
The Macronutrient Composition of Infant Formula Produces Differences in Gut Microbiota Maturation That Associate with Weight Gain Velocity and Weight Status

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants and Trial Design
2.1.1. Study Cohort
2.1.2. Composition of Infant Formula Diets
2.1.3. Clinical Phenotypes
2.2. Methodology
2.2.1. Shotgun Metagenomics
2.2.2. Targeted Metabolomics
2.3. Statistical Analyses
3. Results
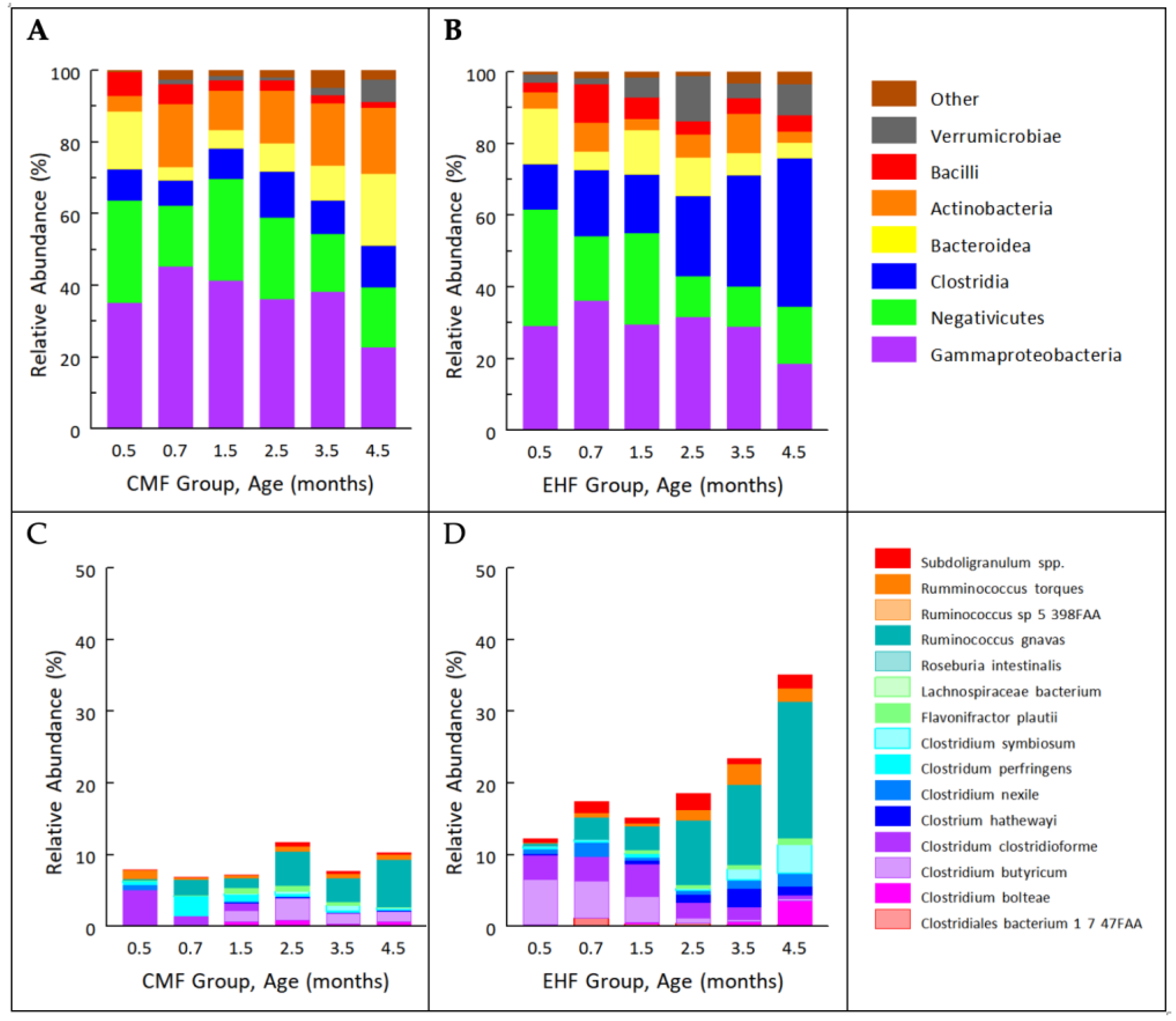
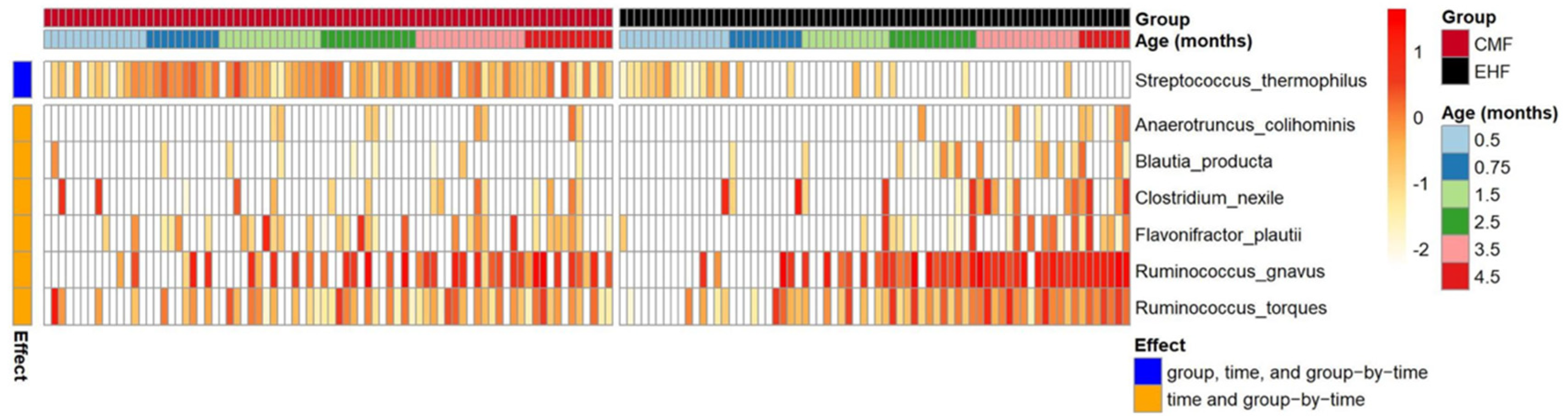
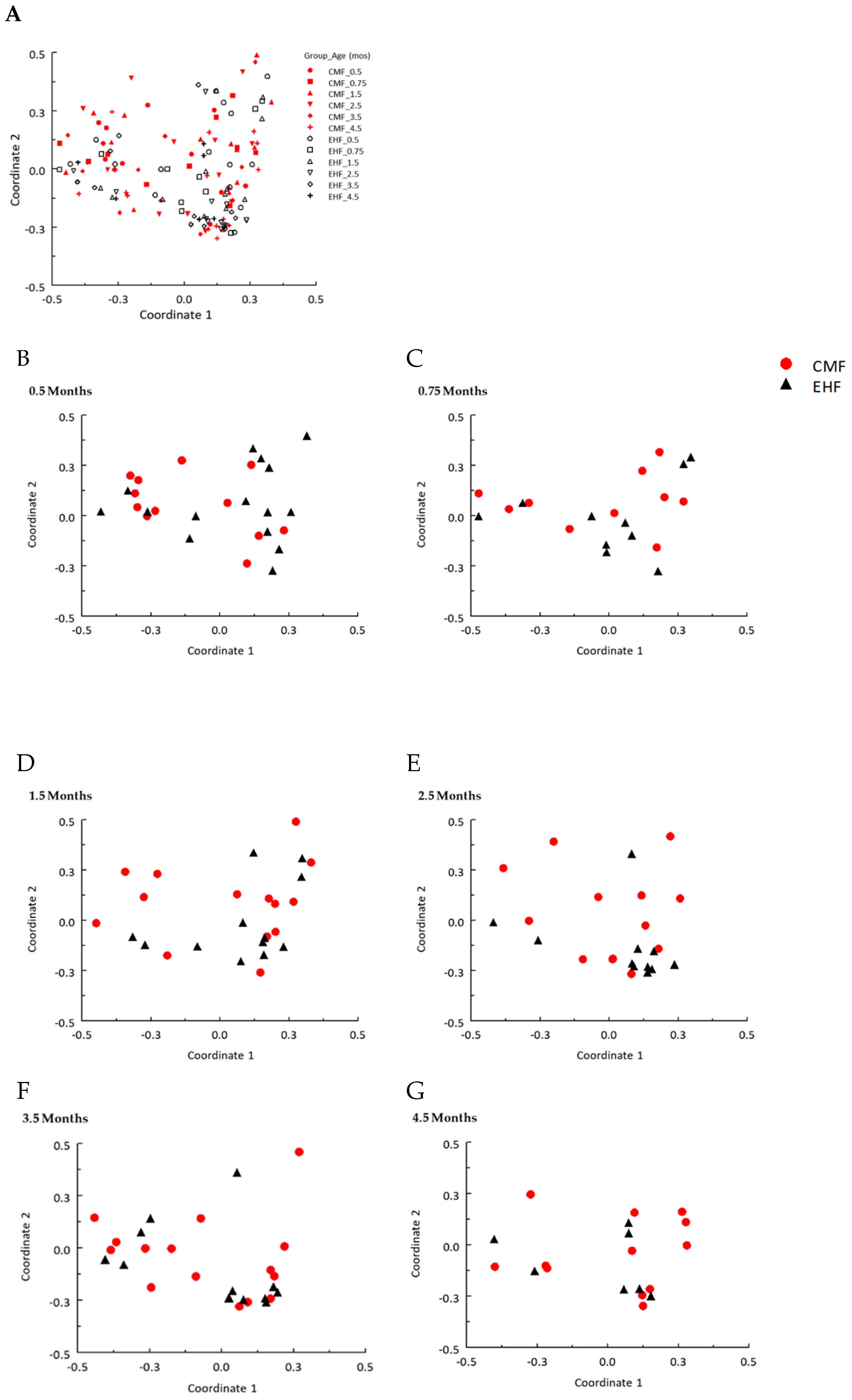
3.1. Outgrowth of Ruminococcus Gnavus and Other Clostridia Species Driven by Formula-Induced Differences in Gut Microbiota
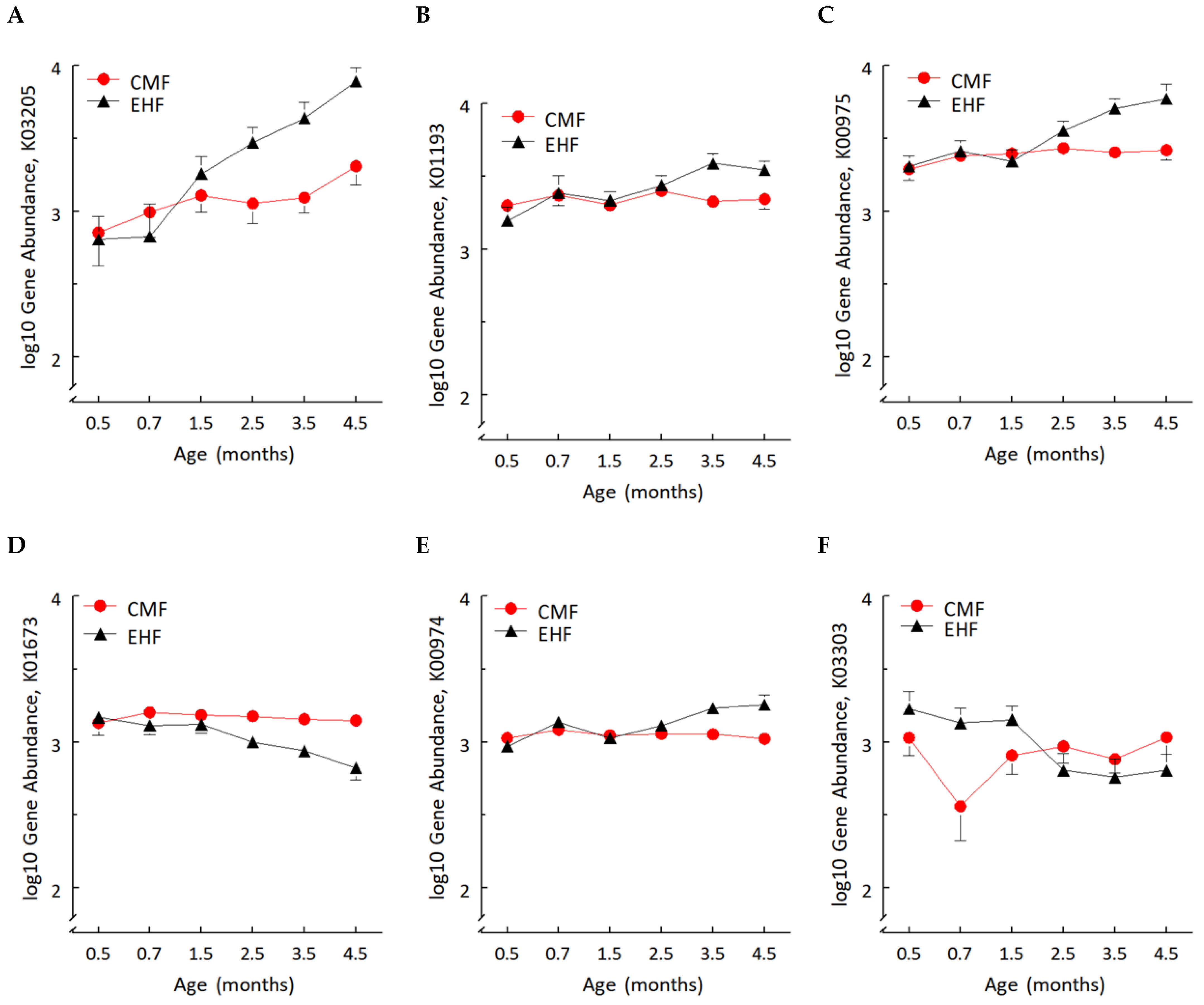
3.2. Formula-Induced Differences in Expression of Genes Related to Carbohydrate Metabolism
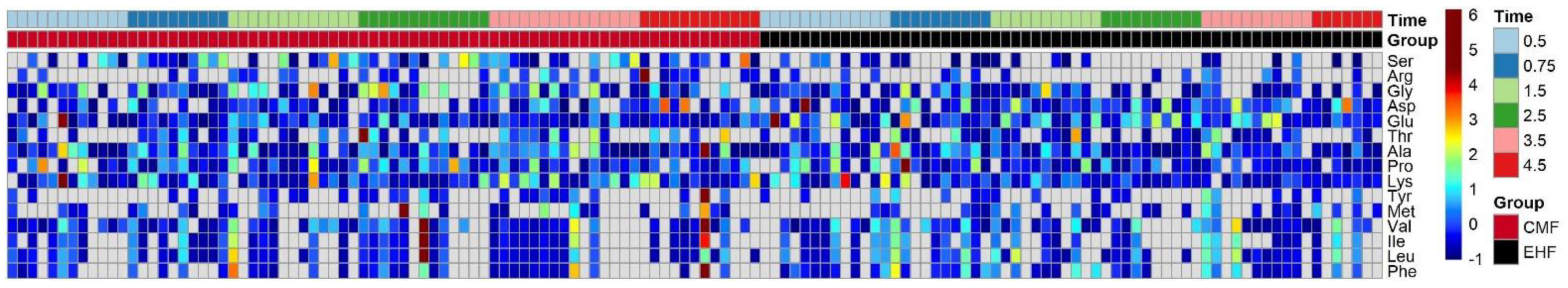
3.3. The Two Groups Shared Similar Fecal Amino Acid Concentrations despite Different Free Amino Acid Concentrations in CMF and EHF
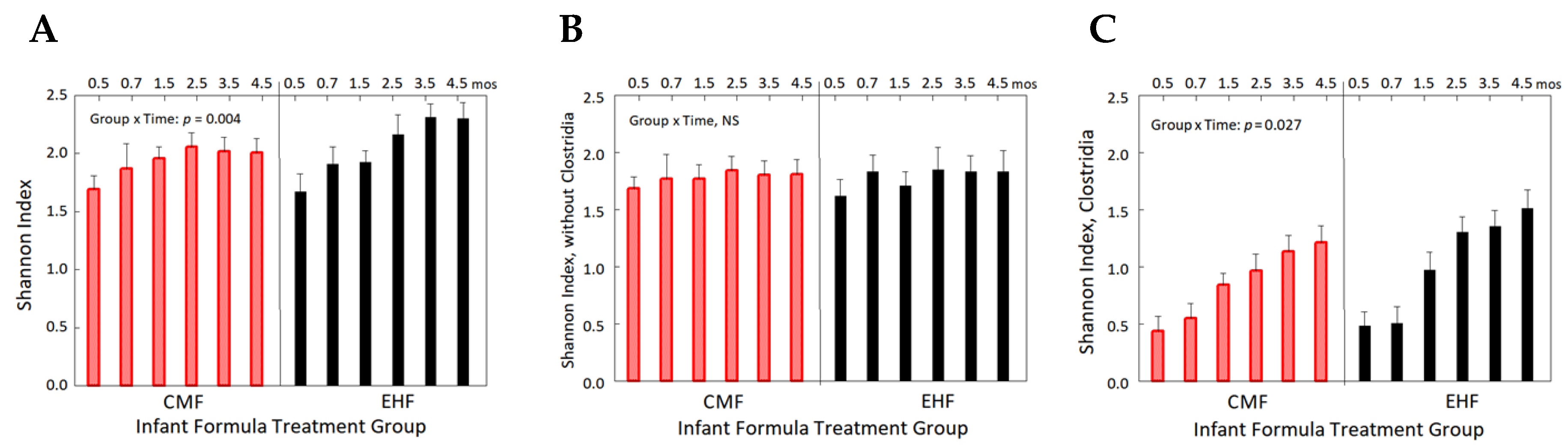
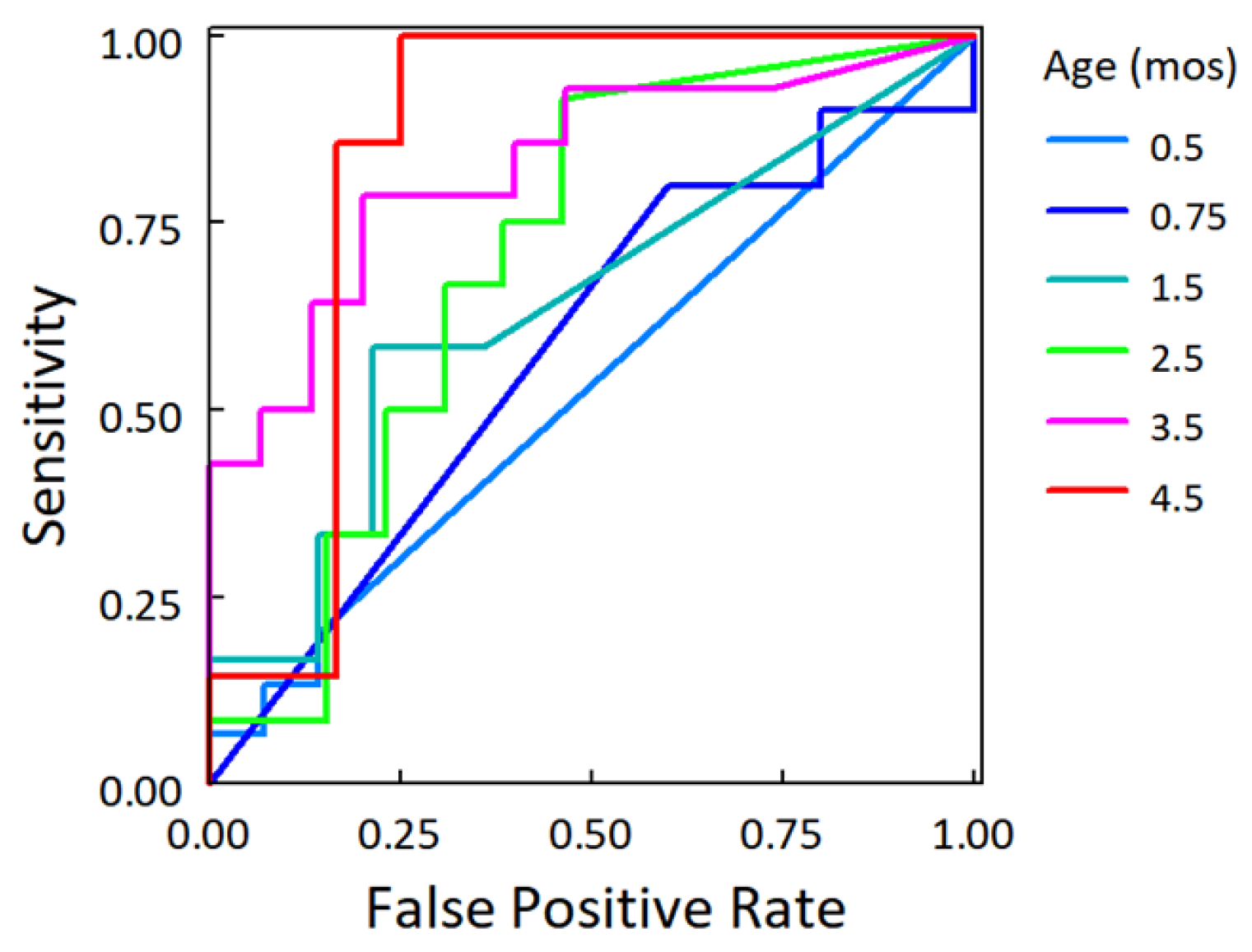
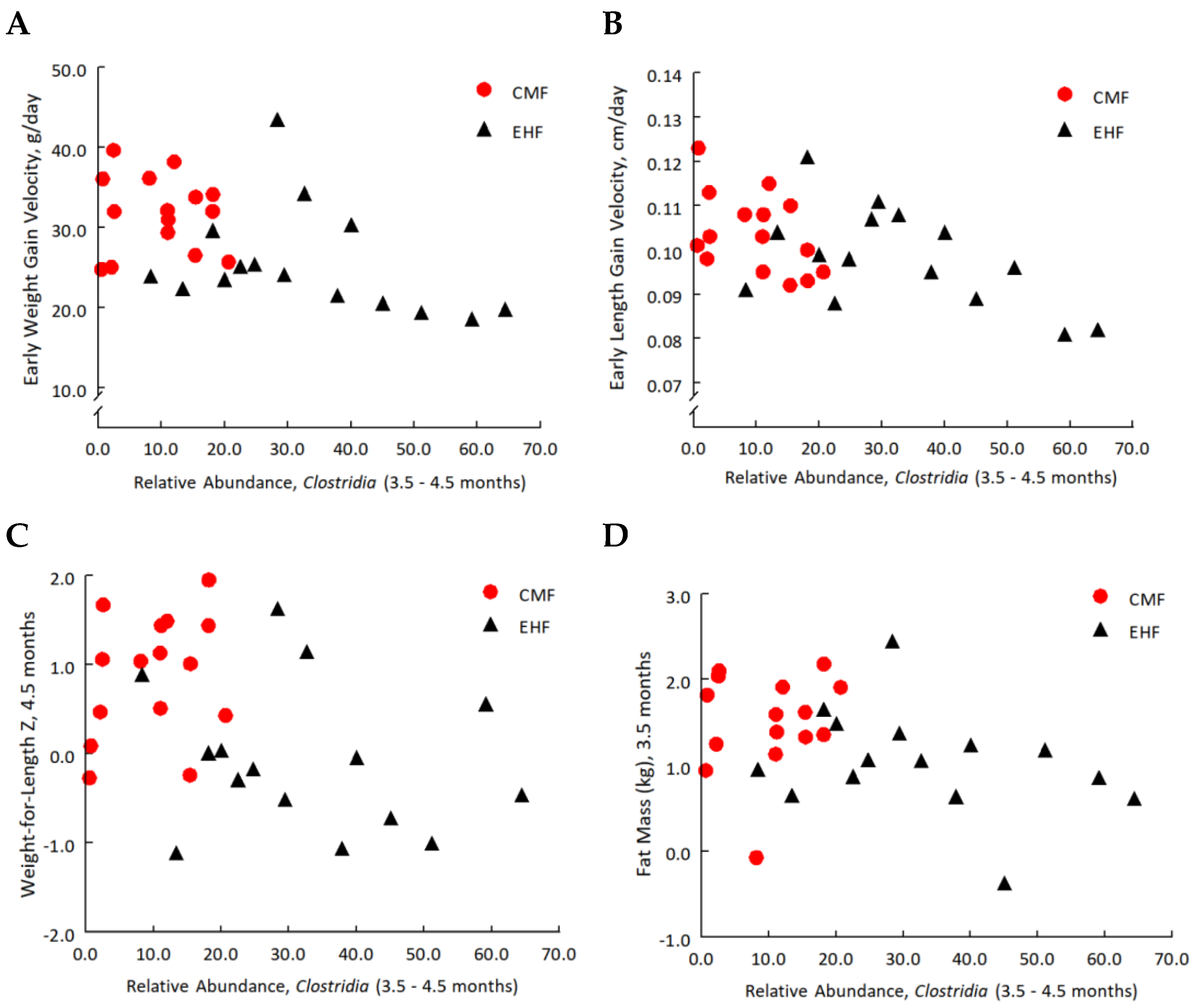
3.4. Increases in the Relative Abundance of Clostridia Related to Leaner Phenotypes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Time, β Coefficient | Time, p-Value |
|---|---|---|
| Actinomyces odontolyticus | 0.12 | 3.4 × 10−5 |
| Akkermansia muciniphila | 0.28 | 3.1 × 10−5 |
| Anaerostipes, unclassified | 0.25 | 6.0 × 10−8 |
| Bacteroides uniformis | 0.07 | 0.02 |
| Bifidobacterium bifidum | 0.35 | 1.0 × 10−5 |
| Bifidobacterium breve | 0.26 | 1.5 × 10−4 |
| Bifidobacterium longum | 0.39 | 3.7 × 10−5 |
| Bifidobacterium pseudocatenulatum | 0.11 | 0.02 |
| Bilophila, unclassified | 0.06 | 0.01 |
| Clostridiales bacterium 1 7 47FAA | 0.23 | 9.0 × 10−6 |
| Clostridium bartlettii | 0.20 | 0.02 |
| Clostridium bolteae | 0.22 | 3.7 × 10−5 |
| Clostridium difficile | 0.28 | 8.3 × 10−8 |
| Clostridium hathewayi | 0.11 | 0.03 |
| Clostridium perfringens | −0.26 | 4.5 × 10−5 |
| Clostridium ramosum | 0.36 | 1.4 × 10−9 |
| Clostridium symbiosum | 0.13 | 3.7 × 10−5 |
| Collinsella aerofaciens | 0.22 | 4.4 × 10−4 |
| Coprobacillus, unclassified | 0.43 | 5.4 × 10−13 |
| Eggerthella, unclassified | 0.12 | 7.8 × 10−5 |
| Enterococcus avium | 0.11 | 1.6 × 10−5 |
| Enterococcus faecalis | −0.12 | 0.02 |
| Erysipelotrichaceae bacterium 21 3 | 0.14 | 3.6 × 10−5 |
| Erysipelotrichaceae bacterium 2 2 44A | 0.12 | 3.0 × 10−3 |
| Erysipelotrichaceae bacterium 6 1 45 | 0.09 | 0.02 |
| Eubacterium limosum | 0.06 | 0.01 |
| Granulicatella, unclassified | 0.08 | 1.2 × 10−3 |
| Klebsiella pneumoniae | 0.09 | 0.01 |
| Lachnospiraceae bacterium 2 1 58FAA | 0.23 | 6.0 × 10−8 |
| Lachnospiraceae bacterium 7 1 58FAA | 0.06 | 0.01 |
| Lachnospiraceae bacterium 9 1 43BFAA | 0.10 | 1.3 × 10−5 |
| Lactobacillus fermentum | −0.13 | 13.5 × 10−4 |
| Lactobacillus gasseri | −0.24 | 3.3 × 10−5 |
| Lactococcus lactis | 0.19 | 3.8 × 10−3 |
| Megasphaera, unclassified | 0.19 | 0.04 |
| Parabacteroides distasonis | 0.58 | 0.03 |
| Solobacterium moorei | 0.06 | 7.1 × 10−5 |
| Staphylococcus hominis | −0.11 | 1.2 × 10−4 |
| Streptococcus mitis oralis pneumoniae | 0.08 | 0.01 |
| Streptococcus peroris | 0.09 | 0.01 |
| Streptococcus salivarius | −0.33 | 8.3 × 10−8 |
| Streptococcus vestibularis | −0.41 | 8.3 × 10−8 |
| Subdoligranulum, unclassified | 0.16 | 9.9 × 10−6 |
| Veillonella atypica | −0.18 | 1.2 × 10−4 |
| Veillonella parvula | −0.13 | 0.02 |
| Veillonella ratti | 0.30 | 4.3 × 10−4 |
| Veillonella, unclassified | −0.15 | 0.01 |
References
- Koleva, P.T.; Kim, J.-S.; Scott, J.; Kozyrskyj, A.L. Microbial programming of health and disease starts during fetal life. Birth Defects Res. Part C Embryo Today Rev. 2015, 105, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and stabilization of the human gut microbiome during the first year of life. Cell Host Microbe 2015, 17, 852. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Fanaro, S.; Chierici, R.; Guerrini, P.; Vigi, V. Intestinal microflora in early infancy: Composition and development. Acta Paediatr. 2007, 92, 48–55. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Sotelo-Orozco, J.; Rudolph, C.; Lönnerdal, B.; Slupsky, C.M. The role of protein and free amino acids on intake, metabolism, and gut microbiome: A comparison between breast-fed and formula-fed rhesus monkey infants. Front. Pediatr. 2020, 7, 563. [Google Scholar] [CrossRef]
- Buddington, R.K.; Sangild, P.T. Companion Animals Symposium: Development of the mammalian gastrointestinal tract, the resident microbiota, and the role of diet in early life. J. Anim. Sci. 2011, 89, 1506–1519. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Schloss, P.D. Dynamics and associations of microbial community types across the human body. Nat. 2014, 509, 357–360. [Google Scholar] [CrossRef]
- Ho, N.T.; Li, F.; Lee-Sarwar, K.A.; Tun, H.M.; Brown, B.; Pannaraj, P.S.; Bender, J.M.; Azad, M.B.; Thompson, A.L.; Weiss, S.T.; et al. Meta-analysis of effects of exclusive breastfeeding on infant gut microbiota across populations. Nat. Commun. 2018, 9, 4169. [Google Scholar] [CrossRef]
- Ma, J.; Li, Z.; Zhang, W.; Zhang, C.; Zhang, Y.; Mei, H.; Zhuo, N.; Wang, H.; Wang, L.; Wu, D. Comparison of gut microbiota in exclusively breast-fed and formula-fed babies: A study of 91 term infants. Sci. Rep. 2020, 10, 15792. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Chung, J.; Battaglia, T.; Henderson, N.; Jay, M.; Li, H.; Lieber, A.D.; Wu, F.; Perez-Perez, G.I.; Chen, Y.; et al. Antibiotics, birth mode, and diet shape microbiome maturation during early life. Sci. Transl. Med. 2016, 8, 343ra82. [Google Scholar] [CrossRef]
- Guaraldi, F.; Salvatori, G. Effect of breast and formula feeding on gut microbiota shaping in newborns. Front. Cell. Infect. Microbiol. 2012, 2, 94. [Google Scholar] [CrossRef] [PubMed]
- Mackie, R.I.; Sghir, A.; Gaskins, H.R. Developmental microbial ecology of the neonatal gastrointestinal tract. Am. J. Clin. Nutr. 1999, 69, 1035s–1045s. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.P.J.G.; Dejong, C.H.C.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Re-print of “Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol. Res. 2013, 69, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Allison, C.; Macfarlane, G.T. Influence of pH, nutrient availability, and growth rate on amine production by Bacteroides fragilis and Clostridium perfringens. Appl. Environ. Microbiol. 1989, 55, 2894–2898. [Google Scholar] [CrossRef] [PubMed]
- Gallier, S.; Van den Abbeele, P.; Prosser, C. Comparison of the bifidogenic effects of goat and cow milk-based infant formulas to human breast milk in an in vitro gut model for 3-month-old infants. Front. Nutr. 2020, 7, 608495. [Google Scholar] [CrossRef]
- Rossen, L.M.; Simon, A.E.; Herrick, K.A. Types of infant formulas consumed in the United States. Clin. Pediatr. 2016, 55, 278–285. [Google Scholar] [CrossRef]
- Ventura, A.K.; San Gabriel, A.; Hirota, M.; Mennella, J.A. Free amino acid content in infant formulas. Nutr. Food Sci. 2012, 42, 271–278. [Google Scholar] [CrossRef]
- Romaní-Pérez, M.; Bullich-Vilarrubias, C.; López-Almela, I.; Liébana-García, R.; Olivares, M.; Sanz, Y. The microbiota and the gut-brain axis in controlling food intake and energy homeostasis. Int. J. Mol. Sci. 2021, 22, 5830. [Google Scholar] [CrossRef]
- Achour, L.; Flourié, B.; Briet, F.; Franchisseur, C.; Bornet, F.; Champ, M.; Rambaud, J.C.; Messing, B. Metabolic effects of digestible and partially indigestible cornstarch: A study in the absorptive and postabsorptive periods in healthy humans. Am. J. Clin. Nutr. 1997, 66, 1151–1159. [Google Scholar] [CrossRef][Green Version]
- Rzehak, P.; Sausenthaler, S.; Koletzko, S.; Reinhardt, D.; Von Berg, A.; Krämer, U.; Berdel, D.; Bollrath, C.; Grübl, A.; Bauer, C.P.; et al. Short- and long-term effects of feeding hydrolyzed protein infant formulas on growth at < or = 6 y of age: Results from the German Infant Nutritional Intervention Study. Am. J. Clin. Nutr. 2009, 89, 1846–1856. [Google Scholar] [CrossRef] [PubMed]
- Mennella, J.A.; Ventura, A.K.; Beauchamp, G.K. Differential growth patterns among healthy infants fed protein hydrolysate or cow-milk formulas. Pediatrics 2011, 127, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Giovannini, M.; Agostoni, C.; Fiocchi, A.; Bellú, R.; Trojan, S.; Riva, E. Antigen-reduced infant formulas versus human milk: Growth and metabolic parameters in the first 6 months of life. J. Am. Coll. Nutr. 1994, 13, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Gappa, M.; Filipiak-Pittroff, B.; Libuda, L.; von Berg, A.; Koletzko, S.; Bauer, C.; Heinrich, J.; Schikowski, T.; Berdel, D.; Standl, M. Long-term effects of hydrolyzed formulae on atopic diseases in the GINI study. Allergy 2021, 76, 1903–1907. [Google Scholar] [CrossRef]
- Laursen, M.F.; Andersen, L.B.B.; Michaelsen, K.F.; Mølgaard, C.; Trolle, E.; Bahl, M.I.; Licht, T.R. Infant gut microbiota development is driven by transition to family foods independent of maternal obesity. mSphere 2016, 1, e00069-15. [Google Scholar] [CrossRef]
- Mennella, J.A.; Inamdar, L.; Pressman, N.; Schall, J.I.; Papas, M.A.; Schoeller, D.; Stallings, V.A.; Trabulsi, J.C. Type of infant formula increases early weight gain and impacts energy balance: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 1015–1025. [Google Scholar] [CrossRef]
- Mennella, J.; Trabulsi, J.; Inamdar, L.; Preedy, V.R.; Watson, R.R.; Zibadi, S. The sensory world of formula-fed infants: Differences among artificial milk feedings in flavor learning and satiation. In Handbook of Dietary and Nutritional Aspects of Bottle Feeding; Preedy, V.R., Watson, R.R., Zibadi, S., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2014; pp. 95–116. [Google Scholar]
- Terheggen-Lagro, S.W.; Khouw, I.M.; Schaafsma, A.; Wauters, E.A. Safety of a new extensively hydrolysed formula in children with cow’s milk protein allergy: A double blind crossover study. BMC Pediatr. 2002, 2, 10. [Google Scholar] [CrossRef]
- WHO Multicentre Growth Reference Study Group. World Health Organization (WHO) Child Growth Standards: Length/Height-for-Age, Weight-for-Age, Weight-for-Length, Weight-for-Height and Body Mass Index-for-Age: Methods and Development; WHO Multicentre Growth Reference Study Group: Geneva, Switzerland, 2006. [Google Scholar]
- Roy, S.M.; Spivack, J.G.; Faith, M.S.; Chesi, A.; Mitchell, J.A.; Kelly, A.; Grant, S.F.A.; McCormack, S.E.; Zemel, B.S. Infant BMI or weight-for-length and obesity risk in early childhood. Pediatrics 2016, 137, e20153492. [Google Scholar] [CrossRef]
- Schoeller, D.A.; Hnilicka, J.M. Reliability of the doubly labeled water method for the measurement of total daily energy ex-penditure in free-living subjects. J. Nutr. 1996, 126, 348S–354S. [Google Scholar]
- Fomon, S.J.; Haschke, F.; Ziegler, E.E.; Nelson, S.E. Body composition of reference children from birth to age 10 years. Am. J. Clin. Nutr. 1982, 35, 1169–1175. [Google Scholar] [CrossRef]
- Racette, S.B.; Schoeller, D.A.; Luke, A.H.; Shay, K.; Hnilicka, J.; Kushner, R.F. Relative dilution spaces of 2H- and 18O-labeled water in humans. Am. J. Physiol. Metab. 1994, 267, E585–E590. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Chen, E.Z.; Baldassano, R.N.; Otley, A.R.; Griffiths, A.M.; Lee, D.; Bittinger, K.; Bailey, A.; Friedman, E.S.; Hoffmann, C.; et al. Inflammation, antibiotics, and diet as environmental stressors of the gut microbiome in pediatric Crohn’s disease. Cell Host Microbe 2015, 18, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Truong, D.T.; Franzosa, E.A.; Tickle, T.L.; Scholz, M.; Weingart, G.; Pasolli, E.; Tett, A.; Huttenhower, C.; Segata, N. Meta, Phl, An2 for enhanced metagenomic taxonomic profiling. Nat. Methods 2015, 12, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef]
- Ni, J.; Shen, T.-C.D.; Chen, E.Z.; Bittinger, K.; Bailey, A.; Roggiani, M.; Sirota-Madi, A.; Friedman, E.S.; Chau, L.; Lin, A.; et al. A role for bacterial urease in gut dysbiosis and Crohn’s disease. Sci. Transl. Med. 2017, 9, eaah6888. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef]
- Mennella, J.A.; Reiter, A.; Brewer, B.; Pohlig, R.T.; Stallings, V.A.; Trabulsi, J.C. Early weight gain forecasts accelerated eruption of deciduous teeth and later overweight status during the first year. J. Pediatr. 2020, 225, 174–181. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: http://www.R-project.org (accessed on 22 September 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Kastl, A.J., Jr.; Terry, N.A.; Wu, G.D.; Albenberg, L.G. The structure and function of the human small intestinal microbiota: Current understanding and future directions. Cell. Mol. Gastroenterol. Hepatol. 2020, 9, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Roswall, J.; Olsson, L.M.; Kovatcheva-Datchary, P.; Nilsson, S.; Tremaroli, V.; Simon, M.-C.; Kiilerich, P.; Akrami, R.; Krämer, M.; Uhlén, M.; et al. Developmental trajectory of the healthy human gut microbiota during the first 5 years of life. Cell Host Microbe 2021, 29, 765–776.e3. [Google Scholar] [CrossRef] [PubMed]
- Trabulsi, J.C.; Smethers, A.D.; Eosso, J.R.; Papas, M.A.; Stallings, V.A.; Mennella, J.A. Impact of early rapid weight gain on odds for overweight at one year differs between breastfed and formula-fed infants. Pediatr. Obes. 2020, 15, 12688. [Google Scholar] [CrossRef] [PubMed]
- Johnson, C.L.; Versalovic, J. The human microbiome and its potential importance to pediatrics. Pediatrics 2012, 129, 950–960. [Google Scholar] [CrossRef] [PubMed]
- Houghteling, P.; Walker, W.A. Why is initial bacterial colonization of the intestine important to infants’ and children’s health? J. Pediatr. Gastroenterol. Nutr. 2015, 60, 294–307. [Google Scholar] [CrossRef]
- Bittinger, K.; Zhao, C.; Li, Y.; Ford, E.; Friedman, E.S.; Ni, J.; Kulkarni, C.V.; Cai, J.; Tian, Y.; Liu, Q.; et al. Bacterial colonization reprograms the neonatal gut metabolome. Nat. Microbiol. 2020, 5, 838–847. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Scaria, J.; Suzuki, H.; Ptak, C.P.; Chen, J.-W.; Zhu, Y.; Guo, X.-K.; Chang, Y.-F. Comparative genomic and phenomic analysis of Clostridium difficile and Clostridium sordellii, two related pathogens with differing host tissue preference. BMC Genom. 2015, 16, 448. [Google Scholar] [CrossRef]
- Tracy, B.P.; Jones, S.W.; Fast, A.G.; Indurthi, D.C.; Papoutsakis, E.T. Clostridia: The importance of their exceptional substrate and metabolite diversity for biofuel and biorefinery applications. Curr. Opin. Biotechnol. 2012, 23, 364–381. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Palmas, V.; Pisanu, S.; Madau, V.; Casula, E.; Deledda, A.; Cusano, R.; Uva, P.; Vascellari, S.; Loviselli, A.; Manzin, A.; et al. Gut microbiota markers associated with obesity and overweight in Italian adults. Sci. Rep. 2021, 11, 5532. [Google Scholar] [CrossRef] [PubMed]
- Mihatsch, W.A.; Högel, J.; Pohlandt, F. Hydrolysed protein accelerates the gastrointestinal transport of formula in preterm infants. Acta Paediatr. 2001, 90, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Roager, H.M.; Hansen, L.B.S.; Bahl, M.I.; Frandsen, H.L.; Carvalho, V.; Gøbel, R.J.; Dalgaard, M.D.; Plichta, D.R.; Sparholt, M.H.; Vestergaard, H.; et al. Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat. Microbiol. 2016, 1, 16093. [Google Scholar] [CrossRef] [PubMed]
- Mead, G.C. The Amino Acid-fermenting Clostridia. J. Gen. Microbiol. 1971, 67, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wu, A.C.; Go, R.M.; Malouf, J.; Turner, M.S.; Malde, A.K.; Mark, A.E.; Gilbert, R.G. The characterization of modified starch branching enzymes: Toward the control of starch chain-length distributions. PLoS ONE 2015, 10, e0125507. [Google Scholar] [CrossRef] [PubMed]
- Crost, E.H.; Tailford, L.E.; Le Gall, G.; Fons, M.; Henrissat, B.; Juge, N. Utilisation of mucin glycans by the human gut symbiont Ruminococcus gnavus is strain-dependent. PLoS ONE 2013, 8, e76341. [Google Scholar] [CrossRef]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.; Florin, T.H.J. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353.e1321. [Google Scholar] [CrossRef]
- Walker, A.; Ince, J.; Duncan, S.H.; Webster, L.M.; Holtrop, G.; Ze, X.; Brown, D.; Stares, M.D.; Scott, P.; Bergerat, A.; et al. Dominant and diet-responsive groups of bacteria within the human colonic microbiota. ISME J. 2010, 5, 220–230. [Google Scholar] [CrossRef]
- Wright, H.D. Direct fermentation of disaccharides and variation in sugar utilization by Streptococcus thermophilus. J. Path Bacteriol. 1936, 43, 487–501. [Google Scholar] [CrossRef]
- Hutkins, R.; Morris, H.A.; McKay, L.L. Galactose transport in Streptococcus thermophilus. Appl. Environ. Microbiol. 1985, 50, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Sato, Y.; Kawashima, M. KEGG mapping tools for uncovering hidden features in biological data. Protein Sci. 2021, 31, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, P.O.A.; Victora, C. Rapid growth in infancy and childhood and obesity in later life—A systematic review. Obes. Rev. 2005, 6, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Pereira Indiani, C.M.; Rizzardi, K.F.; Castelo, P.M.; Ferraz, L.F.C.; Darrieux, M.; Parisotto, T. Childhood obesity and Firmicutes/Bacteroidetes ratio in the gut microbiota: A systematic review. Child. Obes. 2018, 14, 501–509. [Google Scholar] [CrossRef]
- Castaner, O.; Goday, A.; Park, Y.-M.; Lee, S.-H.; Magkos, F.; Shiow, S.-A.T.E.; Schröder, H. The gut microbiome profile in obesity: A systematic review. Int. J. Endocrinol. 2018, 2018, 4095789. [Google Scholar] [CrossRef]
- Mennella, J.A.; Papas, M.A.; Reiter, A.R.; Stallings, V.A.; Trabulsi, J.C. Early rapid weight gain among formula-fed infants: Impact of formula type and maternal feeding styles. Pediatr. Obes. 2019, 14, e12503. [Google Scholar] [CrossRef]
- Petersen, C.; Bell, R.; Klag, K.A.; Lee, S.-H.; Soto, R.; Ghazaryan, A.; Buhrke, K.; Ekiz, H.A.; Ost, K.S.; Boudina, S.; et al. T cell-mediated regulation of the microbiota protects against obesity. Science 2019, 365, eaat9351. [Google Scholar] [CrossRef]
- Von Berg, A.; Filipiak-Pittroff, B.; Kramer, U.; Link, E.; Heinrich, J.; Koletzko, S.; Grubl, A.; Hoffmann, U.; Beckmann, C.; Reinhardt, D.; et al. The German Infant Nutritional Intervention Study (GINI) for the preventive effect of hydrolyzed infant formulas in infants at high risk for allergic diseases. Design and selected results. Allergol. Select. 2017, 1, 28–38. [Google Scholar] [CrossRef]
- Ecklu-Mensah, G.; Gilbert, J.; Devkota, S. Dietary selection pressures and their impact on the gut microbiome. Cell. Mol. Gastroenterol. Hepatol. 2021, 13, 7–18. [Google Scholar] [CrossRef]
- Alexander, M.; Turnbaugh, P.J. Deconstructing mechanisms of diet-microbiome-immune interactions. Immunity 2020, 53, 264–276. [Google Scholar] [CrossRef] [PubMed]
- Ventura, A.K.; Beauchamp, G.K.; Mennella, J.A. Infant regulation of intake: The effect of free glutamate content in infant formulas. Am. J. Clin. Nutr. 2012, 95, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, B.; von Kries, R.; Monasterolo, R.C.; Subias, J.E.; Scaglioni, S.; Giovannini, M.; Beyer, J.; Demmelmair, H.; Anton, B.; Gruszfeld, D.; et al. Infant feeding and later obesity risk. Adv. Exp. Med. Biol. 2009, 646, 15–29. [Google Scholar] [CrossRef] [PubMed]
- Wade, K.H.; Hall, L.J. Improving causality in microbiome research: Can human genetic epidemiology help? Wellcome Open Res. 2020, 4, 199. [Google Scholar] [CrossRef]







| Infant Formula Treatment Group | |||
|---|---|---|---|
| Characteristics | CMF (n = 15) | EHF (n = 15) | p Value |
| Age in months | 0.39 ± 0.02 | 0.40 ± 0.02 | 0.90 |
| Female, n (%) | 6 (40%) | 6 (40%) | 1.00 |
| Race/ethnicity, n (%) | |||
| Black | 9 (60%) | 8 (53%) | 0.88 |
| White | 4 (27%) | 4 (27%) | |
| More than one race/ethnicity | 2 (13%) | 3 (20%) | |
| Anthropometry, Z score; 0.5 mos | |||
| Weight for age (WAZ) | −0.54 ± 0.21 | −0.37 ± 0.21 | 0.57 |
| Length for age (LAZ) | −0.73 ± 0.27 | −0.65 ± 0.67 | 0.85 |
| Weight for length (WLZ) | −0.25 ± 0.24 | −0.06 ± 0.24 | 0.59 |
| Body composition 1, 0.75 mos | |||
| Fat mass (kg) | 0.43 ± 0.08 | 0.49 ± 0.08 | 0.60 |
| Percent body fat (%) | 11.6 ± 2.0 | 12.8 ± 2.2 | 0.70 |
| Infant Formula Treatment Group | |||
|---|---|---|---|
| Characteristics | CMF (n = 15) | EHF (n = 15) | p Value |
| Anthropometry, Z scores; 4.5 months | |||
| Weight for age (WAZ) | 0.30 ± 0.23 | −0.55 ± 0.23 | 0.02 |
| Length for age (LAZ) | −0.47 ± 0.27 | −0.66 ± 0.27 | 0.61 |
| Weight for length (WLZ) | 0.88 ± 0.20 | −0.07 ± 0.20 | <0.001 |
| Weight-gain velocity (g/day), 0.5–4.5 months | 31.77 ± 1.49 | 25.49 ± 1.49 | <0.001 |
| Length-gain velocity (cm/day), 0.5–4.5 months | 0.104 ± 0.003 | 0.094 ± 0.003 | 0.18 |
| Body composition, 3.5 months 1 | |||
| Fat mass (kg) | 1.61 ± 0.08 | 1.15 ± 0.12 | 0.01 |
| Percent body fat (%) | 24.3 ± 1.5 | 19.0 ± 1.5 | 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mennella, J.A.; Li, Y.; Bittinger, K.; Friedman, E.S.; Zhao, C.; Li, H.; Wu, G.D.; Trabulsi, J.C. The Macronutrient Composition of Infant Formula Produces Differences in Gut Microbiota Maturation That Associate with Weight Gain Velocity and Weight Status. Nutrients 2022, 14, 1241. https://doi.org/10.3390/nu14061241
Mennella JA, Li Y, Bittinger K, Friedman ES, Zhao C, Li H, Wu GD, Trabulsi JC. The Macronutrient Composition of Infant Formula Produces Differences in Gut Microbiota Maturation That Associate with Weight Gain Velocity and Weight Status. Nutrients. 2022; 14(6):1241. https://doi.org/10.3390/nu14061241
Chicago/Turabian StyleMennella, Julie A., Yun Li, Kyle Bittinger, Elliot S. Friedman, Chunyu Zhao, Hongzhe Li, Gary D. Wu, and Jillian C. Trabulsi. 2022. "The Macronutrient Composition of Infant Formula Produces Differences in Gut Microbiota Maturation That Associate with Weight Gain Velocity and Weight Status" Nutrients 14, no. 6: 1241. https://doi.org/10.3390/nu14061241
APA StyleMennella, J. A., Li, Y., Bittinger, K., Friedman, E. S., Zhao, C., Li, H., Wu, G. D., & Trabulsi, J. C. (2022). The Macronutrient Composition of Infant Formula Produces Differences in Gut Microbiota Maturation That Associate with Weight Gain Velocity and Weight Status. Nutrients, 14(6), 1241. https://doi.org/10.3390/nu14061241