.4 (92.4; 120.3) | |||
| Propionic acid | Wheat | 0.65 (0.56; 0.74) | 0.62 (0.54; 0.71) | 0.62 (0.53; 0.71) | 0.2014 | 0.1640 |
| Rye | 0.60 (0.53; 0.69) | 0.67 (0.59; 0.77) | 0.68 (0.59; 0.78) | |||
| Butyric acid | Wheat | 0.84 (0.68; 1.02) | 0.69 (0.57; 0.83) | 0.78 (0.63; 0.95) | <0.0001 | 0.0270 |
| Rye | 0.68 (0.56; 0.82) | 0.99 (0.83; 1.19) | 0.93 (0.77; 1.12) | |||
| Isobutyric acid | Wheat | 0.17 (0.15; 0.19) | 0.16 (0.14; 0.18) | 0.17 (0.15; 0.18) | 0.8106 | 0.3958 |
| Rye | 0.16 (0.14; 0.17) | 0.16 (0.14; 0.17) | 0.15 (0.14; 0.17) | |||
| Succinic acid | Wheat | 3.82 (3.57; 4.08) | 3.82 (3.56; 4.09) | 3.81 (3.55; 4.1) | 0.5560 | 0.9472 |
| Rye | 3.81 (3.58; 4.06) | 3.91 (3.66; 4.18) | 3.80 (3.54; 4.07) | |||
| Valeric acid | Wheat | 0.24 (0.21; 0.28) | 0.21 (0.18; 0.24) | 0.22 (0.19; 0.26) | 0.2574 | 0.6612 |
| Rye | 0.20 (0.18; 0.23) | 0.22 (0.19; 0.26) | 0.20 (0.17; 0.23) | |||
| Isovaleric acid | Wheat | 1.08 (0.98; 1.18) | 0.93 (0.83; 1.04) | 0.98 (0.88; 1.08) | 0.7735 | 0.8242 |
| Rye | 0.94 (0.86; 1.02) | 0.86 (0.77; 0.96) | 0.94 (0.86; 1.04) | |||
| Caproic acid | Wheat | 0.47 (0.42; 0.52) | 0.45 (0.4; 0.5) | 0.47 (0.42; 0.53) | 0.5455 | 0.3119 |
| Rye | 0.43 (0.39; 0.47) | 0.45 (0.4; 0.5) | 0.42 (0.37; 0.47) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iversen, K.N.; Dicksved, J.; Zoki, C.; Fristedt, R.; Pelve, E.A.; Langton, M.; Landberg, R. The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients 2022, 14, 1669. https://doi.org/10.3390/nu14081669
Iversen KN, Dicksved J, Zoki C, Fristedt R, Pelve EA, Langton M, Landberg R. The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients. 2022; 14(8):1669. https://doi.org/10.3390/nu14081669
Chicago/Turabian StyleIversen, Kia Nøhr, Johan Dicksved, Camille Zoki, Rikard Fristedt, Erik A. Pelve, Maud Langton, and Rikard Landberg. 2022. "The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study)" Nutrients 14, no. 8: 1669. https://doi.org/10.3390/nu14081669
APA StyleIversen, K. N., Dicksved, J., Zoki, C., Fristedt, R., Pelve, E. A., Langton, M., & Landberg, R. (2022). The Effects of High Fiber Rye, Compared to Refined Wheat, on Gut Microbiota Composition, Plasma Short Chain Fatty Acids, and Implications for Weight Loss and Metabolic Risk Factors (the RyeWeight Study). Nutrients, 14(8), 1669. https://doi.org/10.3390/nu14081669