
Beyond the Calorie Paradigm: Taking into Account in Practice the Balance of Fat and Carbohydrate Oxidation during Exercise?

 ,
, 
Abstract
1. Introduction: Can Exercise Be Defined on the Basis of the Fuel It Oxidizes?
2. The Balance of Substrates during Exercise
2.1. Balance of Substrate Oxidation during Exercise: The “Crossover Concept”
2.2. Measuring the Maximal Lipid Oxidation during Exercise
2.3. Reproducibility and Stability over Time
2.4. Measuring the Kinetics of Carbohydrate Oxidation during Exercise
3. Physiological Relevance and Determinants of LIPOXmax
3.1. The LIPOXmax Is the Level of Exercise Spontaneously Selected for Prolonged Exercise: The Hypothesis of the ‘Healthy Primitive Lifestyle’
3.2. The Balance of Substrates as a Window on Mitochondrial Function and Metabolic Flexibility
3.3. Carbohydrate Breakdown Controls Lipid Oxidation Rate
3.4. Agreement between the Balance of Substrates and Lactate/Ventilatory Threshold
3.5. The LIPOXmax as a Marker of Functional Capacity
4. Various Influences
4.1. Genetics
4.2. Effects of Exercise and Training on Lipid Oxidation
4.3. Age and Gender
4.4. Feeding and Dietary Habits
4.5. Hormones
5. Targeting Exercise Training on Lipid Oxidation
5.1. Exercise (Regardless of Its Mode or Intensity) Is a Powerful Therapeutic Tool in Metabolic Diseases
5.2. The Surprising Efficacy of Low-to Moderate Intensity Training
5.3. Training at the LIPOXmax for the Treatment of Obesity
5.3.1. Effects of LIPOXmax Training on Obesity and Associated Disorders in Obese and Overweight Subjects
5.3.2. Sparing Effects of LIPOXmax Training on Fat-Free Mass (FFM)
5.3.3. Metabolic Effects of LIPOXmax Training
5.3.4. LIPOXmax Training Improves Inflammatory Status
5.3.5. LIPOXmax Training Also Decreases Blood Viscosity
5.4. LIPOXmax Training in Adolescents Suffering from Obesity
5.5. Bariatric Surgery
5.6. Diabetes
5.7. LIPOXmax Training in Patients Gaining Weight under Psychotropic Drugs
5.8. LIPOXmax Training in Cancer
5.9. Osteoarthritis
5.10. How to Follow This Low-Intensity Training?
6. How Can We Explain the Unexpected Efficacy of Low Volumes of Low-Intensity Exercise?
6.1. LIPOXmax Training Improves Mitochondrial Respiration and the Krebs Cycle Function
6.2. Secretion of Chemical Messengers (‘Myometabokines’) by Muscle
6.3. Sedentarity and Exercise as Epigenetic Programmers of Various Functions in the Body
6.4. Sedentarity and Exercise Effects on Appetite
7. Other Strategies Based on the Balance of Substrates during Exercise
7.1. Targeting Exercise at the Level of the “Point of Crossover”
7.2. Optimal Level of Fat/Carbohydrate Oxidation Ratio (OLORFOX)
8. Concluding Remarks
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Perez-Martin, A.; Mercier, J. Stress tests and exercise training program for diabetics-Initial metabolic evaluation. Ann. Endocrinol. 2001, 62, 291–293. [Google Scholar]
- Dériaz, O.; Dumont, M.; Bergeron, N.; Després, J.P.; Brochu, M.; Prud’homme, D. Skeletal muscle low attenuation area and maximal fat oxidation rate during submaximal exercise in male obese individuals. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 1579–1584. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Achten, J.; Gleeson, M.; Jeukendrup, A.E. Determination of the exercise intensity that elicits maximal fat oxidation. Med. Sci. Sports Exerc. 2002, 34, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E. Regulation of endogenous fat and carbohydrate metabolism in relation to exercise intensity and duration. Am. J. Physiol. 1993, 265, E380–E391. [Google Scholar] [CrossRef]
- Bergman, B.C.; Brooks, G.A. Respiratory gas-exchange ratios during graded exercise in fed and fasted trained and untrained men. J. Appl. Physiol. 1999, 86, 479–487. [Google Scholar] [CrossRef]
- Friedlander, A.L.; Jacobs, K.A.; Fattor, J.A.; Horning, M.A.; Hagobian, T.A.; Bauer, T.A. Contributions of working muscle to whole body lipid metabolism are altered by exercise intensity and training. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E107–E116. [Google Scholar] [CrossRef]
- Dumortier, M.; Pérez-Martin, A.; Pierrisnard, E.; Mercier, J.; Brun, J.F. Regular exercise (3 × 45 min/wk) decreases plasma viscosity in sedentary obese, insulin resistant patients parallel to an improvement in fitness and a shift in substrate oxidation balance. Clin. Hemorheol. Microcirc. 2002, 26, 219–229. [Google Scholar]
- Brandou, F.; Dumortier, M.; Garandeau, P.; Mercier, J.; Brun, J.F. Effects of a two-month rehabilitation program on substrate utilization during exercise in obese adolescents. Diabetes Metab. 2003, 29, 20–27. [Google Scholar] [CrossRef]
- Achten, J.; Jeukendrup, A.E. Optimizing fat oxidation through exercise and diet. Nutrition 2004, 20, 716–727. [Google Scholar] [CrossRef]
- Petridou, A.; Siopi, A.; Mougios, V. Exercise in the management of obesity. Metabolism 2019, 92, 163–169. [Google Scholar] [CrossRef]
- Swift, D.L.; McGee, J.E.; Earnest, C.P.; Carlisle, E.; Nygard, M.; Johannsen, N.M. The Effects of Exercise and Physical Activity on Weight Loss and Maintenance. Prog. Cardiovasc. Dis. 2018, 61, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Ohkawara, K.; Tanaka, S.; Miyachi, M.; Ishikawa-Takata, K.; Tabata, I. A dose-response relation between aerobic exercise and visceral fat reduction: Systematic review of clinical trials. Int. J. Obes. 2007, 31, 1786–1797. [Google Scholar] [CrossRef]
- Amaro-Gahete, F.J.; Sanchez-Delgado, G.; Jurado-Fasoli, L.; De-la-O, A.; Castillo, M.J.; Helge, J.W.; Ruiz, J.R. Assessment of maximal fat oxidation during exercise: A systematic review. Scand. J. Med. Sci. Sports 2019, 29, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Maunder, E.; Plews, D.J.; Kilding, A.E. Contextualizing maximal fat oxidation during exercise: Determinants and normative values. Front. Physiol. 2018, 9, 599. [Google Scholar] [CrossRef]
- Calonne, J.; Fares, E.J.; Montani, J.P.; Schutz, Y.; Dulloo, A.; Isacco, L. Dynamics of fat oxidation from sitting at rest to light exercise in inactive young humans. Metabolites 2021, 11, 334. [Google Scholar] [CrossRef] [PubMed]
- Chávez-Guevara, I.A.; Urquidez-Romero, R.; Pérez-León, J.A.; González-Rodríguez, E.; Moreno-Brito, V.; Ramos-Jiménez, A. Chronic effect of fatmax training on body weight, fat mass, and cardiorespiratory fitness in obese subjects: A meta-analysis of randomized clinical trials. Int. J. Environ. Res. Public Health 2020, 17, 7888. [Google Scholar] [CrossRef] [PubMed]
- Drapier, E.; Cherif, A.; Richou, M.; Bughin, F.; Fedou, C.; Mercier, J.; Avignon, A.; Brun, J.F. Long term (3 years) weight loss after low-intensity endurance training targeted at the level of maximal muscular lipid oxidation. Integr. Obes. Diabetes 2018, 4, 1–6. [Google Scholar] [CrossRef]
- Hammoudi, L.; Brun, J.F.; Noirez, P.; Bui, G.; Chevalier, C.; Gimet, F.; Mercier, J.; de Mauverger, E.R. Effects of 2 years endurance training targeted at the level of maximal lipid oxidation on body composition. Sci. Sports 2020, 35, 350–357. [Google Scholar] [CrossRef]
- Slentz, C.A.; Aiken, L.B.; Houmard, J.A.; Bales, C.W.; Johnson, J.L.; Tanner, C.J.; Duscha, B.D.; Kraus, W.E. Inactivity, exercise, and visceral fat. STRRIDE: A randomized, controlled study of exercise intensity and amount. J. Appl. Physiol. 2005, 99, 1613–1618. [Google Scholar] [CrossRef]
- McGarrah, R.W.; Slentz, C.A.; Kraus, W.E. The effect of vigorous-versus moderate-intensity aerobic exercise on insulin action. Curr. Cardiol. Rep. 2016, 18, 117. [Google Scholar] [CrossRef]
- Slentz, C.A.; Houmard, J.A.; Kraus, W.E. Modest exercise prevents the progressive disease associated with physical inactivity. Exerc. Sport Sci. Rev. 2007, 35, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Huffman, K.M.; Koves, T.R.; Hubal, M.J.; Abouassi, H.; Beri, N.; Bateman, L.A.; Stevens, R.D.; Ilkayeva, O.R.; Hoffman, E.P.; Muoio, D.M.; et al. Metabolite signatures of exercise training in human skeletal muscle relate to mitochondrial remodeling and cardiometabolic fitness. Diabetologia 2014, 57, 2282–2295. [Google Scholar] [CrossRef] [PubMed]
- Seaborne, R.A.; Sharples, A.P. The interplay between exercise metabolism, epigenetics, and skeletal muscle remodeling. Exerc. Sport Sci. Rev. 2020, 48, 188–200. [Google Scholar] [CrossRef] [PubMed]
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 2003, 133, 2485S–2493S. [Google Scholar] [CrossRef]
- Huh, J.Y. The role of exercise-induced myokines in regulating metabolism. Arch. Pharmacal. Res. 2018, 41, 14–29. [Google Scholar] [CrossRef]
- Maurer, J.; Hoene, M.; Weigert, C. Signals from the circle: Tricarboxylic acid cycle intermediates as myometabokines. Metabolites 2021, 11, 474. [Google Scholar] [CrossRef]
- Tanianskii, D.A.; Jarzebska, N.; Birkenfeld, A.L.; O’Sullivan, J.F.; Rodionov, R.N. Beta-aminoisobutyric acid as a novel regulator of carbohydrate and lipid metabolism. Nutrients 2019, 11, 524. [Google Scholar] [CrossRef]
- Iwayama, K.; Kurihara, R.; Nabekura, Y.; Kawabuchi, R.; Park, I.; Kobayashi, M.; Ogata, H.; Kayaba, M.; Satoh, M.; Tokuyama, K. Exercise increases 24-h fat oxidation only when it is performed before breakfast. EBioMedicine 2015, 2, 2003–2009. [Google Scholar] [CrossRef]
- Iwayama, K.; Kawabuchi, R.; Nabekura, Y.; Kurihara, R.; Park, I.; Kobayashi, M.; Ogata, H.; Kayaba, M.; Omi, N.; Satoh, M.; et al. Exercise before breakfast increases 24-h fat oxidation in female subjects. PLoS ONE 2017, 12, e0180472. [Google Scholar] [CrossRef]
- Guiraudou, M.; Cherif, A.; Richou, M.; Fidani, T.; Romain, A.J.; Mercier, J.; de Mauverger, E.R.; Brun, J.F. Effects over 24Hr of exercise targeted on lipid versus carbohydrate oxidation on eating behaviour in normal weight volunteers. Int. J. Sports Sci. Med. 2018, 2, 031–035. [Google Scholar]
- Brun, J.F. Exercise makes more than an energy deficit: Toward improved protocols for the management of obesity? EBioMedicine 2015, 2, 1862–1863. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Pérez-Martin, A.; Dumortier, M.; Raynaud, E.; Brun, J.F.; Fédou, C.; Bringer, J. Balance of substrate oxidation during submaximal exercise in lean and obese people. Diabetes Metab. 2001, 27, 466–474. [Google Scholar] [PubMed]
- Brun, J.F.; Myzia, J.; Varlet-Marie, E.; Mercier, J.; de Mauverger, E.R. The weight-lowering effect of low-intensity endurance training targeted at the level of maximal lipid oxidation (LIPOXmax) lasts for more than 8 years, and is associated with improvements in body composition and blood pressure. Sci. Sports, 2022; in press. [Google Scholar]
- Brun, J.F.; Jean, E.; Ghanassia, E.; Flavier, S.; Mercier, J. Metabolic training: New paradigms of exercise training for metabolic diseases with exercise calorimetry targeting individuals. Ann. Readapt. Med. Phys. 2007, 50, 528–534. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Romain, A.J.; Mercier, J. Maximal lipid oxidation during exercise (Lipoxmax): From physiological measurements to clinical applications. Facts and uncertainties. Sci. Sports 2011, 26, 57–71. [Google Scholar] [CrossRef]
- Brun, J.F.; Varlet-Marie, E.; Romain, A.J.; Mercier, J. Measurement and Physiological Relevance of the Maximal Lipid Oxidation Rate during Exercise (LIPOXmax); INTECH Open Access Publisher: London, UK, 2012; ISBN 979-953-307-096-3. [Google Scholar]
- Brun, J.F.; Malatesta, D.; Sartorio, A. Maximal lipid oxidation during exercise: A target for individualizing endurance training in obesity and diabetes? J. Endocrinol. Investig. 2012, 35, 686–691. [Google Scholar]
- Zuntz, N. Über die Bedeutung der verschiedenen Nährstoffe als Erzeuger der Muskelkraft. Arch. Die Gesamte Physiol. Menschen Tiere 1901, 83, 557–571. [Google Scholar] [CrossRef]
- Krogh, A.; Linhardt, J. The relative value of fat and carbohydrate as sources of muscular energy. Biochem. J. 1920, 14, 290–363. [Google Scholar] [CrossRef]
- Christensen, E.H.; Hansen, O. Arbeitsfähigkeit und Ernährung. Scand. Arch. Physiol. 1939, 81, 60–171. [Google Scholar] [CrossRef]
- Gibala, M.J. Protein metabolism and endurance exercise. Sports Med. 2007, 37, 337–340. [Google Scholar] [CrossRef]
- Sahlin, K.; Sallstedt, E.; Bishop, D.; Tonkonogi, M. Turning down lipid oxidation during heavy exercise—What is the mechanism? J. Physiol. Pharmacol. 2008, 59 (Suppl. S7), 19–30. [Google Scholar] [PubMed]
- Brooks, G.A.; Mercier, J. Balance of carbohydrate and lipid utilization during exercise: The “crossover” concept. J. Appl. Physiol. 1994, 76, 2253–2261. [Google Scholar] [CrossRef]
- Manetta, J.; Perez-Martin, A.; Brun, J.F.; Callis, A.; Prefaut, C.; Mercier, J. Fuel oxidation during exercise in middle-aged men: Effect of training and glucose disposal. Med. Sci. Sports Exerc. 2002, 34, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Manetta, J.; Brun, J.F.; Prefaut, C.; Mercier, J. Substrate oxidation during exercise at moderate and hard intensity in middle-aged and young athletes versus sedentary men. Metabolism 2005, 54, 1411–1419. [Google Scholar] [CrossRef]
- Aucouturier, J.; Rance, M.; Meyer, M.; Isacco, L.; Thivel, D.; Fellmann, N. Determination of the maximal fat oxidation point in obese children and adolescents: Validity of methods to assess maximal aerobic power. Eur. J. Appl. Physiol. 2009, 105, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Frayn, K.N. Calculation of substrate oxidation rates in vivo from gaseous exchange. J. Appl. Physiol. 1983, 55, 628–634. [Google Scholar] [CrossRef]
- Tappy, L.; Schneiter, P. Measurement of substrate oxidation in man. Diabetes Metab. 1997, 23, 435–442. [Google Scholar]
- Péronnet, F.; Massicotte, D. Table of nonprotein respiratory quotient: An update. Can. J. Sport Sci. 1991, 16, 23–29. [Google Scholar]
- Jeukendrup, A.E.; Wallis, G.A. Measurement of substrate oxidation during exercise by means of gas exchange measurements. Int. J. Sports Med. 2005, 26 (Suppl. S1), S28–S37. [Google Scholar] [CrossRef]
- MacRae, H.S.; Noakes, T.D.; Dennis, S.C. Role of decreased carbohydrate oxidation on slower rises in ventilation with increasing exercise intensity after training. Eur. J. Appl. Physiol. Occup. Physiol. 1995, 71, 523–529. [Google Scholar] [CrossRef]
- Bordenave, S.; Flavier, S.; Fedou, C.; Brun, J.F.; Mercier, J. Exercise calorimetry in sedentary patients: Procedures based on short 3 min steps underestimate carbohydrate oxidation and overestimate lipid oxidation. Diabetes Metab. 2007, 33, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Meyer, T.; Folz, C.; Rosenberger, F.; Kindermann, W. The reliability of fat. Scand. J. Med. Sci. Sports 2009, 19, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Croci, I.; Borrani, F.; Byrne, N.M.; Wood, R.E.; Hickman, I.J.; Chenevière, X.; Malatesta, D. Reproducibility of Fatmax and fat oxidation rates during exercise in recreationally trained males. PLoS ONE 2014, 9, e97930. [Google Scholar] [CrossRef]
- Crisp, N.A.; Guelfi, K.J.; Licari, M.K.; Braham, R.; Fournier, P.A. Does exercise duration affect Fatmax in overweight boys? Eur. J. Appl. Physiol. 2012, 112, 2557–2564. [Google Scholar] [CrossRef] [PubMed]
- Chrzanowski-Smith, O.J.; Edinburgh, R.M.; Betts, J.A.; Stokes, K.A.; Gonzalez, J.T. Evaluation of a graded exercise test to determine peak fat oxidation in individuals with low cardiorespiratory fitness. Appl. Physiol. Nutr. Metab. 2018, 43, 1288–1297. [Google Scholar] [CrossRef]
- Chenevière, X.; Malatesta, D.; Peters, E.M.; Borrani, F. A mathematical model to describe fat oxidation kinetics during graded exercise. Med. Sci. Sports Exerc. 2009, 41, 1615–1625. [Google Scholar] [CrossRef]
- Amaro-Gahete, F.J.; Sanchez-Delgado, G.; Alcantara, J.M.; Martinez-Tellez, B.; Acosta, F.M.; Helge, J.W.; Ruiz, J.R. Impact of data analysis methods for maximal fat oxidation estimation during exercise in sedentary adults. Eur. J. Sport Sci. 2019, 19, 1230–1239. [Google Scholar] [CrossRef]
- González-Haro, C.; Galilea, P.A.; González-de-Suso, J.M.; Drobnic, F.; Escanero, J.F. Maximal lipidic power in high competitive level triathletes and cyclists. Br. J. Sports Med. 2007, 41, 23–28. [Google Scholar] [CrossRef]
- Watt, M.J.; Heigenhauser, G.J.; Spriet, L.L. Effects of dynamic exercise intensity on the activation of hormone-sensitive lipase in human skeletal muscle. J. Physiol. 2003, 547 Pt 1, 301–308. [Google Scholar] [CrossRef]
- Randell, R.K.; Carter, J.M.; Jeukendrup, A.E.; Lizarraga, M.A.; Yanguas, J.I.; Rollo, I. Fat oxidation rates in professional soccer players. Med. Sci. Sports Exerc. 2019, 51, 1677–1683. [Google Scholar] [CrossRef]
- Lavault, P.; Deaux, S.; Romain, A.J.; Fédou, C.; Mercier, J.; Brun, J.F. Intérêt de la quantification de la masse musculaire pour interpréter la calorimétrie d’effort. Sci. Sports 2011, 26, 88–91. [Google Scholar] [CrossRef]
- Achten, J.; Venables, M.C.; Jeukendrup, A.E. Fat oxidation rates are higher during running compared with cycling over a wide range of intensities. Metabolism 2003, 52, 747–752. [Google Scholar] [CrossRef]
- Michallet, A.; Bricout, V.; Wuyam, B.; Guinot, M.; Favre-Juvin, A.; Lévy, P. Aspects méthodologiques de la mesure du PCGL et du Lipoxmax. Rev. Mal. Respir. 2006, 23, 396. [Google Scholar] [CrossRef]
- Gmada, N.; Marzouki, H.; Haboubi, M.; Tabka, Z.; Shephard, R.J.; Bouhlel, E. Crossover and maximal fat-oxidation points in sedentary healthy subjects: Methodological issues. Diabetes Metab. 2012, 38, 40–45. [Google Scholar] [CrossRef] [PubMed]
- De Souza Silveira, R.; Carlsohn, A.; Langen, G.; Mayer, F.; Scharhag-Rosenberger, F. Reliability and day-to-day variability of peak fat oxidation during treadmill ergometry. J. Int. Soc. Sports Nutr. 2016, 13, 4. [Google Scholar] [CrossRef]
- Chrzanowski-Smith, O.J.; Edinburgh, R.M.; Thomas, M.P.; Haralabidis, N.; Williams, S.; Betts, J.A.; Gonzalez, J.T. The day-to-day reliability of peak fat oxidation and FATMAX. Eur. J. Appl. Physiol. 2020, 120, 1745–1759. [Google Scholar] [CrossRef]
- Marzouki, H.; Farhani, Z.; Bouhlel, E. Relative and absolute reliability of the crossover and maximum fat oxidation points during treadmill running. Sci. Sports 2014, 29, e107–e114. [Google Scholar] [CrossRef]
- Brun, J.F.; Romain, A.J.; Guiraudou, M.; Fédou, C.; Mercier, J. Stability over time of the level of maximal lipid oxidation during exercise (Stabilité dans le temps du niveau maximal d’oxydation des lipides à l’exercice). Sci. Sports 2014, 29, 168–171. [Google Scholar] [CrossRef]
- Brooks, G.A. Importance of the ‘crossover’ concept in exercise metabolism. Clin. Exp. Pharmacol. Physiol. 1997, 24, 889–895. [Google Scholar] [CrossRef]
- Brooks, G.A. The Precious Few Grams of Glucose During Exercise. Int. J. Mol. Sci. 2020, 21, 5733. [Google Scholar] [CrossRef]
- Aloulou, I. Le coût glucidique du watt sur ergocycle: Une constante biologique? Carbohydrate cost of the watt on ergocycle: A reproducible biological parameter? Sci. Sport 2002, 17, 315–317. [Google Scholar] [CrossRef]
- Brun, J.F.; Dumortier, M.; Fedou, C.; Mercier, J. Exercise hypoglycemia in nondiabetic subjects. Diabetes Metab. 2001, 27 Pt 1, 92–106. [Google Scholar] [PubMed]
- Jean, E.; Flavier, S.; Mercier, J.; Brun, J.F. Exercise calorimetry with 6 min steps closely predicts the lipid oxidation flow rate of a 45 min steady state targeted training session at the level of maximal lipid oxidation (LIPOXmax). Fund. Clin. Pharmacol. 2007, 21, 95. [Google Scholar]
- Meyer, T.; Gässler, N.; Kindermann, W. Determination of “Fatmax” with 1 h cycling protocols of constant load. Appl. Physiol. Nutr. Metab. 2007, 32, 249–256. [Google Scholar] [CrossRef]
- Özdemir, Ç.; Özgünen, K.; Günaştı, Ö.; Eryılmaz, S.K.; Kılcı, A.; Kurdak, S.S. Changes in substrate utilization rates during 40 min of walking within the Fatmax range. Physiol. Int. 2019, 106, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Dasilva, S.G.; Guidetti, L.; Buzzachera, C.F.; Elsangedy, H.M.; Krinski, K.; De Campos, W.; Goss, F.L.; Baldari, C. Gender-based differences in substrate use during exercise at a self-selected pace. J. Strength Cond. Res. 2011, 25, 2544–2551. [Google Scholar] [CrossRef]
- Brun, J.F.; Romain, A.J.; Pollatz, M.; Chevance, G.; Fédou, C.; Mercier, J. L’intensité spontanément sélectionnée pour un exercice prolongé se situe dans la zone d’oxydation lipidique maximale. Nutr. Clin. Metab. 2013, 27 (Suppl. S1), S106–S107. [Google Scholar]
- Brun, J.F.; Guiraudou, M.; Romain, A.J.; Pollatz, M.; Fédou, C.; Mercier, J. Le niveau d’oxydation maximale des lipides (LIPOXmax), un niveau d’exercice adapté par ses propriétés énergétiques et comportementales au «génotype économe» du syndrome métabolique: L’hypothèse du «healthy primitive lifestyle». Sci. Sports 2015, 30, 74–81. [Google Scholar] [CrossRef]
- Neel, J.V. Diabetes mellitus: A “thrifty” genotype rendered detrimental by “progress”? Am. J. Hum. Genet. 1962, 14, 353–362. [Google Scholar]
- Chakravarthy, M.V.; Booth, F.W. Eating, exercise, and “thrifty” genotypes: Connecting the dots toward an evolutionary understanding of modern chronic diseases. J. Appl. Physiol. 2004, 96, 3–10. [Google Scholar] [CrossRef]
- O’Keefe, J.H.; Vogel, R.; Lavie, C.J.; Cordain, L. Exercise like a hunter-gatherer: A prescription for organic physical fitness. Prog. Cardiovasc. Dis. 2011, 53, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Curtis, J.P.; Selter, J.G.; Wang, Y.; Rathore, S.S.; Jovin, I.S.; Jadbabaie, F.; Kosiborod, M.; Portnay, E.L.; Sokol, S.I.; Bader, F.; et al. The obesity paradox: Body mass index and outcomes in patients with heart failure. Arch. Intern. Med. 2005, 165, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Macpherson, J.; Gray, S.R.; Gill, J.M.; Welsh, P.; Celis-Morales, C.; Sattar, N.; Pell, J.P.; Ho, F.K. Are people with metabolically healthy obesity really healthy? A prospective cohort study of 381,363 UK Biobank participants. Diabetologia 2021, 64, 1963–1972. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Varlet-Marie, E.; Chevance, G.; Pollatz, M.; Fedou, C.; de Mauverger, E.R. Versatility of ‘hemorheologic fitness’ according to exercise intensity: Emphasis on the “healthy primitive lifestyle”. Korea-Aust. Rheol. J. 2014, 26, 249–253. [Google Scholar] [CrossRef]
- Bordenave, S.; Metz, L.; Flavier, S.; Lambert, K.; Ghanassia, E.; Dupuy, A. Training-induced improvement in lipid oxidation in type 2 diabetes mellitus is related to alterations in muscle mitochondrial activity. Effect of endurance training in type 2 diabetes. Diabetes Metab. 2008, 34, 162–168. [Google Scholar] [CrossRef]
- Nordby, P.; Saltin, B.; Helge, J.W. Whole-body fat oxidation determined by graded exercise and indirect calorimetry: A role for muscle oxidative capacity? Scand. J. Med. Sci. Sports 2006, 16, 209–214. [Google Scholar] [CrossRef]
- Rosenkilde, M.; Reichkendler, M.H.; Auerbach, P.; Bonne, T.C.; Sjödin, A.; Ploug, T.; Stallknecht, B.M. Changes in peak fat oxidation in response to different doses of endurance training. Scand. J. Med. Sci. Sports 2015, 25, 41–52. [Google Scholar] [CrossRef]
- Maunder, E.; Plews, D.J.; Wallis, G.A.; Brick, M.J.; Leigh, W.B.; Chang, W.L.; Stewart, T.; Watkins, C.M.; Kilding, A.E. Peak fat oxidation is positively associated with vastus lateralis CD36 content, fed-state exercise fat oxidation, and endurance performance in trained males. Eur. J. Appl. Physiol. 2022, 122, 93–102. [Google Scholar] [CrossRef]
- Sahlin, K.; Mogensen, M.; Bagger, M.; Fernström, M.; Pedersen, P.K. The potential for mitochondrial fat oxidation in human skeletal muscle influences whole body fat oxidation during low-intensity exercise. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E223–E230. [Google Scholar] [CrossRef][Green Version]
- Daussin, F.N.; Zoll, J.; Ponsot, E.; Dufour, S.P.; Doutreleau, S.; Lonsdorfer, E.; Ventura-Clapier, R.; Mettauer, B.; Piquard, F.; Geny, B.; et al. Training at high exercise intensity promotes qualitative adaptations of mitochondrial function in human skeletal muscle. J. Appl. Physiol. 2008, 104, 1436–1441. [Google Scholar] [CrossRef]
- Kelley, D.E. Skeletal muscle fat oxidation: Timing and flexibility are everything. J. Clin. Investig. 2005, 115, 1699–1702. [Google Scholar] [CrossRef] [PubMed]
- Galgani, J.E.; Moro, C.; Ravussin, E. Metabolic flexibility and insulin resistance. Am. J. Physiol. Endocrinol. Metab. 2008, 295, E1009–E1017. [Google Scholar] [CrossRef] [PubMed]
- Galgani, J.E.; Fernández-Verdejo, R. Pathophysiological role of metabolic flexibility on metabolic health. Obes. Rev. 2021, 22, e13131. [Google Scholar] [CrossRef]
- Corpeleijn, E.; Saris, W.H.; Blaak, E.E. Metabolic flexibility in the development of insulin resistance and type 2 diabetes: Effects of lifestyle. Obes. Rev. 2009, 10, 178–193. [Google Scholar] [CrossRef]
- San-Millán, I.; Brooks, G.A. Assessment of metabolic flexibility by means of measuring blood lactate, fat, and carbohydrate oxidation responses to exercise in professional endurance athletes and less-fit individuals. Sports Med. 2018, 48, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Moro, C.; Pillard, F.; de Glisezinski, I.; Crampes, F.; Thalamas, C.; Harant, I.; Marques, M.A.; Lafontan, M.; Berlan, M. Sex differences in lipolysis-regulating mechanisms in overweight subjects: Effect of exercise intensity. Obesity 2007, 15, 2245–2255. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, S.; Codecasa, F.; Cornacchia, M.; Maestrini, S.; Salvadori, A.; Brunani, A.; Malatesta, D. Fat oxidation, hormonal and plasma metabolite kinetics during a submaximal incremental test in lean and obese adults. PLoS ONE 2014, 9, e88707. [Google Scholar] [CrossRef]
- Starritt, E.C.; Howlett, R.A.; Heigenhauser, G.J.; Spriet, L.L. Sensitivity of CPT I to malonyl-CoA in trained and untrained human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E462–E468. [Google Scholar] [CrossRef]
- Coyle, E.F.; Jeukendrup, A.E.; Wagenmakers, A.J.; Saris, W.H. Fatty acid oxidation is directly regulated by carbohydrate metabolism during exercise. Am. J. Physiol. 1997, 273, E268–E275. [Google Scholar] [CrossRef]
- Aïssa Benhaddad, A.; Pérez-Martin, A.; Brun, J.F.; Fédou, C.; Raynaud, E.; Mercier, J. Concordance entre le seuil ventilatoire et le “point de cross-over” chez des patients en surpoids présentant des troubles de la glycorégulation. Diabetes Metab. 1999, 25 (Suppl. S1), 39. [Google Scholar]
- Ceugniet, F.; Pérez-Martin, A.; Brun, J.F.; Mercier, J.; Préfaut, C. Comparison between ventilatory threshold and crossover point of substrate utilization during exercise in obese patients with OSAS. Eur. J. Physiol. 2001, 442, R130. [Google Scholar]
- Astorino, T.A. Is the ventilatory threshold coincident with maximal fat oxidation during submaximal exercise in women? J. Sports Med. Phys. Fit. 2000, 40, 209–216. [Google Scholar]
- Peric, R.; Meucci, M.; Bourdon, P.C.; Nikolovski, Z. Does the aerobic threshold correlate with the maximal fat oxidation rate in short stage treadmill tests? J. Sports Med. Phys. Fit. 2018, 58, 1412–1417. [Google Scholar] [CrossRef] [PubMed]
- Emerenziani, G.P.; Ferrari, D.; Marocco, C.; Greco, E.A.; Migliaccio, S.; Lenzi, A.; Baldari, C.; Guidetti, L. Relationship between individual ventilatory threshold and maximal fat oxidation (MFO) over different obesity classes in women. PLoS ONE 2019, 14, e0215307. [Google Scholar] [CrossRef] [PubMed]
- Tolfrey, K.; Jeukendrup, A.E.; Batterham, A.M. Group- and individual-level coincidence of the ‘Fatmax’ and lactate accumulation in adolescents. Eur. J. Appl. Physiol. 2010, 109, 1145–1153. [Google Scholar] [CrossRef]
- Metz, L.; Sirvent, P.; Py, G.; Brun, J.F.; Fédou, C.; Raynaud, E.; Mercier, J. Relationship between blood lactate concentration and substrate utilization during exercise in type 2 diabetic postmenopausal women. Metabolism 2005, 54, 1102–1107. [Google Scholar] [CrossRef]
- Poole, D.C.; Rossiter, H.B.; Brooks, G.A.; Gladden, L.B. The anaerobic threshold: 50+ years of controversy. J. Physiol. 2021, 599, 737–767. [Google Scholar] [CrossRef]
- Makni, E.; Moalla, W.; Trabelsi, Y.; Lac, G.; Brun, J.F.; Tabka, Z.; Elloumi, M. Six-minute walking test predicts maximal fat oxidation in obese children. Int. J. Obes. 2012, 36, 908–913. [Google Scholar] [CrossRef]
- Purdom, T.; Kravitz, L.; Dokladny, K.; Mermier, C. Understanding the factors that effect maximal fat oxidation. J. Int. Soc. Sports Nutr. 2018, 15, 3. [Google Scholar] [CrossRef]
- Jeukendrup, A.E. Modulation of carbohydrate and fat utilization by diet, exercise and environment. Biochem. Soc. Trans. 2003, 31, 1270–1273. [Google Scholar] [CrossRef]
- Dumortier, M.; Thöni, G.; Brun, J.F.; Mercier, J. Substrate oxidation during exercise: Impact of time interval from the last meal in obese women. Int. J. Obes. 2005, 29, 966–974. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Attalin, V.; Romain, J.; Desplan, M.; Carayol, M.; Ninot, G.; Fédou, C.; Brun, J.F. Effet d’un petit déjeuner sur le LIPOXmax. Diabetes Metab. 2011, 37, A19. [Google Scholar]
- Fletcher, G.; Eves, F.F.; Glover, E.I.; Robinson, S.L.; Vernooij, C.A.; Thompson, J.L.; Wallis, G.A. Dietary intake is independently associated with the maximal capacity for fat oxidation during exercise. Am. J. Clin. Nutr. 2017, 105, 864–872. [Google Scholar] [CrossRef] [PubMed]
- Roberts, J.D.; Willmott, A.G.; Beasley, L.; Boal, M.; Davies, R.; Martin, L.; Chichger, H.; Gautam, L.; Del Coso, J. The impact of decaffeinated green tea extract on fat oxidation, body composition and cardio-metabolic health in overweight, recreationally active individuals. Nutrients 2021, 13, 764. [Google Scholar] [CrossRef]
- Blicher, S.; Bartholomae, E.; Kressler, J. Effects of chronic decaffeinated green tea extract supplementation on lipolysis and substrate utilization during upper body exercise. J. Sport Health Sci. 2021, 10, 237–242. [Google Scholar] [CrossRef]
- Desai, T.; Bottoms, L.; Roberts, M. The effects of Montmorency tart cherry juice supplementation and FATMAX exercise on fat oxidation rates and cardio-metabolic markers in healthy humans. Eur. J. Appl. Physiol. 2018, 118, 2523–2539. [Google Scholar] [CrossRef]
- Coyle, E.F.; Jeukendrup, A.E.; Oseto, M.C.; Hodgkinson, B.J.; Zderic, T.W. Low-fat diet alters intramuscular substrates and reduces lipolysis and fat oxidation during exercise. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E391. [Google Scholar] [CrossRef]
- Chenevière, X.; Borrani, F.; Ebenegger, V.; Gojanovic, B.; Malatesta, D. Effect of a 1-hour single bout of moderate-intensity exercise on fat oxidation kinetics. Metabolism 2009, 58, 1778–1786. [Google Scholar] [CrossRef]
- Brandou, F.; Savy-Pacaux, A.M.; Marie, J.; Brun, J.F.; Mercier, J. Comparison of the type of substrate oxidation during exercise between pre and post pubertal markedly obese boys. Int. J. Sports Med. 2006, 27, 407–414. [Google Scholar] [CrossRef]
- Riddell, M.C.; Jamnik, V.K.; Iscoe, K.E.; Timmons, B.W.; Gledhill, N. Fat oxidation rate and the exercise intensity that elicits maximal fat oxidation decreases with pubertal status in young male subjects. J. Appl. Physiol. 2008, 105, 742–748. [Google Scholar] [CrossRef]
- Zakrzewski, J.K.; Tolfrey, K. Comparison of fat oxidation over a range of intensities during treadmill and cycling exercise in children. Eur. J. Appl. Physiol. 2011, 112, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Aucouturier, J.; Baker, J.S.; Duché, P. Fat and carbohydrate metabolism during submaximal exercise in children. Sports Med. 2008, 38, 213–238. [Google Scholar] [CrossRef] [PubMed]
- Bouhlel, E.; Salhi, Z.; Bouhlel, H.; Mdella, S.; Amamou, A.; Zaouali, M.; Mercier, J.; Bigard, X.; Tabka, Z.; Zbidi, A.; et al. Effect of Ramadan fasting on fuel oxidation during exercise in trained male rugby players. Diabetes Metab. 2006, 32, 617–624. [Google Scholar] [CrossRef]
- Friedlander, A.L.; Casazza, G.A.; Horning, M.A.; Huie, M.J.; Piacentini, M.F.; Trimmer, J.K. Training-induced alterations of carbohydrate metabolism in women: Women respond differently from men. J. Appl. Physiol. 1998, 85, 1175–1186. [Google Scholar] [CrossRef]
- Frandsen, J.; Amaro-Gahete, F.J.; Landgrebe, A.; Dela, F.; Ruiz, J.R.; Helge, J.W.; Larsen, S. The influence of age, sex and cardiorespiratory fitness on maximal fat oxidation rate. Appl. Physiol. Nutr. Metab. 2021, 46, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Chenevière, X.; Borrani, F.; Sangsue, D.; Gojanovic, B.; Malatesta, D. Gender differences in whole-body fat oxidation kinetics during exercise. Appl. Physiol. Nutr. Metab. 2011, 36, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Boegner, C.; Raynaud, E.; Mercier, J. Contrairement à une idée reçue, les femmes n’oxydent pas plus de lipides à l’effort que les hommes, mais leur LIPOXmax survient à une puissance plus élevée. Sci. Sports 2009, 24, 45–48. [Google Scholar] [CrossRef]
- Frandsen, J.; Hansen, I.M.D.; Wismann, J.F.; Olsen, M.H.; Brage-Andersen, M.R.; Sahl, R.E.; Hansen, M.; Ingersen, A.; Modvig, J.L.; Schmücker, M.; et al. Maximal fat oxidation rate is higher in fit women and unfit women with obesity, compared to normal-weight unfit women. J. Clin. Endocrinol. Metab. 2021, 106, e4389–e4399. [Google Scholar] [CrossRef]
- Isacco, L.; Duché, P.; Boisseau, N. Influence of hormonal status on substrate utilization at rest and during exercise in the female population. Sports Med. 2012, 42, 327–342. [Google Scholar] [CrossRef]
- Boisseau, N.; Isacco, L. Substrate metabolism during exercise: Sexual dimorphism and women’s specificities. Eur. J. Sport Sci. 2021, 17, 1–21. [Google Scholar] [CrossRef]
- Willett, H.N.; Koltun, K.J.; Hackney, A.C. Influence of menstrual cycle estradiol-β-17 fluctuations on energy substrate utilization-oxidation during aerobic, endurance exercise. Int. J. Environ. Res. Public Health 2021, 18, 7209. [Google Scholar] [CrossRef] [PubMed]
- Febbraio, M.A.; Snow, R.J.; Hargreaves, M.; Stathis, C.G.; Martin, I.K.; Carey, M.F. Muscle metabolism during exercise and heat stress in trained men: Effect of acclimation. J. Appl. Physiol. 1994, 76, 589–597. [Google Scholar] [CrossRef] [PubMed]
- Del Coso, J.; Hamouti, N.; Ortega, J.F.; Mora-Rodriguez, R. Aerobic fitness determines whole-body fat oxidation rate during exercise in the heat. Appl. Physiol. Nutr. Metab. 2010, 35, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Jeukendrup, A.E. Maximal fat oxidation during exercise in trained men. Int. J. Sports Med. 2003, 24, 603–608. [Google Scholar]
- Venables, M.C.; Achten, J.; Jeukendrup, A.E. Determinants of fat oxidation during exercise in healthy men and women: A cross-sectional study. J. Appl. Physiol. 2005, 98, 160–167. [Google Scholar] [CrossRef]
- Varlet-Marie, E.; Brun, J.F.; Fedou, C.; Mercier, J. Balance of substrates used for oxidation at exercise in ahletes: Lipodependent vs glucodependent sports. Fund. Clin. Pharmacol. 2006, 20, 220. [Google Scholar]
- Sardinoux, M.; Brun, J.F.; Lefebvre, P.; Bringer, J.; Fabre, G.; Salsano, V. Influence of bariatric surgery on exercice maximal lipid oxydation point in grade 3 obese patients. Fund. Clin. Pharmacol. 2009, 23, 57. [Google Scholar]
- Griffiths, A.; Shannon, O.M.; Matu, J.; King, R.; Deighton, K.; O’Hara, J.P. The effects of environmental hypoxia on substrate utilisation during exercise: A meta-analysis. J. Int. Soc. Sports Nutr. 2019, 16, 10. [Google Scholar] [CrossRef]
- Montes-de-Oca-García, A.; Perez-Bey, A.; Corral-Pérez, J.; Velázquez-Díaz, D.; Opazo-Díaz, E.; Fernandez-Santos, J.R.; Rebollo-Ramos, M.; Amaro-Gahete, F.J.; Cuenca-García, M.; Ponce-González, J.G. Maximal fat oxidation capacity is associated with cardiometabolic risk factors in healthy young adults. Eur. J. Sport Sci. 2021, 21, 907–917. [Google Scholar] [CrossRef]
- Amaro-Gahete, F.J.; Sanchez-Delgado, G.; Jurado-Fasoli, L.; Ruiz, J.R. Uncertain association between maximal fat oxidation during exercise and cardiometabolic risk factors in healthy sedentary adults. Eur. J. Sport Sci. 2021, 22, 1–11. [Google Scholar] [CrossRef]
- Amaro-Gahete, F.J.; Acosta, F.M.; Migueles, J.H.; Ponce González, J.G.; Ruiz, J.R. Association of sedentary and physical activity time with maximal fat oxidation during exercise in sedentary adults. Scand. J. Med. Sci. Sports 2020, 30, 1605–1614. [Google Scholar] [CrossRef] [PubMed]
- Croci, I.; Hickman, I.J.; Wood, R.E.; Borrani, F.; Macdonald, G.A.; Byrne, N.M. Fat oxidation over a range of exercise intensities: Fitness versus fatness. Appl. Physiol. Nutr. Metab. 2014, 39, 1352–1359. [Google Scholar] [CrossRef] [PubMed]
- Malin, S.K.; Stephens, B.R.; Sharoff, C.G.; Hagobian, T.A.; Chipkin, S.R.; Braun, B. Metformin’s effect on exercise and postexercise substrate oxidation. Int. J. Sport Nutr. Exerc. Metab. 2010, 20, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Ghanassia, E.; Brun, J.F.; Fedou, C.; Raynaud, E.; Mercier, J. Substrate oxidation during exercise: Type 2 diabetes is associated with a decrease in lipid oxidation and an earlier shift towards carbohydrate utilization. Diabetes Metab. 2006, 32, 604–610. [Google Scholar] [CrossRef]
- Mogensen, M.; Vind, B.F.; Højlund, K.; Beck-Nielsen, H.; Sahlin, K. Maximal lipid oxidation in patients with type 2 diabetes is normal and shows an adequate increase in response to aerobic training. Diabetes Obes. Metab. 2009, 11, 874–883. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Fedou, C.; Grubka, E.; Karafiat, M.; Varlet-Marie, E.; Mercier, J. Moindre utilisation des lipides à l’exercice chez le diabétique de type 1. Sci. Sports 2008, 23, 198–200. [Google Scholar] [CrossRef]
- Fel, S.; Rochette, E.; Walther, G.; Echaubard, S.; Pereira, B.; Merlin, E.; Terral, D.; Duché, P. Maximal fat oxidation during exercise is already impaired in pre-pubescent children with type 1 diabetes mellitus. Front. Physiol. 2021, 12, 664211. [Google Scholar] [CrossRef]
- Desplan, M.; Avignon, A.; Mestejanot, C.; Outters, M.; Sardinoux, M.; Fedou, C. Impaired ability to oxidize lipids at exercise in severe vs. mild to moderate sleep apnea/hypopnea obstructive sleep syndrome (OSAS). Fund. Clin. Pharmacol. 2010, 24, 66. [Google Scholar]
- Frandsen, J.; Poggi, A.I.; Ritz, C.; Larsen, S.; Dela, F.; Helge, J.W. Peak fat oxidation rate is closely associated with plasma free fatty acid concentrations in women; similar to men. Front. Physiol. 2021, 12, 696261. [Google Scholar] [CrossRef]
- Soenen, S.; Plasqui, G.; Smeets, A.J.; Westerterp-Plantenga, M.S. Protein intake induced an increase in exercise stimulated fat oxidation during stable body weight. Physiol. Behav. 2010, 101, 770–774. [Google Scholar] [CrossRef]
- Soriano, E.; Pascual, O.; Fédou, C.; Brun, J.F. Oxydation des substrats énergétiques à l’effort dans l’anorexie mentale. Diabetes Metab. 2011, 37, A36. [Google Scholar] [CrossRef]
- Brun, J.F.; Hammoudi, L.; Gimet, F.; Noirez, P.; de Mauverger, E.R.; Mercier, J. Oxydation des lipides au cours de l’exercice: Influence du sexe, de la corpulence et de l’âge à partir d’une base de données de 5258 calorimetries d’effort. Nutr. Clin. Métabolisme 2020, 34, 38. [Google Scholar] [CrossRef]
- Swain, D.P.; Leutholtz, B.C.; King, M.E.; Haas, L.A.; Branch, J.D. Relationship between % heart rate reserve and % VO2 reserve in treadmill exercise. Med. Sci. Sports Exerc. 1998, 30, 318–321. [Google Scholar] [CrossRef] [PubMed]
- Swain, D.P.; Leutholtz, B.C. Heart rate reserve is equivalent to %VO2 reserve, not to %VO2max. Med. Sci. Sports Exerc. 1997, 29, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, J.E.; Rottensteiner, M.; Wiklund, P.; Hämäläinen, K.; Laakkonen, E.K.; Kaprio, J.; Kainulainen, H.; Kujala, U.M. Fat oxidation at rest and during exercise in male monozygotic twins. Eur. J. Appl. Physiol. 2019, 119, 2711–2722. [Google Scholar] [CrossRef]
- Bellou, V.; Belbasis, L.; Tzoulaki, I.; Evangelou, E. Risk factors for type 2 diabetes mellitus: An exposure-wide umbrella review of meta-analyses. PLoS ONE 2018, 13, e0194127. [Google Scholar] [CrossRef]
- Ahlborg, G.; Felig, P.; Hagenfeldt, L.; Hendler, R.; Wahren, J. Substrate turnover during prolonged exercise in man: Splanchnic and leg metabolism of glucose, free fatty acids, and amino acids. J. Clin. Investig. 1974, 53, 1080–1090. [Google Scholar] [CrossRef]
- Watt, M.J.; Steinberg, G.R.; Heigenhauser, G.J.; Spriet, L.L.; Dyck, D.J. Hormone-sensitive lipase activity and triacylglycerol hydrolysis are decreased in rat soleus muscle by cyclopiazonic acid. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E412–E419. [Google Scholar] [CrossRef]
- Coyle, E.F. Substrate utilization during exercise in active people. Am. J. Clin. Nutr. 1995, 61 (Suppl. S4), 968S–979S. [Google Scholar] [CrossRef]
- Coyle, E.F.; Coggan, A.R.; Hemmert, M.K.; Ivy, J.L. Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. Appl. Physiol. 1986, 61, 165–172. [Google Scholar] [CrossRef]
- Coggan, A.R.; Coyle, E.F. Carbohydrate ingestion during prolonged exercise: Effects on metabolism and performance. Exerc. Sport Sci. Rev. 1991, 19, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Bensimhon, D.R.; Kraus, W.E.; Donahue, M.P. Obesity and physical activity: A review. Am. Heart J. 2006, 151, 598–603. [Google Scholar] [CrossRef] [PubMed]
- Melanson, E.L.; Sharp, T.A.; Seagle, H.M.; Horton, T.J.; Donahoo, W.T.; Grunwald, G.K. Effect of exercise intensity on 24-h energy expenditure and nutrient oxidation. J. Appl. Physiol. 2002, 92, 1045–1052. [Google Scholar] [CrossRef] [PubMed]
- Folch, N.; Péronnet, F.; Massicotte, D.; Duclos, M.; Lavoie, C.; Hillaire-Marcel, C. Metabolic response to small and large 13C-labelled pasta meals following rest or exercise in man. Br. J. Nutr. 2001, 85, 671–680. [Google Scholar] [CrossRef]
- Malatesta, D.; Werlen, C.; Bulfaro, S.; Chenevière, X.; Borrani, F. Effect of high-intensity interval exercise on lipid oxidation during postexercise recovery. Med. Sci. Sports Exerc. 2009, 41, 364–374. [Google Scholar] [CrossRef]
- Lazzer, S.; Lafortuna, C.; Busti, C.; Galli, R.; Tinozzi, T.; Agosti, F. Fat oxidation rate during and after a low- or high-intensity exercise in severely obese Caucasian adolescents. Eur. J. Appl. Physiol. 2010, 108, 383–391. [Google Scholar] [CrossRef]
- Henderson, G.C.; Fattor, J.A.; Horning, M.A.; Faghihnia, N.; Johnson, M.L.; Mau, T.L.; Luke-Zeitoun, M.; Brooks, G.A. Lipolysis and fatty acid metabolism in men and women during the postexercise recovery period. J. Physiol. 2007, 584 Pt 3, 963–981. [Google Scholar] [CrossRef]
- Kuo, C.C.; Fattor, J.A.; Henderson, G.C.; Brooks, G.A. Lipid oxidation in fit young adults during postexercise recovery. J. Appl. Physiol. 2005, 99, 349–356. [Google Scholar] [CrossRef]
- Warren, A.; Howden, E.J.; Williams, A.D.; Fell, J.W.; Johnson, N.A. Postexercise fat oxidation: Effect of exercise duration, intensity, and modality. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 607–623. [Google Scholar] [CrossRef]
- Talanian, J.L.; Galloway, S.D.; Heigenhauser, G.J.; Bonen, A.; Spriet, L.L. Two weeks of high-intensity aerobic interval training increases the capacity for fat oxidation during exercise in women. J. Appl. Physiol. 2007, 102, 1439–1447. [Google Scholar] [CrossRef]
- Barwell, N.D.; Malkova, D.; Leggate, M.; Gill, J.M. Individual responsiveness to exercise-induced fat loss is associated with change in resting substrate utilization. Metabolism 2009, 58, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Iwayama, K.; Ogawa, A.; Tanaka, Y.; Yajima, K.; Park, I.; Ando, A.; Ogata, H.; Kayaba, M.; Zhang, S.; Tanji, F.; et al. Effects of exercise before breakfast on plasma free fatty acid profile and 24-h fat oxidation. Metab. Open 2020, 8, 100067. [Google Scholar] [CrossRef] [PubMed]
- Frampton, J.; Edinburgh, R.M.; Ogden, H.B.; Gonzalez, J.T.; Chambers, E.S. The acute effect of fasted exercise on energy intake, energy expenditure, subjective hunger and gastrointestinal hormone release compared to fed exercise in healthy individuals: A systematic review and network meta-analysis. Int. J. Obes. 2022, 46, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Fédou, C.; Chaze, D.; Pérez-Martin, A.; Lumbroso, S.; Sultan, C.; Mercier, J. Relations entre les proportions respectives de lipides et de glucides oxydés à différents niveaux d’exercice et l’axe GH-IGF-I-IGFBPs chez des sportifs entraînés. Ann. Endocrinol. 1999, 60, 366. [Google Scholar]
- Randell, R.K.; Rollo, I.; Roberts, T.J.; Dalrymple, K.J.; Jeukendrup, A.E.; Carter, J.M. Maximal fat oxidation rates in an athletic population. Med. Sci. Sports Exerc. 2017, 49, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Coquart, J.B.; Grosbois, J.M.; Nycz, M.; Bart, F.; Garcin, M. Influence du niveau de performance sur le seuil d’oxydation maximale des lipides (Lipoxmax) chez des cyclistes. Sci. Sports 2011, 26, 32–37. [Google Scholar] [CrossRef]
- Chenevière, X.; Malatesta, D.; Gojanovic, B.; Borrani, F. Differences in whole-body fat oxidation kinetics between cycling and running. Eur. J. Appl. Physiol. 2010, 109, 1037–1045. [Google Scholar] [CrossRef]
- King, L.; Sillers, W.; McCarthy, K.; Louis, P.; Astorino, T.A. Higher fat oxidation during treadmill walking versus cycle ergometry in active women at equal RPE: A pilot study. J. Sports Med. Phys. Fit. 2016, 56, 1298–1303. [Google Scholar]
- Mendelson, M.; Jinwala, K.; Wuyam, B.; Levy, P.; Flore, P. Can crossover and maximal fat oxidation rate points be used equally for ergocycling and walking/running on a track? Diabetes Metab. 2012, 38, 264–270. [Google Scholar] [CrossRef]
- Amaro-Gahete, F.J.; Sanchez-Delgado, G.; Ruiz, J.R. Commentary: Contextualising maximal fat oxidation during exercise: Determinants and normative values. Front. Physiol. 2018, 9, 1460. [Google Scholar] [CrossRef]
- Zunquin, G.; Theunynck, D.; Sesboüé, B.; Arhan, P.; Bougle, D. Evolution of fat oxidation during exercise in obese pubertal boys: Clinical implications. J. Sports Sci. 2009, 27, 315–318. [Google Scholar] [CrossRef] [PubMed]
- Zunquin, G.; Theunynck, D.; Sesboüé, B.; Arhan, P.; Bougle, D. Comparison of fat oxidation during exercise in lean and obese pubertal boys: Clinical implications. Br. J. Sports Med. 2009, 43, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Helge, J.W.; Wu, B.J.; Willer, M.; Daugaard, J.R.; Storlien, L.H.; Kiens, B. Training affects muscle phospholipid fatty acid composition in humans. J. Appl. Physiol. 2001, 90, 670–677. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Whitfield, J.; Heikura, I.A.; Ross, M.L.R.; Tee, N.; Forbes, S.F.; Hall, R.; McKay, A.K.A.; Wallett, A.M.; Sharma, A.P. Adaptation to a low carbohydrate high fat diet is rapid but impairs endurance exercise metabolism and performance despite enhanced glycogen availability. J. Physiol. 2021, 599, 771–790. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M. Re-examining high-fat diets for sports performance: Did we call the ‘nail in the coffin’ too soon? Sports Med. 2015, 45 (Suppl. S1), S33–S49. [Google Scholar] [CrossRef]
- Storlien, L.H.; Pan, D.A.; Kriketos, A.D.; Baur, L.A. High fat diet-induced insulin resistance. Lessons and implications from animal studies. Ann. N. Y. Acad. Sci. 1993, 683, 82–90. [Google Scholar] [CrossRef]
- Bergman, R.N.; Ader, M. Free fatty acids and pathogenesis of type 2 diabetes mellitus. Trends Endocrinol. Metab. 2000, 11, 351–356. [Google Scholar] [CrossRef]
- Kim, S.P.; Ellmerer, M.; Van Citters, G.W.; Bergman, R.N. Primacy of hepatic insulin resistance in the development of the metabolic syndrome induced by an isocaloric moderate-fat diet in the dog. Diabetes 2003, 52, 2453–2460. [Google Scholar] [CrossRef]
- Pereira, S.; Breen, D.M.; Naassan, A.E.; Wang, P.Y.; Uchino, H.; Fantus, I.G.; Carpentier, A.C.; Gutierrez-Juarez, R.; Brindley, D.N.; Lam, T.K.; et al. In vivo effects of polyunsaturated, monounsaturated, and saturated fatty acids on hepatic and peripheral insulin sensitivity. Metabolism 2015, 64, 315–322. [Google Scholar] [CrossRef]
- Drapier, E.; Brun, J.F.; Fédou, C.; de Mauverger, E.R. Synergie entre les effets amaigrissants de l’activité musculaire ciblée au LIPOXmax et d’un enrichissement modéré en protéines du blanc d’œuf. Sci. Sports 2016, 31, 103–106. [Google Scholar] [CrossRef]
- Kjaer, M.; Secher, N.H.; Galbo, H. Physical stress and catecholamine release. Baillieres Clin. Endocrinol. Metab. 1987, 1, 279–298. [Google Scholar] [CrossRef]
- Molinoff, P.B. Alpha-and beta-adrenergic receptor subtypes properties, distribution and regulation. Drugs 1984, 28 (Suppl. S2), 1–15. [Google Scholar] [CrossRef] [PubMed]
- DeFronzo, R.A.; Thorin, D.; Felber, J.P.; Simonson, D.C.; Thiebaud, D.; Jequier, E.; Golay, A. Effect of beta and alpha adrenergic blockade on glucose-induced thermogenesis in man. J. Clin. Investig. 1984, 73, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Kim, H.S.; Seo, H.S.; Na, J.O.; Jang, Y.N.; Han, Y.M.; Kim, H.M. Stimulation of alpha1-adrenergic receptor ameliorates cellular functions of multiorgans beyond vasomotion through PPARδ. PPAR Res. 2020, 2020, 3785137. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.; Papay, R.S.; Perez, D.M. The role of α1-adrenergic receptors in regulating metabolism: Increased glucose tolerance, leptin secretion and lipid oxidation. J. Recept. Signal Transduct. 2017, 37, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Hargreaves, M. Effect of epinephrine on glucose disposal during exercise in humans: Role of muscle glycogen. Am. J. Physiol. Endocrinol. Metab. 2002, 283, E578–E583. [Google Scholar] [CrossRef][Green Version]
- Djurhuus, C.B.; Gravholt, C.H.; Nielsen, S.; Pedersen, S.B.; Møller, N.; Schmitz, O. Additive effects of cortisol and growth hormone on regional and systemic lipolysis in humans. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E488–E494. [Google Scholar] [CrossRef]
- Richter, W.O.; Naudé, R.J.; Oelofsen, W.; Schwandt, P. In vitro lipolytic activity of beta-endorphin and its partial sequences. Endocrinology 1987, 120, 1472–1476. [Google Scholar] [CrossRef]
- Møller, N.; Jørgensen, J.O.; Schmitz, O.; Møller, J.; Christiansen, J.; Alberti, K.G. Effects of a growth hormone pulse on total and forearm substrate fluxes in humans. Am. J. Physiol. 1990, 258, E86–E91. [Google Scholar] [CrossRef]
- Vijayakumar, A.; Novosyadlyy, R.; Wu, Y.; Yakar, S.; LeRoith, D. Biological effects of growth hormone on carbohydrate and lipid metabolism. Growth Horm. IGF Res. 2010, 20, 1–7. [Google Scholar] [CrossRef]
- Møller, N.; Jørgensen, J.O.; Alberti, K.G.; Flyvbjerg, A.; Schmitz, O. Short-term effects of growth hormone on fuel oxidation and regional substrate metabolism in normal man. J. Clin. Endocrinol. Metab. 1990, 70, 1179–1186. [Google Scholar] [CrossRef] [PubMed]
- Brandou, F.; Aloulou, I.; Razimbaud, A.; Fédou, C.; Mercier, J.; Brun, J.F. Lower ability to oxidize lipids in adult patients with growth hormone (GH) deficiency: Reversal under GH treatment. Clin. Endocrinol. 2006, 65, 423–428. [Google Scholar] [CrossRef] [PubMed]
- Guha, N.; Sönksen, P.H.; Holt, R.I. IGF-I abuse in sport: Current knowledge and future prospects for detection. Growth Horm. IGF Res. 2009, 19, 408–411. [Google Scholar] [CrossRef] [PubMed]
- Hoene, M.; Weigert, C. The role of interleukin-6 in insulin resistance, body fat distribution and energy balance. Obes. Rev. 2008, 9, 20–29. [Google Scholar] [CrossRef]
- Dyck, D.J. Adipokines as regulators of muscle metabolism and insulin sensitivity. Appl. Physiol. Nutr. Metab. 2009, 34, 396–402. [Google Scholar] [CrossRef]
- Romain, A.J.; Carayol, M.; Desplan, M.; Fedou, C.; Ninot, G.; Mercier, J.; Avignon, A.; Brun, J.F. Physical activity targeted at maximal lipid oxidation: A meta-analysis. J. Nutr. Metab. 2012, 2012, 285395. [Google Scholar] [CrossRef] [PubMed]
- Houmard, J.A.; Tanner, C.J.; Slentz, C.A.; Duscha, B.D.; McCartney, J.S.; Kraus, W.E. Effect of the volume and intensity of exercise training on insulin sensitivity. J. Appl. Physiol. 2004, 96, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Dumortier, M.; Brandou, F.; Perez-Martin, A.; Fedou, C.; Mercier, J.; Brun, J.F. Low-intensity endurance exercise targeted for lipid oxidation improves body composition and insulin sensitivity in patients with the metabolic syndrome. Diabetes Metab. 2003, 29, 509–518. [Google Scholar] [CrossRef]
- Brandou, F.; Savy-Pacaux, A.M.; Marie, J.; Bauloz, M.; Maret-Fleuret, I.; Borrocoso, S. Impact of high- and low-intensity targeted exercise training on the type of substrate utilization in obese boys submitted to a hypocaloric diet. Diabetes Metab. 2005, 31, 327–335. [Google Scholar] [CrossRef]
- Jean, E.; Grubka, E.; Karafiat, M.; Flavier, S.; Fédou, C.; Mercier, J. Effets d’un entraînement en endurance ciblé par la calorimétrie à l’effort chez des diabétiques de type 2. Ann. Endocrinol. 2006, 67, 462. [Google Scholar] [CrossRef]
- Ben Ounis, O.; Elloumi, M.; Ben Chiekh, I.; Zbidi, A.; Amri, M.; Lac, G. Effects of two-month physical-endurance and diet-restriction programmes on lipid profiles and insulin resistance in obese adolescent boys. Diabetes Metab. 2008, 34, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Fédou, C.; Fabre, J.; Baillat, V.; Reynes, J.; Brun, J.F.; Mercier, J. Balance des substrats à l’exercice chez des patients infectés par le VIH 1 et présentant un syndrome lipodystrophique: Effet d’un réentraînement ciblé par la calorimétrie d’effort. Sci. Sports 2008, 23, 189–192. [Google Scholar] [CrossRef]
- Venables, M.C.; Jeukendrup, A.E. Endurance Training and Obesity. Med. Sci. Sports Exerc. 2008, 40, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Ben Ounis, O.; Elloumi, M.; Amri, M.; Zbidi, A.; Tabka, Z.; Lac, G. Impact of diet, exercise and diet combined with exercise programs on plasma lipoprotein and adiponectin levels in obese girls. J. Sport Sci. Med. 2008, 7, 437–445. [Google Scholar]
- Ben Ounis, O.; Elloumi, M.; Amri, M.; Trabelsi, Y.; Lac, G.; Tabka, Z. Impact of training and hypocaloric diet on fat oxidation and body composition in obese adolescents. Sci. Sports 2009, 24, 178–185. [Google Scholar] [CrossRef]
- Ben Ounis, O.; Elloumi, M.; Lac, G.; Makni, E.; Van Praagh, E.; Zouhal, H. Two-month effects of individualized exercise training with or without caloric restriction on plasma adipocytokine levels in obese female adolescents. Ann. Endocrinol. 2009, 70, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Romain, A.J.; Attal, J.; Hermès, A.; Capdevielle, D.; Raimondi, R.; Boulenger, J. Effets d’un réentraînement en endurance au LIPOXmax chez des patients psychiatriques traités par psychotropes. Sci. Sports 2009, 24, 265–268. [Google Scholar] [CrossRef]
- Elloumi, M.; Ben Ounis, O.; Makni, E.; Van Praagh, E.; Tabka, Z.; Lac, G. Effect of individualized weight-loss programmes on adiponectin, leptin and resistin levels in obese adolescent boys. Acta Paediatr. 2009, 98, 1487–1493. [Google Scholar] [CrossRef]
- Maurie, J.; Brun, J.F.; Jean, E.; Romain, A.J.; Mercier, J. Comparaison de deux modalités différentes d’activité physique (SWEET et Lipoxmax) chez des diabétiques de type 2. Sci. Sports 2011, 26, 92–96. [Google Scholar] [CrossRef]
- Tan, S.; Wang, X.; Wang, J. Effects of supervised exercise training at the intensity of maximal fat oxidation in overweight young women. J. Exerc. Sci. Fit. 2012, 10, 64–69. [Google Scholar] [CrossRef]
- Besnier, F.; Lenclume, V.; Gérardin, P.; Fianu, A.; Martinez, J.; Naty, N.; Porcherat, S.; Boussaid, K.; Schneebeli, S.; Jarlet, E. Individualized exercise training at maximal fat oxidation combined with fruit and vegetable-rich diet in overweight or obese women: The LIPOXmax-réunion randomized controlled trial. PLoS ONE 2015, 10, e0139246. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Wang, J.; Cao, L.; Guo, Z.; Wang, Y. Positive effect of exercise training at maximal fat oxidation intensity on body composition and lipid metabolism in overweight middle-aged women. Clin. Physiol. Funct. Imaging 2016, 36, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Wang, J.; Cao, L. Exercise training at the intensity of maximal fat oxidation in obese boys. Appl. Physiol. Nutr. Metab. 2016, 41, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Du, P.; Zhao, W.; Pang, J.; Wang, J. Exercise training at maximal fat oxidation intensity for older women with type 2 diabetes. Int. J. Sports Med. 2018, 39, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Cao, L.; Jiang, Y.; Li, Q.; Wang, J.; Tan, S. Exercise training at maximal fat oxidation intensity for overweight or obese older women: A randomized study. J. Sport Sci. Med. 2019, 18, 413–418. [Google Scholar]
- Zeng, J.; Peng, L.; Zhao, Q.; Chen, Q.G. Effects over 12 weeks of different types and durations of exercise intervention on body composition of young women with obesity. Sci. Sports 2021, 36, 45–52. [Google Scholar] [CrossRef]
- Jiang, Y.; Tan, S.; Wang, Z.; Guo, Z.; Li, Q.; Wang, J. Aerobic exercise training at maximal fat oxidation intensity improves body composition, glycemic control, and physical capacity in older people with type 2 diabetes. J. Exerc. Sci. Fit. 2020, 18, 7–13. [Google Scholar] [CrossRef]
- Brun, J.F.; Myzia, J.; Bui, G.; de Mauverger, E.R.; Mercier, J. The weight-lowering effect of low-intensity endurance training targeted on the level of maximal lipid oxidation (LIPOX MAX) persists over more than 6 years and is associated with improvements in body composition and lipid oxidation. Clin. Nutr. ESPEN 2020, 40, 653. [Google Scholar] [CrossRef]
- Guedjati, M.R.; Silini, S. Effets d’un réentraînement dans la zone du LIPOXmax sur la composition corporelle de femmes obèses d’âge périménopausique. Sci. Sports 2021, 36, 193–201. [Google Scholar] [CrossRef]
- Bughin, F.; Bui, G.; Ayoub, B.; Blervaque, L.; Saey, D.; Avignon, A.; Brun, J.F.; Molinari, N.; Pomies, P.; Mercier, J.; et al. Impact of a mobile telerehabilitation solution on metabolic health outcomes and rehabilitation adherence in patients with obesity: Randomized controlled trial. JMIR mHealth uHealth 2021, 9, e28242. [Google Scholar] [CrossRef]
- Lindström, J.; Ilanne-Parikka, P.; Peltonen, M.; Aunola, S.; Eriksson, J.G.; Hemiö, K. Sustained reduction in the incidence of type 2 diabetes by lifestyle intervention: Follow-up of the Finnish Diabetes Prevention Study. Lancet 2006, 368, 1673–1679. [Google Scholar] [CrossRef]
- Gilis-Januszewska, A.; Lindström, J.; Tuomilehto, J.; Piwońska-Solska, B.; Topór-Mądry, R.; Szybiński, Z.; Peltonen, M.; Schwarz, P.E.; Windak, A.; Hubalewska-Dydejczyk, A. Sustained diabetes risk reduction after real life and primary health care setting implementation of the diabetes in Europe prevention using lifestyle, physical activity and nutritional intervention (DE-PLAN) project. BMC Public Health 2017, 17, 198. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Yang, Y.; Li, Y.; Han, R. Physical exercise as therapy for type 2 diabetes mellitus: From mechanism to orientation. Ann. Nutr. Metab. 2019, 74, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Wake, A.D. Antidiabetic effects of physical activity: How it helps to control Type 2 diabetes. Diabetes Metab. Syndr. Obes. 2020, 13, 2909–2923. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.; Dagnone, D.; Jones, P.J.; Smith, H.; Paddags, A.; Hudson, R. Reduction in obesity and related comorbid conditions after diet-induced weight loss or exercise-induced weight loss in men: A randomized, controlled trial. Ann. Intern. Med. 2000, 133, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Slentz, C.A.; Duscha, B.D.; Johnson, J.L.; Ketchum, K.; Aiken, L.B.; Samsa, G.P. Effects of the amount of exercise on body weight, body composition, and measures of central obesity: STRRIDE--a randomized controlled study. Arch. Intern. Med. 2004, 164, 31–39. [Google Scholar] [CrossRef]
- Baillot, A.; Romain, A.J.; Boisvert-Vigneault, K.; Audet, M.; Baillargeon, J.P.; Dionne, I.J.; Valiquette, L.; Chakra, C.N.; Avignon, A.; Langlois, M.F. Effects of lifestyle interventions that include a physical activity component in class II and III obese individuals: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0119017. [Google Scholar] [CrossRef]
- Kramer, A. An overview of the beneficial effects of exercise on health and performance. Adv. Exp. Med. Biol. 2020, 1228, 3–22. [Google Scholar]
- Tian, Q.; Corkum, A.E.; Moaddel, R.; Ferrucci, L. Metabolomic profiles of being physically active and less sedentary: A critical review. Metabolomics 2021, 17, 68. [Google Scholar] [CrossRef]
- Tremblay, M.S.; Warburton, D.E.; Janssen, I.; Paterson, D.H.; Latimer, A.E.; Rhodes, R.E. New Canadian physical activity guidelines. Appl. Physiol. Nutr. Metab. 2011, 36, 36–46. [Google Scholar] [CrossRef]
- Borgundvaag, E.; Janssen, I. Objectively measured physical activity and mortality risk in American adults. Am. J. Prev. Med. 2017, 52, e25–e31. [Google Scholar] [CrossRef] [PubMed]
- Hupin, D.; Raffin, J.; Barth, N.; Berger, M.; Garet, M.; Stampone, K.; Celle, S.; Pichot, V.; Bongue, B.; Barthelemy, J.C.; et al. Even a previous light-active physical activity at work still reduces late myocardial infarction and stroke in retired adults aged >65 years by 32%: The PROOF Cohort Study. Front. Public Health 2019, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Kokkinos, P.; Myers, J.; Faselis, C.; Panagiotakos, D.B.; Doumas, M.; Pittaras, A.; Manolis, A.; Kokkinos, J.P.; Karasik, P.; Greenberg, M.; et al. Exercise capacity and mortality in older men: A 20-year follow-up study. Circulation 2010, 122, 790–797. [Google Scholar] [CrossRef] [PubMed]
- Fritz, T.; Krämer, D.K.; Karlsson, H.K.; Galuska, D.; Engfeldt, P.; Zierath, J.R.; Krook, A. Low-intensity exercise increases skeletal muscle protein expression of PPARdelta and UCP3 in type 2 diabetic patients. Diabetes Metab. Res. Rev. 2006, 22, 492–498. [Google Scholar] [CrossRef]
- Fu, Q.; Levine, B.D. Exercise and the autonomic nervous system. Handb. Clin. Neurol. 2013, 117, 147–160. [Google Scholar]
- Izquierdo-Alventosa, R.; Inglés, M.; Cortés-Amador, S.; Gimeno-Mallench, L.; Chirivella-Garrido, J.; Kropotov, J.; Serra-Añó, P. Low-intensity physical exercise improves pain catastrophizing and other psychological and physical aspects in women with fibromyalgia: A randomized controlled trial. Int. J. Environ. Res. Public Health 2020, 17, 3634. [Google Scholar] [CrossRef]
- Dipietro, L.; Evenson, K.R.; Bloodgood, B.; Sprow, K.; Troiano, R.P.; Piercy, K.L.; Vaux-Bjerke, A.; Powell, K.E. Benefits of physical activity during pregnancy and postpartum: An umbrella review. Med. Sci. Sports Exerc. 2019, 51, 1292–1302. [Google Scholar] [CrossRef]
- Peters, T.M.; Brazeau, A.S. Exercise in pregnant women with diabetes. Curr. Diabetes Rep. 2019, 19, 80. [Google Scholar] [CrossRef]
- Baggetta, R.; D’Arrigo, G.; Torino, C.; El Hafeez, S.A.; Manfredini, F.; Mallamaci, F.; Zoccali, C.; Tripepi, G. EXCITE Working group. Effect of a home based, low-intensity, physical exercise program in older adults dialysis patients: A secondary analysis of the EXCITE trial. BMC Geriatr. 2018, 18, 248. [Google Scholar] [CrossRef]
- Villanego, F.; Naranjo, J.; Vigara, L.A.; Cazorla, J.M.; Montero, M.E.; García, T.; Torrado, J.; Mazuecos, A. Impact of physical exercise in patients with chronic kidney disease: Sistematic review and meta-analysis. Nefrologia 2020, 40, 237–252. [Google Scholar] [CrossRef]
- Armour, M.; Ee, C.C.; Naidoo, D.; Ayati, Z.; Chalmers, K.J.; Steel, K.A.; de Manincor, M.J.; Delshad, E. Exercise for dysmenorrhoea. Cochrane Database Syst. Rev. 2019, 9, CD004142. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S. Prescription of exercise training for hypertensives. Hypertens. Res. 2020, 43, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Kistler-Fischbacher, M.; Weeks, B.K.; Beck, B.R. The effect of exercise intensity on bone in postmenopausal women (part 2): A meta-analysis. Bone 2021, 143, 115697. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.; Li, Y.; Yang, J.; Li, X.; Zeng, N.; Martin, R.L. The effectiveness of low-intensity exercise and blood flow restriction without exercise on exercise induced muscle damage: A systematic review. Phys. Ther. Sport 2020, 46, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Varlet-Marie, E.; Brun, J.F.; Myzia, F.; Bui, G.; Chérif, A.; Javernaud, E.; Hammoudi, L.; Mercier, J.; de Mauverger, E.R. Exercise targeted on maximal lipid oxidation has long term weight-reducing effects but improves insulin secretion and lipid oxidation before reducing fat mass. Sci. Sports 2022. [Google Scholar] [CrossRef]
- Dandanell, S.; Skovborg, C.; Præst, C.B.; Kristensen, K.B.; Nielsen, M.G.; Lionett, S.; Jørgensen, S.D.; Vigelsø, A.; Dela, F.; Helge, J.W. Maintaining a clinical weight loss after intensive lifestyle intervention is the key to cardiometabolic health. Obes. Res. Clin. Pract. 2017, 11, 489–498. [Google Scholar] [CrossRef]
- Dandanell, S.; Husted, K.; Amdisen, S.; Vigelsø, A.; Dela, F.; Larsen, S.; Helge, J.W. Influence of maximal fat oxidation on long-term weight loss maintenance in humans. J. Appl. Physiol. 2017, 123, 267–274. [Google Scholar] [CrossRef]
- Brun, J.F.; Maurie, J.; Jean, E.; Romain, A.; Mercier, J. Comparison of square-wave endurance exercise test (SWEET) training with endurance training targeted at the level of maximal lipid oxidation in type 2 diabetics. Sci. Sports 2011, 26, 92–96. [Google Scholar]
- Schoenfeld, B.J. The mechanisms of muscle hypertrophy and their application to resistance training. J. Strength Cond. Res. 2010, 24, 2857–2872. [Google Scholar] [CrossRef]
- Krzysztofik, M.; Wilk, M.; Wojdała, G.; Gołaś, A. Maximizing muscle hypertrophy: A systematic review of advanced resistance training techniques and methods. Int. J. Environ. Res. Public Health 2019, 16, 4897. [Google Scholar] [CrossRef]
- Nicklas, B.J.; Wang, X.; You, T.; Lyles, M.F.; Demons, J.; Easter, L. Effect of exercise intensity on abdominal fat loss during calorie restriction in overweight and obese postmenopausal women: A randomized, controlled trial. Am. J. Clin. Nutr. 2009, 89, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, B.; Phillips, S.M.; Atherton, P.J.; Patel, R.; Yarasheski, K.E.; Tarnopolsky, M.A. Differential effects of resistance and endurance exercise in the fed state on signalling molecule phosphorylation and protein synthesis in human muscle. J. Physiol. 2008, 586, 3701–3717. [Google Scholar] [CrossRef] [PubMed]
- Howarth, K.R.; Phillips, S.M.; MacDonald, M.J.; Richards, D.; Moreau, N.A.; Gibala, M.J. Effect of glycogen availability on human skeletal muscle protein turnover during exercise and recovery. J. Appl. Physiol. 2010, 109, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Perreault, K.; Lagacé, J.C.; Brochu, M.; Dionne, I.J. Association between fat free mass and glucose homeostasis: Common knowledge revisited. Ageing Res. Rev. 2016, 28, 46–61. [Google Scholar] [CrossRef]
- Lagacé, J.C.; Brochu, M.; Dionne, I.J. A counterintuitive perspective for the role of fat-free mass in metabolic health. J. Cachexia Sarcopenia Muscle 2020, 11, 343–347. [Google Scholar] [CrossRef]
- Lambert, K.; Kitzmann, M.; Aguer, C.; Mannarino, A.; Fédou, C.; de Mauverger, E.R.; Mercier, J.; Brun, J.F. Chez des obèses, l’aptitude à oxyder les lipides à l’exercice est corrélée à l’insulinosensibilité et à la capacité oxydative musculaire. Nutr. Clin. Métab. 2016, 30, 277. [Google Scholar] [CrossRef]
- Lambert, K.; Aguer, C.; Kitzmann, M.; Mannarino, A.; Fedou, C.; de Mauverger, E.R.; Mercier, J.; Brun, J.F. Whole-body lipid oxidation during exercise is correlated to insulin sensitivity and mitochondrial function in middle-aged obese men. Austin Diabetes Res. 2017, 2, 1013. [Google Scholar]
- Robinson, S.L.; Hattersley, J.; Frost, G.S.; Chambers, E.S.; Wallis, G.A. Maximal fat oxidation during exercise is positively associated with 24-hour fat oxidation and insulin sensitivity in young, healthy men. J. Appl. Physiol. 2015, 118, 1415–1422. [Google Scholar] [CrossRef]
- Gaggini, M.; Morelli, M.; Buzzigoli, E.; DeFronzo, R.A.; Bugianesi, E.; Gastaldelli, A. Non-alcoholic fatty liver disease (NAFLD) and its connection with insulin resistance, dyslipidemia, atherosclerosis and coronary heart disease. Nutrients 2013, 5, 1544–1560. [Google Scholar] [CrossRef]
- Malin, S.K.; Solomon, T.P.; Blaszczak, A. Pancreatic beta-cell function increases in a linear dose-response manner following exercise training in adults with prediabetes. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1248–E1254. [Google Scholar] [CrossRef]
- Michishita, R.; Shono, N.; Kasahara, T.; Tsuruta, T. Effects of low-intensity exercise therapy on early phase insulin secretion in overweight subjects with impaired glucose tolerance and type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2008, 82, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Solomon, T.P.; Malin, S.K.; Karstoft, K. Pancreatic beta-cell function is a stronger predictor of changes in glycemic control after an aerobic exercise intervention than insulin sensitivity. J. Clin. Endocrinol. Metab. 2013, 98, 4176–4186. [Google Scholar] [CrossRef] [PubMed]
- Rutti, S.; Arous, C.; Schvartz, D.; Timper, K.; Sanchez, J.C.; Dermitzakis, E.; Donath, M.Y.; Halban, P.A.; Bouzakri, K. Fractalkine (CX3CL1), a new factor protecting β-cells against TNFα. Mol. Metab. 2014, 3, 731–741. [Google Scholar] [CrossRef] [PubMed]
- Heiskanen, M.A.; Motiani, K.K.; Mari, A.; Saunavaara, V.; Eskelinen, J.J.; Virtanen, K.A.; Koivumäki, M.; Löyttyniemi, E.; Nuutila, P.; Kalliokoski, K.K.; et al. Exercise training decreases pancreatic fat content and improves beta cell function regardless of baseline glucose tolerance: A randomised controlled trial. Diabetologia 2018, 61, 1817–1828. [Google Scholar] [CrossRef]
- Slentz, C.A.; Tanner, C.J.; Bateman, L.A.; Durheim, M.T.; Huffman, K.M.; Houmard, J.A.; Kraus, W.E. Effects of exercise training intensity on pancreatic beta-cell function. Diabetes Care 2009, 32, 1807–1811. [Google Scholar] [CrossRef]
- Langlois, A.; Forterre, A.; Pinget, M.; Bouzakri, K. Impact of moderate exercise on fatty acid oxidation in pancreatic β-cells and skeletal muscle. J. Endocrinol. Investig. 2021, 44, 1815–1825. [Google Scholar] [CrossRef]
- Mattusch, F.; Dufaux, B.; Heine, O.; Mertens, I.; Rost, R. Reduction of the plasma concentration of C-reactive protein following nine months of endurance training. Int. J. Sports Med. 2000, 21, 21–24. [Google Scholar] [CrossRef]
- Stewart, L.K.; Flynn, M.G.; Campbell, W.W.; Craig, B.A.; Robinson, J.P.; Timmerman, K.L. The influence of exercise training on inflammatory cytokines and C-reactive protein. Med. Sci. Sports Exerc. 2007, 39, 1714–1719. [Google Scholar] [CrossRef]
- Goldhammer, E.; Tanchilevitch, A.; Maor, I.; Beniamini, Y.; Rosenschein, U.; Sagiv, M. Exercise training modulates cytokines activity in coronary heart disease patients. Int. J. Cardiol. 2005, 100, 93–99. [Google Scholar] [CrossRef]
- Martins, R.A.; Neves, A.P.; Coelho-Silva, M.J.; Veríssimo, M.T.; Teixeira, A.M. The effect of aerobic versus strength-based training on high-sensitivity C-reactive protein in older adults. Eur. J. Appl. Physiol. 2010, 110, 161–169. [Google Scholar] [CrossRef]
- Ben Ounis, O.; Elloumi, M.; Zouhal, H.; Makni, E.; Denguezli, M.; Amri, M. Effect of individualized exercise training combined with diet restriction on inflammatory markers and IGF-1/IGFBP-3 in obese children. Ann. Nutr. Metab. 2010, 56, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Fedou, C.; Bordenave, S.; Metz, L.; Lambert, K.; Dupuy, A. L’amélioration de l’oxydation des lipides induite par l’entrainement ciblé au LIPOXmax chez des diabétiques de type 2 s’accompagne d’une diminution de la protéine C-réactive (CRP). Ann. Endocrinol. Paris 2010, 71, 416. [Google Scholar]
- Gram, A.S.; Bladbjerg, E.M.; Quist, J.S.; Petersen, M.B.; Rosenkilde, M.; Stallknecht, B. Anti-inflammatory effects of active commuting and leisure time exercise in overweight and obese women and men: A randomized controlled trial. Atherosclerosis 2017, 265, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Varlet-Marie, E.; Connes, P.; Aloulou, I. Hemorheological alterations related to training and overtraining. Biorheology 2010, 47, 95–115. [Google Scholar] [CrossRef] [PubMed]
- Lazzer, S.; Busti, C.; Agosti, F.; De Col, A.; Pozzo, R.; Sartorio, A. Optimizing fat oxidation through exercise in severely obese Caucasian adolescents. Clin. Endocrinol. 2007, 67, 582–588. [Google Scholar] [CrossRef]
- Lazzer, S.; Tringali, G.; Caccavale, M. Effects of high-intensity interval training on physical capacities and substrate oxidation rate in obese adolescents. J. Endocrinol. Investig. 2017, 40, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Brun, J.F.; Sardinoux, M.; Nocca, D.; Salsano, V.; Fabre, G.; Bringer, J.; Lefebvre, P.; Avignon, A.; Mercier, J. Weight loss induced by bariatric surgery improves the ability to oxidize lipids at exercise but shifts the power of maximal lipid oxidation (LIPOXmax) to the left. Diabetologia 2009, 52, S273. [Google Scholar]
- Drapier, E.; Fédou, C.; Ghanassia, E.; Nocca, D.; Mercier, J. L’exercice en endurance à faible intensité renforce-t-il l’efficacité amaigrissante d’une gastrectomie en manchon? Sci. Sports 2016, 31, 162–165. [Google Scholar] [CrossRef]
- Brun, J.F.; Lasteyrie, V.; Hammoudi, L.; Nocca, D.; Ghanassia, E. Exercise targeted at the level of maximal lipid oxidation (LIPOXmax) improves weight loss, decreases orexigenic pulsions and increases satiety after sleeve gastrectomy. Glob. J. Obes. Diabetes Metab. Syndr. 2019, 6, 017–021. [Google Scholar]
- Filou, V.; Richou, M.; Bughin, F.; Fédou, C.; de Mauverger, E.R.; Mercier, J.; Brun, J.F. Complementarity of bariatric surgery and physical activity. Sci. Sports 2019, 33, 65–72. [Google Scholar]
- Umpierre, D.; Ribeiro, P.A.; Kramer, C.K.; Leitão, C.B.; Zucatti, A.T.; Azevedo, M.J.; Gross, J.L.; Ribeiro, J.P.; Schaan, B.D. Physical activity advice only or structured exercise training and association with HbA1c levels in type 2 diabetes: A systematic review and meta-analysis. JAMA 2011, 305, 1790–1799. [Google Scholar] [CrossRef] [PubMed]
- Savikj, M.; Zierath, J.R. Train like an athlete: Applying exercise interventions to manage type 2 diabetes. Diabetologia 2020, 63, 1491–1499. [Google Scholar] [CrossRef]
- Nguyen, J.M.; Richou, M.; Bughin, F.; Mercier, J.; de Mauverger, E.R.; Brun, J.F. Effet à 3 ans sur le poids et l’équilibre glycémique de l'exercice en endurance ciblé sur l’oxydation des lipides (LIPOXmax) chez des diabétiques de type 2 comparés à des non-diabétiques. Nutr. Clin. Métab. 2019, 33, 55–56. [Google Scholar] [CrossRef]
- Francescato, M.P.; Cattin, L.; Geat, M.; Tosoratti, E.; Lazzer, S.; Noacco, C.; di Prampero, P.E. Glucose Pulse: A simple method to estimate the amount of glucose oxidized during exercise in type 1 diabetic patients. Diabetes Care 2005, 28, 2028–2030. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Brun, J.F.; Traverso, M.; Fédou, C.; Renard, E.; Mercier, J. Deux mythes concernant l’exercice chez le diabétique de type 1: Le seuil hyperglycémique de 2, 5 g/L de contre-indication à l’effort et le “glucose pulse”. Sci. Sports 2009, 24, 108–110. [Google Scholar] [CrossRef]
- Marinari, G.; Espitalier-Rivière, C.; Fédou, C.; Romain, A.J.; Brun, J.F. Balance des substrats à l’exercice déterminée par calorimétrie d’effort chez des patientes entreprenant un réentraînement après traitement d’un cancer mammaire. Sci. Sports 2018, 33, 185–189. [Google Scholar] [CrossRef]
- Marinari, G.; Espitalier-Rivière, C.; Fédou, C.; Romain, A.J.; Brun, J.F. Activité physique, obésité et cancer du sein: Quelles conclusions pratiques? Physical activity, obesity and breast cancer: Which practical conclusions? Sci. Sports 2019, 34, 191–198. [Google Scholar] [CrossRef]
- Golightly, Y.M.; Allen, K.D.; Caine, D.J. A comprehensive review of the effectiveness of different exercise programs for patients with osteoarthritis. Physician Sportsmed. 2012, 40, 52–65. [Google Scholar] [CrossRef]
- Wellsandt, E.; Golightly, Y. Exercise in the management of knee and hip osteoarthritis. Curr. Opin. Rheumatol. 2018, 30, 151–159. [Google Scholar] [CrossRef]
- Mauro, M.; Taylor, V.; Wharton, S.; Sharma, A.M. Barriers to obesity treatment. Eur. J. Intern. Med. 2008, 19, 173–180. [Google Scholar] [CrossRef]
- Svetkey, L.P.; Stevens, V.J.; Brantley, P.J.; Appel, L.J.; Hollis, J.F.; Loria, C.M. Comparison of strategies for sustaining weight loss. J. Am. Med. Assoc. 2008, 299, 1139–1148. [Google Scholar] [CrossRef] [PubMed]
- Mons, U.; Hahmann, H.; Brenner, H. A reverse J-shaped association of leisure time physical activity with prognosis in patients with stable coronary heart disease: Evidence from a large cohort with repeated measurements. Heart 2014, 100, 1043–1049. [Google Scholar] [CrossRef] [PubMed]
- Ochner, C.N.; Barrios, D.M.; Lee, C.D.; Pi-Sunyer, F.X. Biological mechanisms that promote weight regain following weight loss in obese humans. Physiol. Behav. 2013, 120, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.T.; Astrup, A.; Sjödin, A. Are dietary proteins the key to successful body weight management? A systematic review and meta-analysis of studies assessing body weight outcomes after interventions with increased dietary protein. Nutrients 2021, 13, 3193. [Google Scholar] [CrossRef]
- Dutheil, F.; Lac, G.; Courteix, D.; Doré, E.; Chapier, R.; Roszyk, L.; Sapin, V.; Lesourd, B. Treatment of metabolic syndrome by combination of physical activity and diet needs an optimal protein intake: A randomized controlled trial. Nutr. J. 2012, 11, 72. [Google Scholar] [CrossRef]
- Soenen, S.; Martens, E.A.; Hochstenbach-Waelen, A.; Lemmens, S.G.; Westerterp-Plantenga, M.S. Normal protein intake is required for body weight loss and weight maintenance, and elevated protein intake for additional preservation of resting energy expenditure and fat free mass. J. Nutr. 2013, 143, 591–596. [Google Scholar] [CrossRef]
- Koulmann, N.; Bigard, A.X. Interaction between signalling pathways involved in skeletal muscle responses to endurance exercise. Pflugers Arch. 2006, 452, 125–139. [Google Scholar] [CrossRef]
- Hart, D.A.; Zernicke, R.F. Optimal human functioning requires exercise across the lifespan: Mobility in a 1 g environment is intrinsic to the integrity of multiple biological systems. Front. Physiol. 2020, 11, 156. [Google Scholar] [CrossRef]
- Bruhn, L.; Kjøbsted, R.; Quist, J.S.; Gram, A.S.; Rosenkilde, M.; Faerch, K.; Wojtaszewski, J.F.; Stallknecht, B.; Blond, M.B. Effect of exercise training on skeletal muscle protein expression in relation to insulin sensitivity: Per-protocol analysis of a randomized controlled trial (GO-ACTIWE). Physiol. Rep. 2021, 9, e14850. [Google Scholar] [CrossRef]
- Stamatakis, E.; Gale, J.; Bauman, A.; Ekelund, U.; Hamer, M.; Ding, D. Sitting time, physical activity, and risk of mortality in adults. J. Am. Coll. Cardiol. 2019, 73, 2062–2072. [Google Scholar] [CrossRef]
- Stamatakis, E.; Ekelund, U.; Ding, D.; Hamer, M.; Bauman, A.E.; Lee, I.M. Is the time right for quantitative public health guidelines on sitting? A narrative review of sedentary behaviour research paradigms and findings. Br. J. Sports Med. 2019, 53, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Katzmarzyk, P.T.; Powell, K.E.; Jakicic, J.M.; Troiano, R.P.; Piercy, K.; Tennant, B. Sedentary behavior and health: Update from the 2018 Physical Activity Guidelines Advisory Committee. Med. Sci. Sports Exerc. 2019, 51, 1227–1241. [Google Scholar] [CrossRef] [PubMed]
- Cabanas-Sánchez, V.; Guallar-Castillón, P.; Higueras-Fresnillo, S.; Rodríguez-Artalejo, F.; Martínez-Gómez, D. Changes in sitting time and cardiovascular mortality in older adults. Am. J. Prev. Med. 2018, 54, 419–422. [Google Scholar] [CrossRef] [PubMed]
- Rees-Punia, E.; Evans, E.M.; Schmidt, M.D.; Gay, J.L.; Matthews, C.E.; Gapstur, S.M.; Patel, A.V. Mortality risk reductions for replacing sedentary time with physical activities. Am. J. Prev. Med. 2019, 56, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Spreitzer, A.; Haber, P. Fat loss depends on energy deficit only, independently of the method for weight loss. Ann. Nutr. Metab. 2007, 51, 428–432. [Google Scholar] [CrossRef]
- Slentz, C.A.; Bateman, L.A.; Willis, L.H.; Shields, A.T.; Tanner, C.J.; Piner, L.W.; Hawk, V.H.; Muehlbauer, M.J.; Samsa, G.P.; Nelson, R.C.; et al. Effects of aerobic vs. resistance training on visceral and liver fat stores, liver enzymes, and insulin resistance by HOMA in overweight adults from STRRIDE AT/RT. Am. J. Physiol. Endocrinol. Metab. 2011, 301, E1033–E1039. [Google Scholar] [CrossRef]
- Aguer, C.; Mercier, J.; Yong Wai Man, C.; Metz, L.; Bordenave, S.; Lambert, K.; Jean, E.; Lantier, L.; Bounoua, L.; Brun, J.F.; et al. Intramyocellular lipid accumulation is associated with permanent relocation ex vivo and in vitro of fatty acid translocase (FAT)/CD36 in obese patients. Diabetologia 2010, 53, 1151–1163. [Google Scholar] [CrossRef]
- Berggren, J.R.; Boyle, K.E.; Chapman, W.H.; Houmard, J.A. Skeletal muscle lipid oxidation and obesity: Influence of weight loss and exercise. Am. J. Physiol. Endocrinol. Metab. 2008, 294, E726–E732. [Google Scholar] [CrossRef]
- Houmard, J.A.; Pories, W.J.; Dohm, G.L. Is there a metabolic program in the skeletal muscle of obese individuals? J. Obes. 2011, 2011, 250496. [Google Scholar] [CrossRef]
- Houmard, J.A.; Pories, W.J.; Dohm, G.L. Severe obesity: Evidence for a deranged metabolic program in skeletal muscle? Exerc. Sport Sci. Rev. 2012, 40, 204–210. [Google Scholar] [CrossRef]
- Cavalli, G.; Heard, E. Advances in epigenetics link genetics to the environment and disease. Nature 2019, 571, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, S.J.; Molloy, P.L.; Varinli, H.; Morrison, J.L.; Muhlhausler, B.S.; Members of Epi-SCOPE. Epigenetics and human obesity. Int. J. Obes. 2015, 39, 85–97. [Google Scholar] [CrossRef] [PubMed]
- Demerath, E.W.; Guan, W.; Grove, M.L. Epigenome-wide association study (EWAS) of BMI, BMI change and waist circumference in African American adults identifies multiple replicated loci. Hum. Mol. Genet. 2015, 24, 4464–4479. [Google Scholar] [CrossRef] [PubMed]
- Dick, K.J.; Nelson, C.P.; Tsaprouni, L. DNA methylation and body-mass index: A genome-wide analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef]
- Ling, C.; Rönn, T. Epigenetics in human obesity and Type 2 diabetes. Cell Metab. 2019, 29, 1028–1044. [Google Scholar] [CrossRef]
- Booth, F.W.; Roberts, C.K.; Thyfault, J.P.; Ruegsegger, G.N.; Toedebusch, R.G. Role of inactivity in chronic diseases: Evolutionary insight and pathophysiological mechanisms. Physiol. Rev. 2017, 97, 1351–1402. [Google Scholar] [CrossRef]
- Jacobsen, S.C.; Brøns, C.; Bork-Jensen, J. Effects of short-term high-fat overfeeding on genome-wide DNA methylation in the skeletal muscle of healthy young men. Diabetologia 2012, 55, 3341–3349. [Google Scholar] [CrossRef]
- Alibegovic, A.C.; Sonne, M.P.; Højbjerre, L.; Bork-Jensen, J.; Jacobsen, S.; Nilsson, E.; Faerch, K.; Hiscock, N.; Mortensen, B.; Friedrichsen, M.; et al. Insulin resistance induced by physical inactivity is associated with multiple transcriptional changes in skeletal muscle in young men. Am. J. Physiol. Endocrinol. Metab. 2010, 299, E752–E763. [Google Scholar] [CrossRef]
- Egan, B.; Zierath, J.R. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef]
- Barres, R.; Yan, J.; Egan, B.; Treebak, J.T.; Rasmussen, M.; Fritz, T.; Caidahl, K.; Krook, A.; O’Gorman, D.J.; Zierath, J.R. Acute exercise remodels promoter methylation in human skeletal muscle. Cell Metab. 2012, 15, 405–411. [Google Scholar] [CrossRef]
- Voisin, S.; Eynon, N.; Yan, X.; Bishop, D.J. Exercise training and DNA methylation in humans. Acta Physiol. 2015, 213, 39–59. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, M.E.; Marabita, F.; Gomez-Cabrero, D. An integrative analysis reveals coordinated reprogramming of the epigenome and the transcriptome in human skeletal muscle after training. Epigenetics 2014, 9, 1557. [Google Scholar] [CrossRef]
- Papaioannou, F.; Karatzanos, E.; Chatziandreou, I.; Philippou, A.; Nanas, S.; Dimopoulos, S. Epigenetic effects following acute and chronic exercise in cardiovascular disease: A systematic review. Int. J. Cardiol. 2021, 341, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Tarnowski, M.; Kopytko, P.; Piotrowska, K. Epigenetic regulation of inflammatory responses in the context of physical activity. Genes 2021, 12, 1313. [Google Scholar] [CrossRef] [PubMed]
- Widmann, M.; Nieß, A.M.; Munz, B. Physical exercise and epigenetic modifications in skeletal muscle. Sports Med. 2019, 49, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.G.; Twomey, L.C.; Custaud, M.A.; Turner, J.D.; Moyna, N.; Cummins, P.M.; Murphy, R.P. The role of epigenetics in cardiovascular health and ageing: A focus on physical activity and nutrition. Mech. Ageing Dev. 2018, 174, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Nitert, M.D.; Dayeh, T.; Volkov, P.; Elgzyri, T.; Hall, E.; Nilsson, E.; Yang, B.T.; Lang, S.; Parikh, H.; Wessman, Y.; et al. Impact of an exercise intervention on DNA methylation in skeletal muscle from first-degree relatives of patients with type 2 diabetes. Diabetes 2012, 61, 3322–3332. [Google Scholar] [CrossRef]
- King, N.A.; Caudwell, P.P.; Hopkins, M.; Stubbs, J.R.; Naslund, E.; Blundell, J.E. Dual-process action of exercise on appetite control: Increase in orexigenic drive but improvement in meal-induced satiety. Am. J. Clin. Nutr. 2009, 90, 921–927. [Google Scholar] [CrossRef]
- King, N.M.; Hopkins, M.; Caudwell, P.P.; Stubbs, R.J.; Blundell, J.E. Individual variability following 12 weeks of supervised exercise: Identification and characterization of compensation for exercise-induced weight loss. Int. J. Obes. 2008, 32, 177–184. [Google Scholar] [CrossRef]
- Beaulieu, K.; Blundell, J.E.; van Baak, M.A.; Battista, F.; Busetto, L.; Carraça, E.V.; Dicker, D.; Encantado, J.; Ermolao, A.; Farpour-Lambert, N.; et al. Effect of exercise training interventions on energy intake and appetite control in adults with overweight or obesity: A systematic review and meta-analysis. Obes. Rev. 2021, 22 (Suppl. S2), e13251. [Google Scholar] [CrossRef]
- Mayer, J.; Roy, P.; Mitra, K.P. Relation between caloric intake, body weight, and physical work: Studies in an industrial male population in West Bengal. Am. J. Clin. Nutr. 1956, 4, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Pollatz, M.; Guiraudou, M.; Romain, A.J.; Fédou, C.; Brun, J.F.; Mercier, J. Les effets d’un réentraînement de 45 minutes au LIPOXmax sur le comportement alimentaire de patients obèses sont analogues le matin et l’après-midi. Sci. Sports 2014, 29, 57–61. [Google Scholar] [CrossRef]
- Guiraudou, M.; Romain, A.J.; Mawunu, M.; Bedjih, K.; Fédou, C.; Brun, J.F. Effets chroniques de l’exercice ciblé au niveau d’oxydation maximale des lipides (LIPOXmax) sur le comportement alimentaire de sujets obèses sédentaires. Sci. Sports 2016, 31, 13–18. [Google Scholar] [CrossRef]
- Brun, J.F.; Romain, A.J.; Sferlazza, A.; Fédou, C.; de Mauverger, E.R.; Mercier, J. Which individuals become fatter when they practice exercise? Sci. Sports 2016, 31, 214–218. [Google Scholar] [CrossRef]
- Quist, J.; Blond, M.B.; Gram, A.S.; Steenholt, C.B.; Janus, C.; Holst, J.J.; Rehfeld, J.F.; Sjödin, A.; Stallknecht, B.; Rosenkilde, M. Effects of active commuting and leisure-time exercise on appetite in individuals with overweight and obesity. J. Appl. Physiol. 2019, 126, 941–951. [Google Scholar] [CrossRef]
- Anderson, K.C.; Zieff, G.; Paterson, C.; Stoner, L.; Weltman, A.; Allen, J.D. The effect of acute exercise on pre-prandial ghrelin levels in healthy adults: A systematic review and meta-analysis. Peptides 2021, 145, 170625. [Google Scholar] [CrossRef]
- Gonzalez-Gil, A.M.; Elizondo-Montemayor, L. The role of exercise in the interplay between myokines, hepatokines, osteokines, adipokines, and modulation of inflammation for energy substrate redistribution and fat mass loss: A review. Nutrients 2020, 12, 1899. [Google Scholar] [CrossRef]
- Murphy, M.P.; O’Neill, L.A. Krebs cycle reimagined: The emerging roles of succinate and itaconate as signal transducers. Cell 2018, 174, 780–784. [Google Scholar] [CrossRef]
- Alghannam, A.F.; Ghaith, M.M.; Alhussain, M.H. Regulation of energy substrate metabolism in endurance exercise. Int. J. Environ. Res. Public Health 2021, 18, 4963. [Google Scholar] [CrossRef]
- Coquart, J.B.; Boitel, G.; Garcin, M. Effects of a training program at the crossover point on the cluster of metabolic abnormalities and cardiovascular risk factors. J. Exerc. Sci. Fit. 2014, 12, 73–79. [Google Scholar] [CrossRef]
- Myzia, J.; Brun, J.F.; Varlet-Marie, E.; de Mauverger, E.R.; Mercier, J. Endurance training minimizing carbohydrate oxidation by targeting the optimal level of fat/carbohydrate oxidation ratio (OLORFOX)? Sci. Sports, 2022; in press. [Google Scholar]
- Brun, J.F.; Metrat, S.; Nguyen, J.M.; Richou, M.; Rabta, F.M. Subjects with substituted hypothyroidism oxidize more lipids and carbohydrates during exercise. Ann. Musculoskelet. Med. 2018, 2, 013–016. [Google Scholar]











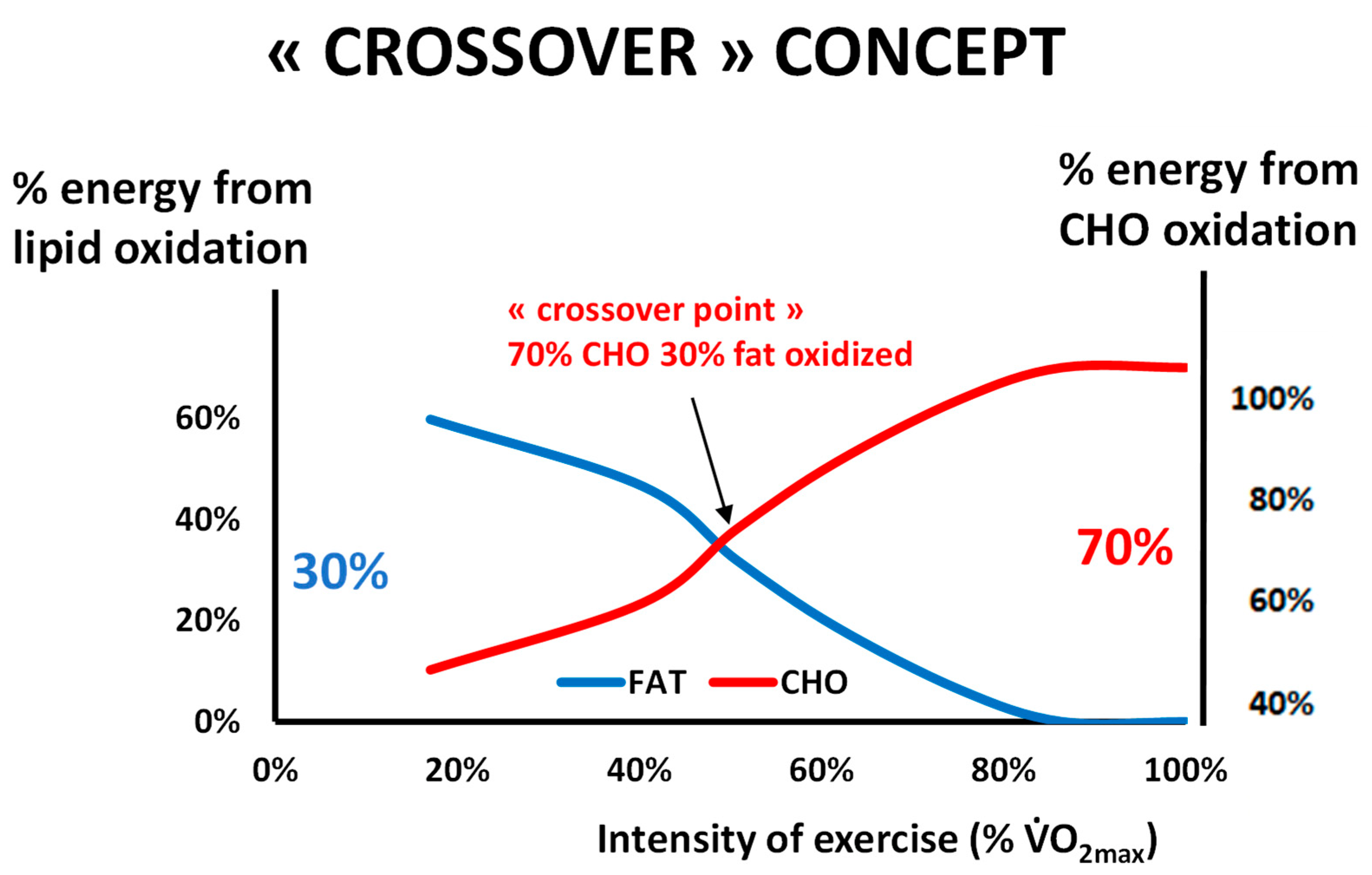{kind=link}
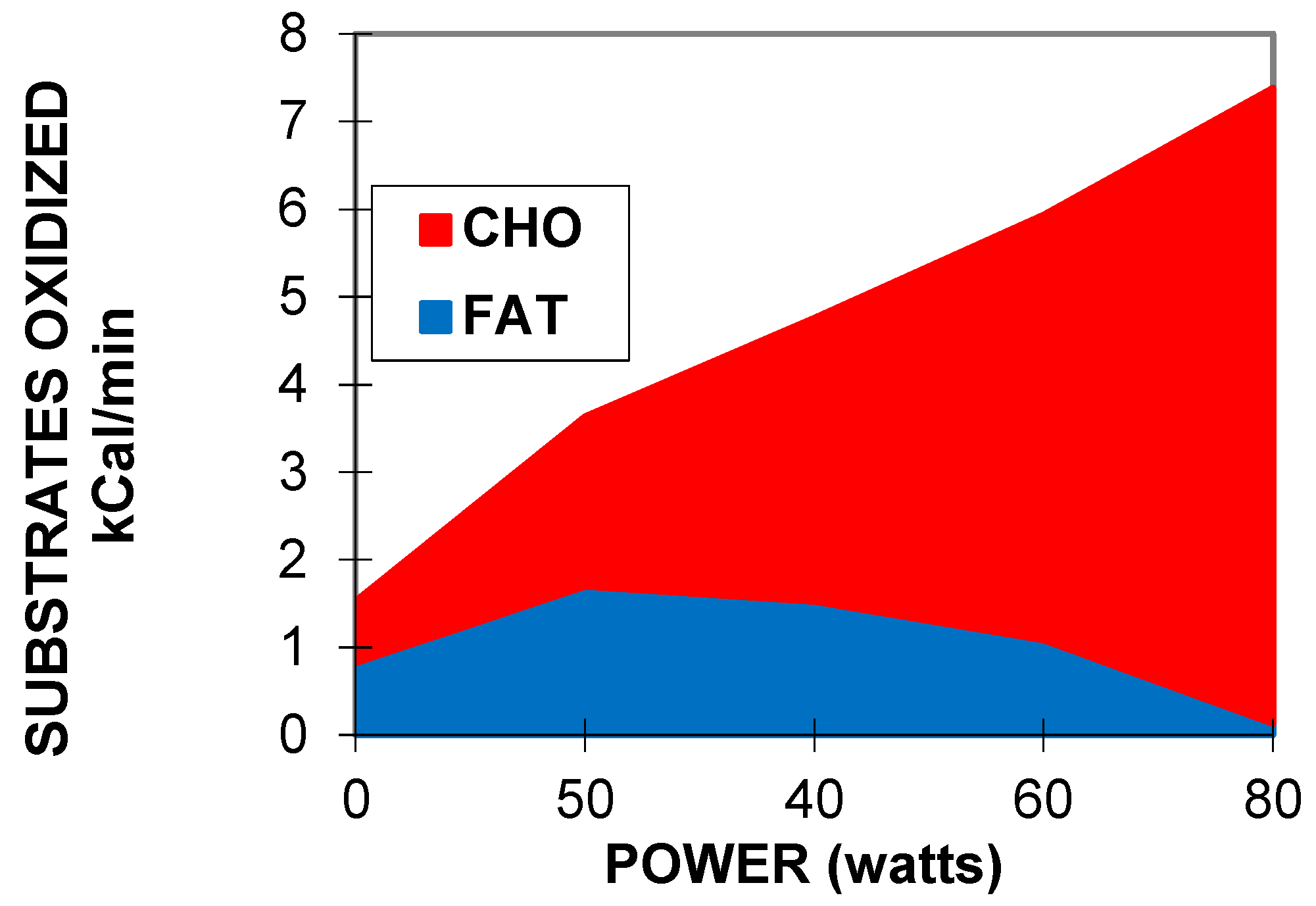{kind=link}
{kind=link}
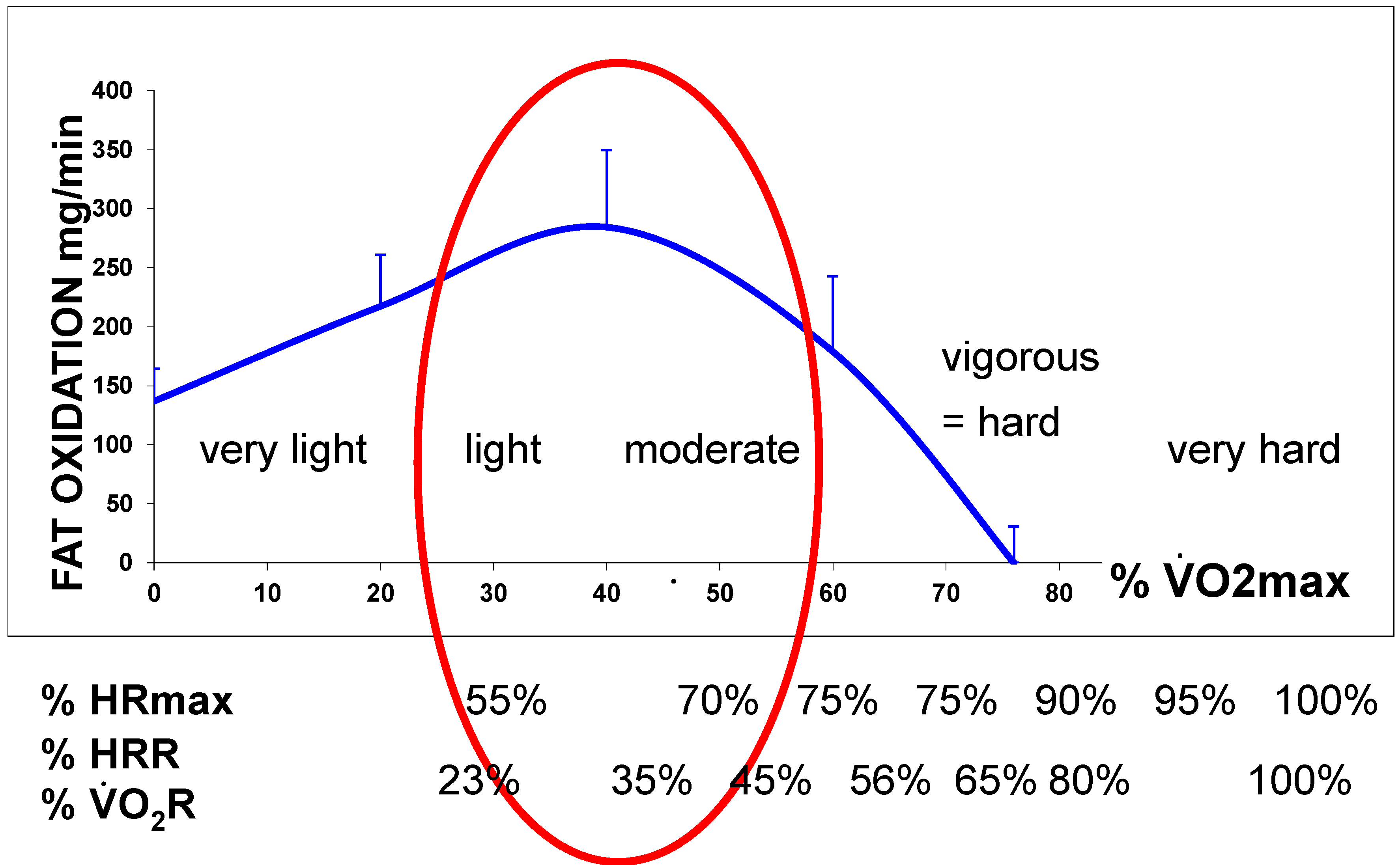{kind=link}
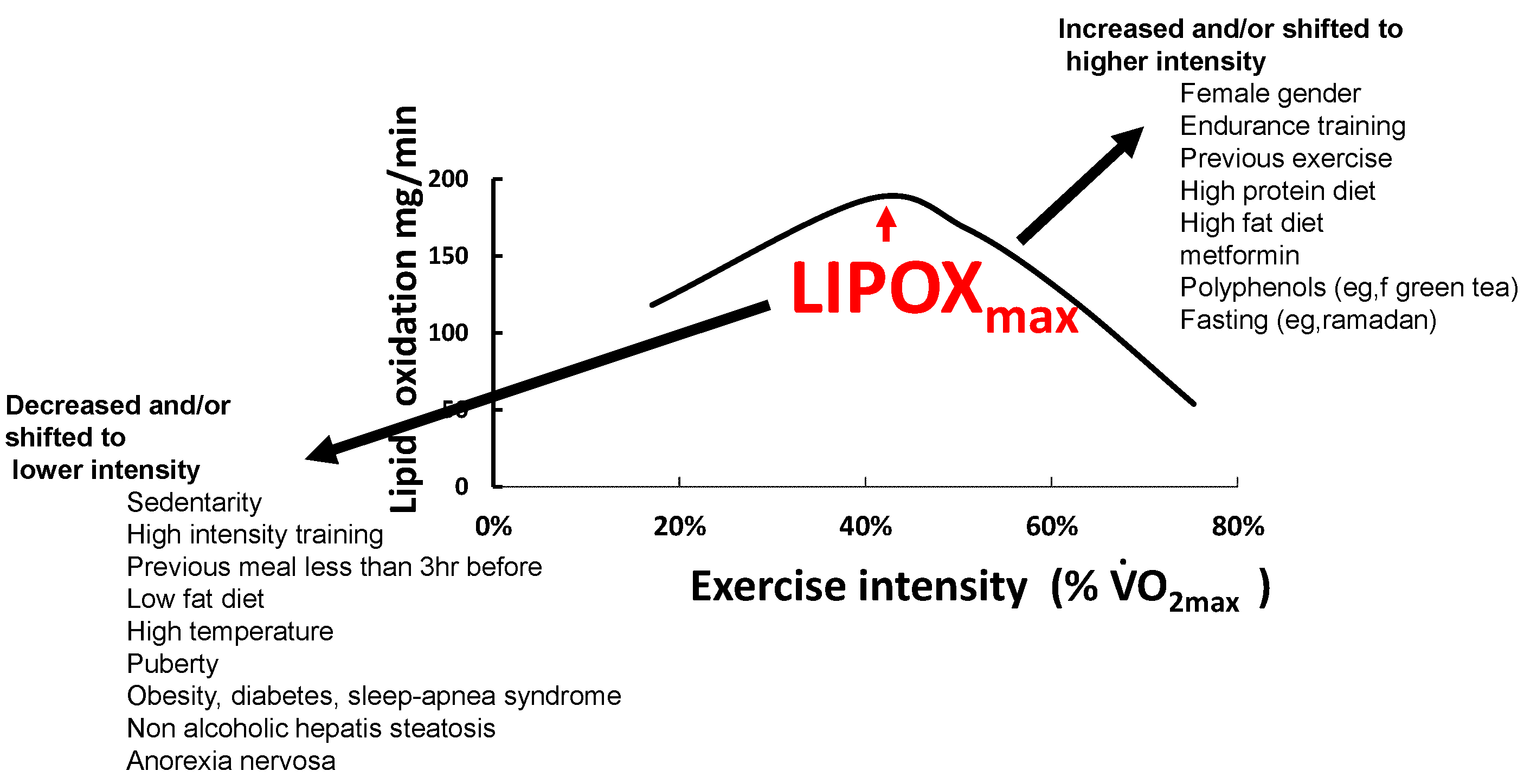{kind=link}
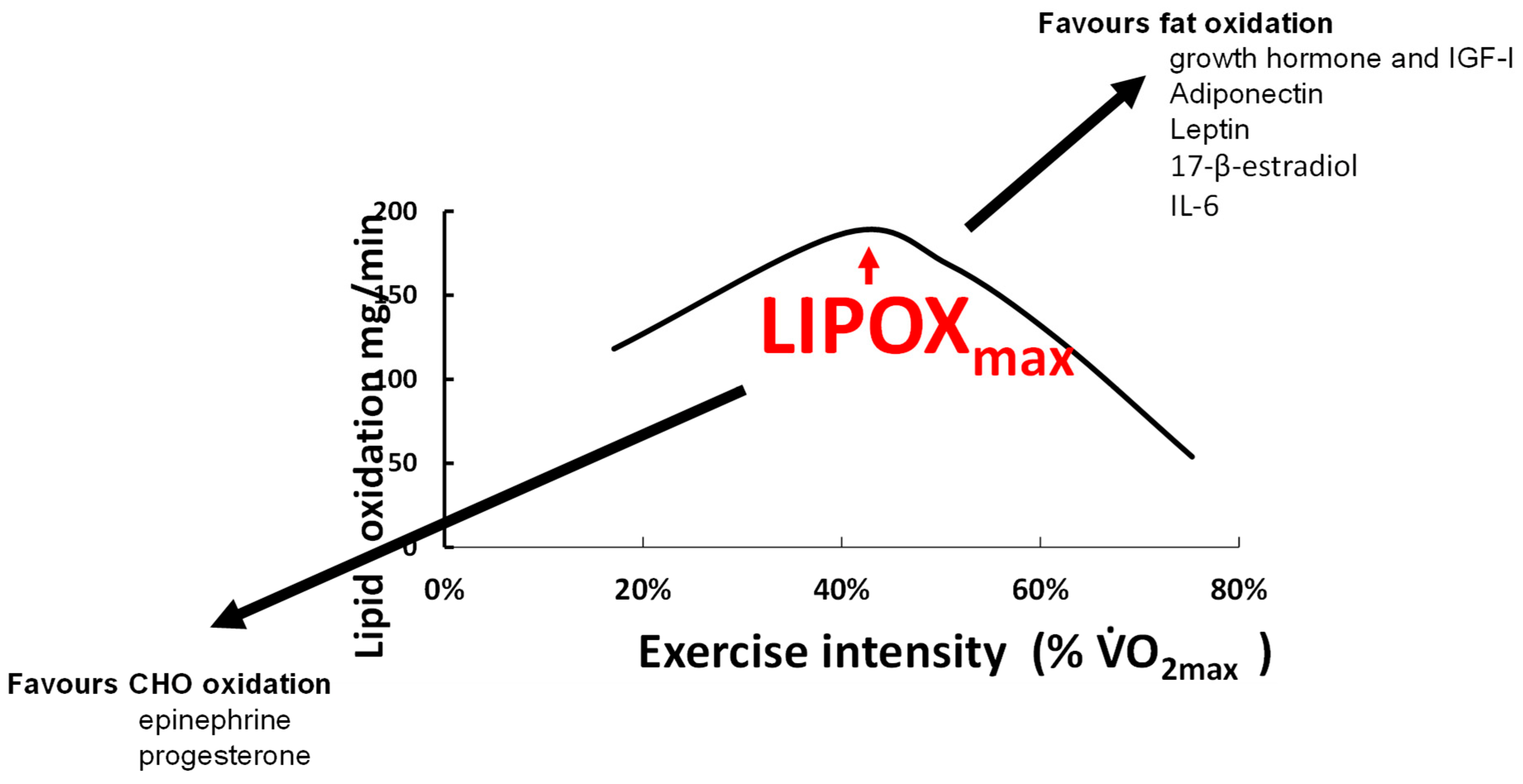{kind=link}
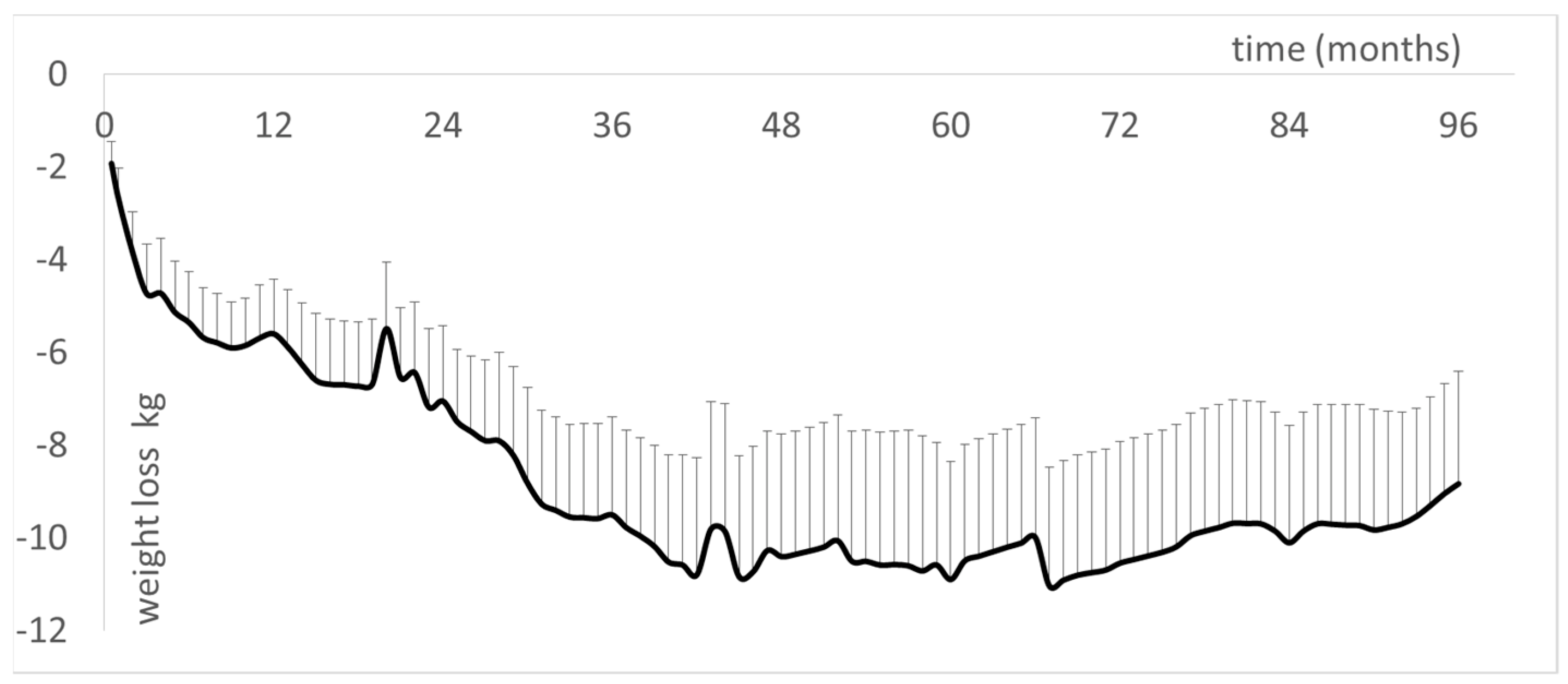{kind=link}
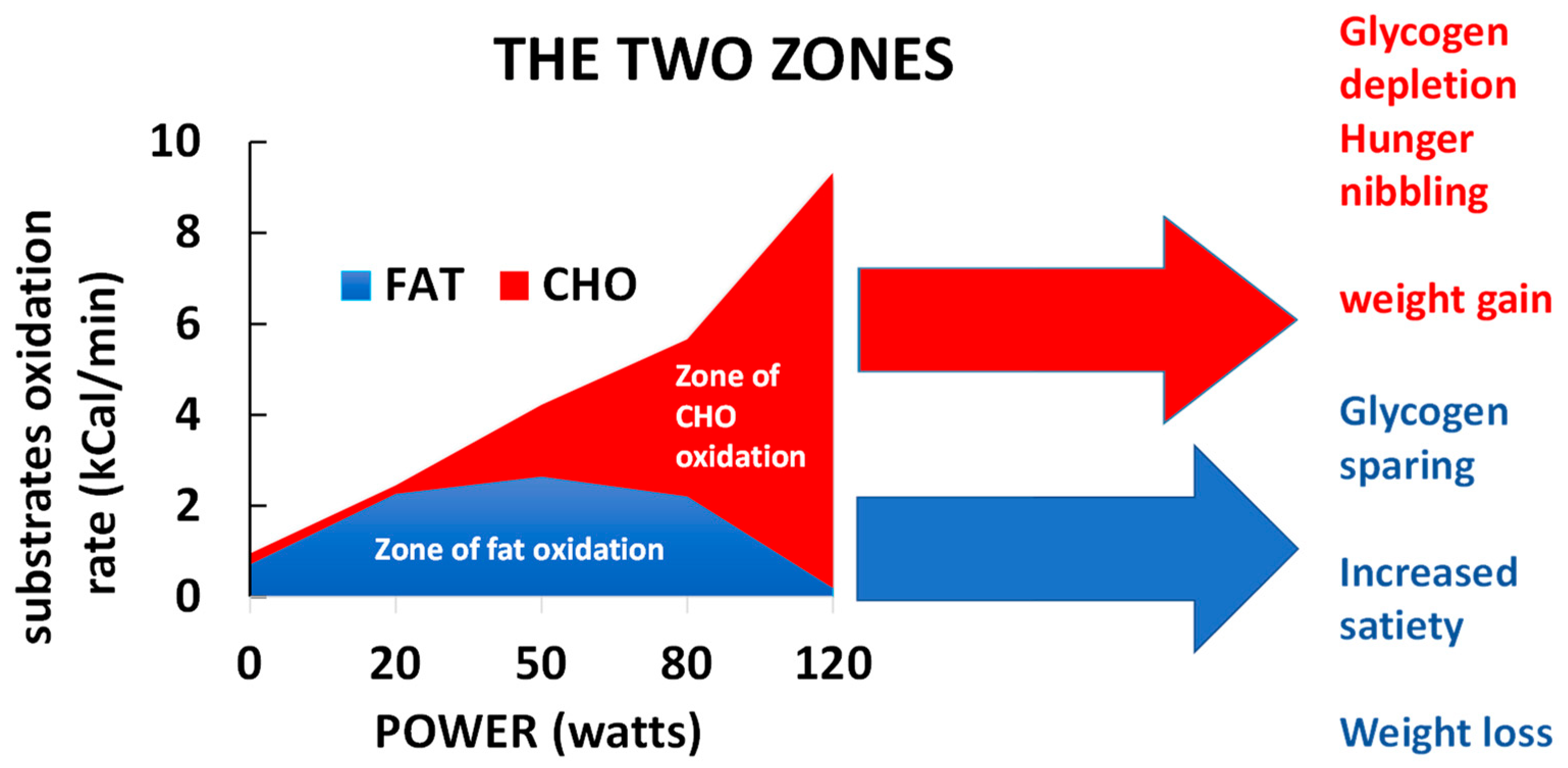{kind=link}
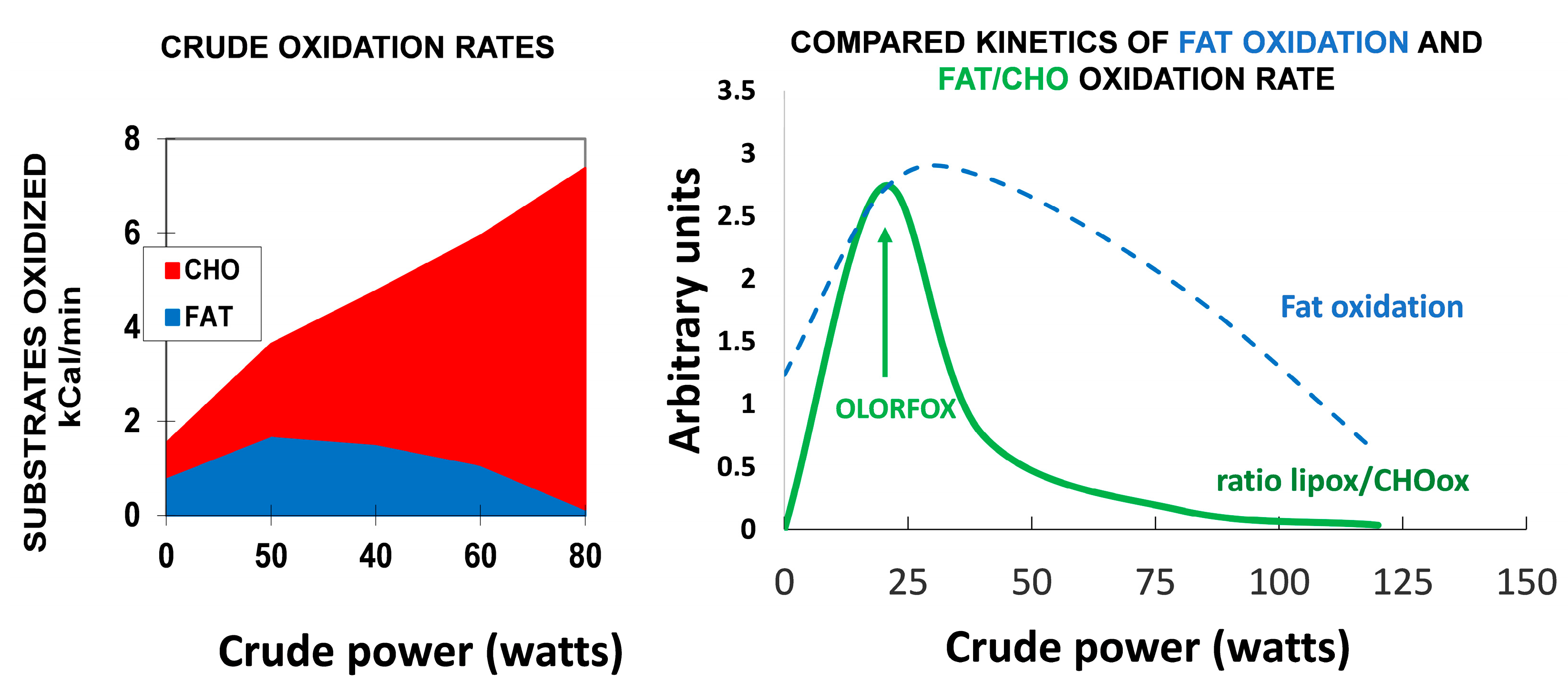{kind=link}
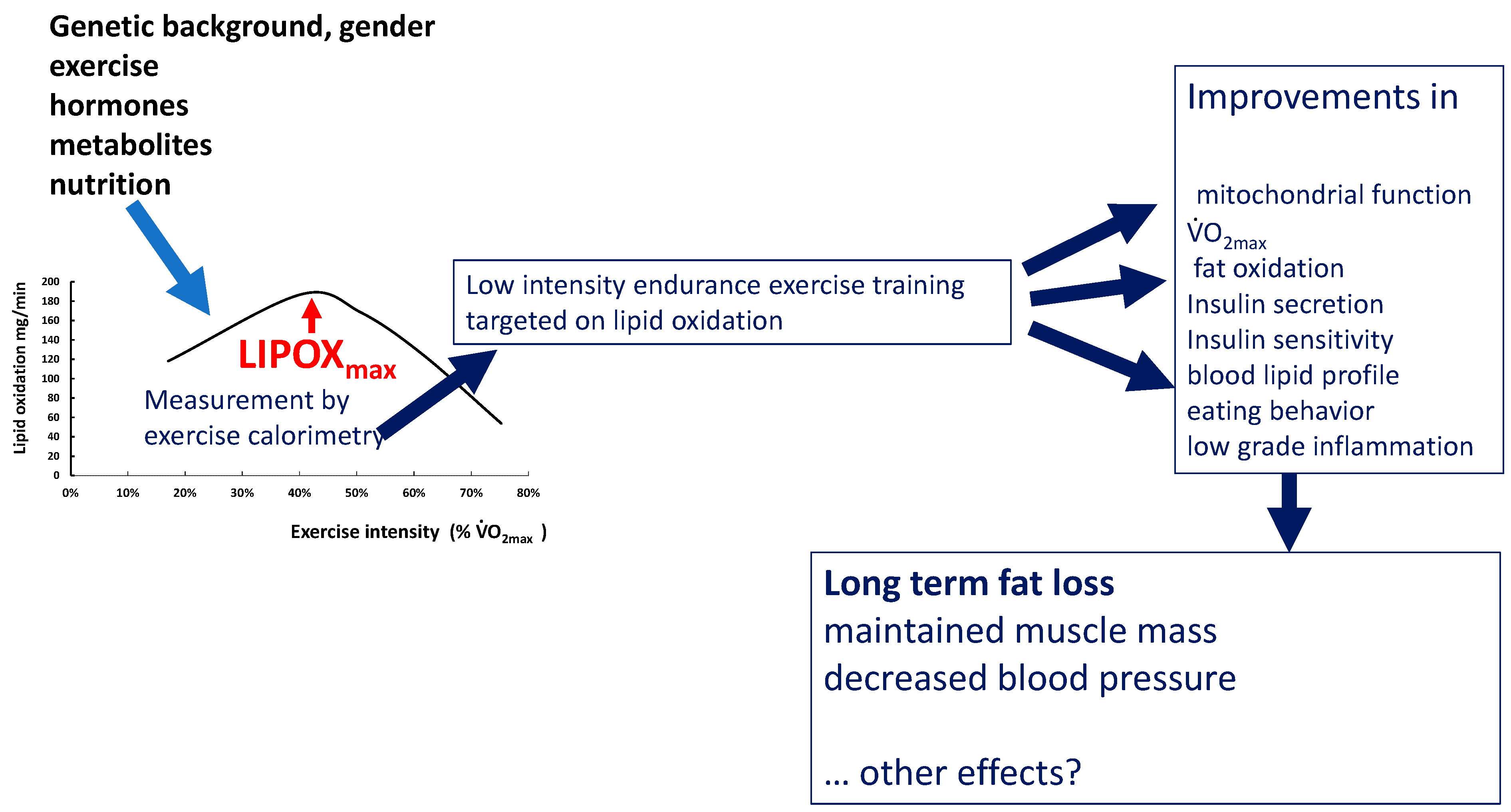{kind=link}
| Modifying Factor | Effect | References |
|---|---|---|
| Previous meal taken less than 3 h before | Decreased MFO and shifted LIPOXmax to a slightly lower intensity | [5,6,111,112,113] |
| Dietary carbohydrate and fat intake | Dietary carbohydrate and fat intake make modest but independent contributions to the interindividual variability in the capacity to oxidize fat during exercise. | [114] |
| Polyphenols of green tea | They increase maximal fat oxidation and shifts the point where fat is no longer oxidized to higher intensity levels | [115,116] |
| Anthocyanins from Prunus cerasus L. | Reported to be capable of augmenting fat oxidation but do not modify MFO | [117] |
| Low-fat diet | Decreases fat oxidation during exercise | [118] |
| Previous exercise performed just before the exercise calorimetry | 1 h single bout of moderate-intensity exercise slightly increases MFO | [119] |
| Puberty | LIPOXmax and MFO are higher in prepubertal children and gradually decrease throughout puberty to reach adult values at the end of puberty | [120,121,122,123] |
| Type of exercise | Higher during running than cycling in adults and in pre- to early pubertal children | [63,122] |
| Ramadan | At the end of Ramadan, subjects had increased their fat utilization during exercise, with a right-shift of the cross-over point and the LIPOXmax and an increase in MFO | [124] |
| Gender | Women oxidize slightly more lipids, and on average, their LIPOXmax occurs at higher relative output. Estradiol and progesterone seem to have opposite effects, with estradiol eliciting greater lipid oxidation. Greater use of fat and reduced amount of CHO usage during the luteal vs. follicular phase, directly related to the change in estradiol. | [122,125,126,127,128,129,130,131,132] |
| Temperature | Shift to preferential CHO oxidation during exercise in hot environments. Reversal after acclimation and training. | [133,134] |
| Highly trained athletes | Most of them exhibit a markedly high ability to oxidize lipids during exercise but in some sports such as soccer, preferential use of CHO is often observed | [5,59,135,136,137] |
| Obesity and diabetes | LIPOXmax values markedly shifted to lower power intensities and MFO decreased. After bariatric surgery the LIPOXmax is shifted to lower intensities. | [32,138] |
| Hypoxia | Exposure to hypoxia did not induce a consistent change in the balance of substrates during exercise compared with normoxia | [139] |
| Cardiometabolic risk factors | Increased waist circumference and plasma triacylglycerols are associated with impaired lipid oxidation | [140] |
| Non-alcoholic hepatic steatosis | Inverse correlation between LIPOXmax and the fatty liver index | [141] |
| Sedentary time, and physical activity time | Sedentary impairs, while physical activity improves, MFO and LIPOXmax. More studies needed. | [142] |
| Cardiorespiratory fitness (CRF) | MFO and Fatmax are positively correlated with VO2max | [143] |
| Metformin | Increases fat oxidation during exercise and decreases its postexercise rise | [144] |
| Type 2 diabetes | Lower ability to oxidize lipids when compared to subjects matched for body mass index (difference not found by all authors) | [145,146] |
| Type 1 diabetes | Lower ability to oxidize lipids | [147,148] |
| Sleep apnea syndrome | Lower ability to oxidize lipids during exercise. Training improves both apnea index and lipid oxidation during exercise (MFO and LIPOXmax values) | [149] |
| Fasting | Peak fat oxidation increased in prolonged fasted state and this was highly correlated with plasma free fatty acids concentrations. | [150] |
| Protein intake | A moderate increase in daily protein intake up to 1 g·kg−1·d−1 improves the maximal ability to oxidize fat during exercise. | [151] |
| Anorexia nervosa | Lower ability to oxidize fat during exercise due to the decrease in muscle mass | [152] |
| Author | Population | Duration of Study | Average Weight Loss |
|---|---|---|---|
| Dumortier et al. (2002) [7] | 21 metabolic syndrome | 2 months | −2.5 kg |
| Brandou et al. (2003) [8] | 14 obese adolescents | 2 months | −3.72 kg |
| Dumortier et al. (2003) [209] | 28 metabolic syndrome | 2 months | −2.6 kg |
| Brandou et al. (2005) [210] | 7 obese adolescents | 3 months | −5.2 kg |
| Jean et al. (2006) [211] | 28 type 2 diabetics | 3 months | −1.3 kg |
| Ben Ounis et al. (2008) [212] | 8 obese adolescents | 2 months | −11.5 kg |
| Fedou et al. (2008) [213] | 10 HIV-infected patients under antiretroviral therapy | 12 months | −0.92 kg |
| Bordenave et al. (2008) [86] | 11 type 2 diabetics | 2 months | No weight change |
| Venables et al. (2008) [214] | 8 obese adults | 2 months | −0.2 kg |
| Ben Ounis et al. (2008) [212] | 8 obese boys | 2 months | −1.90 kg |
| Ben Ounis et al. (2008) [215] | 6 obese girls | 2 months | −1.40 kg |
| Ben Ounis et al. (2009) [216] | 18 obese adolescents | 2 months | −6 kg |
| Ben Ounis et al. (2009) [217] | 9 obese adolescents | 2 months | −9.5 kg |
| Romain et al. (2009) [218] | 17 psychiatric patients under neuroleptics | 3 months | −2.9 kg |
| Mogensen et al. (2009) [146] | 12 type 2 diabetic patients | 2.5 months | No weight change |
| Elloumi et al. (2009) [219] | 7 obese adolescent boys | 2 months | −1.7 kg |
| Elloumi et al. (2009) [219] | 7 obese adolescents | 2 months | −12.3 kg |
| Maurie et al. (2011) [220] | 39 type 2 diabetics | 3 months | −2.23 kg |
| Tan et al. (2016) [221] | 29 obese women (20–23) | 10 weeks | −4 kg |
| Besnier et al. (2015) [222] | 33 overweight and obese women | 5 months | −5 kg |
| Tan et al. (2016) [223] | 15 middle aged women | 10 weeks | −3 kg |
| Tan et al. (2016) [224] | 11 boys (9.0 ± 1.0) | 10 weeks | −1 kg |
| Tan et al. (2018) [225] | 16 elderly women with T2D | 8 weeks | −2.4 kg |
| Cao et al. (2019) [226] | 13 overweight and obese women | 10 weeks | −4.6 kg |
| Zeng et al. (2020) [227] | 18 young obese women | 12 weeks | NR |
| Jiang et al. (2020) [228] | 13 elderly women with T2D | 16 weeks | −2.10 kg |
| Jiang et al. (2020) [228] | 14 elderly men with T2D | 16 weeks | −3.3 kg |
| Hammoudi et al. (2020) [18] | 61 obese women | 2 years | −6 kg |
| Brun et al. (2020) [229] | 49 obese | 6 years | −6.7 kg |
| Guedjati et al. (2020) [230] | 21 middle aged obese women | 3 weeks | −2 kg |
| Bughin et al. (2021) [231] | 14 obese followed by telerehabilitation | 12 weeks | −0.8 kg |
| Brun et al. (2022) [33] | 10 obese | 8 years | −9.1 kg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brun, J.-F.; Myzia, J.; Varlet-Marie, E.; Raynaud de Mauverger, E.; Mercier, J. Beyond the Calorie Paradigm: Taking into Account in Practice the Balance of Fat and Carbohydrate Oxidation during Exercise? Nutrients 2022, 14, 1605. https://doi.org/10.3390/nu14081605
Brun J-F, Myzia J, Varlet-Marie E, Raynaud de Mauverger E, Mercier J. Beyond the Calorie Paradigm: Taking into Account in Practice the Balance of Fat and Carbohydrate Oxidation during Exercise? Nutrients. 2022; 14(8):1605. https://doi.org/10.3390/nu14081605
Chicago/Turabian StyleBrun, Jean-Frédéric, Justine Myzia, Emmanuelle Varlet-Marie, Eric Raynaud de Mauverger, and Jacques Mercier. 2022. "Beyond the Calorie Paradigm: Taking into Account in Practice the Balance of Fat and Carbohydrate Oxidation during Exercise?" Nutrients 14, no. 8: 1605. https://doi.org/10.3390/nu14081605
APA StyleBrun, J.-F., Myzia, J., Varlet-Marie, E., Raynaud de Mauverger, E., & Mercier, J. (2022). Beyond the Calorie Paradigm: Taking into Account in Practice the Balance of Fat and Carbohydrate Oxidation during Exercise? Nutrients, 14(8), 1605. https://doi.org/10.3390/nu14081605