Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method

Abstract
:1. Introduction
2. Materials and Methods
3. Results
3.1. Sources of Astaxanthin
3.2. Differential Activity of Astaxanthin Stereoisomers
3.3. Mechanisms of Astaxanthin Bioactivity
3.4. Evolution of Carotenoids
4. Discussion
4.1. Mitochondrial Targeting Makes Astaxanthin an Exceptionally Effective Antioxidant
4.2. Mitochondrial Dysfunction Is at the Core of Many Contemporary Diseases
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef]
- Ambati, R.R.; Moi, P.S.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Oslan, S.N.H.; Tan, J.S.; Oslan, S.N.; Matanjun, P.; Mokhtar, R.A.M.; Shapawi, R.; Huda, N. Haematococcus pluvialis as a Potential Source of Astaxanthin with Diverse Applications in Industrial Sectors: Current Research and Future Directions. Molecules 2021, 26, 6470. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Bird, M.I.; Crabtree, S.A.; Haig, J.; Ulm, S.; Wurster, C.M. A global carbon and nitrogen isotope perspective on modern and ancient human diet. Proc. Nat. Acad. Sci. USA 2021, 118, e2024642118. [Google Scholar] [CrossRef]
- Kim, S.H.; Kim, H. Astaxanthin modulation of signaling pathways that regulate autophagy. Mar. Drugs 2019, 17, 546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stachowiak, B.; Szulc, P. Astaxanthin for the food industry. Molecules 2021, 26, 2666. [Google Scholar] [CrossRef] [PubMed]
- Perera, C.O.; Yen, G.M. Functional properties of carotenoids in human health. Int. J. Food Prop. 2007, 10, 201–230. [Google Scholar] [CrossRef]
- Domínguez-Bocanegra, A.R.; Guerrero Legarreta, I.; Martinez Jeronimo, F.; Tomasini Campocosio, A. Influence of environmental and nutritional factors in the production of astaxanthin from Haematococcus pluvialis. Bioresour. Technol. 2004, 92, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Tsubokura, A.; Yoneda, H.; Mizuta, H. Paracoccus carotinifaciens sp. nov., a new aerobic gram-negative astaxanthin-producing bacterium. Int. J. Syst. Evol. Microbiol. 1999, 49, 277–282. [Google Scholar] [CrossRef]
- Pan, C.H.; Chien, Y.H. Concentration and composition of astaxanthin in black tiger prawn Penaeus monodon post-larvae fed Artemia sp. Nauplii or mauxia Shrimsp acetes intermedius. J. World Aquac. Soc. 2003, 34, 57–65. [Google Scholar] [CrossRef]
- Parisenti, J.; Beirão, L.H.; Maraschin, M.; Mourino, J.L.; Nascimento Vieira, F.D.; Bedin, L.H.; Rodrigues, E. Pigmentation and carotenoid content of shrimp fed with Haematococcus pluvialis and soy lecithin. Aquac. Nutr. 2011, 17, e530–e535. [Google Scholar] [CrossRef]
- Capelli, B.; Bagchi, D.; Cysewski, G.R. Synthetic astaxanthin is significantly inferior to algal-based astaxanthin as an antioxidant and may not be suitable as a human nutraceutical supplement. Nutrafoods 2013, 12, 145–152. [Google Scholar] [CrossRef]
- Kusdiyantini, E.; Gaudin, P.; Goma, G.; Blanc, P.J. Growth kinetics and astaxanthin production of Phaffia rhodozyma on glycerol as a carbon source during batch fermentation. Biotechnol. Lett. 1998, 20, 929–934. [Google Scholar] [CrossRef]
- Visioli, J.F.; Artaria, C. Astaxanthin in cardiovascular health and disease: Mechanisms of action, therapeutic merits, and knowledge gaps. Food Funct. 2017, 8, 39–63. [Google Scholar] [CrossRef] [PubMed]
- Shah, M.M.R.; Liang, Y.; Cheng, J.J.; Daroch, M. Astaxanthin-producing green microalga Haematococcus pluvialis: From single cell to high value commercial products. Front. Plant Sci. 2016, 7, 531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mascia, F.; Girolomoni, L.; Alcocer, M.J.P.; Bargigia, I.; Perozeni, F.; Cazzaniga, S.; Cerullo, G.; D’Andrea, C.; Ballottari, M. Functional analysis of photosynthetic pigment binding complexes in the green alga Haematococcus pluvialis reveals distribution of astaxanthin in photosystems. Sci. Rep. 2017, 7, 16319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Ye, Z.; Wang, M.; Manzoor, M.F.; Aadil, R.M.; Tan, X.; Liu, Z. Comparison of different methods for extracting the astaxanthin from Haematococcus pluvialis: Chemical composition and biological activity. Molecules 2021, 26, 3569. [Google Scholar] [CrossRef]
- Yang, L.; Qiao, X.; Gu, J.; Li, X.; Cao, Y.; Xu, J.; Xue, C. Influence of molecular structure of astaxanthin esters on their stability and bioavailability. Food Chem. 2021, 343, 128497. [Google Scholar] [CrossRef] [PubMed]
- Carberry, J.; Carberry, M.J.; Tagrin, D.S. Sustainable Nutrition. U.S. Patent 10227557B2, 12 March 2019. [Google Scholar]
- Ding, L.; Yang, J.; Yin, K.; Cheng, H.; Li, J.; Xue, C. The spatial arrangement of astaxanthin in bilayers greatly influenced the structural stability of DPPC liposomes. Colloids Surf. B Biointerfaces 2022, 212, 112383. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, F.; Zhang, Y.; Tan, X.; Lou, P.; Liu, H. Analysis of astaxanthin molecular targets based on network pharmacological strategies. J. Food Biochem. 2021, 45, e13717. [Google Scholar] [CrossRef] [PubMed]
- Snell, T.W.; Harder, S.E.; Carberry, M.; Carberry, J. Importance of astaxanthin as a dietary supplement in aquaculture. Hatch. Feed. Manag. 2019, 7, 31–33. [Google Scholar]
- Snell, T.W.; Carberry, M.; Carberry, J.; Wilson, T. Astaxanthin: An important micro-ingredient in aquaculture feeds. Aquafeed 2022, in press. [Google Scholar]
- Johnston, R.K.; Siegfried, E.J.; Snell, T.W.; Carberry, J.; Carberry, M.; Brown, C.; Farooq, S. Effects of astaxanthin on Brachionus manjavacas (Rotifera) population growth. Aquac. Res. 2018, 49, 2278–2287. [Google Scholar] [CrossRef] [Green Version]
- Mashhadi, N.S.; Zakerkish, M.; Mohammadiasl, J.; Zarei, M.; Mohammadshahi, M.; Haghighizadeh, M.H. Astaxanthin improves glucose metabolism and reduces blood pressure in patients with type 2 diabetes mellitus. Asia Pac. J. Clin. Nutr. 2018, 27, 341–346. [Google Scholar] [PubMed]
- Park, J.; Chyun, J.; Kim, Y.; Line, L.L.; Chew, B.P. Astaxanthin decreased oxidative stress and inflammation and enhanced immune response in humans. Nutr. Metab. 2010, 7, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNulty, H.P.; Byun, J.; Lockwood, S.F.; Jacob, R.F.; Mason, R.P. Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochim. Biophys. Acta Biomembr. 2007, 1768, 167–174. [Google Scholar] [CrossRef] [Green Version]
- Wolf, A.M.; Asoh, S.; Hiranuma, H.; Ohsawa, I.; Iio, K.; Satou, A.; Ishikura, M.; Ohta, S. Astaxanthin protects mitochondrial redox state and functional integrity against oxidative stress. J. Nutr. Biochem. 2010, 21, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Kim, H. Inhibitory Effect of Astaxanthin on Oxidative Stress-Induced Mitochondrial Dysfunction-A Mini-Review. Nutrients 2018, 10, 1137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landon, R.; Gueguen, V.; Petite, H.; Letourneur, D.; Pavon-Djavid, G.; Anagnostou, F. Impact of astaxanthin on diabetes pathogenesis and chronic complications. Mar. Drugs 2020, 18, 357. [Google Scholar] [CrossRef] [PubMed]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of coevolution towards plant photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef]
- Coesel, S.; Obornı´k, M.; Varela, J.; Falciatore, A.; Bowler, C. Evolutionary origins and functions of the carotenoid biosynthetic pathway in marine diatoms. PLoS ONE 2008, 3, e2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Kim, J.H.; Kim, S.W. Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects. Mar. Drugs 2014, 12, 4810–4832. [Google Scholar] [CrossRef]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef] [Green Version]
- Le Moal, E.; Juban, G.; Bernard, A.S.; Varga, T.; Policar, C.; Chazaud, B.; Mounier, R. Macrophage-derived superoxide production and antioxidant response following skeletal muscle injury. Free Radic. Biol. Med. 2018, 120, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Hecht, K.A.; Schnackenberg, J.; Nair, A.; Lignell, A. Astaxanthin for improved muscle function and enhanced physical performance. In Global Perspectives on Astaxanthin; Academic Press: Cambridge, MA, USA, 2021. [Google Scholar] [CrossRef]
- Ferro, M.; Graubard, A.; Beauchesne, T. Astaxanthin: The possible effects of this carotenoid on disease, inflammation and aging control. A meta-analysis. Int. J. Food Nutr. Res. 2021, 5, 44. [Google Scholar]
- Sztretye, M.; Dienes, B.; Gönczi, M.; Czirják, T.; Csernoch, L.; Dux, L.; Szentesi, P.; Keller-Pintér, A. Astaxanthin: A Potential Mitochondrial Targeted Antioxidant Treatment in Diseases and with Aging. Oxid Med. Cell Longev. 2019, 2019, 3849692. [Google Scholar] [CrossRef] [Green Version]
- Krestinina, O.; Baburina, Y.; Krestinin, R.; Odinokova, I.; Fadeva, I.; Sotnikova, L. Astaxanthin Prevents Mitochondrial Impairment Induced by Isoproterenol in Isolated Rat Heart Mitochondria. Antioxidants 2020, 9, 262. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.D.; Kang, H.E.; Yang, S.H.; Lee, M.G.; Shin, W.G. Pharmacokinetics and first-pass metabolism of astaxanthin in rats. Br. J. Nutr. 2011, 105, 220–227. [Google Scholar] [CrossRef] [Green Version]
- Ekpe, L.; Inaku, K.; Ekpe, V. Antioxidant effects of astaxanthin in various diseases—A review. J. Mol. Pathophysiol. 2018, 7, 1–6. [Google Scholar] [CrossRef]
- Kidd, P. Astaxanthin- cell membrane and nutrient with diverse clinical benefits and anti-ageing potentials. Altern. Med. Rev. 2011, 16, 255–264. [Google Scholar]
- Fassett, R.G.; Coombes, J.S. Astaxanthin in cardiovascular health and diseases. Molecules 2012, 17, 2030–2048. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Yin, J.; Chen, J.; Ma, X.; Wu, M.; Liu, G.; Yao, G.; Tan, B.; Yin, Y. Mitochondria-Targeted Antioxidants: A Step towards Disease Treatment. Oxidative Med. Cell. Longev. 2020, 2020, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Kohandel, Z.; Farkhondeh, T.; Aschner, M.; Pourbagher-Shahri, A.M.; Samarghandian, S. Anti-inflammatory action of astaxanthin and its use in the treatment of various diseases. Biomed. Pharmacother. 2021, 145, 112179. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; De Bittencourt, P.I.H. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhu, J.; Yin, W.; Ding, X. Astaxanthin improves cognitive deficits from oxidative stress, nitric oxide synthase and inflammation through upregulation of PI3K/Akt in diabetes rat. Int. J. Clin. Exp. Pathol. 2015, 8, 6083–6094. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Stereoisomer | Primary Source | Localization | Bioavailability | Bioactivity |
|---|---|---|---|---|
3S,3′S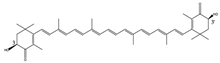 | Haematococcus pluvialis [10] Paracoccus carotinifaciens [11] Salmo salar [2] | Mitochondria, membrane insertion | high | Anti-oxidation Anti-inflammatory Facilitates mitochondria function Strengthens immunity Promotes fertility UV protection Colorant |
3R,3′S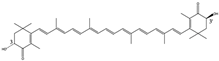 | Penaeus monodon [12] Litopenaeus vannamei [13] Synthetic—petroleum [14] | Float freely in cells | low | Anti-oxidation Colorant |
3R,3′R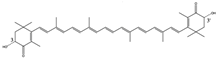 | Phaffia rhodozyma [15] Euphausia superba [16] | Float freely in cells | low | Anti-oxidation Colorant |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Snell, T.W.; Carberry, J. Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method. Nutrients 2022, 14, 1522. https://doi.org/10.3390/nu14071522
Snell TW, Carberry J. Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method. Nutrients. 2022; 14(7):1522. https://doi.org/10.3390/nu14071522
Chicago/Turabian StyleSnell, Terry W., and John Carberry. 2022. "Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method" Nutrients 14, no. 7: 1522. https://doi.org/10.3390/nu14071522
APA StyleSnell, T. W., & Carberry, J. (2022). Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method. Nutrients, 14(7), 1522. https://doi.org/10.3390/nu14071522