
Food Contamination: An Unexplored Possible Link between Dietary Habits and Parkinson’s Disease


Abstract
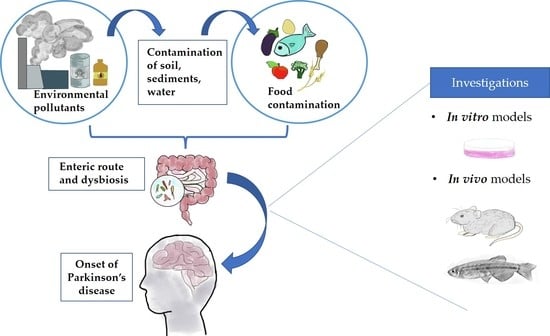
1. Introduction
2. The Impact of Food Contamination on the Onset of Parkinson’s Disease
2.1. Metals
2.1.1. Mercury
2.1.2. Manganese
2.2. Pesticides
2.2.1. Herbicides
2.2.2. Insecticides
2.3. Flame Retardants
Polybrominated Diphenyl Ethers
3. Evidence and Mechanisms of Action
3.1. Metals
3.2. Pesticides
4. The Hypothesis of the Enteric Route: From the Ingestion to the Brain
Gut Microbiome in Parkinson’s Disease: Is It Susceptible to Pollutants?
5. Models to Study the Role of Nutrition in Parkinson’s Disease
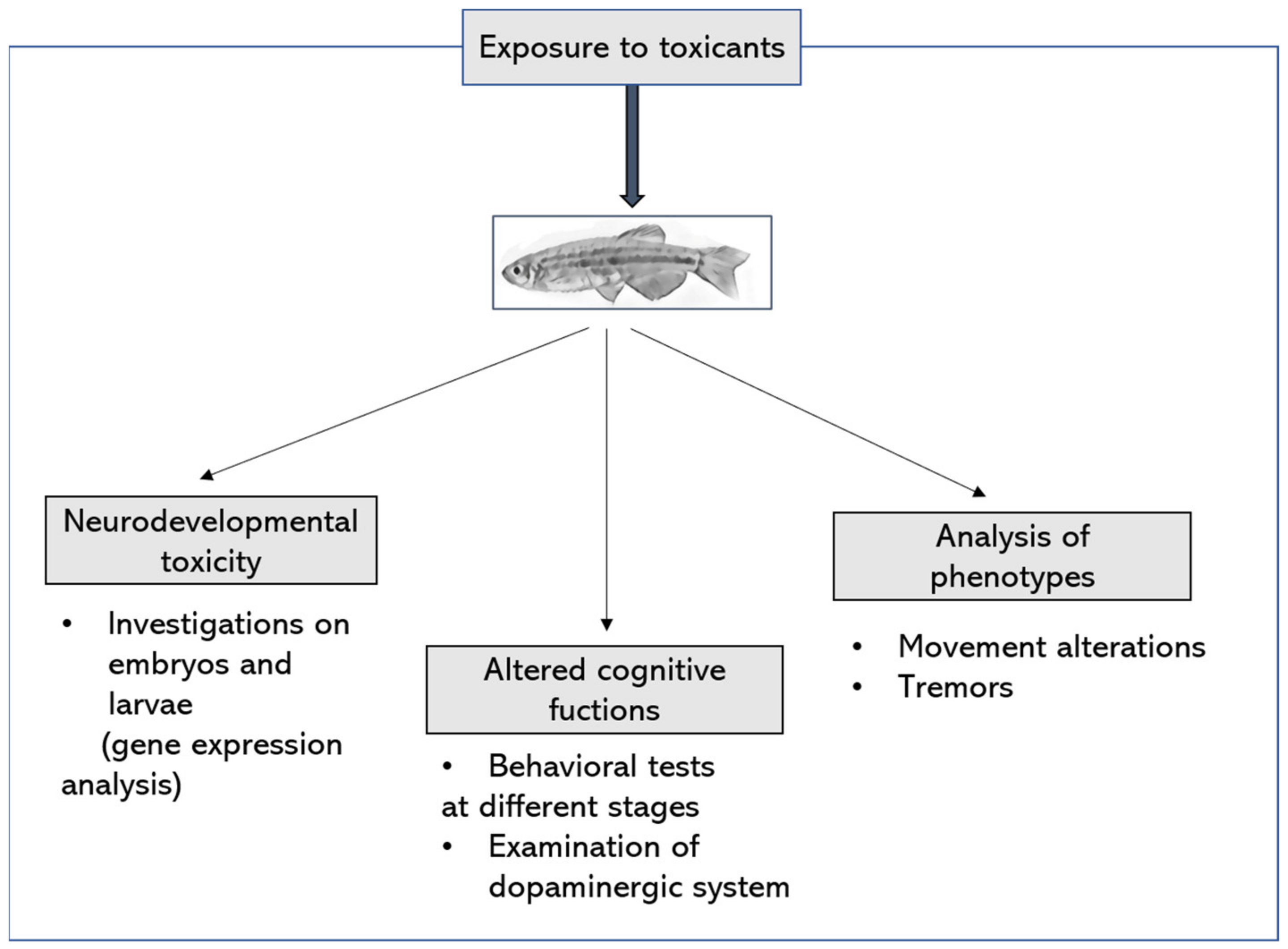
Zebrafish between Food Safety Research and Parkinson’s Disease Modelling
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Francis, H.M.; Stevenson, R.J. Potential for Diet to Prevent and Remediate Cognitive Deficits in Neurological Disorders. Nutr. Rev. 2018, 76, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Scarmeas, N.; Anastasiou, C.A.; Yannakoulia, M. Nutrition and Prevention of Cognitive Impairment. Lancet Neurol. 2018, 17, 1006–1015. [Google Scholar] [CrossRef]
- Béjot, Y.; Yaffe, K. Ageing Population: A Neurological Challenge. NED 2019, 52, 76–77. [Google Scholar] [CrossRef] [PubMed]
- Rather, I.A.; Koh, W.Y.; Paek, W.K.; Lim, J. The Sources of Chemical Contaminants in Food and Their Health Implications. Front. Pharmacol. 2017, 8, 830. [Google Scholar] [CrossRef]
- Riess, O.; Krüger, R. Parkinson’s Disease—A Multifactorial Neurodegenerative Disorder. J. Neural Transm. Suppl. 1999, 56, 113–125. [Google Scholar] [CrossRef]
- Antony, P.M.A.; Diederich, N.J.; Krüger, R.; Balling, R. The Hallmarks of Parkinson’s Disease. FEBS J. 2013, 280, 5981–5993. [Google Scholar] [CrossRef]
- Forno, L.S. Neuropathology of Parkinson’s Disease. J. Neuropathol. Exp. Neurol. 1996, 55, 259–272. [Google Scholar] [CrossRef]
- Jope, R.S.; Yuskaitis, C.J.; Beurel, E. Glycogen Synthase Kinase-3 (GSK3): Inflammation, Diseases, and Therapeutics. Neurochem. Res. 2007, 32, 577–595. [Google Scholar] [CrossRef]
- Li, D.-W.; Liu, Z.-Q.; Chen, W.; Yao, M.; Li, G.-R. Association of Glycogen Synthase Kinase-3β with Parkinson’s Disease (Review). Mol. Med. Rep. 2014, 9, 2043–2050. [Google Scholar] [CrossRef]
- Wang, W.; Yang, Y.; Ying, C.; Li, W.; Ruan, H.; Zhu, X.; You, Y.; Han, Y.; Chen, R.; Wang, Y.; et al. Inhibition of Glycogen Synthase Kinase-3β Protects Dopaminergic Neurons from MPTP Toxicity. Neuropharmacology 2007, 52, 1678–1684. [Google Scholar] [CrossRef]
- King, T.D.; Clodfelder-Miller, B.; Barksdale, K.A.; Bijur, G.N. Unregulated Mitochondrial GSK3β Activity Results in NADH:Ubiquinone Oxidoreductase Deficiency. Neurotox. Res. 2008, 14, 367–382. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Ritz, B. The Search for Environmental Causes of Parkinson’s Disease: Moving Forward. J. Parkinson’s Dis. 2018, 8, S9–S17. [Google Scholar] [CrossRef] [PubMed]
- Ball, N.; Teo, W.-P.; Chandra, S.; Chapman, J. Parkinson’s Disease and the Environment. Front. Neurol. 2019, 10, 218. [Google Scholar] [CrossRef] [PubMed]
- Toro, R.; Downward, G.S.; van der Mark, M.; Brouwer, M.; Huss, A.; Peters, S.; Hoek, G.; Nijssen, P.; Mulleners, W.M.; Sas, A.; et al. Parkinson’s Disease and Long-Term Exposure to Outdoor Air Pollution: A Matched Case-Control Study in the Netherlands. Environ. Int. 2019, 129, 28–34. [Google Scholar] [CrossRef]
- Tanner, C.M. Advances in Environmental Epidemiology. Mov. Disord. 2010, 25, S58–S62. [Google Scholar] [CrossRef] [PubMed]
- Bro-Rasmussen, F. Contamination by Persistent Chemicals in Food Chain and Human Health. Sci. Total Environ. 1996, 188, S45–S60. [Google Scholar] [CrossRef]
- Chia, S.J.; Tan, E.-K.; Chao, Y.-X. Historical Perspective: Models of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 72464. [Google Scholar] [CrossRef]
- Chojnacka, K.; Mikulewicz, M. Bioaccumulation. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 456–460. ISBN 978-012-386-455-0. [Google Scholar]
- Ritz, B.; Lee, P.-C.; Hansen, J.; Lassen, C.F.; Ketzel, M.; Sørensen, M.; Raaschou-Nielsen, O. Traffic-Related Air Pollution and Parkinson’s Disease in Denmark: A Case–Control Study. Environ. Health Perspect. 2016, 124, 351–356. [Google Scholar] [CrossRef]
- Tanner, C.M.; Chen, B.; Wang, W.; Peng, M.; Liu, Z.; Liang, X.; Kao, L.C.; Gilley, D.W.; Goetz, C.G.; Schoenberg, B.S. Environmental Factors and Parkinson’s Disease: A Case-Control Study in China. Neurology 1989, 39, 660–664. [Google Scholar] [CrossRef]
- Emurotu, J.E.; Onianwa, P.C. Bioaccumulation of Heavy Metals in Soil and Selected Food Crops Cultivated in Kogi State, North Central Nigeria. Environ. Syst. Res. 2017, 6, 21. [Google Scholar] [CrossRef]
- Yousafzai, A.M.; Ullah, F.; Bari, F.; Raziq, S.; Riaz, M.; Khan, K.; Nishan, U.; Sthanadar, I.A.; Shaheen, B.; Shaheen, M.; et al. Bioaccumulation of Some Heavy Metals: Analysis and Comparison of Cyprinus carpio and Labeo rohita from Sardaryab, Khyber Pakhtunkhwa. Biomed. Res. Int. 2017, 2017, 5801432. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Khisroon, M.; Khan, A.; Gulfam, N.; Siraj, M.; Zaidi, F.; Ahmadullah; Abidullah; Fatima, S.H.; Noreen, S.; et al. Bioaccumulation of Heavy Metals in Water, Sediments, and Tissues and Their Histopathological Effects on Anodonta cygnea (Linea, 1876) in Kabul River, Khyber Pakhtunkhwa, Pakistan. BioMed Res. Int. 2018, 2018, e1910274. [Google Scholar] [CrossRef] [PubMed]
- Jakimska, A.; Konieczka, P.; Skóra, K.; Namieśnik, J. Bioaccumulation of Metals in Tissues of Marine Animals, Part II: Metal Concentrations in Animal Tissues. Pol. J. Environ. Stud. 2011, 20, 1127–1146. [Google Scholar]
- Elsayed, H.; Yigiterhan, O.; Al-Ansari, E.M.A.S.; Al-Ashwel, A.A.; Elezz, A.A.; Al-Maslamani, I.A. Methylmercury Bioaccumulation among Different Food Chain Levels in the EEZ of Qatar (Arabian Gulf). Reg. Stud. Mar. Sci. 2020, 37, 101334. [Google Scholar] [CrossRef]
- Dos Anjos, M.R.; Machado, N.G.; da Silva, M.E.P.; Bastos, W.R.; Miranda, M.R.; de Carvalho, D.P.; Mussy, M.H.; de Holanda, I.B.B.; Biudes, M.S.; Fulan, J.Â. Bioaccumulation of Methylmercury in Fish Tissue from the Roosevelt River, Southwestern Amazon Basin. Rev. Ambient. Água 2016, 11, 508–518. [Google Scholar] [CrossRef]
- Liu, M.; Xiao, W.; Zhang, Q.; Shi, L.; Wang, X.; Xu, Y. Methylmercury Bioaccumulation in Deepest Ocean Fauna: Implications for Ocean Mercury Biotransport through Food Webs. Environ. Sci. Technol. Lett. 2020, 7, 469–476. [Google Scholar] [CrossRef]
- Harada, M. Minamata Disease: Methylmercury Poisoning in Japan Caused by Environmental Pollution. Crit. Rev. Toxicol. 1995, 25, 1–24. [Google Scholar] [CrossRef]
- Shao, Y.; Figeys, D.; Ning, Z.; Mailloux, R.; Chan, H.M. Methylmercury Can Induce Parkinson’s-like Neurotoxicity Similar to 1-Methyl-4- Phenylpyridinium: A Genomic and Proteomic Analysis on MN9D Dopaminergic Neuron Cells. J. Toxicol. Sci. 2015, 40, 817–828. [Google Scholar] [CrossRef][Green Version]
- Nabi, S. Methylmercury and Parkinson’s Disease. In Toxic Effects of Mercury; Nabi, S., Ed.; Springer India: New Delhi, India, 2014; pp. 211–218. ISBN 978-813-221-922-4. [Google Scholar]
- Ho, P.W.; Ho, J.W.; Liu, H.-F.; So, D.H.; Tse, Z.H.; Chan, K.-H.; Ramsden, D.B.; Ho, S.-L. Mitochondrial Neuronal Uncoupling Proteins: A Target for Potential Disease-Modification in Parkinson’s Disease. Transl. Neurodegener. 2012, 1, 3. [Google Scholar] [CrossRef]
- Petersen, M.S.; Halling, J.; Bech, S.; Wermuth, L.; Weihe, P.; Nielsen, F.; Jørgensen, P.J.; Budtz-Jørgensen, E.; Grandjean, P. Impact of Dietary Exposure to Food Contaminants on the Risk of Parkinson’s Disease. NeuroToxicology 2008, 29, 584–590. [Google Scholar] [CrossRef]
- Costa, F.d.N.; Korn, M.G.A.; Brito, G.B.; Ferlin, S.; Fostier, A.H. Preliminary Results of Mercury Levels in Raw and Cooked Seafood and Their Public Health Impact. Food Chem. 2016, 192, 837–841. [Google Scholar] [CrossRef] [PubMed]
- Takeda, A. Manganese Action in Brain Function. Brain Res. Rev. 2003, 41, 79–87. [Google Scholar] [CrossRef]
- Couper, J. On the effects of black oxide of manganese when inhaled into the lungs. Br. Ann. Med. Pharmacol. 1837, 1, 41–42. [Google Scholar]
- Guilarte, T.R. Manganese and Parkinson’s Disease: A Critical Review and New Findings. Environ. Health Perspect. 2010, 118, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Song, H.-J.; Lee, J.-J.; Seo, J.-H.; Kim, J.-H.; Lee, H.J.; Kim, H.J.; Kim, Y.; Ahn, J.-H.; Park, S.-J.; et al. Neuroplastic Changes within the Brains of Manganese-Exposed Welders: Recruiting Additional Neural Resources for Successful Motor Performance. Occup. Environ. Med. 2010, 67, 809–815. [Google Scholar] [CrossRef] [PubMed]
- Smargiassi, A.; Baldwin, M.; Savard, S.; Kennedy, G.; Mergler, D.; Zayed, J. Assessment of Exposure to Manganese in Welding Operations during the Assembly of Heavy Excavation Machinery Accessories. Appl. Occup. Environ. Hyg. 2000, 15, 746–750. [Google Scholar] [CrossRef] [PubMed]
- Santamaria, A.B.; Cushing, C.A.; Antonini, J.M.; Finley, B.L.; Mowat, F.S. State-of-the-Science Review: Does Manganese Exposure during Welding Pose a Neurological Risk? J. Toxicol. Environ. Health B Crit. Rev. 2007, 10, 417–465. [Google Scholar] [CrossRef]
- Sjögren, B.; Gustavsson, P.; Hogstedt, C. Neuropsychiatric Symptoms among Welders Exposed to Neurotoxic Metals. Occup. Environ. Med. 1990, 47, 704–707. [Google Scholar] [CrossRef]
- Powers, K.M.; Smith-Weller, T.; Franklin, G.M.; Longstreth, W.T.; Swanson, P.D.; Checkoway, H. Parkinson’s Disease Risks Associated with Dietary Iron, Manganese, and Other Nutrient Intakes. Neurology 2003, 60, 1761–1766. [Google Scholar] [CrossRef]
- Chua, A.C.; Morgan, E.H. Effects of Iron Deficiency and Iron Overload on Manganese Uptake and Deposition in the Brain and Other Organs of the Rat. Biol. Trace Elem. Res. 1996, 55, 39–54. [Google Scholar] [CrossRef]
- Eriguchi, M.; Iida, K.; Ikeda, S.; Osoegawa, M.; Nishioka, K.; Hattori, N.; Nagayama, H.; Hara, H. Parkinsonism Relating to Intoxication with Glyphosate. Intern. Med. 2019, 58, 1935–1938. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, E.R.; Leiros da Costa, M.D.; Bacheschi, L.A.; Scaff, M.; Leite, C.C. Parkinsonism after Glycine-Derivate Exposure. Mov. Disord. 2001, 16, 565–568. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Fan, X.-N.; Tan, Y.-Y.; Cheng, Q.; Chen, S.-D. Parkinsonism after Chronic Occupational Exposure to Glyphosate. Parkinsonism Relat. Disord. 2011, 17, 486–487. [Google Scholar] [CrossRef] [PubMed]
- Cattani, D.; de Liz Oliveira Cavalli, V.L.; Heinz Rieg, C.E.; Domingues, J.T.; Dal-Cim, T.; Tasca, C.I.; Mena Barreto Silva, F.R.; Zamoner, A. Mechanisms Underlying the Neurotoxicity Induced by Glyphosate-Based Herbicide in Immature Rat Hippocampus: Involvement of Glutamate Excitotoxicity. Toxicology 2014, 320, 34–45. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, I.; Sung, J.-M.; Song, J. Parkinson’s Disease in a Worker Exposed to Insecticides at a Greenhouse. Ann. Occup. Environ. Med. 2021, 33, e6. [Google Scholar] [CrossRef]
- Wang, A.; Cockburn, M.; Ly, T.T.; Bronstein, J.; Ritz, B. The Association Between Ambient Exposure to Organophosphates and Parkinson’s Disease Risk. Occup. Environ. Med. 2014, 71, 275–281. [Google Scholar] [CrossRef]
- Lukaszewicz-Hussain, A. Role of Oxidative Stress in Organophosphate Insecticide Toxicity—Short Review. Pestic. Biochem. Physiol. 2010, 98, 145–150. [Google Scholar] [CrossRef]
- Norkaew, S.; Lertmaharit, S.; Wilaiwan, W.; Siriwong, W.; Pérez, H.M.; Robson, M.G. An Association between Organophosphate Pesticides Exposure and Parkinsonism amongst People in an Agricultural Area in Ubon Ratchathani Province, Thailand. Rocz. Panstw. Zakl. Hig. 2015, 66, 21–26. [Google Scholar]
- Slotkin, T.A.; Seidler, F.J. Developmental Exposure to Organophosphates Triggers Transcriptional Changes in Genes Associated with Parkinson’s Disease In Vitro and In Vivo. Brain Res. Bull. 2011, 86, 340–347. [Google Scholar] [CrossRef]
- Lee, P.-C.; Rhodes, S.L.; Sinsheimer, J.S.; Bronstein, J.; Ritz, B. Functional Paraoxonase 1 Variants Modify the Risk of Parkinson’s Disease Due to Organophosphate Exposure. Environ. Int. 2013, 56, 42–47. [Google Scholar] [CrossRef]
- Feng, M.; Li, Y.; Qu, R.; Wang, L.; Wang, Z. Oxidative Stress Biomarkers in Freshwater Fish Carassius auratus Exposed to Decabromodiphenyl Ether and Ethane, or Their Mixture. Ecotoxicology 2013, 22, 1101–1110. [Google Scholar] [CrossRef] [PubMed]
- Caudle, W.M.; Guillot, T.S.; Lazo, C.R.; Miller, G.W. Industrial Toxicants and Parkinson’s Disease. Neurotoxicology 2012, 33, 178–188. [Google Scholar] [CrossRef]
- Costa, L.G.; Giordano, G. Developmental Neurotoxicity of Polybrominated Diphenyl Ether (PBDE) Flame Retardants. Neurotoxicology 2007, 28, 1047–1067. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kannan, K.; Moon, H.-B. Assessment of Exposure to Polybrominated Diphenyl Ethers (PBDEs) via Seafood Consumption and Dust Ingestion in Korea. Sci. Total Environ. 2013, 443, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Malisch, R.; Kotz, A. Dioxins and PCBs in Feed and Food—Review from European Perspective. Sci. Total Environ. 2014, 491–492, 2–10. [Google Scholar] [CrossRef] [PubMed]
- Giannico, O.V.; Desiante, F.; Basile, F.C.; Franco, E.; Baldacci, S.; Fragnelli, G.R.; Diletti, G.; Conversano, M. Dioxins and PCBs Contamination in Mussels from Taranto (Ionian Sea, Southern Italy): A Seven Years Spatio-Temporal Monitoring Study. Ann. Ist. Super. Sanità 2020, 56, 452–461. [Google Scholar] [CrossRef]
- Weber, R.; Herold, C.; Hollert, H.; Kamphues, J.; Ungemach, L.; Blepp, M.; Ballschmiter, K. Life Cycle of PCBs and Contamination of the Environment and of Food Products from Animal Origin. Environ. Sci. Pollut. Res. Int. 2018, 25, 16325–16343. [Google Scholar] [CrossRef]
- Dron, J.; Wafo, E.; Boissery, P.; Dhermain, F.; Bouchoucha, M.; Chamaret, P.; Lafitte, D. Trends of Banned Pesticides and PCBs in Different Tissues of Striped Dolphins (Stenella coeruleoalba) Stranded in the Northwestern Mediterranean Reflect Changing Contamination Patterns. Mar. Pollut. Bull. 2021, 174, 113198. [Google Scholar] [CrossRef]
- Rybicki, B.A.; Johnson, C.C.; Uman, J.; Gorell, J.M. Parkinson’s Disease Mortality and the Industrial Use of Heavy Metals in Michigan. Mov. Disord. 1993, 8, 87–92. [Google Scholar] [CrossRef]
- Gorell, J.M.; Johnson, C.C.; Rybicki, B.A.; Peterson, E.L.; Kortsha, G.X.; Brown, G.G.; Richardson, R.J. Occupational Exposures to Metals as Risk Factors for Parkinson’s Disease. Neurology 1997, 48, 650–658. [Google Scholar] [CrossRef]
- Zecca, L.; Youdim, M.B.H.; Riederer, P.; Connor, J.R.; Crichton, R.R. Iron, Brain Ageing and Neurodegenerative Disorders. Nat. Rev. Neurosci. 2004, 5, 863–873. [Google Scholar] [CrossRef]
- Jellinger, K.A. The Role of Iron in Neurodegeneration: Prospects for Pharmacotherapy of Parkinson’s Disease. Drugs Aging 1999, 14, 115–140. [Google Scholar] [CrossRef] [PubMed]
- Fernandes Azevedo, B.; Barros Furieri, L.; Peçanha, F.M.; Wiggers, G.A.; Frizera Vassallo, P.; Ronacher Simões, M.; Fiorim, J.; Rossi de Batista, P.; Fioresi, M.; Rossoni, L.; et al. Toxic Effects of Mercury on the Cardiovascular and Central Nervous Systems. J. Biomed. Biotechnol. 2012, 2012, e949048. [Google Scholar] [CrossRef] [PubMed]
- Bjorklund, G.; Stejskal, V.; Urbina, M.A.; Dadar, M.; Chirumbolo, S.; Mutter, J. Metals and Parkinson’s Disease: Mechanisms and Biochemical Processes. Curr. Med. Chem. 2018, 25, 2198–2214. [Google Scholar] [CrossRef]
- Gorell, J.M.; Rybicki, B.A.; Cole Johnson, C.; Peterson, E.L. Occupational Metal Exposures and the Risk of Parkinson’s Disease. Neuroepidemiology 1999, 18, 303–308. [Google Scholar] [CrossRef]
- Shao, Y.; Chan, H.M. Effects of Methylmercury on Dopamine Release in MN9D Neuronal Cells. Toxicol. Mech. Methods 2015, 25, 637–644. [Google Scholar] [CrossRef]
- Prashanth, L.K.; Taly, A.B.; Sinha, S.; Arunodaya, G.R.; Swamy, H.S. Wilson’s Disease: Diagnostic Errors and Clinical Implications. J. Neurol. Neurosurg. Psychiatry 2004, 75, 907–909. [Google Scholar] [CrossRef]
- Johnson, S. Is Parkinson’s Disease the Heterozygote Form of Wilson’s Disease: PD = 1/2 WD? Med. Hypotheses 2001, 56, 171–173. [Google Scholar] [CrossRef]
- Dusek, P.; Litwin, T.; Członkowska, A. Neurologic Impairment in Wilson Disease. Ann. Transl. Med. 2019, 7 (Suppl. 2), S64. [Google Scholar] [CrossRef]
- Ortiz, J.F.; Cox, Á.M.; Tambo, W.; Eskander, N.; Wirth, M.; Valdez, M.; Niño, M. Neurological Manifestations of Wilson’s Disease: Pathophysiology and Localization of Each Component. Cureus 2020, 12, e11509. [Google Scholar] [CrossRef]
- Royer, A.; Sharman, T. Copper Toxicity. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Kalita, J.; Kumar, V.; Misra, U.K.; Bora, H.K. Movement Disorder in Copper Toxicity Rat Model: Role of Inflammation and Apoptosis in the Corpus Striatum. Neurotox. Res. 2020, 37, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Castillo-Gonzalez, J.A.; Loera-Arias, M.D.J.; Saucedo-Cardenas, O.; Montes-de-Oca-Luna, R.; Garcia-Garcia, A.; Rodriguez-Rocha, H. Phosphorylated α-Synuclein-Copper Complex Formation in the Pathogenesis of Parkinson’s Disease. Parkinson’s Dis. 2017, 2017, 9164754. [Google Scholar] [CrossRef]
- Binolfi, A.; Lamberto, G.R.; Duran, R.; Quintanar, L.; Bertoncini, C.W.; Souza, J.M.; Cerveñansky, C.; Zweckstetter, M.; Griesinger, C.; Fernández, C.O. Site-Specific Interactions of Cu(II) with Alpha and Beta-Synuclein: Bridging the Molecular Gap between Metal Binding and Aggregation. J. Am. Chem. Soc. 2008, 130, 11801–11812. [Google Scholar] [CrossRef] [PubMed]
- Paris, I.; Perez-Pastene, C.; Couve, E.; Caviedes, P.; LeDoux, S.; Segura-Aguilar, J. Copper·Dopamine Complex Induces Mitochondrial Autophagy Preceding Caspase-Independent Apoptotic Cell Death. J. Biol. Chem. 2009, 284, 13306–13315. [Google Scholar] [CrossRef]
- Caudle, W.M. Occupational Metal Exposure and Parkinsonism. In Neurotoxicity of Metals; Aschner, M., Costa, L.G., Eds.; Advances in Neurobiology; Springer International Publishing: Cham, Switzerland, 2017; Volume 18, pp. 143–158. ISBN 978-331-960-188-5. [Google Scholar]
- Gomes, C.M.; Santos, R. Neurodegeneration in Friedreich’s Ataxia: From Defective Frataxin to Oxidative Stress. Oxid. Med. Cell. Longev. 2013, 2013, 487534. [Google Scholar] [CrossRef]
- Parkinson, M.H.; Boesch, S.; Nachbauer, W.; Mariotti, C.; Giunti, P. Clinical Features of Friedreich’s Ataxia: Classical and Atypical Phenotypes. J. Neurochem. 2013, 126, 103–117. [Google Scholar] [CrossRef]
- Uversky, V.N.; Li, J.; Fink, A.L. Metal-Triggered Structural Transformations, Aggregation, and Fibrillation of Human Alpha-Synuclein. A Possible Molecular NK between Parkinson’s Disease and Heavy Metal Exposure. J. Biol. Chem. 2001, 276, 44284–44296. [Google Scholar] [CrossRef]
- Chen, P.; Parmalee, N.; Aschner, M. Genetic Factors and Manganese-Induced Neurotoxicity. Front. Genet. 2014, 5, 265. [Google Scholar] [CrossRef]
- Mason, L.H.; Harp, J.P.; Han, D.Y. Pb Neurotoxicity: Neuropsychological Effects of Lead Toxicity. BioMed Res. Int. 2014, 2014, e840547. [Google Scholar] [CrossRef]
- Coon, S.; Stark, A.; Peterson, E.; Gloi, A.; Kortsha, G.; Pounds, J.; Chettle, D.; Gorell, J. Whole-Body Lifetime Occupational Lead Exposure and Risk of Parkinson’s Disease. Environ. Health Perspect. 2006, 114, 1872–1876. [Google Scholar] [CrossRef]
- Richardson, J.R.; Fitsanakis, V.; Westerink, R.H.S.; Kanthasamy, A.G. Neurotoxicity of Pesticides. Acta Neuropathol. 2019, 138, 343–362. [Google Scholar] [CrossRef] [PubMed]
- Caldas, E.D. Toxicological Aspects of Pesticides. In Sustainable Agrochemistry: A Compendium of Technologies; Vaz, S., Jr., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 275–305. ISBN 978-303-017-891-8. [Google Scholar]
- Hassaan, M.A.; El Nemr, A. Pesticides Pollution: Classifications, Human Health Impact, Extraction and Treatment Techniques. Egypt. J. Aquat. Res. 2020, 46, 207–220. [Google Scholar] [CrossRef]
- Langston, J.W.; Ballard, P.; Tetrud, J.W.; Irwin, I. Chronic Parkinsonism in Humans Due to a Product of Meperidine-Analog Synthesis. Science 1983, 219, 979–980. [Google Scholar] [CrossRef] [PubMed]
- Ritz, B.R.; Paul, K.C.; Bronstein, J.M. Of Pesticides and Men: A California Story of Genes and Environment in Parkinson’s Disease. Curr. Environ. Health Rep. 2016, 3, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Kamel, F.; Hoppin, J.A. Association of Pesticide Exposure with Neurologic Dysfunction and Disease. Environ. Health Perspect. 2004, 112, 950–958. [Google Scholar] [CrossRef]
- Cole, D.C.; Carpio, F.; Julian, J.; Léon, N. Assessment of Peripheral Nerve Function in an Ecuadorian Rural Population Exposed to Pesticides. J. Toxicol. Environ. Health A 1998, 55, 77–91. [Google Scholar] [CrossRef]
- Ruijten, M.W.; Sallé, H.J.; Verberk, M.M.; Smink, M. Effect of Chronic Mixed Pesticide Exposure on Peripheral and Autonomic Nerve Function. Arch. Environ. Health 1994, 49, 188–195. [Google Scholar] [CrossRef]
- Muñoz-Quezada, M.T.; Lucero, B.A.; Iglesias, V.P.; Muñoz, M.P.; Cornejo, C.A.; Achu, E.; Baumert, B.; Hanchey, A.; Concha, C.; Brito, A.M.; et al. Chronic Exposure to Organophosphate (OP) Pesticides and Neuropsychological Functioning in Farm Workers: A Review. Int. J. Occup. Environ. Health 2016, 22, 68–79. [Google Scholar] [CrossRef]
- Voorhees, J.R.; Rohlman, D.S.; Lein, P.J.; Pieper, A.A. Neurotoxicity in Preclinical Models of Occupational Exposure to Organophosphorus Compounds. Front. Neurosci. 2016, 10, 590. [Google Scholar] [CrossRef]
- Narayan, S.; Liew, Z.; Bronstein, J.M.; Ritz, B. Occupational Pesticide Use and Parkinson’s Disease in the Parkinson Environment Gene (PEG) Study. Environ. Int. 2017, 107, 266–273. [Google Scholar] [CrossRef]
- Vaccari, C.; El Dib, R.; de Camargo, J.L.V. Paraquat and Parkinson’s Disease: A Systematic Review Protocol According to the OHAT Approach for Hazard Identification. Syst. Rev. 2017, 6, 98. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Freyberger, A.; Lawrenz, B.; Schladt, L.; Schmuck, G.; Ellinger-Ziegelbauer, H. Mechanistic Investigations of the Mitochondrial Complex I Inhibitor Rotenone in the Context of Pharmacological and Safety Evaluation. Sci. Rep. 2017, 7, 45465. [Google Scholar] [CrossRef] [PubMed]
- Kara, M.; Oztas, E.; Ramazanoğulları, R.; Kouretas, D.; Nepka, C.; Tsatsakis, A.M.; Veskoukis, A.S. Benomyl, a Benzimidazole Fungicide, Induces Oxidative Stress and Apoptosis in Neural Cells. Toxicol. Rep. 2020, 7, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Braak, H.; Rüb, U.; Gai, W.P.; Del Tredici, K. Idiopathic Parkinson’s Disease: Possible Routes by Which Vulnerable Neuronal Types May Be Subject to Neuroinvasion by an Unknown Pathogen. J. Neural Transm. 2003, 110, 517–536. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, C.H.; Del Tredici, K.; Braak, H. Parkinson’s Disease: A Dual-hit Hypothesis. Neuropathol. Appl. Neurobiol. 2007, 33, 599–614. [Google Scholar] [CrossRef] [PubMed]
- Rietdijk, C.D.; Perez-Pardo, P.; Garssen, J.; van Wezel, R.J.A.; Kraneveld, A.D. Exploring Braak’s Hypothesis of Parkinson’s Disease. Front. Neurol. 2017, 8, 37. [Google Scholar] [CrossRef]
- Pan-Montojo, F.; Anichtchik, O.; Dening, Y.; Knels, L.; Pursche, S.; Jung, R.; Jackson, S.; Gille, G.; Spillantini, M.G.; Reichmann, H.; et al. Progression of Parkinson’s Disease Pathology Is Reproduced by Intragastric Administration of Rotenone in Mice. PLoS ONE 2010, 5, e8762. [Google Scholar] [CrossRef]
- Holmqvist, S.; Chutna, O.; Bousset, L.; Aldrin-Kirk, P.; Li, W.; Björklund, T.; Wang, Z.-Y.; Roybon, L.; Melki, R.; Li, J.-Y. Direct Evidence of Parkinson Pathology Spread from the Gastrointestinal Tract to the Brain in Rats. Acta Neuropathol. 2014, 128, 805–820. [Google Scholar] [CrossRef]
- Anselmi, L.; Bove, C.; Coleman, F.H.; Le, K.; Subramanian, M.P.; Venkiteswaran, K.; Subramanian, T.; Travagli, R.A. Ingestion of Subthreshold Doses of Environmental Toxins Induces Ascending Parkinsonism in the Rat. NPJ Parkinson’s Dis. 2018, 4, 30. [Google Scholar] [CrossRef]
- Devos, D.; Lebouvier, T.; Lardeux, B.; Biraud, M.; Rouaud, T.; Pouclet, H.; Coron, E.; Bruley des Varannes, S.; Naveilhan, P.; Nguyen, J.-M.; et al. Colonic Inflammation in Parkinson’s Disease. Neurobiol. Dis. 2013, 50, 42–48. [Google Scholar] [CrossRef]
- Forsyth, C.B.; Shannon, K.M.; Kordower, J.H.; Voigt, R.M.; Shaikh, M.; Jaglin, J.A.; Estes, J.D.; Dodiya, H.B.; Keshavarzian, A. Increased Intestinal Permeability Correlates with Sigmoid Mucosa Alpha-Synuclein Staining and Endotoxin Exposure Markers in Early Parkinson’s Disease. PLoS ONE 2011, 6, e28032. [Google Scholar] [CrossRef] [PubMed]
- Banks, W.A.; Dohgu, S.; Lynch, J.L.; Fleegal-DeMotta, M.A.; Erickson, M.A.; Nakaoke, R.; Vo, T.Q. Nitric Oxide Isoenzymes Regulate Lipopolysaccharide-Enhanced Insulin Transport across the Blood-Brain Barrier. Endocrinology 2008, 149, 1514–1523. [Google Scholar] [CrossRef]
- Banks, W.A.; Erickson, M.A. The Blood-Brain Barrier and Immune Function and Dysfunction. Neurobiol. Dis. 2010, 37, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, C.; Antonioli, L.; Colucci, R.; Blandizzi, C.; Fornai, M. Interplay among Gut Microbiota, Intestinal Mucosal Barrier and Enteric Neuro-Immune System: A Common Path to Neurodegenerative Diseases? Acta Neuropathol. 2018, 136, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, J.M.; Murphy, K.; Stanton, C.; Ross, R.P.; Kober, O.I.; Juge, N.; Avershina, E.; Rudi, K.; Narbad, A.; Jenmalm, M.C.; et al. The Composition of the Gut Microbiota throughout Life, with an Emphasis on Early Life. Microb. Ecol. Health Dis. 2015, 26, 26050. [Google Scholar] [CrossRef]
- Phelps, D.; Brinkman, N.E.; Keely, S.P.; Anneken, E.M.; Catron, T.R.; Betancourt, D.; Wood, C.E.; Espenschied, S.T.; Rawls, J.F.; Tal, T. Microbial Colonization Is Required for Normal Neurobehavioral Development in Zebrafish. Sci. Rep. 2017, 7, 11244. [Google Scholar] [CrossRef]
- Keshavarzian, A.; Green, S.J.; Engen, P.A.; Voigt, R.M.; Naqib, A.; Forsyth, C.B.; Mutlu, E.; Shannon, K.M. Colonic Bacterial Composition in Parkinson’s Disease. Mov. Disord. 2015, 30, 1351–1360. [Google Scholar] [CrossRef]
- Scheperjans, F.; Aho, V.; Pereira, P.A.B.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut Microbiota Are Related to Parkinson’s Disease and Clinical Phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.-M.; et al. Enterotypes of the Human Gut Microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- Defois, C.; Ratel, J.; Denis, S.; Batut, B.; Beugnot, R.; Peyretaillade, E.; Engel, E.; Peyret, P. Environmental Pollutant Benzo[a]Pyrene Impacts the Volatile Metabolome and Transcriptome of the Human Gut Microbiota. Front. Microbiol. 2017, 8, 1562. [Google Scholar] [CrossRef]
- Cruz, R.; Palmeira, J.D.; Martins, Z.E.; Faria, M.A.; Ferreira, H.; Marques, A.; Casal, S.; Cunha, S.C. Multidisciplinary Approach to Determine the Effect of Polybrominated Diphenyl Ethers on Gut Microbiota. Environ. Pollut. 2020, 260, 113920. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Ruzafa, L.; Cruz, F.; Roman, P.; Cardona, D. Gut Microbiota and Neurological Effects of Glyphosate. NeuroToxicology 2019, 75, 1–8. [Google Scholar] [CrossRef]
- Tang, Q.; Tang, J.; Ren, X.; Li, C. Glyphosate Exposure Induces Inflammatory Responses in the Small Intestine and Alters Gut Microbial Composition in Rats. Environ. Pollut. 2020, 261, 114129. [Google Scholar] [CrossRef] [PubMed]
- Aitbali, Y.; Ba-M’hamed, S.; Elhidar, N.; Nafis, A.; Soraa, N.; Bennis, M. Glyphosate Based- Herbicide Exposure Affects Gut Microbiota, Anxiety and Depression-like Behaviors in Mice. Neurotoxicol. Teratol. 2018, 67, 44–49. [Google Scholar] [CrossRef]
- Arun, K.B.; Madhavan, A.; Sindhu, R.; Emmanual, S.; Binod, P.; Pugazhendhi, A.; Sirohi, R.; Reshmy, R.; Awasthi, M.K.; Gnansounou, E.; et al. Probiotics and Gut Microbiome—Prospects and Challenges in Remediating Heavy Metal Toxicity. J. Hazard. Mater. 2021, 420, 126676. [Google Scholar] [CrossRef] [PubMed]
- Seki, N.; Akiyama, M.; Yamakawa, H.; Hase, K.; Kumagai, Y.; Kim, Y.-G. Adverse Effects of Methylmercury on Gut Bacteria and Accelerated Accumulation of Mercury in Organs Due to Disruption of Gut Microbiota. J. Toxicol. Sci. 2021, 46, 91–97. [Google Scholar] [CrossRef]
- Pinto, D.V.; Raposo, R.S.; Matos, G.A.; Alvarez-Leite, J.I.; Malva, J.O.; Oriá, R.B. Methylmercury Interactions With Gut Microbiota and Potential Modulation of Neurogenic Niches in the Brain. Front. Neurosci. 2020, 14, 576543. [Google Scholar] [CrossRef]
- Wang, H.-T.; Ding, J.; Xiong, C.; Zhu, D.; Li, G.; Jia, X.-Y.; Zhu, Y.-G.; Xue, X.-M. Exposure to Microplastics Lowers Arsenic Accumulation and Alters Gut Bacterial Communities of Earthworm Metaphire Californica. Environ. Pollut. 2019, 251, 110–116. [Google Scholar] [CrossRef]
- Van de Wiele, T.; Gallawa, C.M.; Kubachk, K.M.; Creed, J.T.; Basta, N.; Dayton, E.A.; Whitacre, S.; Laing, G.D.; Bradham, K. Arsenic Metabolism by Human Gut Microbiota upon In Vitro Digestion of Contaminated Soils. Environ. Health Perspect. 2010, 118, 1004–1009. [Google Scholar] [CrossRef]
- Wang, D.; Yan, J.; Teng, M.; Yan, S.; Zhou, Z.; Zhu, W. In Utero and Lactational Exposure to BDE-47 Promotes Obesity Development in Mouse Offspring Fed a High-Fat Diet: Impaired Lipid Metabolism and Intestinal Dysbiosis. Arch. Toxicol. 2018, 92, 1847–1860. [Google Scholar] [CrossRef]
- Iszatt, N.; Janssen, S.; Lenters, V.; Dahl, C.; Stigum, H.; Knight, R.; Mandal, S.; Peddada, S.; González, A.; Midtvedt, T.; et al. Environmental Toxicants in Breast Milk of Norwegian Mothers and Gut Bacteria Composition and Metabolites in Their Infants at 1 Month. Microbiome 2019, 7, 34. [Google Scholar] [CrossRef] [PubMed]
- Scoville, D.K.; Li, C.Y.; Wang, D.; Dempsey, J.L.; Raftery, D.; Mani, S.; Gu, H.; Cui, J.Y. Polybrominated Diphenyl Ethers and Gut Microbiome Modulate Metabolic Syndrome-Related Aqueous Metabolites in Mice. Drug. Metab. Dispos. 2019, 47, 928–940. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.; Qiao, N.; Li, T.; Yu, R.; Zhai, Q.; Tian, F.; Zhao, J.; Zhang, H.; Chen, W. Dietary Supplementation with Probiotics Regulates Gut Microbiota Structure and Function in Nile Tilapia Exposed to Aluminum. PeerJ 2019, 7, e6963. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Guo, R.; Yang, Y.; Ding, J.; Zhang, Y. Long-Term Effect of Heavy-Metal Pollution on Diversity of Gastrointestinal Microbial Community of Bufo Raddei. Toxicol. Lett. 2016, 258, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, J.; Sun, J.-L. Oral Nickel Changes of Intestinal Microflora in Mice. Curr. Microbiol. 2019, 76, 590–596. [Google Scholar] [CrossRef]
- Wang, N.; Jiang, M.; Zhang, P.; Shu, H.; Li, Y.; Guo, Z.; Li, Y. Amelioration of Cd-Induced Bioaccumulation, Oxidative Stress and Intestinal Microbiota by Bacillus cereus in Carassius auratus Gibelio. Chemosphere 2020, 245, 125613. [Google Scholar] [CrossRef]
- Lin, X.; Zhao, J.; Zhang, W.; He, L.; Wang, L.; Chang, D.; Cui, L.; Gao, Y.; Li, B.; Chen, C.; et al. Acute Oral Methylmercury Exposure Perturbs the Gut Microbiome and Alters Gut-Brain Axis Related Metabolites in Rats. Ecotoxicol. Environ. Saf. 2020, 190, 110130. [Google Scholar] [CrossRef]
- Samsel, A.; Seneff, S. Glyphosate’s Suppression of Cytochrome P450 Enzymes and Amino Acid Biosynthesis by the Gut Microbiome: Pathways to Modern Diseases. Entropy 2013, 15, 1416–1463. [Google Scholar] [CrossRef]
- Martínez-Morales, P.L.; Liste, I. Stem Cells as In Vitro Model of Parkinson’s Disease. Stem Cells Int. 2012, 2012, 980941. [Google Scholar] [CrossRef]
- Pfisterer, U.; Kirkeby, A.; Torper, O.; Wood, J.; Nelander, J.; Dufour, A.; Björklund, A.; Lindvall, O.; Jakobsson, J.; Parmar, M. Direct Conversion of Human Fibroblasts to Dopaminergic Neurons. Proc. Natl. Acad. Sci. USA 2011, 108, 10343–10348. [Google Scholar] [CrossRef]
- Biedler, J.L.; Roffler-Tarlov, S.; Schachner, M.; Freedman, L.S. Multiple Neurotransmitter Synthesis by Human Neuroblastoma Cell Lines and Clones. Cancer Res. 1978, 38, 3751–3757. [Google Scholar] [PubMed]
- Wiatrak, B.; Kubis-Kubiak, A.; Piwowar, A.; Barg, E. PC12 Cell Line: Cell Types, Coating of Culture Vessels, Differentiation and Other Culture Conditions. Cells 2020, 9, 958. [Google Scholar] [CrossRef] [PubMed]
- Xicoy, H.; Wieringa, B.; Martens, G.J.M. The SH-SY5Y Cell Line in Parkinson’s Disease Research: A Systematic Review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Konnova, E.A.; Swanberg, M. Animal Models of Parkinson’s Disease. In Parkinson’s Disease: Pathogenesis and Clinical Aspects; Stoker, T.B., Greenland, J.C., Eds.; Codon Publications: Brisbane, QLD, Australia, 2018; ISBN 978-099-443-816-4. [Google Scholar]
- Breger, L.S.; Fuzzati Armentero, M.T. Genetically Engineered Animal Models of Parkinson’s Disease: From Worm to Rodent. Eur. J. Neurosci. 2019, 49, 533–560. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Vila, M.; Lincoln, S.; McCormack, A.; Picciano, M.; LaFrancois, J.; Yu, X.; Dickson, D.; Langston, W.J.; McGowan, E.; et al. Lack of Nigral Pathology in Transgenic Mice Expressing Human Alpha-Synuclein Driven by the Tyrosine Hydroxylase Promoter. Neurobiol. Dis. 2001, 8, 535–539. [Google Scholar] [CrossRef]
- Miyazaki, I.; Isooka, N.; Imafuku, F.; Sun, J.; Kikuoka, R.; Furukawa, C.; Asanuma, M. Chronic Systemic Exposure to Low-Dose Rotenone Induced Central and Peripheral Neuropathology and Motor Deficits in Mice: Reproducible Animal Model of Parkinson’s Disease. Int. J. Mol. Sci. 2020, 21, 93254. [Google Scholar] [CrossRef]
- Kimmel, C.B. Patterning the Brain of the Zebrafish Embryo. Annu. Rev. Neurosci. 1993, 16, 707–732. [Google Scholar] [CrossRef]
- Rupp, B.; Reichert, H.; Wullimann, M.F. The Zebrafish Brain: A Neuroanatomical Comparison with the Goldfish. Anat. Embryol. 1996, 194, 187–203. [Google Scholar] [CrossRef]
- Ali, S.; Champagne, D.L.; Spaink, H.P.; Richardson, M.K. Zebrafish Embryos and Larvae: A New Generation of Disease Models and Drug Screens. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 115–133. [Google Scholar] [CrossRef]
- Howe, K.; Clark, M.D.; Torroja, C.F.; Torrance, J.; Berthelot, C.; Muffato, M.; Collins, J.E.; Humphray, S.; McLaren, K.; Matthews, L.; et al. The Zebrafish Reference Genome Sequence and Its Relationship to the Human Genome. Nature 2013, 496, 498–503. [Google Scholar] [CrossRef]
- Vaz, R.L.; Outeiro, T.F.; Ferreira, J.J. Zebrafish as an Animal Model for Drug Discovery in Parkinson’s Disease and Other Movement Disorders: A Systematic Review. Front. Neurol. 2018, 9, 347. [Google Scholar] [CrossRef] [PubMed]
- Razali, K.; Othman, N.; Mohd Nasir, M.H.; Doolaanea, A.A.; Kumar, J.; Ibrahim, W.N.; Mohamed Ibrahim, N.; Mohamed, W.M.Y. The Promise of the Zebrafish Model for Parkinson’s Disease: Today’s Science and Tomorrow’s Treatment. Front. Genet. 2021, 12, 655550. [Google Scholar] [CrossRef] [PubMed]
- D’Amora, M.; Giordani, S. The Utility of Zebrafish as a Model for Screening Developmental Neurotoxicity. Front. Neurosci. 2018, 12, 976. [Google Scholar] [CrossRef] [PubMed]
- Caballero, M.V.; Candiracci, M. Zebrafish as Screening Model for Detecting Toxicity and Drugs Efficacy. J. Unexplor. Med. Data 2018, 3, 4. [Google Scholar] [CrossRef]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A Preclinical Model for Drug Screening. Assay Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef]
- Pitchai, A.; Rajaretinam, R.K.; Freeman, J.L. Zebrafish as an Emerging Model for Bioassay-Guided Natural Product Drug Discovery for Neurological Disorders. Medicines 2019, 6, 61. [Google Scholar] [CrossRef]
- Sun, Z.; Gitler, A.D. Discovery and Characterization of Three Novel Synuclein Genes in Zebrafish. Dev. Dyn. 2008, 237, 2490–2495. [Google Scholar] [CrossRef]
- Christensen, C.; Þorsteinsson, H.; Maier, V.H.; Karlsson, K.Æ. Multi-Parameter Behavioral Phenotyping of the MPP+ Model of Parkinson’s Disease in Zebrafish. Front. Behav. Neurosci. 2020, 14, 247. [Google Scholar] [CrossRef]
- Bailone, R.L.; de Aguiar, L.K.; Roca, R.d.O.; Borra, R.C.; Corrêa, T.; Janke, H.; Fukushima, H.C.S. Zebrafish as an Animal Model for Food Safety Research: Trends in the Animal Research. Food Biotechnol. 2019, 33, 283–302. [Google Scholar] [CrossRef]
- Fishman, M.C. Zebrafish—The Canonical Vertebrate. Science 2001, 294, 1290–1291. [Google Scholar] [CrossRef]
- Tsay, H.-J.; Wang, Y.-H.; Chen, W.-L.; Huang, M.-Y.; Chen, Y.-H. Treatment with Sodium Benzoate Leads to Malformation of Zebrafish Larvae. Neurotoxicol. Teratol. 2007, 29, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Dambal, V.Y.; Selvan, K.P.; Lite, C.; Barathi, S.; Santosh, W. Developmental Toxicity and Induction of Vitellogenin in Embryo-Larval Stages of Zebrafish (Danio rerio) Exposed to Methyl Paraben. Ecotoxicol. Environ. Saf. 2017, 141, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Simmons, A.E.; Karimi, I.; Talwar, M.; Simmons, T.W. Effects of Nitrite on Development of Embryos and Early Larval Stages of the Zebrafish (Danio rerio). Zebrafish 2012, 9, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Caioni, G.; Merola, C.; Perugini, M.; d’Angelo, M.; Cimini, A.M.; Amorena, M.; Benedetti, E. An Experimental Approach to Study the Effects of Realistic Environmental Mixture of Linuron and Propamocarb on Zebrafish Synaptogenesis. Int. J. Environ. Res. Public Health 2021, 18, 4664. [Google Scholar] [CrossRef] [PubMed]
- Bridi, D.; Altenhofen, S.; Gonzalez, J.B.; Reolon, G.K.; Bonan, C.D. Glyphosate and Roundup® Alter Morphology and Behavior in Zebrafish. Toxicology 2017, 392, 32–39. [Google Scholar] [CrossRef]
- Adelsbach, T.L.; Tjeerdema, R.S. Chemistry and Fate of Fenvalerate and Esfenvalerate. Rev. Environ. Contam. Toxicol. 2003, 176, 137–154. [Google Scholar] [CrossRef]
- Awoyemi, O.M.; Kumar, N.; Schmitt, C.; Subbiah, S.; Crago, J. Behavioral, Molecular and Physiological Responses of Embryo-Larval Zebrafish Exposed to Types I and II Pyrethroids. Chemosphere 2019, 219, 526–537. [Google Scholar] [CrossRef]
- Corcellas, C.; Feo, M.L.; Torres, J.P.; Malm, O.; Ocampo-Duque, W.; Eljarrat, E.; Barceló, D. Pyrethroids in Human Breast Milk: Occurrence and Nursing Daily Intake Estimation. Environ. Int. 2012, 47, 17–22. [Google Scholar] [CrossRef]
- Amaraneni, M.; Pang, J.; Mortuza, T.B.; Muralidhara, S.; Cummings, B.S.; White, C.A.; Vorhees, C.V.; Zastre, J.; Bruckner, J.V. Brain Uptake of Deltamethrin in Rats as a Function of Plasma Protein Binding and Blood-Brain Barrier Maturation. Neurotoxicology 2017, 62, 24–29. [Google Scholar] [CrossRef]
- Hansen, M.R.H.; Jørs, E.; Lander, F.; Condarco, G.; Debes, F.; Tirado Bustillos, N.; Schlünssen, V. Neurological Deficits After Long-Term Pyrethroid Exposure. Environ. Health Insights 2017, 11, 1178630217700628. [Google Scholar] [CrossRef]
- Nasuti, C.; Brunori, G.; Eusepi, P.; Marinelli, L.; Ciccocioppo, R.; Gabbianelli, R. Early Life Exposure to Permethrin: A Progressive Animal Model of Parkinson’s Disease. J. Pharmacol. Toxicol. Methods 2017, 83, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Elwan, M.A.; Richardson, J.R.; Guillot, T.S.; Caudle, W.M.; Miller, G.W. Pyrethroid Pesticide-Induced Alterations in Dopamine Transporter Function. Toxicol. Appl. Pharmacol. 2006, 211, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Xia, R.; Liu, Z.; Shen, J.; Gong, X.; Hu, Y.; Chen, H.; Yu, Y.; Gao, W.; Wang, C.; et al. Fenvalerate Triggers Parkinson-like Symptom during Zebrafish Development through Initiation of Autophagy and P38 MAPK/MTOR Signaling Pathway. Chemosphere 2020, 243, 125336. [Google Scholar] [CrossRef] [PubMed]
- Akomea-Frempong, S.; Ofosu, I.W.; Owusu-Ansah, E.d.-G.J.; Darko, G. Health Risks Due to Consumption of Pesticides in Ready-to-Eat Vegetables (Salads) in Kumasi, Ghana. Int. J. Food Contam. 2017, 4, 13. [Google Scholar] [CrossRef]
- Nardelli, V.; D’Amico, V.; Ingegno, M.; Della Rovere, I.; Iammarino, M.; Casamassima, F.; Calitri, A.; Nardiello, D.; Li, D.; Quinto, M. Pesticides Contamination of Cereals and Legumes: Monitoring of Samples Marketed in Italy as a Contribution to Risk Assessment. Appl. Sci. 2021, 11, 7283. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| References | |
|---|---|
| Easy maintenance, manipulation, and no time-consuming experimental procedures | [145] |
| Genomic homology with humans | [146] |
| Basic anatomical and physiological pattern conserved | [147,148] |
| Common molecular pathways | [149] |
| Fewer ethical issues | |
| Facilitation in drug administration (dissolution in water) | [147] |
| Faster determination of toxicological endpoints | [150,151] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caioni, G.; Cimini, A.; Benedetti, E. Food Contamination: An Unexplored Possible Link between Dietary Habits and Parkinson’s Disease. Nutrients 2022, 14, 1467. https://doi.org/10.3390/nu14071467
Caioni G, Cimini A, Benedetti E. Food Contamination: An Unexplored Possible Link between Dietary Habits and Parkinson’s Disease. Nutrients. 2022; 14(7):1467. https://doi.org/10.3390/nu14071467
Chicago/Turabian StyleCaioni, Giulia, Annamaria Cimini, and Elisabetta Benedetti. 2022. "Food Contamination: An Unexplored Possible Link between Dietary Habits and Parkinson’s Disease" Nutrients 14, no. 7: 1467. https://doi.org/10.3390/nu14071467
APA StyleCaioni, G., Cimini, A., & Benedetti, E. (2022). Food Contamination: An Unexplored Possible Link between Dietary Habits and Parkinson’s Disease. Nutrients, 14(7), 1467. https://doi.org/10.3390/nu14071467