
Dietary Complex and Slow Digestive Carbohydrates Promote Bone Mass and Improve Bone Microarchitecture during Catch-Up Growth in Rats

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Housing
2.2. Experimental Procedures
2.3. Biochemical Parameteres
2.4. Ex Vivo Densitometry Analysis
2.5. Micro-CT Analysis
2.6. Statistical Analysis
3. Results
3.1. Effect of Food Restriction on Body Weight, Length, Bone Densitometry, and Microstructure
3.2. Effect of the Nutritional Intervention on Body Weight, Body Length, BMD, and BMC, during the Refeeding Period
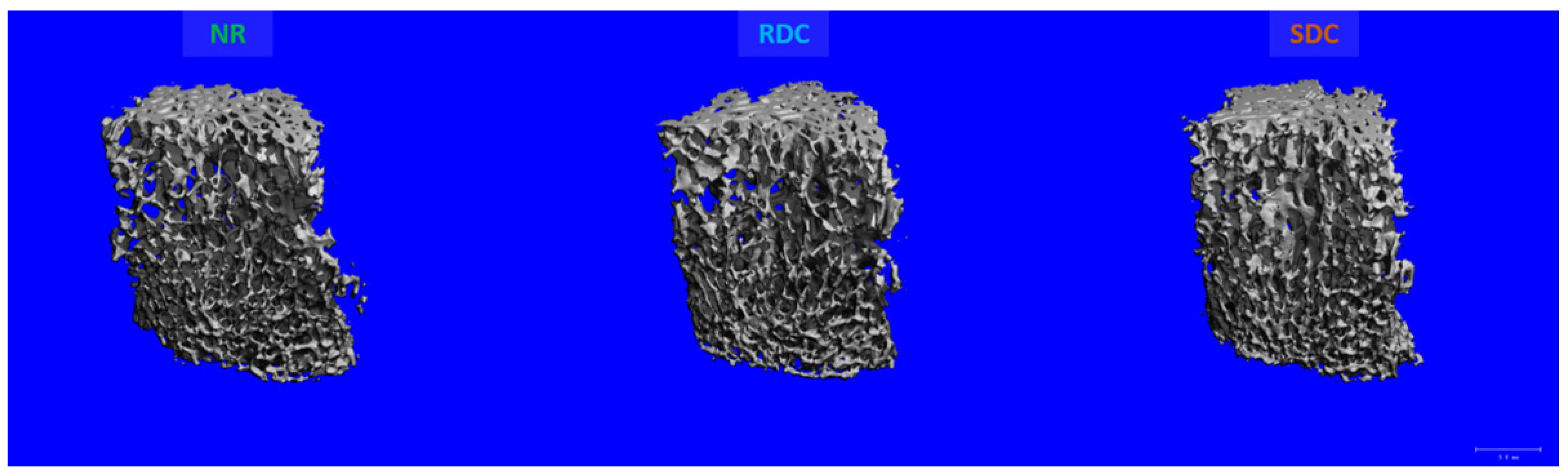
3.3. Effects of the Nutritional Intervention on Trabecular and Cortical Microstructure
3.4. Effects of the Nutritional Intervention on Growth Plate
3.5. Effects of the Nutritional Intervention on Serum Metabolites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Onis, M.; Branca, F. Childhood stunting: A global perspective. Matern. Child Nutr. 2016, 12 (Suppl. 1), 12–26. [Google Scholar] [CrossRef] [PubMed]
- Leroy, J.L.; Frongillo, E.A. Perspective: What Does Stunting Really Mean? A Critical Review of the Evidence. Adv. Nutr. 2019, 10, 196–204. [Google Scholar] [CrossRef]
- Prendergast, A.J.; Humphrey, J.H. The stunting syndrome in developing countries. Paediatr. Int. Child Health 2014, 34, 250–265. [Google Scholar] [CrossRef]
- Kueper, J.; Beyth, S.; Liebergall, M.; Kaplan, L.; Schroeder, J.E. Evidence for the adverse effect of starvation on bone quality: A review of the literature. Int. J. Endocrinol. 2015, 2015, 628740. [Google Scholar] [CrossRef]
- Scheffler, C.; Hermanussen, M.; Bogin, B.; Liana, D.S.; Taolin, F.; Cempaka, P.; Irawan, M.; Ibbibah, L.F.; Mappapa, N.K.; Payong, M.K.E.; et al. Stunting is not a synonym of malnutrition. Eur. J. Clin. Nutr. 2020, 74, 377–386. [Google Scholar] [CrossRef]
- Wit, J.M.; Boersma, B. Catch-up growth: Definition, mechanisms, and models. J. Pediatr. Endocrinol. Metab. 2002, 15 (Suppl. 5), 1229–1241. [Google Scholar] [PubMed]
- Singhal, A. Long-Term Adverse Effects of Early Growth Acceleration or Catch-Up Growth. Ann. Nutr. Metab. 2017, 70, 236–240. [Google Scholar] [CrossRef] [PubMed]
- Golden, N.H.; Abrams, S.A.; Committeeon, N. Optimizing bone health in children and adolescents. Pediatrics 2014, 134, e1229-43. [Google Scholar] [CrossRef]
- Pando, R.; Masarwi, M.; Shtaif, B.; Idelevich, A.; Monsonego-Ornan, E.; Shahar, R.; Phillip, M.; Gat-Yablonski, G. Bone quality is affected by food restriction and by nutrition-induced catch-up growth. J. Endocrinol. 2014, 223, 227–239. [Google Scholar] [CrossRef][Green Version]
- Yoshimura, T.; Tohya, T.; Onoda, C.; Okamura, H. Poor nutrition in prepubertal Japanese children at the end of World War II suppressed bone development. Maturitas 2005, 52, 32–34. [Google Scholar] [CrossRef]
- Masarwi, M.; Gabet, Y.; Dolkart, O.; Brosh, T.; Shamir, R.; Phillip, M.; Gat-Yablonski, G. Skeletal effect of casein and whey protein intake during catch-up growth in young male Sprague-Dawley rats. Br. J. Nutr. 2016, 116, 59–69. [Google Scholar] [CrossRef]
- Bar-Maisels, M.; Gabet, Y.; Shamir, R.; Hiram-Bab, S.; Pasmanik-Chor, M.; Phillip, M.; Bar-Yoseph, F.; Gat-Yablonski, G. Beta Palmitate Improves Bone Length and Quality during Catch-Up Growth in Young Rats. Nutrients 2017, 9, 764. [Google Scholar] [CrossRef] [PubMed]
- Ruppert, K.; Cauley, J.; Lian, Y.; Zgibor, J.C.; Derby, C.; Solomon, D.H. The effect of insulin on bone mineral density among women with type 2 diabetes: A SWAN Pharmacoepidemiology study. Osteoporos Int. 2018, 29, 347–354. [Google Scholar] [CrossRef]
- Liu, Q.; Yang, Z.; Xie, C.; Ling, L.; Hu, H.; Cao, Y.; Huang, Y.; Zhu, Q.; Hua, Y. The Hyperglycemia and Hyperketonemia Impaired Bone Microstructures: A Pilot Study in Rats. Front. Endocrinol. 2020, 11, 590575. [Google Scholar] [CrossRef]
- Grysman, A.; Carlson, T.; Wolever, T.M. Effects of sucromalt on postprandial responses in human subjects. Eur. J. Clin. Nutr. 2008, 62, 1364–1371. [Google Scholar] [CrossRef]
- Holub, I.; Gostner, A.; Theis, S.; Nosek, L.; Kudlich, T.; Melcher, R.; Scheppach, W. Novel findings on the metabolic effects of the low glycaemic carbohydrate isomaltulose (Palatinose). Br. J. Nutr. 2010, 103, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Schwingshackl, L.; Hoffmann, G. Long-term effects of low glycemic index/load vs. high glycemic index/load diets on parameters of obesity and obesity-associated risks: A systematic review and meta-analysis. Nutr. Metab. Cardiovasc Dis. 2013, 23, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Brand-Miller, J.; Hayne, S.; Petocz, Z.; Colagiuri, S. Low–Glycemic Index Diets in the Management of Diabetes. Diabetes Care 2003, 8, 2261–2267. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Y.; Zhang, Y.H.; Wang, P.; Qin, L.Q. Meta-analysis of dietary glycemic load and glycemic index in relation to risk of coronary heart disease. Am. J. Cardiol. 2012, 109, 1608–1613. [Google Scholar] [CrossRef]
- Salto, R.; Giron, M.D.; Ortiz-Moral, C.; Manzano, M.; Vilchez, J.D.; Reche-Perez, F.J.; Bueno-Vargas, P.; Rueda, R.; Lopez-Pedrosa, J.M. Dietary Complex and Slow Digestive Carbohydrates Prevent Fat Deposits During Catch-Up Growth in Rats. Nutrients 2020, 12, 2568. [Google Scholar] [CrossRef]
- Fricke, O.; Schoenau, E. The ’Functional Muscle-Bone Unit’: Probing the relevance of mechanical signals for bone development in children and adolescents. Growth Horm IGF Res. 2007, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rauch, F.; Bailey, D.A.; Baxter-Jones, A.; Mirwald, R.; Faulkner, R. The ’muscle-bone unit’ during the pubertal growth spurt. Bone 2004, 34, 771–775. [Google Scholar] [CrossRef]
- Vandewalle, S.; Taes, Y.; Fiers, T.; Toye, K.; Van Caenegem, E.; Roggen, I.; De Schepper, J.; Kaufman, J.M. Associations of sex steroids with bone maturation, bone mineral density, bone geometry, and body composition: A cross-sectional study in healthy male adolescents. J. Clin. Endocrinol. Metab. 2014, 99, E1272–E1282. [Google Scholar] [CrossRef] [PubMed]
- Hildebrand, T.R.P. A new method for the model-independent assessment of thickness in three-dimensional images. J. Microsc. 1997, 185, 67–75. [Google Scholar] [CrossRef]
- Masrour Roudsari, J.; Mahjoub, S. Quantification and comparison of bone-specific alkaline phosphatase with two methods in normal and paget’s specimens. Casp. J. Intern. Med. 2012, 3, 478–483. [Google Scholar]
- Turan, S.; Topcu, B.; Gokce, I.; Guran, T.; Atay, Z.; Omar, A.; Akcay, T.; Bereket, A. Serum alkaline phosphatase levels in healthy children and evaluation of alkaline phosphatase z-scores in different types of rickets. J. Clin. Res. Pediatr. Endocrinol. 2011, 3, 7–11. [Google Scholar] [CrossRef]
- Boyer, P.M.; Compagnucci, G.E.; Olivera, M.I.; Bozzini, C.; Roig, M.C.; Compagnucci, C.V.; Alippi, R.M. Bone status in an animal model of chronic sub-optimal nutrition: A morphometric, densitometric and mechanical study. Br. J. Nutr. 2005, 93, 663–669. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Cloutier, A.M.; Thomas, N.A.; Panus, D.A.; Lotinun, S.; Pinz, I.; Baron, R.; Rosen, C.J.; Bouxsein, M.L. Caloric restriction leads to high marrow adiposity and low bone mass in growing mice. J. Bone Miner. Res. 2010, 25, 2078–2088. [Google Scholar] [CrossRef] [PubMed]
- Guedes, P.; Zamarioli, A.; Botega, I.I.; Silva, R.; Issa, J.P.M.; Butezloff, M.M.; Sousa, Y.; Ximenez, J.P.B.; Volpon, J.B. Undernutrition impairs the quality of growth plate and trabecular and cortical bones in growing rats1. Acta Cir. Bras. 2019, 34, e201900301. [Google Scholar] [CrossRef]
- Bryk, G.; Coronel, M.Z.; Lugones, C.; Mandalunis, P.; Rio, M.E.; Gualtieri, A.F.; de Portela, M.L.; Zeni, S.N. Effect of a mixture of GOS/FOS((R)) on calcium absorption and retention during recovery from protein malnutrition: Experimental model in growing rats. Eur. J. Nutr. 2016, 55, 2445–2458. [Google Scholar] [CrossRef]
- Alasagheirin, M.H.; Clark, M.K. Skeletal growth, body composition, and metabolic risk among North Sudanese immigrant children. Public Health Nurs. 2018, 35, 91–99. [Google Scholar] [CrossRef]
- Misra, M.; Prabhakaran, R.; Miller, K.K.; Goldstein, M.A.; Mickley, D.; Clauss, L.; Lockhart, P.; Cord, J.; Herzog, D.B.; Katzman, D.K.; et al. Weight gain and restoration of menses as predictors of bone mineral density change in adolescent girls with anorexia nervosa-1. J. Clin. Endocrinol. Metab. 2008, 93, 1231–1237. [Google Scholar] [CrossRef]
- Faje, A.T.; Karim, L.; Taylor, A.; Lee, H.; Miller, K.K.; Mendes, N.; Meenaghan, E.; Goldstein, M.A.; Bouxsein, M.L.; Misra, M.; et al. Adolescent girls with anorexia nervosa have impaired cortical and trabecular microarchitecture and lower estimated bone strength at the distal radius. J. Clin. Endocrinol. Metab. 2013, 98, 1923–1929. [Google Scholar] [CrossRef] [PubMed]
- Most, J.; Tosti, V.; Redman, L.M.; Fontana, L. Calorie restriction in humans: An update. Ageing Res. Rev. 2017, 39, 36–45. [Google Scholar] [CrossRef]
- Gordon, C.M.; Zemel, B.S.; Wren, T.A.; Leonard, M.B.; Bachrach, L.K.; Rauch, F.; Gilsanz, V.; Rosen, C.J.; Winer, K.K. The Determinants of Peak Bone Mass. J. Pediatr. 2017, 180, 261–269. [Google Scholar] [CrossRef] [PubMed]
- Villareal, D.T.; Fontana, L.; Weiss, E.P.; Racette, S.B.; Steger-May, K.; Schechtman, K.B.; Klein, S.; Holloszy, J.O. Bone mineral density response to caloric restriction–induced weight loss or exercise-induced weight loss: A randomized controlled trial. Arch. Intern. Med. 2006, 166, 2502–2510. [Google Scholar] [CrossRef] [PubMed]
- Shapses, S.A.; Sukumar, D. Bone metabolism in obesity and weight loss. Annu Rev. Nutr. 2012, 32, 287–309. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The Laboratory Rat: Relating Its Age With Human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar]
- Fonseca, H.; Moreira-Goncalves, D.; Coriolano, H.J.; Duarte, J.A. Bone quality: The determinants of bone strength and fragility. Sports Med. 2014, 44, 37–53. [Google Scholar] [CrossRef]
- Chen, L.L.; Wang, S.X.; Dai, Y.; Buckoreelall, P.; Zhang, P.; Zhang, H.H.; Kong, W. Effect of catch-up growth by various dietary patterns and resveratrol intervention on bone status. Exp. Biol. Med. 2012, 237, 297–304. [Google Scholar] [CrossRef]
- Berends, L.M.; Fernandez-Twinn, D.S.; Martin-Gronert, M.S.; Cripps, R.L.; Ozanne, S.E. Catch-up growth following intra-uterine growth-restriction programmes an insulin-resistant phenotype in adipose tissue. Int. J. Obes. 2013, 37, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.S.; Yeh, T.H.; King, S.; Mathers, J.; Rybchyn, M.S.; Neist, E.; Cameron, M.; Tacey, A.; Girgis, C.M.; Levinger, I.; et al. Distinct Effects of a High Fat Diet on Bone in Skeletally Mature and Developing Male C57BL/6J Mice. Nutrients 2021, 13, 1666. [Google Scholar] [CrossRef] [PubMed]
- Syggelos, N.; Augoulea, A.; Armeni, E.; Kaparos, G.; Vafaei, H.; Dontas, I.; Marinou, K.; Vaggos, G.; Raptou, P.; Lambrinoudaki, I.; et al. Impact of prenatal and postnatal nutritional manipulation on bone quality in adult Wistar rats offspring. Clin. Nutr. Open Sci. 2021, 35, 34–47. [Google Scholar] [CrossRef]
- Cianfarani, S.; Germani, D.; Branca, F. Low birthweight and adult insulin resistance: The “catch-up growth” hypothesis. Arch. Dis. Child. Fetal Neonatal. Ed. 1999, 81, F71–F73. [Google Scholar] [CrossRef]
- Bilinski, W.J.; Paradowski, P.T.; Sypniewska, G. Bone health and hyperglycemia in pediatric populations. Crit Rev. Clin. Lab. Sci. 2020, 57, 444–457. [Google Scholar] [CrossRef] [PubMed]
- Shi, P.; Hou, A.; Li, C.; Wu, X.; Jia, S.; Cen, H.; Hu, X.; Gong, H. Continuous subcutaneous insulin infusion ameliorates bone structures and mechanical properties in type 2 diabetic rats by regulating bone remodeling. Bone 2021, 153, 116101. [Google Scholar] [CrossRef]
- Sangeetha, P.T.; Ramesh, M.N.; Prapulla, S.G. Recent trends in the microbial production, analysis and application of Fructooligosaccharides. Trends Food Sci. Technol. 2005, 16, 442–457. [Google Scholar] [CrossRef]
- Shyam, S.; Ramadas, A.; Chang, S.K. Isomaltulose: Recent evidence for health benefits. J. Funct. Foods 2018, 48, 173–178. [Google Scholar] [CrossRef]
- Sorndech, W.; Nakorn, K.N.; Tongta, S.; Blennow, A. Isomalto-oligosaccharides: Recent insights in production technology and their use for food and medical applications. Lwt 2018, 95, 135–142. [Google Scholar] [CrossRef]
- Costa, G.T.; Vasconcelos, Q.; Abreu, G.C.; Albuquerquue, A.O.; Vilar, J.L.; Aragao, G.F. Systematic review of the ingestion of fructooligosaccharides on the absorption of minerals and trace elements versus control groups. Clin. Nutr. ESPEN 2021, 41, 68–76. [Google Scholar] [CrossRef]
- Bueno-Vargas, P.; Manzano, M.; Diaz-Castro, J.; Lopez-Aliaga, I.; Rueda, R.; Lopez-Pedrosa, J.M. Maternal Dietary Supplementation with Oligofructose-Enriched Inulin in Gestating/Lactating Rats Preserves Maternal Bone and Improves Bone Microarchitecture in Their Offspring. PLoS ONE 2016, 11, e0154120. [Google Scholar] [CrossRef]
- Rizzoli, R.; Biver, E.; Brennan-Speranza, T.C. Nutritional intake and bone health. Lancet Diabetes Endocrinol. 2021, 9, 606–621. [Google Scholar] [CrossRef]
- Kruger, M.; Gericke, G.; White, Z. Micronutrients and bone growth in preadolescent children from developing countries. Open Nutr. J. 2016, 10, 13–22. [Google Scholar] [CrossRef]
- Rizzoli, R. Nutritional influence on bone: Role of gut microbiota. Aging Clin. Exp. Res. 2019, 31, 743–751. [Google Scholar] [CrossRef]
- Yang, Z.D.; Guo, Y.S.; Huang, J.S.; Gao, Y.F.; Peng, F.; Xu, R.Y.; Su, H.H.; Zhang, P.J. Isomaltulose Exhibits Prebiotic Activity, and Modulates Gut Microbiota, the Production of Short Chain Fatty Acids, and Secondary Bile Acids in Rats. Molecules 2021, 26, 2464. [Google Scholar] [CrossRef]
- Tyagi, A.M.; Yu, M.; Darby, T.M.; Vaccaro, C.; Li, J.Y.; Owens, J.A.; Hsu, E.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; et al. The Microbial Metabolite Butyrate Stimulates Bone Formation via T Regulatory Cell-Mediated Regulation of WNT10B Expression. Immunity 2018, 49, 1116–1131.e7. [Google Scholar] [CrossRef]
- Chen, T.H.; Chen, W.M.; Hsu, K.H.; Kuo, C.D.; Hung, S.C. Sodium butyrate activates ERK to regulate differentiation of mesenchymal stem cells. Biochem. Biophys Res. Commun. 2007, 355, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Lucas, S.; Omata, Y.; Hofmann, J.; Bottcher, M.; Iljazovic, A.; Sarter, K.; Albrecht, O.; Schulz, O.; Krishnacoumar, B.; Kronke, G.; et al. Short-chain fatty acids regulate systemic bone mass and protect from pathological bone loss. Nat. Commun. 2018, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Li, J.Y.; Yu, M.; Pal, S.; Tyagi, A.M.; Dar, H.; Adams, J.; Weitzmann, M.N.; Jones, R.M.; Pacifici, R. Parathyroid hormone-dependent bone formation requires butyrate production by intestinal microbiota. J. Clin. Investig. 2020, 130, 1767–1781. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, N. Subchapter 26A—Parathyroid Hormone. In Handbook of Hormones; Takei, Y., Ando, H., Tsutsui, K., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 224–226. [Google Scholar]
- Clarke, B. Normal bone anatomy and physiology. Clin. J. Am. Soc. Nephrol. 2008, 3 (Suppl. 3), S131–S139. [Google Scholar] [CrossRef]
- Lui, J.C.; Nilsson, O.; Baron, J. Growth Plate Senescence and Catch-Up Growth. In 4th ESPE Advanced Seminar in Developmental Endocrinology, Stockholm, June–July 2010; Camacho-Hübner, C., Nilsson, O., Sävendahl, L., Eds.; Karger: Stockholm, Sweden, 2010. [Google Scholar]
- Nilsson, O.; Baron, J. Fundamental limits on longitudinal bone growth: Growth plate senescence and epiphyseal fusion. Trends Endocrinol. Metab. 2004, 15, 370–374. [Google Scholar] [CrossRef]
- Hunziker, E.B. Elongation of the Long Bones in Humans by the Growth Plates. Nestlé Nutr. Inst. Workshop Ser. 2018, 89, 13–23. [Google Scholar] [PubMed]
- Martin, E.A.; Ritman, E.L.; Turner, R.T. Time course of epiphyseal growth plate fusion in rat tibiae. Bone 2003, 32, 261–267. [Google Scholar] [CrossRef]
- Gat-Yablonski, G.; Shtaif, B.; Abraham, E.; Phillip, M. Nutrition-induced catch-up growth at the growth plate. J. Pediatr Endocrinol. Metab. 2008, 21, 879–893. [Google Scholar] [CrossRef] [PubMed]
- Roach, H.I.; Mehta, G.; Oreffo, R.O.C.; Clarke, N.M.P.; Cooper, C. Temporal Analysis of Rat Growth Plates: Cessation of Growth with Age Despite Presence of a Physis. J. Histochem. Cytochem. 2003, 51, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Masarwi, M.; Shamir, R.; Phillip, M.; Gat-Yablonski, G. Leptin stimulates aromatase in the growth plate: Limiting catch-up growth efficiency. J. Endocrinol. 2018, 237, 229–242. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Peng, Y.; Sharrow, A.C.; Iqbal, J.; Zhang, Z.; Papachristou, D.J.; Zaidi, S.; Zhu, L.L.; Yaroslavskiy, B.B.; Zhou, H.; et al. FSH directly regulates bone mass. Cell 2006, 125, 247–260. [Google Scholar] [CrossRef]
- Zaidi, M.; Lizneva, D.; Kim, S.M.; Sun, L.; Iqbal, J.; New, M.I.; Rosen, C.J.; Yuen, T. FSH, Bone Mass, Body Fat, and Biological Aging. Endocrinology 2018, 159, 3503–3514. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | AIN93M | RDC | SDC |
|---|---|---|---|
| NR | RDC | SDC | |
| CHO (g/100 g diet) | 78.5 | 67.0 | 67.0 |
| Sucrose (g/100 g CHO) | 13.0 | 32.0 | |
| Isomaltulose (g/100 g CHO) | 26.4 | ||
| Sucromalt® (g/100 g CHO) | 22.1 | ||
| Cornstarch (g/100 g CHO) | 61.0 | ||
| MDs (g/100 g CHO) | 20.0 | 64.0 | 23.0 |
| IMOs (g/100 g CHO) | 11.5 | ||
| Resistant MD (g/100 g CHO) | 10.0 | ||
| FOS or Inulin enriched FOS (g/100 g CHO) | 4.0 | 7.0 | |
| Cellulose (g/100 g CHO) | 5.0 | ||
| Protein (g/100 g diet) | 14.0 | 15.0 | 15.0 |
| Fat (g/100 g diet) | 4.0 | 9.8 | 9.8 |
| Micro-CT Parameter | Tibia | Vertebra | ||||
|---|---|---|---|---|---|---|
| NR | RDC | SDC | NR | RDC | SDC | |
| BV/TV (ratio) | 0.124 ± 0.010 | 0.118 ± 0.005 * | 0.164 ± 0.026 *# | 0.261 ± 0.007 | 0.265 ± 0.003 | 0.307 ± 0.014 *# |
| Tb.Th (mm) | 0.064 ± 0.001 | 0.054 ± 0.001 * | 0.051 ± 0.001 *# | 0.076 ± 0.001 | 0.073 ± 0.001 * | 0.072 ± 0.001 * |
| Tb.N (1/mm) | 2.484 ± 0.215 | 2.765 ± 0.179 * | 4.425 ± 0.305 *# | 3.448 ± 0.072 | 3.667 ± 0.168 | 4.064 ± 0.116 *# |
| Tb.Sp (mm) | 0.417 ± 0.039 | 0.374 ± 0.028 * | 0.208 ± 0.013 *# | 0.269 ± 0.006 | 0.251 ± 0.004 * | 0.226 ± 0.008 *# |
| Conn.D (1/mm3) | 59.52 ± 6.703 | 71.31 ± 4.068 * | 141.9 ± 14.10 *# | 69.62 ± 2.901 | 89.62 ± 3.359 * | 111.3 ± 6.682 *# |
| Ct.Th (mm) | 0.576 ± 0.007 | 0.508 ± 0.008 | 0.515 ± 0.010 | 0.259 ± 0.006 | 0.221 ± 0.005 * | 0.242 ± 0.006 *# |
| Tt.Ar (mm2) | 4.749 ± 0.115 | 3.641 ± 0.074 * | 3.686 ± 0.102 * | 3.103 ± 0.08 | 2.400 ± 0.054 * | 2.661 ± 0.104 *# |
| Ct.Ar (mm2) | 4.687 ± 0.103 | 3.609 ± 0.073 * | 3.660 ± 0.102 * | 3.007 ± 0.078 | 2.292 ± 0.058 * | 2.555 ± 0.105 *# |
| Ct.Ar/Tt.Ar (ratio) | 0.993 ± 0.001 | 0.991 ± 0.001 | 0.993 ± 0.001 | 0.970 ± 0.001 | 0.9577 ± 0.003 * | 0.963 ± 0.001 * |
| pMOI (mm4) | 7.216 ± 0.352 | 4.344 ± 0.170 * | 4.423 ± 0.264 * | - | - | - |
| Metabolite | Not Restricted Group | RDC Group | SDC Group |
|---|---|---|---|
| Calcium (mmol/L) | 2.443 ± 0.020 | 2.461 ± 0.018 | 2.467 ± 0.111 |
| Magnesium (mg/dL) | 2.181 ± 0.081 | 2.260 ± 0.090 | 2.327 ± 0.065 |
| Phosphorus (mg/dL) | 5.388 ± 0.111 | 7.122 ± 0.418 * | 6.525 ± 0.211 * |
| ALP (UI/L) | 110.4 ± 6.696 | 160.5 ± 16.140 * | 175.6 ± 6.793 * |
| PTH (pg/mL) | 28.81 ± 4.524 | 45.85 ± 8.638 | 34.27 ±7.557 |
| OPG (pg/mL) | 716.7 ± 67.14 | 618.8 ± 39.04 | 705.5 ± 30.52 |
| GH (pg/mL) | 2957 ± 957.8 | 9327 ± 1840.0 * | 8162 ± 2.152 * |
| Insulin (pg/mL) | 662.1 ± 138.9 | 787.1 ± 159.9 | 589.7 ± 148.1 |
| Leptin (pg/mL) | 5413 ± 618.6 | 6738 ± 790.7 | 3855 ± 620.0 # |
| LH (pg/mL) | 505.5 ± 88.41 | 730.0 ± 127.7 | 644.0 ± 111.1 |
| FSH (pg/mL) | 6635 ± 426.4 | 8077 ± 481.7 * | 6607 ± 307.3 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bueno-Vargas, P.; Manzano, M.; Pérez-Castillo, Í.M.; Rueda, R.; López-Pedrosa, J.M. Dietary Complex and Slow Digestive Carbohydrates Promote Bone Mass and Improve Bone Microarchitecture during Catch-Up Growth in Rats. Nutrients 2022, 14, 1303. https://doi.org/10.3390/nu14061303
Bueno-Vargas P, Manzano M, Pérez-Castillo ÍM, Rueda R, López-Pedrosa JM. Dietary Complex and Slow Digestive Carbohydrates Promote Bone Mass and Improve Bone Microarchitecture during Catch-Up Growth in Rats. Nutrients. 2022; 14(6):1303. https://doi.org/10.3390/nu14061303
Chicago/Turabian StyleBueno-Vargas, Pilar, Manuel Manzano, Íñigo M. Pérez-Castillo, Ricardo Rueda, and José M. López-Pedrosa. 2022. "Dietary Complex and Slow Digestive Carbohydrates Promote Bone Mass and Improve Bone Microarchitecture during Catch-Up Growth in Rats" Nutrients 14, no. 6: 1303. https://doi.org/10.3390/nu14061303
APA StyleBueno-Vargas, P., Manzano, M., Pérez-Castillo, Í. M., Rueda, R., & López-Pedrosa, J. M. (2022). Dietary Complex and Slow Digestive Carbohydrates Promote Bone Mass and Improve Bone Microarchitecture during Catch-Up Growth in Rats. Nutrients, 14(6), 1303. https://doi.org/10.3390/nu14061303