
The Tobacco β-Cembrenediol: A Prostate Cancer Recurrence Suppressor Lead and Prospective Scaffold via Modulation of Indoleamine 2,3-Dioxygenase and Tryptophan Dioxygenase

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
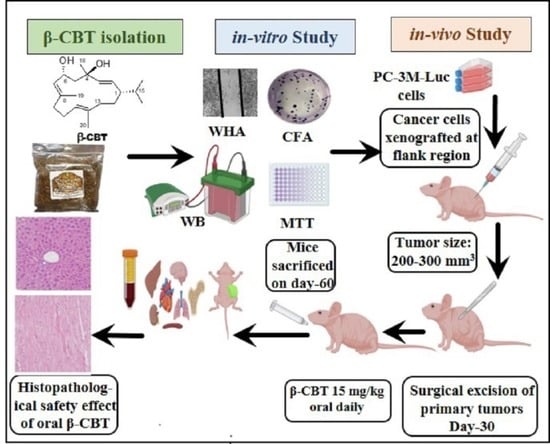
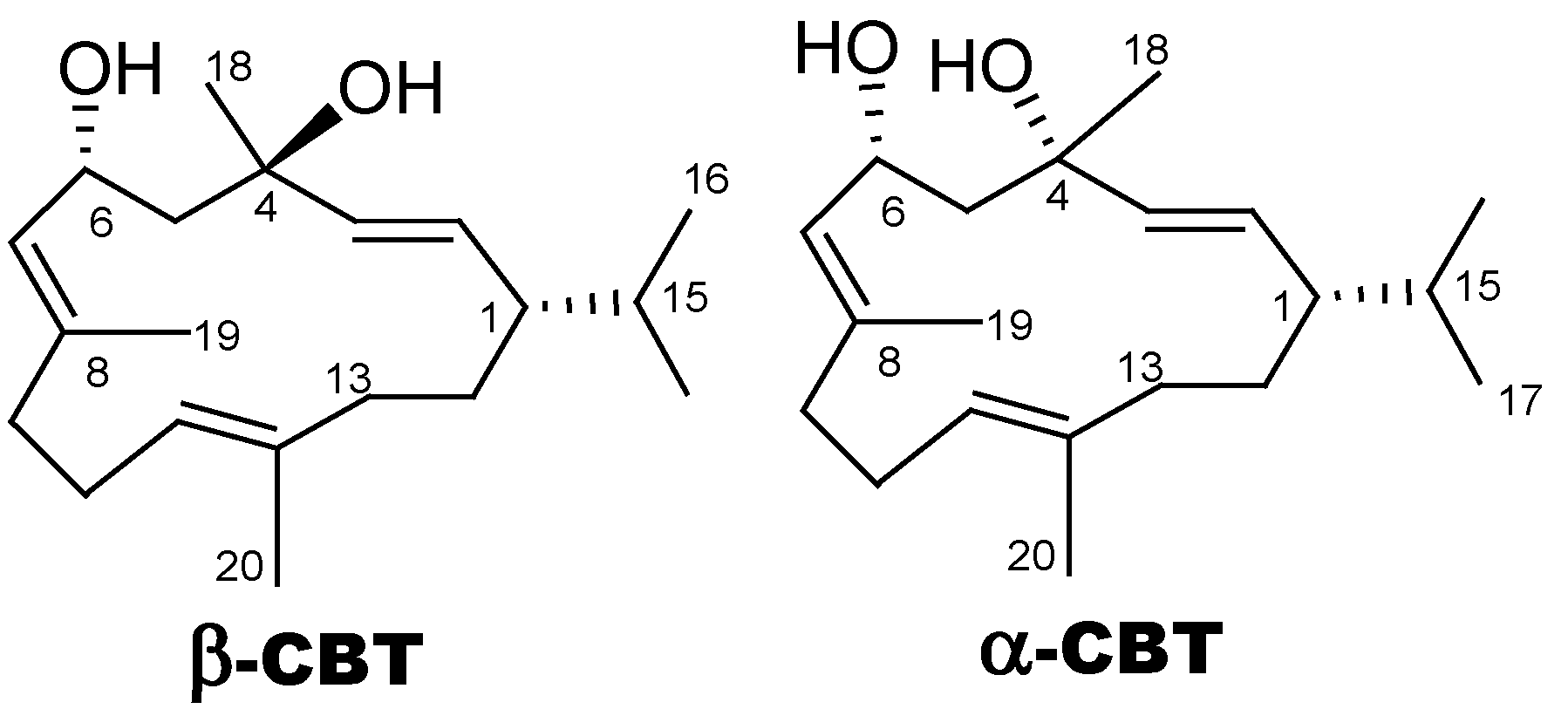
2.1. β-CBT Isolation
2.2. Cell Lines and Culture Conditions
2.3. Compound Preparation and Stock Solution
2.4. Assesment of β-CBT Effects on PC Cells Viability
2.5. Wound-Healing Assay
2.6. Colony Formation Assay
2.7. Western Blot Analysis
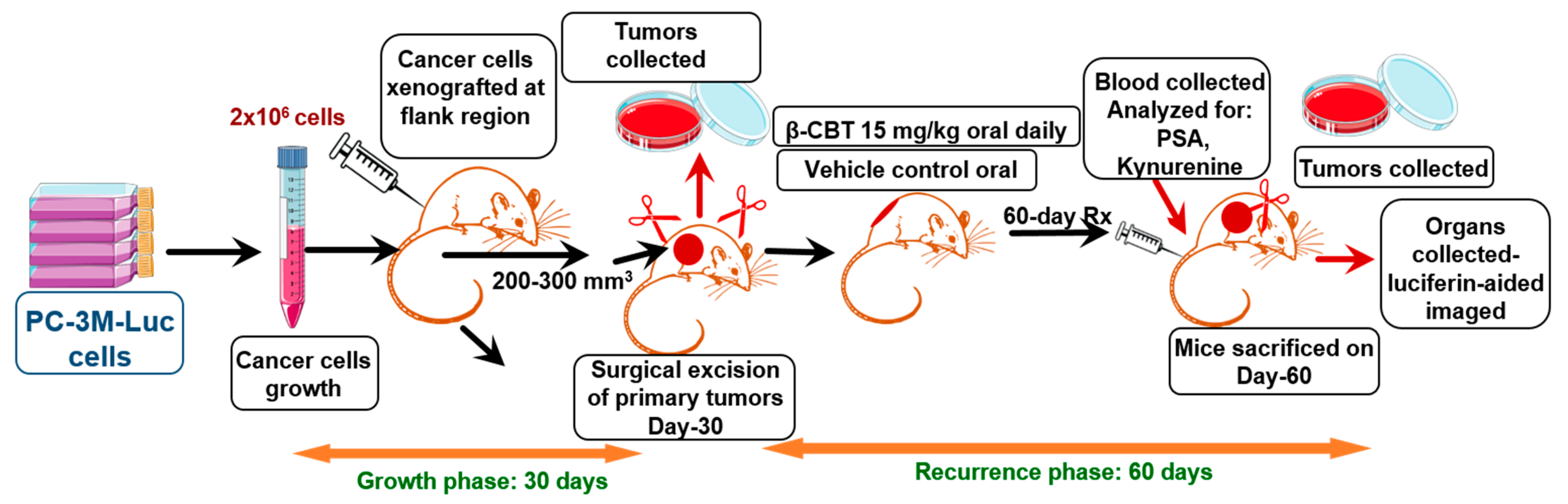
2.8. Animal Model and Treatment Mode
2.9. Aimals Plasma PSA Level
2.10. Animals Plasma Kyn Level
2.11. Statisticcal Analysis
3. Results
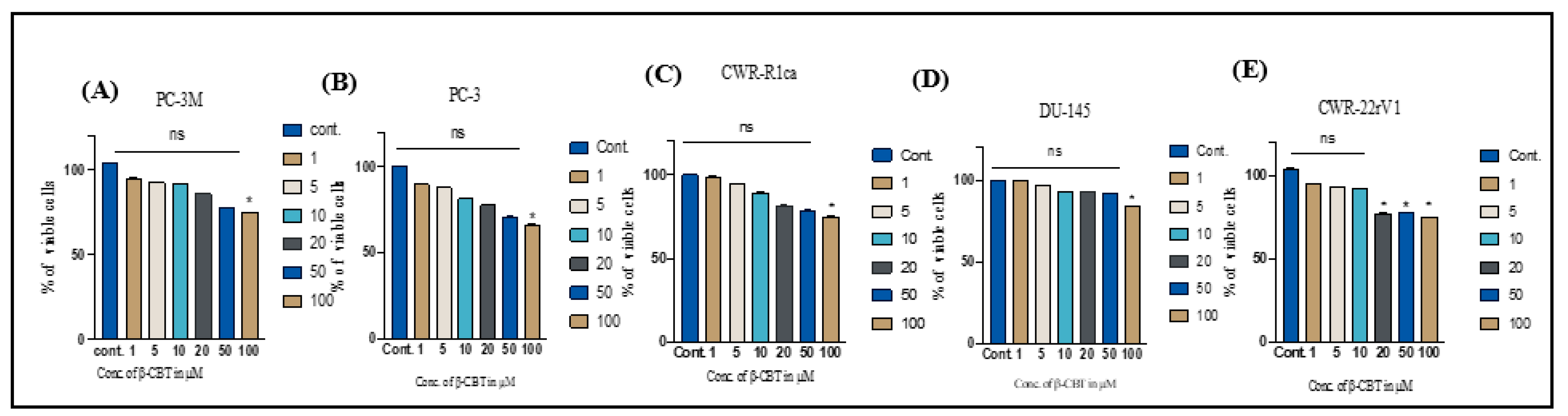
3.1. Effects of β-CBT on the Viability of Different Human PC Cells
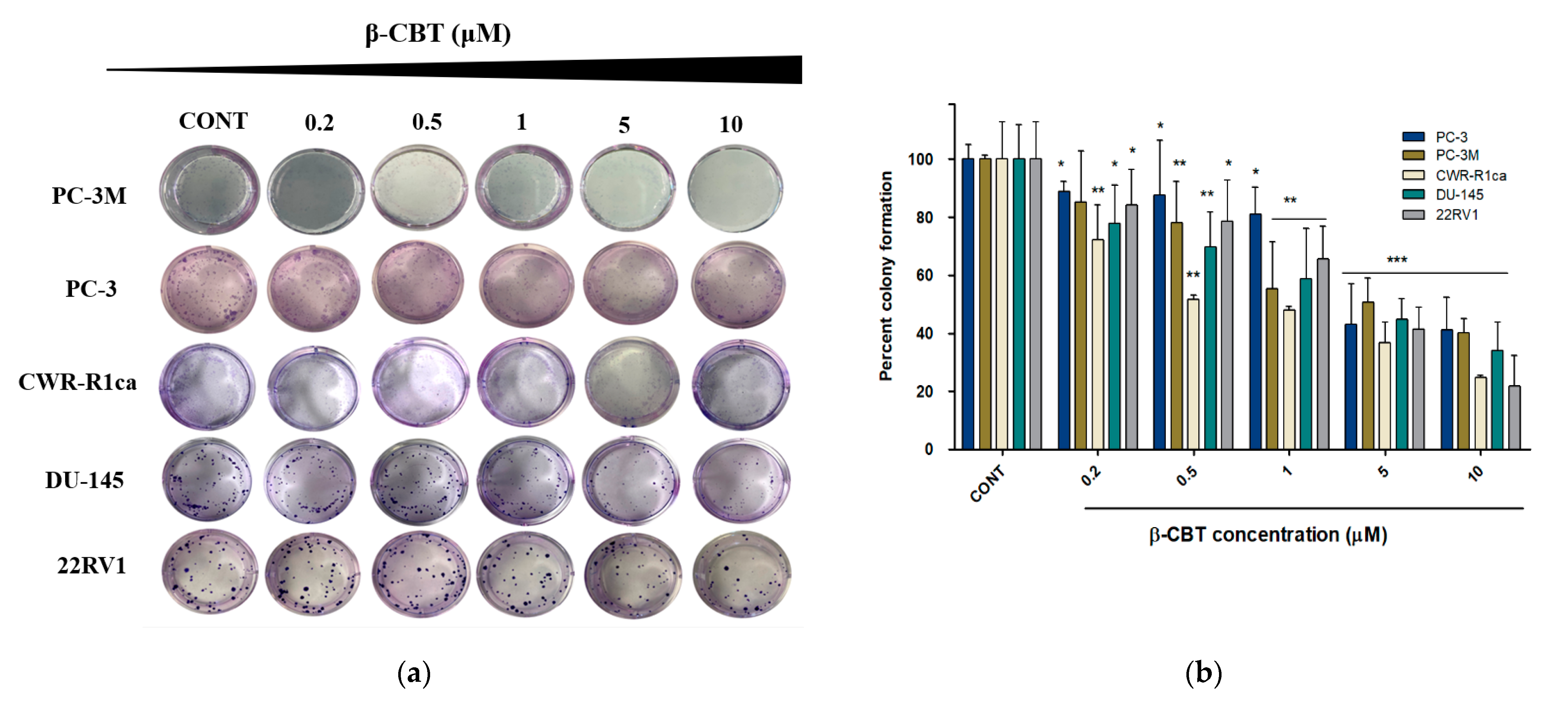
3.2. β-CBT Reduced the Migration and Colony Formation in Different PC Cells
3.3. β-CBT Reduced IDO1 and TDO2 Expression In Vitro in PC-3M Cell Line
3.4. β-CBT Inhibited PC-3M-Cells Locoregional Recurrences after Primary Tumor Surgical Excision and Tumor Distant Recurrence
3.5. β-CBT Reduced Mice Plasma Total Prostate-Specific Antigen (PSA)
3.6. β-CBT Reduced Mice Plasma Kynurenine Level
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- American Cancer Society. Cancer Facts & Figures. 2022. Available online: https://www.cancer.org/latest-news/facts-and-figures-2022.html (accessed on 14 February 2022).
- Smith, S.M.; Wachter, K.; Burris, H.A., III; Schilsky, R.L.; George, D.J.; Peterson, D.E.; Johnson, M.L.; Markham, M.J.; Mileham, K.F.; Beg, M.S.; et al. Clinical Cancer Advances 2021: ASCO’s Report on Progress against Cancer. J. Clin. Oncol. 2021, 39, 1165–1184. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, K.A.; Sylvester, P.W. Biocatalytic and semisynthetic studies of the anticancer tobacco cembranoids. Expert Opin. Investig. Drugs 2007, 16, 877–887. [Google Scholar] [CrossRef]
- Wahlberg, I.; Enzell, C.R. Tobacco cembranoids. Beitr. Zur Tab. Int. 1984, 12, 93–104. [Google Scholar] [CrossRef][Green Version]
- Wahlberg, A.I.; Eklund, A.M. Cembrane-derived aroma compounds in tobacco and their biogenesis. In Trends in Flavour Research; Maarse, H., van der Heji, D.G., Eds.; Elsevier Science B.V.: Oxford, UK, 1994; pp. 449–462. [Google Scholar]
- Olsson, E.; Berg, J.E.; Wahlberg, I. Tobacco chemistry. 79. Eight new cembranoids from tobacco. Structural elucidation and conformational studies. Tetrahedron 1993, 49, 4975–4992. [Google Scholar] [CrossRef]
- Ferchmin, P.A.; Hao, J.; Perez, D.; Penzo, M.; Maldonado, H.M.; Gonzalez, M.T.; Rodriguez, A.D.; Jean, D.V. Tobacco cembranoids protect the function of acute hippocampal slices against NMDA by a mechanism mediated by α4β2 nicotinic receptors. J. Neurosci. Res. 2005, 82, 631–641. [Google Scholar] [CrossRef]
- Vélez-Carrasco, W.; Green, C.E.; Catz, P.; Furimsky, A.; O’Loughlin, K.; Eterović, V.A.; Ferchmin, P. Pharmacokinetics and metabolism of 4R-cembranoid. PLoS ONE 2015, 10, e0121540. [Google Scholar] [CrossRef]
- Aqil, F.; Zahin, M.; El Sayed, K.A.; Ahmad, I.; Orabi, K.Y.; Arif, J.M. Antimicrobial, antioxidant, and antimutagenic activities of selected marine natural products and tobacco cembranoids. Drug Chem. Toxicol. 2011, 34, 167–179. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Shi, J.; Xue, S.J.; Zhang, Z. Analyses of effects of α-cembratrien-diol on cell morphology and transcriptome of Valsa mali var. mali. Food Chem. 2017, 214, 110–118. [Google Scholar] [CrossRef]
- Saito, Y.; Takayawa, H.; Konishi, S.; Yoshida, D.; Mizusaki, S. Identification of cembratriene-4,6-diol as anti-tumor-promoting agent from cigarette smoke condensate. Carcinogenesis 1985, 6, 1189–1194. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Nishino, H.; Yoshida, D.; Mizusaki, S.; Ohnishi, A. 12-O-tetradecanoyl-phorbol-13-acetate-stimulated 32Pi incorporation into phospholipids and protein phosphorylation by 2,7,11-cembratriene-4,6-diol, an antitumor-promoting agent. Oncology 1988, 45, 122–126. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Tsujino, Y.; Kaneko, H.; Yoshida, D.; Mizusaki, S. Inhibitory effects of cembratriene-4,6-diol derivatives on the induction of Epstein-Barr virus early antigen by 12-O-tetradecanoylphorbol-13-acetate. Agric. Biol. Chem. 1987, 51, 941–943. [Google Scholar]
- Yuan, X.-L.; Mao, X.-X.; Du, Y.-M.; Yan, P.-Z.; Hou, X.-D.; Zhang, Z.-F. Anti-tumor activity of cembranoid-type diterpenes isolated from Nicotiana tabacum L. Biomolecules 2019, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, H.Y.; Moheyldin, M.M.; Hailat, M.M.; El Sayed, K.A. (1S,2E,4S,7E,11E)-2,7,11-Cembratriene-4,6-diol semisynthetic analogs as novel c-Met inhibitors for the control of c-Met-Dependent breast malignancies. Bioorg. Med. Chem. 2016, 24, 5748–5761. [Google Scholar] [CrossRef]
- Hailat, M.; Ebrahim, H.Y.; Siddique, A.; Goda, A.; El Sayed, K.A. (1S,2E,4S,7E,11E)-2,7,11-cembratriene-4,6-diol as a novel angiogenesis inhibitor for the control of triple negative breast malignancies. Bioorg. Med. Chem. 2017, 25, 3911–3921. [Google Scholar] [CrossRef] [PubMed]
- El Sayed, K.A.; Laphookhieo, S.; Baraka, H.N.; Yousaf, M.; Hebert, A.; Bagaley, D.; Rainey, F.A.; Muralidharan, A.; Thomas, S.; Shah, G.V. Biocatalytic and semisynthetic optimization of the anti-invasive tobacco (1S,2E,4R,6R,7E,11E)-2,7,11-cembratriene-4,6-diol. Bioorg. Med. Chem. 2008, 16, 2886–2893. [Google Scholar] [CrossRef] [PubMed]
- Baraka, H.N.; Khanfar, M.A.; Williams, J.C.; El-Giar, E.M.; El Sayed, K.A. Bioactive natural, biocatalytic, and semisynthetic tobacco cembranoids. Planta Med. 2011, 77, 467–476. [Google Scholar] [CrossRef]
- Opitz, C.A.; Patterson, L.F.S.; Mohapatra, S.R.; Dewi, D.L.; Sadik, A.; Platten, M.; Trump, S. The therapeutic potential of targeting tryptophan catabolism in cancer. Br. J. Cancer 2020, 122, 30–44. [Google Scholar] [CrossRef]
- Chen, J.Y.; Li, C.F.; Kuo, C.C.; Tsai, K.K.; Hou, M.F.; Hung, W.C. Cancer/stroma interplay via cyclooxygenase-2 and indoleamine 2,3-dioxygenase promotes breast cancer progression. Breast Cancer Res. 2014, 16, 410. [Google Scholar] [CrossRef]
- Ebata, T.; Shimizu, T.; Fujiwara, Y.; Tamura, K.; Kondo, S.; Iwasa, S.; Yonemori, K.; Shimomura, A.; Kitano, S.; Koyama, T.; et al. Phase I study of the indoleamine 2,3-dioxygenase 1 inhibitor navoximod (GDC-0919) as monotherapy and in combination with the PD-L1 inhibitor atezolizumab in Japanese patients with advanced solid tumours. Investig. New Drugs 2020, 38, 468–477. [Google Scholar] [CrossRef]
- Zhai, L.; Ladomersky, E.; Lenzen, A.; Nguyen, B.; Patel, R.; Lauing, K.L.; Wu, M.; Wainwright, D.A. IDO1 in cancer: A Gemini of immune checkpoints. Cell Mol. Immunol. 2018, 15, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Moretti, S.; Nucci, N.; Menicali, E.; Morelli, S.; Bini, V.; Colella, R.; Mandarano, M.; Sidoni, A.; Puxeddu, E. The aryl hydrocarbon receptor is expressed in thyroid carcinoma and appears to mediate epithelial-mesenchymal-transition. Cancers 2020, 12, 145. [Google Scholar] [CrossRef] [PubMed]
- Zahm, C.D.; Johnson, L.E.; McNeel, D.G. Increased indoleamine 2,3-dioxygenase activity and expression in prostate cancer following targeted immunotherapy. Cancer Immunol. Immunother. 2019, 68, 1661–1669. [Google Scholar] [CrossRef] [PubMed]
- Ala, M. The footprint of kynurenine pathway in every cancer: A new target for chemotherapy. Eur. J. Pharmacol. 2021, 896, 173921. [Google Scholar] [CrossRef]
- Awasthi, S.; Gerke, T.; Park, J.Y.; Asamoah, F.A.; Williams, V.L.; Fink, A.K.; Balkrishnan, R.; Lee, D.I.; Malkowicz, S.B.; Lal, P.; et al. Optimizing time-to-treatment to achieve durable biochemical disease control after surgery in prostate cancer—A multi-institutional cohort study. Cancer Epidemiol. Biomark. Prev. 2019, 28, 570–577. [Google Scholar] [CrossRef]
- Martins, A.H.; Hu, J.; Xu, Z.; Mu, C.; Alvarez, P.; Ford, B.D.; El Sayed, K.A.; Eterovic, V.A.; Ferchmin, P.A.; Hao, J. Neuroprotective activity of (1S,2E,4R,6R,7E,11E)-2,7,11-cembratriene-4,6-diol (4R) in vitro and in vivo in rodent models of brain ischemia. Neuroscience 2015, 291, 250–259. [Google Scholar] [CrossRef]
- Siddique, A.B.; Kilgore, P.C.; Tajmim, A.; Singh, S.S.; Meyer, S.A.; Jois, S.D.; Cvek, U.; Trutschl, M.; El Sayed, K.A. (−)-Oleocanthal as a dual c-MET-COX2 inhibitor for the control of lung cancer. Nutrients 2020, 12, 1749. [Google Scholar] [CrossRef]
- Abdelwahed, K.S.; Siddique, A.B.; Qusa, M.H.; King, J.A.; Souid, S.; Abd Elmageed, Z.Y.; El Sayed, K.A. PCSK9 Axis-targeting pseurotin A as a novel prostate cancer recurrence suppressor lead. ACS Pharmacol. Transl. Sci. 2021, 4, 1771–1781. [Google Scholar] [CrossRef]
- Abdelwahed, K.S.; Siddique, A.B.; Mohyeldin, M.M.; Qusa, M.H.; Goda, A.A.; Singh, S.S.; Ayoub, N.M.; King, J.A.; Jois, S.D.; El Sayed, K.A. Pseurotin A as a novel suppressor of hormone dependent breast cancer progression and recurrence by inhibiting PCSK9 secretion and interaction with LDL receptor. Pharmacol. Res. 2020, 158, 104847. [Google Scholar] [CrossRef]
- Siddique, A.B.; Ayoub, N.M.; Tajmim, A.; Meyer, S.A.; Hill, R.A.; El Sayed, K.A. (−)-Oleocanthal prevents breast cancer locoregional recurrence after primary tumor surgical excision and neoadjuvant targeted therapy in orthotopic nude mouse models. Cancers 2019, 11, 637. [Google Scholar] [CrossRef]
- Qusa, M.H.; Siddique, A.B.; Nazzal, S.; El Sayed, K.A. Novel olive oil phenolic (−)-oleocanthal (+)-xylitol-based solid dispersion formulations with potent oral anti-breast cancer activities. Int. J. Pharm. 2019, 569, 118596. [Google Scholar] [CrossRef] [PubMed]
- Brzozowska, B.; Gałecki, M.; Tartas, A.; Ginter, J.; Kaźmierczak, U.; Lundholm, L. Freeware tool for analysing numbers and sizes of cell colonies. Rad. Environ. Bioph. 2019, 58, 109–117. [Google Scholar] [CrossRef] [PubMed]
- Zelefsky, M.J.; Eastham, J.A.; Cronin, A.M.; Fuks, Z.; Zhang, Z.; Yamada, Y.; Vickers, A.; Scardino, P.T. Metastasis after radical prostatectomy or external beam radiotherapy for patients with clinically localized prostate cancer: A comparison of clinical cohorts adjusted for case mix. J. Clin. Oncol. 2010, 28, 1508–1513. [Google Scholar] [CrossRef] [PubMed]
- Damen, M.P.F.; van Rheenen, J.; Scheele, C.L.G.J. Targeting dormant tumor cells to prevent cancer recurrence. FEBS J. 2021, 288, 6286–6303. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Speck-Planche, A. Multiple perspectives in anti-cancer drug discovery: From old targets and natural products to innovative computational approaches. Anti-Cancer Agents Med. Chem. 2019, 19, 146–147. [Google Scholar] [CrossRef]
- Cabral, C.; Efferth, T.; Pires, I.M.; Severino, P.; Lemos, M.F.L. Natural products as a source for new leads in cancer research and treatment. Evid.-Based Complement. Altern. Med. 2018, 2018, 8243680. [Google Scholar] [CrossRef]
- Newman, D.J. Natural product based antibody drug conjugates: Clinical status as of 9 November 2020. J. Nat. Prod. 2021, 84, 917–931. [Google Scholar] [CrossRef]
- Yan, N.; Du, Y.; Liu, X.; Zhang, H.; Liu, Y.; Zhang, Z. A review on bioactivities of tobacco cembranoid diterpenes. Biomolecules 2019, 9, 30. [Google Scholar] [CrossRef]
- Akl, M.R.; Ayoub, N.M.; Mohyeldin, M.M.; Foudah, A.I.; Ayoub, N.M.; Liu, Y.Y.; El Sayed, K.A. Olive phenolics as c-Met inhibitors: (−)-Oleocanthal attenuates cell proliferation, invasiveness, and tumor growth in breast cancer models. PLoS ONE 2014, 9, e97622. [Google Scholar] [CrossRef]
- Retsky, M.; Demicheli, R.; Hrushesky, W.; Baum, M.; Gukas, I. Surgery triggers outgrowth of latent distant disease in breast Cancer: An inconvenient Truth? Cancers 2010, 2, 305–337. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Sengupta, P. Men and mice: Relating their ages. Life Sci. 2016, 152, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Duffy, M.J. Biomarkers for prostate cancer: Prostate-specific antigen and beyond. Clin. Chem. Lab. Med. 2020, 58, 326–339. [Google Scholar] [CrossRef] [PubMed]
- Artibani, W.; Porcaro, A.B.; De Marco, V.; Cerruto, M.A.; Siracusano, S. Management of biochemical recurrence after primary curative treatment for prostate cancer: A review. Urol. Int. 2018, 100, 251–262. [Google Scholar] [CrossRef]
- Moradi, A.; Srinivasan, S.; Clements, J.; Batra, J. Beyond the biomarker role: Prostate-specific antigen (PSA) in the prostate cancer microenvironment. Cancer Metastasis Rev. 2019, 38, 333–346. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prostate Cancer Cell Lines | |||||
|---|---|---|---|---|---|
| PC-3M | PC-3 | CWR-R1ca | DU-145 | CWR-22rv1 | |
| Wound healing assay | 4.50 ± 0.7 | 7.30 ± 0.9 | 9.80 ± 1.1 | 5.30 ± 0.8 | 9.40 ± 0.8 |
| Colony formation assay | 0.49 ± 0.1 | 2.93 ± 0.5 | 0.34 ± 0.1 | 1.21 ± 0.2 | 4.31 ± 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mudhish, E.A.; Siddique, A.B.; Ebrahim, H.Y.; Abdelwahed, K.S.; King, J.A.; El Sayed, K.A. The Tobacco β-Cembrenediol: A Prostate Cancer Recurrence Suppressor Lead and Prospective Scaffold via Modulation of Indoleamine 2,3-Dioxygenase and Tryptophan Dioxygenase. Nutrients 2022, 14, 1505. https://doi.org/10.3390/nu14071505
Mudhish EA, Siddique AB, Ebrahim HY, Abdelwahed KS, King JA, El Sayed KA. The Tobacco β-Cembrenediol: A Prostate Cancer Recurrence Suppressor Lead and Prospective Scaffold via Modulation of Indoleamine 2,3-Dioxygenase and Tryptophan Dioxygenase. Nutrients. 2022; 14(7):1505. https://doi.org/10.3390/nu14071505
Chicago/Turabian StyleMudhish, Ethar A., Abu Bakar Siddique, Hassan Y. Ebrahim, Khaldoun S. Abdelwahed, Judy Ann King, and Khalid A. El Sayed. 2022. "The Tobacco β-Cembrenediol: A Prostate Cancer Recurrence Suppressor Lead and Prospective Scaffold via Modulation of Indoleamine 2,3-Dioxygenase and Tryptophan Dioxygenase" Nutrients 14, no. 7: 1505. https://doi.org/10.3390/nu14071505
APA StyleMudhish, E. A., Siddique, A. B., Ebrahim, H. Y., Abdelwahed, K. S., King, J. A., & El Sayed, K. A. (2022). The Tobacco β-Cembrenediol: A Prostate Cancer Recurrence Suppressor Lead and Prospective Scaffold via Modulation of Indoleamine 2,3-Dioxygenase and Tryptophan Dioxygenase. Nutrients, 14(7), 1505. https://doi.org/10.3390/nu14071505