
Exploratory Analysis of Selected Components of the mTOR Pathway Reveals Potentially Crucial Associations with Childhood Malnutrition

 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Materials and Methods
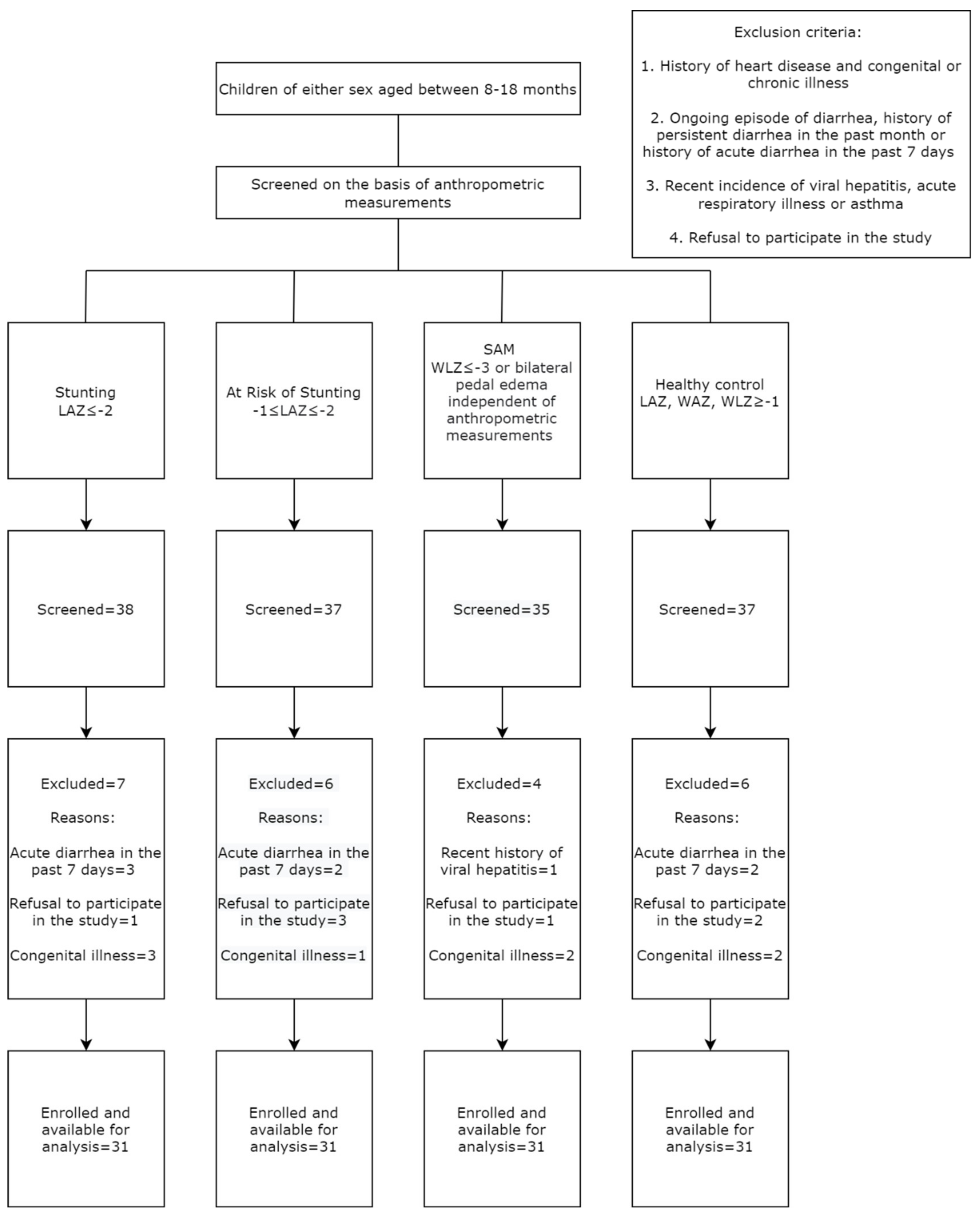
2.1. Study Design, Setting and Population
2.2. Enrolment of Study Participants and Collection of Socio-Demographic Data
2.3. Sample Size
2.4. Sample Collection and Processing
2.5. Extraction of Total Intracellular Proteins
2.6. Assessment of Activity of Phospho-mTORC1 Using ELISA
2.7. Isolation of Total RNA and cDNA Synthesis by Reverse Transcriptase PCR (RT-PCR)
2.8. Expression Analysis of Selected mTOR Pathway Genes
2.9. Statistical Analysis
3. Results
3.1. Socio-Demographic, Anthropometric and Feeding Data of the Study Participants
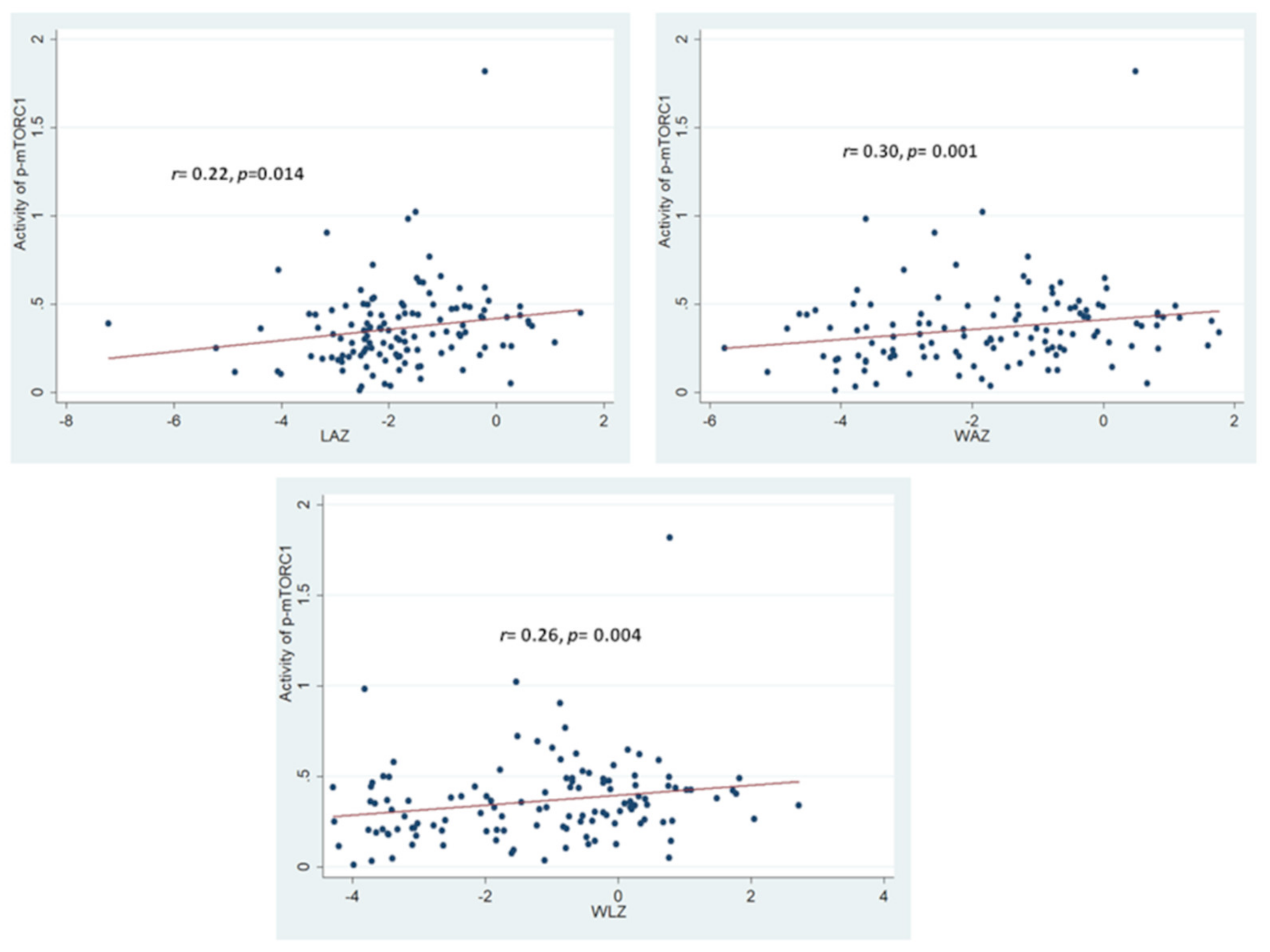
3.2. Assessment of Activity Phospho-mTORC1 and Correlation with Anthropometric Outcomes
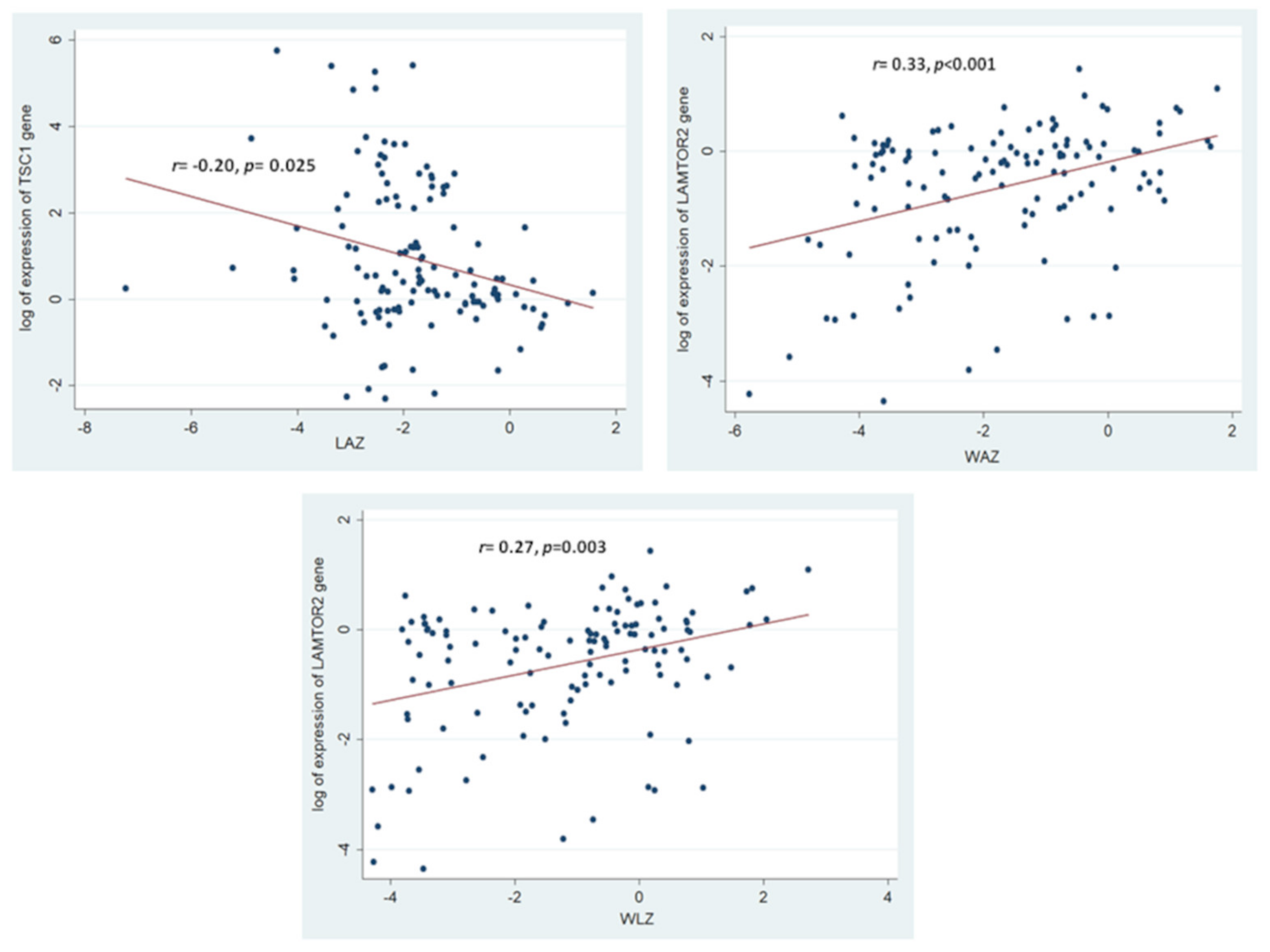
3.3. Association of the Assessed mTOR Pathway Components with Different Forms of Childhood Malnutrition
4. Discussion
Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ibrahim, M.K.; Zambruni, M.; Melby, C.L.; Melby, P.C. Impact of childhood malnutrition on host defense and infection. Clin. Microbiol. Rev. 2017, 30, 919–971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Onis, M.; Onyango, A.W. WHO child growth standards. Lancet 2008, 371, 204. [Google Scholar] [CrossRef] [Green Version]
- De Onis, M.; Branca, F. Childhood stunting: A global perspective. Matern. Child Nutr. 2016, 12, 12–26. [Google Scholar] [CrossRef] [PubMed]
- Mahfuz, M.; Das, S.; Mazumder, R.N.; Rahman, M.M.; Haque, R.; Bhuiyan, M.M.R.; Akhter, H.; Sarker, M.S.A.; Mondal, D.; Muaz, S.S.A.; et al. Bangladesh environmental enteric dysfunction (BEED) study: Protocol for a community-based intervention study to validate non-invasive biomarkers of environmental enteric dysfunction. BMJ Open 2017, 7, e017768. [Google Scholar] [CrossRef] [Green Version]
- Saxton, R.A.; Sabatini, D.M. mTOR signaling in growth, metabolism, and disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Sarbassov, D.D.; Ali, S.M.; Sabatini, D.M. Growing roles for the mTOR pathway. Curr. Opin. Cell Biol. 2005, 17, 596–603. [Google Scholar] [CrossRef]
- Laplante, M.; Sabatini, D.M. mTOR signaling in growth control and disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [Green Version]
- Dillon, E.L. Nutritionally essential amino acids and metabolic signaling in aging. Amino Acids 2013, 45, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Chiang, G.G.; Abraham, R.T. Phosphorylation of mammalian target of rapamycin (mTOR) at Ser-2448 is mediated by p70S6 kinase. J. Biol. Chem. 2005, 280, 25485–25490. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Dibble, C.C.; Matsuzaki, M.; Manning, B.D. The TSC1-TSC2 complex is required for proper activation of mTOR complex 2. Mol. Cell. Biol. 2008, 28, 4104–4115. [Google Scholar] [CrossRef] [Green Version]
- Aylett, C.H.; Sauer, E.; Imseng, S.; Boehringer, D.; Hall, M.N.; Ban, N.; Maier, T. Architecture of human mTOR complex 1. Science 2016, 351, 48–52. [Google Scholar] [CrossRef]
- Scheffler, J.M.; Sparber, F.; Tripp, C.H.; Herrmann, C.; Humenberger, A.; Blitz, J.; Romani, N.; Stoitzner, P.; Huber, L.A. LAMTOR2 regulates dendritic cell homeostasis through FLT3-dependent mTOR signalling. Nat. Commun. 2014, 5, 5138. [Google Scholar] [CrossRef]
- Bahrami, B.F.; Ataie-Kachoie, P.; Pourgholami, M.H.; Morris, D.L. p70 Ribosomal protein S6 kinase (Rps6kb1): An update. J. Clin. Pathol. 2014, 67, 1019–1025. [Google Scholar] [CrossRef]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005, 307, 1098–1101. [Google Scholar] [CrossRef] [Green Version]
- Thul, P.J.; Lindskog, C. The human protein atlas: A spatial map of the human proteome. Protein Sci. 2018, 27, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Jing, Y.; Qiao, J.; Luan, B.; Wang, X.; Wang, L.; Song, Z. Activation of the mTOR signaling pathway is required for asthma onset. Sci. Rep. 2017, 7, 4532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casadei, K.; Kiel, J. Anthropometric Measurement; StatPearls: Treasure Island, FL, USA, 2020. [Google Scholar]
- Charan, J.; Biswas, T. How to calculate sample size for different study designs in medical research? Indian J. Psychol. Med. 2013, 35, 121. [Google Scholar] [CrossRef] [Green Version]
- Turchiano, M.; Nguyen, C.; Fierman, A.; Lifshitz, M.; Convit, A. Impact of blood sample collection and processing methods on glucose levels in community outreach studies. J. Environ. Public Health 2013, 2013, 256151. [Google Scholar] [CrossRef]
- World Health Organization. Guideline: Updates on the Management of Severe Acute Malnutrition in Infants and Children; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Fuss, I.J.; Kanof, M.E.; Smith, P.D.; Zola, H. Isolation of whole mononuclear cells from peripheral blood and cord blood. Curr. Protoc. Immunol. 2009, 85, 7.1.1–7.1.8. [Google Scholar] [CrossRef]
- Louis, K.S.; Siegel, A.C. Cell viability analysis using trypan blue: Manual and automated methods. In Mammalian Cell Viability; Springer: Berlin/Heidelberg, Germany, 2011; pp. 7–12. [Google Scholar]
- Riedhammer, C.; Halbritter, D.; Weissert, R. Peripheral blood mononuclear cells: Isolation, freezing, thawing, and culture. In Multiple Sclerosis; Springer: Berlin/Heidelberg, Germany, 2014; pp. 53–61. [Google Scholar]
- Allen, S.A.; Tomilov, A.; Cortopassi, G.A. Small molecules bind human mTOR protein and inhibit mTORC1 specifically. Biochem. Pharmacol. 2018, 155, 298–304. [Google Scholar] [CrossRef]
- Desjardins, P.; Conklin, D. NanoDrop microvolume quantitation of nucleic acids. JoVE J. Vis. Exp. 2010, 45, e2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Zongrone, A.; Winskell, K.; Menon, P. Infant and young child feeding practices and child undernutrition in Bangladesh: Insights from nationally representative data. Public Health Nutr. 2012, 15, 1697–1704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scherbaum, V.; Srour, M.L. The role of breastfeeding in the prevention of childhood malnutrition. In Hidden Hunger: Malnutrition and the First 1000 Days of Life: Causes, Consequences and Solutions; Karger Publishers: Basel, Switzerland, 2016; Volume 115, pp. 82–97. [Google Scholar]
- Ahmed, T.; Hossain, M.; Mahfuz, M.; Choudhury, N.; Ahmed, S. Imperatives for reducing child stunting in Bangladesh. Matern. Child Nutr. 2016, 12 (Suppl. S1), 242. [Google Scholar] [CrossRef] [Green Version]
- Branca, F.; Ferrari, M. Impact of micronutrient deficiencies on growth: The stunting syndrome. Ann. Nutr. Metab. 2002, 46 (Suppl. S1), 8–17. [Google Scholar] [CrossRef]
- Ramakrishnan, U.; Nguyen, P.; Martorell, R. Effects of micronutrients on growth of children under 5 y of age: Meta-analyses of single and multiple nutrient interventions. Am. J. Clin. Nutr. 2009, 89, 191–203. [Google Scholar] [CrossRef]
- Wang, A.Z.; Shulman, R.J.; Crocker, A.H.; Thakwalakwa, C.; Maleta, K.M.; Devaraj, S.; Manary, M.J.; Trehan, I. A combined intervention of zinc, multiple micronutrients, and albendazole does not ameliorate environmental enteric dysfunction or stunting in rural Malawian children in a double-blind randomized controlled trial. J. Nutr. 2017, 147, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, T.; Auble, D.; Berkley, J.; Black, R.; Ahern, P.P.; Hossain, M.; Hsieh, A.; Ireen, S.; Arabi, M.; Gordon, J.I. An evolving perspective about the origins of childhood undernutrition and nutritional interventions that includes the gut microbiome. Ann. N. Y. Acad. Sci. 2014, 1332, 22–38. [Google Scholar] [CrossRef] [Green Version]
- Duggal, P.; Petri, W.A., Jr. Does malnutrition have a genetic component? Annu. Rev. Genom. Hum. Genet. 2018, 19, 247–262. [Google Scholar] [CrossRef]
- Trehan, I.; Kelly, P.; Shaikh, N.; Manary, M.J. New insights into environmental enteric dysfunction. Arch. Dis. Child. 2016, 101, 741–744. [Google Scholar] [CrossRef]
- De Mello, V.D.F.; Kolehmanien, M.; Schwab, U.; Pulkkinen, L.; Uusitupa, M. Gene expression of peripheral blood mononuclear cells as a tool in dietary intervention studies: What do we know so far? Mol. Nutr. Food Res. 2012, 56, 1160–1172. [Google Scholar] [CrossRef]
- Yohannes, D.A.; De Kauwe, A.; Kaukinen, K.; Kurppa, K.; Mäki, M.; Anderson, R.P.; Linnarsson, S.; Greco, D.; Saavalainen, P. Effects of in vivo gluten challenge on PBMC gene expression profiles in diet treated celiac disease. Front. Immunol. 2020, 11, 594243. [Google Scholar] [CrossRef]
- DeBoer, M.D.; Scharf, R.J.; Leite, A.M.; Férrer, A.; Havt, A.; Pinkerton, R.; Lima, A.A.; Guerrant, R.L. Systemic inflammation, growth factors, and linear growth in the setting of infection and malnutrition. Nutrition 2017, 33, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Millward, D.J. Nutrition, infection and stunting: The roles of deficiencies of individual nutrients and foods, and of inflammation, as determinants of reduced linear growth of children. Nutr. Res. Rev. 2017, 30, 50. [Google Scholar] [CrossRef]
- Majoros, A.; Platanitis, E.; Kernbauer-Hölzl, E.; Rosebrock, F.; Müller, M.; Decker, T. Canonical and non-canonical aspects of JAK–STAT signaling: Lessons from interferons for cytokine responses. Front. Immunol. 2017, 8, 29. [Google Scholar] [CrossRef] [Green Version]
- Batool, R.; Butt, M.S.; Sultan, M.T.; Saeed, F.; Naz, R. Protein–energy malnutrition: A risk factor for various ailments. Crit. Rev. Food Sci. Nutr. 2015, 55, 242–253. [Google Scholar] [CrossRef]
- Semba, R.D.; Shardell, M.; Ashour, F.A.S.; Moaddel, R.; Trehan, I.; Maleta, K.M.; Ordiz, M.I.; Kraemer, K.; Khadeer, M.A.; Ferrucci, L.; et al. Child stunting is associated with low circulating essential amino acids. EBioMedicine 2016, 6, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Nuss, E.T.; Tanumihardjo, S.A. Quality protein maize for Africa: Closing the protein inadequacy gap in vulnerable populations. Adv. Nutr. 2011, 2, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Gazi, M.A.; Das, S.; Siddique, M.A.; Alam, M.A.; Fahim, S.M.; Hasan, M.M.; Hossaini, F.; Kabir, M.M.; Noor, Z.; Haque, R.; et al. Plasma Kynurenine to Tryptophan Ratio Is Negatively Associated with Linear Growth of Children Living in a Slum of Bangladesh: Results from a Community-Based Intervention Study. Am. J. Trop. Med. Hyg. 2021, 104, 766–773. [Google Scholar] [CrossRef]
- Moreau, G.B.; Ramakrishnan, G.; Cook, H.L.; Fox, T.E.; Nayak, U.; Ma, J.Z.; Colgate, E.R.; Kirkpatrick, B.D.; Haque, R.; Petri, W.A., Jr. Childhood growth and neurocognition are associated with distinct sets of metabolites. EBioMedicine 2019, 44, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Lisse, T.S.; Hewison, M. Vitamin D: A New Player in the World of mTOR Signaling; Taylor & Francis: Abingdon, UK, 2011; pp. 937–947. [Google Scholar]
- Fernandes, S.A.; Demetriades, C. The multifaceted role of nutrient sensing and mTORC1 signaling in physiology and aging. Front. Aging 2021, 2, 707372. [Google Scholar] [CrossRef]
- Wyant, G.A.; Abu-Remaileh, M.; Wolfson, R.L.; Chen, W.W.; Freinkman, E.; Danai, L.V.; Heiden, M.G.V.; Sabatini, D.M. mTORC1 activator SLC38A9 is required to efflux essential amino acids from lysosomes and use protein as a nutrient. Cell 2017, 171, 642–654.e12. [Google Scholar] [CrossRef]
- Deval, C.; Chaveroux, C.; Maurin, A.C.; Cherasse, Y.; Parry, L.; Carraro, V.; Milenkovic, D.; Ferrara, M.; Bruhat, A.; Jousse, C.; et al. Amino acid limitation regulates the expression of genes involved in several specific biological processes through GCN2-dependent and GCN2-independent pathways. FEBS J. 2009, 276, 707–718. [Google Scholar] [CrossRef]
- Boukhettala, N.; Claeyssens, S.; Bensifi, M.; Maurer, B.; Abed, J.; Lavoinne, A.; Déchelotte, P.; Coëffier, M. Effects of essential amino acids or glutamine deprivation on intestinal permeability and protein synthesis in HCT-8 cells: Involvement of GCN2 and mTOR pathways. Amino Acids 2012, 42, 375–383. [Google Scholar] [CrossRef]
- Gallinetti, J.; Harputlugil, E.; Mitchell, J.R. Amino acid sensing in dietary-restriction-mediated longevity: Roles of signal-transducing kinases GCN2 and TOR. Biochem. J. 2013, 449, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ye, J.; Palm, W.; Peng, M.; King, B.; Lindsten, T.; Li, M.O.; Koumenis, C.; Thompson, C.B. GCN2 sustains mTORC1 suppression upon amino acid deprivation by inducing Sestrin2. Genes Dev. 2015, 29, 2331–2336. [Google Scholar] [CrossRef] [Green Version]
- Chotechuang, N.; Azzout-Marniche, D.; Bos, C.; Chaumontet, C.; Gausserès, N.; Steiler, T.; Gaudichon, C.; Tomé, D. mTOR, AMPK, and GCN2 coordinate the adaptation of hepatic energy metabolic pathways in response to protein intake in the rat. Am. J. Physiol. Endocrinol. Metab. 2009, 297, E1313–E1323. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Kim, E.; Yuan, H.; Inoki, K.; Goraksha-Hicks, P.; Schiesher, R.L.; Neufeld, T.P.; Guan, K. Regulation of mTORC1 by the Rab and Arf GTPases. J. Biol. Chem. 2010, 285, 19705–19709. [Google Scholar] [CrossRef] [Green Version]
- Semba, R.D.; Trehan, I.; Gonzalez-Freire, M.; Kraemer, K.; Moaddel, R.; Ordiz, M.I.; Ferrucci, L.; Manary, M.J. Perspective: The potential role of essential amino acids and the mechanistic target of rapamycin complex 1 (mTORC1) pathway in the pathogenesis of child stunting. Adv. Nutr. 2016, 7, 853–865. [Google Scholar] [CrossRef] [Green Version]
- Parmigiani, A.; Nourbakhsh, A.; Ding, B.; Wang, W.; Kim, Y.C.; Akopiants, K.; Guan, K.L.; Karin, M.; Budanov, A.V. Sestrins inhibit mTORC1 kinase activation through the GATOR complex. Cell Rep. 2014, 9, 1281–1291. [Google Scholar] [CrossRef] [Green Version]
- Oh, W.J.; Jacinto, E. mTOR complex 2 signaling and functions. Cell Cycle 2011, 10, 2305–2316. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Stunted (n = 31) | At Risk of Stunting (n = 31) | SAM (n = 31) | Healthy Control (n = 31) |
|---|---|---|---|---|
| Age in months | 12 (10, 14) | 12 (8.5, 15.5) | 11 (9, 13) | 12 (9.5, 14.5) |
| Male sex, n (%) | 16 (51.6%) | 20 (64.5%) | 19 (61.3%) | 16 (51.6%) |
| Weight in kg | 7.2 (6.8, 7.6) | 8.15 (7.7, 8.6) | 6.51 (6.2, 6.8) | 9.82 (8.9, 10.7) |
| Length in cm | 69.5 (67.2, 71.8) | 70.3 (66.6, 74) | 71.3 (69.4, 73.3) | 75.4 (72.5, 78.2) |
| Mid Upper Arm Circumference in cm | 135 (128, 142) | 141.5 (136, 147) | 120 (118, 123) | 150 (144, 156) |
| Head circumference in cm | 43.5 (43, 44) | 44.5 (43.3, 45.7) | 44 (42.5, 45.5) | 44.8 (44, 45.6) |
| Birth weight in kg | 2.5 (2.25, 2.75) | 3 (2.55, 3.45) | 2.8 (2.55, 3.05) | 3.1 (2.70, 3.50) |
| Weight for age Z score | −2.42 (−2.96, −1.89) | −1.17 (−1.44, −0.9) | −3.75 (−4, −3.54) | 0.5 (−0.2, 1.2) |
| Length for age Z score | −2.35 (−2.6, −2.1) | −1.62 (−1.8, −1.44) | −2.46 (−3.26, −1.67) | −0.22 (−0.76, 0.30) |
| Weight for length Z score | −1.57 (−2.21, −0.93) | −0.45 (−1.03, −0.13) | −3.66 (−3.81, −3.52) | 0.36 (−0.44, 1.16) |
| Gestational age in weeks | 38 (36.8, 39.3) | 39 (38.5, 39.5) | 38 (37, 39) | 39 (38, 40) |
| Maternal age in years | 23 (19.3, 26.8) | 22.5 (17, 28) | 25 (20, 30) | 24.5 (22.4, 26.6) |
| Duration of EBF in months | 3 (0, 6) | 1 (0, 3.75) | 3 (0, 6) | 3 (0, 6) |
| Currently breastfeeding, n (%) | 28 (90.3%) | 29 (93.6%) | 22 (71%) | 29 (93.6%) |
| Currently formula feeding, n (%) | 15 (48.4%) | 16 (51.6%) | 29 (93.6%) | 19 (61.3%) |
| Currently fed with rice powder or suji, n (%) | 14 (45.2%) | 16 (51.6%) | 28 (90.3%) | 14 (45.2%) |
| Currently fed with cow milk, n (%) | 9 (29%) | 8 (25.8%) | 17 (54.8%) | 6 (19.4%) |
| Peripheral blood mononuclear cell count, million cells/ml | 4 (2.6, 5.3) | 4.35 (1.65, 5.7) | 4.75 (3.85, 5.65) | 4.33 (2.83, 5.83) |
| Expression of mTOR Pathway Gene, 2−∆∆Ct | Stunted vs. Healthy Control | At Risk of Stunting vs. Healthy Control | SAM vs. Healthy Control | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Stunted | Healthy Control | p-Value | At Risk of Stunting | Healthy Control | p-Value | SAM | Healthy Control | p-Value | |
| MTOR | 0.701 (0.233–1.172) | 0.882 (0.369–1.389) | 0.032 | 0.549 (0.151–0.963) | 0.882 (0.369–1.389) | 0.001 | 0.748 (0.331–1.268) | 0.882 (0.369–1.389) | 0.088 |
| TSC1 | 1.822 (1.031–2.638) | 0.972 (0.733–1.202) | 0.016 | 3.331 (3.331–15.902) | 0.972 (0.733–1.202) | 0.001 | 2.062 (2.056–18) | 0.972 (0.733–1.202) | 0.002 |
| LAMTOR2 | 0.688 (0.298–1.078) | 0.991 (0.459–1.522) | 0.003 | 0.823 (0.411–1.221) | 0.991 (0.459–1.522) | 0.037 | 0.627 (0.211–1.062) | 0.991 (0.459–1.522) | 0.007 |
| RPS6K1 | 1.162 (1.162–3.623) | 0.904 (0.411–1.381) | 0.128 | 0.968 (0.242–1.713) | 0.904 (0.411–1.381) | 0.977 | 1.289 (0.532–2.044) | 0.904 (0.411–1.381) | 0.438 |
| RICTOR | 0.731 (0.322–1.131) | 0.869 (0.377–1.364) | 0.109 | 0.473 (0.112–0.828) | 0.869 (0.377–1.364) | 0.013 | 0.854 (0.472–1.619) | 0.869 (0.377–1.364) | 0.603 |
| mTOR Pathway Components Analyzed | Stunted vs. Healthy Control | At Risk of Stunting vs. Healthy Control | SAM vs. Healthy Control | |||
|---|---|---|---|---|---|---|
| Odds Ratio (95% CI) | p-Value | Odds Ratio (95% CI) | p-Value | Odds Ratio (95% CI) | p-Value | |
| phospho-mTORC1 | 0.102 (0.02–0.803) | 0.042 | 0.063 (0.02–2.162) | 0.801 | 0.202 (0.012–0.822) | 0.043 |
| MTOR gene | 0.377 (0.102–1.453) | 0.161 | 0.153 (0.042–0.639) | 0.011 | 0.203 (0.041–1.073) | 0.060 |
| TSC1 gene | 1.622 (1.041–2.513) | 0.031 | 2.509 (1.261–5.011) | 0.009 | 3.151 (0.788–12.511) | 0.012 |
| LAMTOR2 gene | 0.111 (0.023–0.494) | 0.042 | 0.188 (0.061–0.612) | 0.006 | 0.204 (0.042–0.974) | 0.046 |
| RPS6K1 gene | 1.012 (0.968–1.048) | 0.686 | 0.992 (0.943–1.054) | 0.831 | 0.981 (0.842–1.134) | 0.763 |
| RICTOR gene | 1.063 (0.881–1.282) | 0.528 | 0.788 (0.517–1.223) | 0.289 | 0.514 (0.191–1.412) | 0.204 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Palit, P.; Gazi, M.A.; Das, S.; Hasan, M.M.; Noor, Z.; Ferdous, J.; Alam, M.A.; Nuzhat, S.; Islam, M.R.; Mahfuz, M.; et al. Exploratory Analysis of Selected Components of the mTOR Pathway Reveals Potentially Crucial Associations with Childhood Malnutrition. Nutrients 2022, 14, 1612. https://doi.org/10.3390/nu14081612
Palit P, Gazi MA, Das S, Hasan MM, Noor Z, Ferdous J, Alam MA, Nuzhat S, Islam MR, Mahfuz M, et al. Exploratory Analysis of Selected Components of the mTOR Pathway Reveals Potentially Crucial Associations with Childhood Malnutrition. Nutrients. 2022; 14(8):1612. https://doi.org/10.3390/nu14081612
Chicago/Turabian StylePalit, Parag, Md Amran Gazi, Subhasish Das, Md Mehedi Hasan, Zannatun Noor, Jafrin Ferdous, Md Ashraful Alam, Sharika Nuzhat, Md Ridwan Islam, Mustafa Mahfuz, and et al. 2022. "Exploratory Analysis of Selected Components of the mTOR Pathway Reveals Potentially Crucial Associations with Childhood Malnutrition" Nutrients 14, no. 8: 1612. https://doi.org/10.3390/nu14081612
APA StylePalit, P., Gazi, M. A., Das, S., Hasan, M. M., Noor, Z., Ferdous, J., Alam, M. A., Nuzhat, S., Islam, M. R., Mahfuz, M., Haque, R., & Ahmed, T. (2022). Exploratory Analysis of Selected Components of the mTOR Pathway Reveals Potentially Crucial Associations with Childhood Malnutrition. Nutrients, 14(8), 1612. https://doi.org/10.3390/nu14081612