
Targeting the Gut Microbiota of Vertically HIV-Infected Children to Decrease Inflammation and Immunoactivation: A Pilot Clinical Trial

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Methods
2.1. Study Design, Participants, Setting and Eligibility
2.2. Nutritional Intervention
2.3. Laboratory Assessments
2.4. Gut Microbiota Analysis
2.5. Statistical Analysis
3. Results
3.1. Characteristics of the Study Population
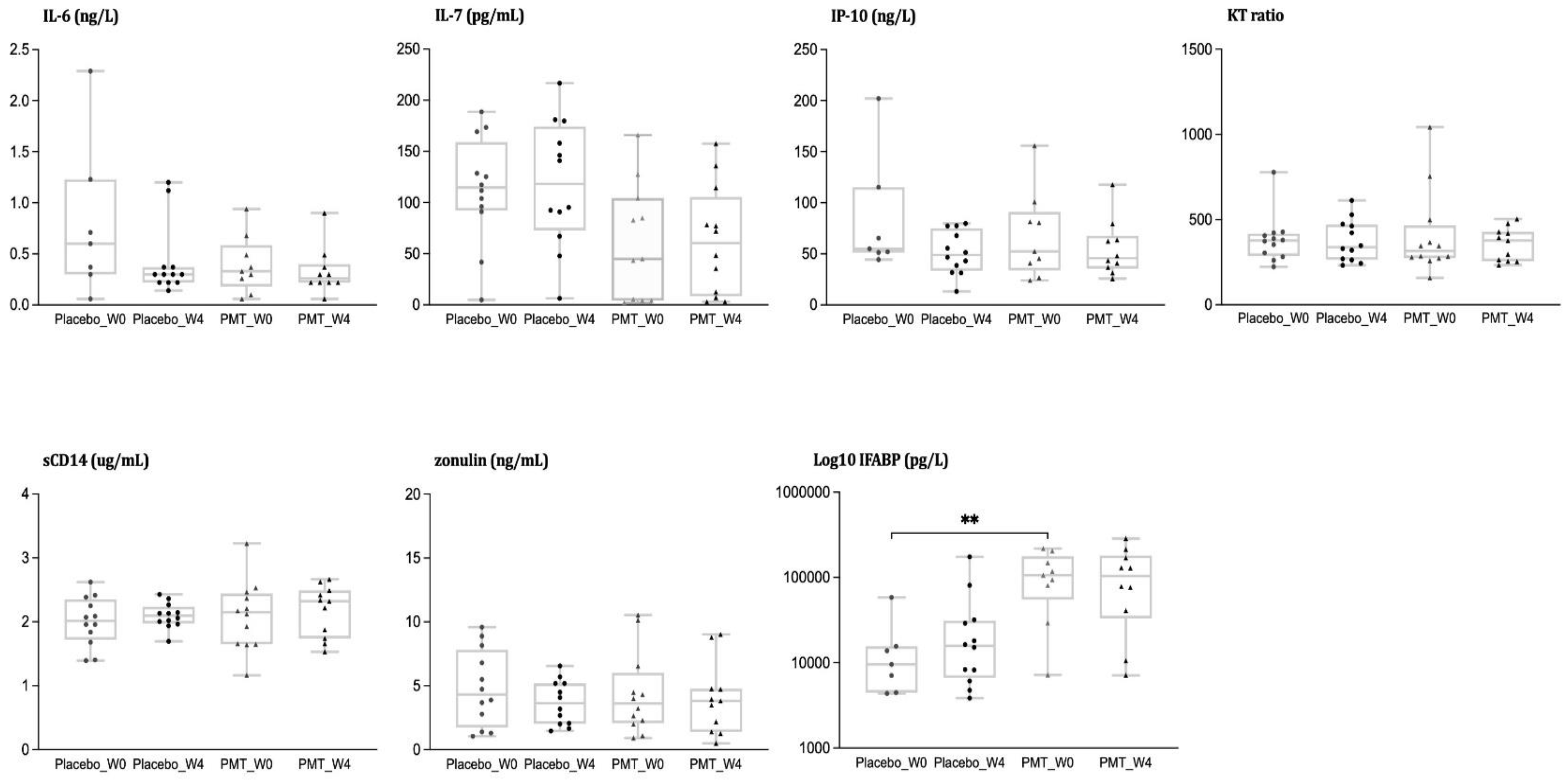
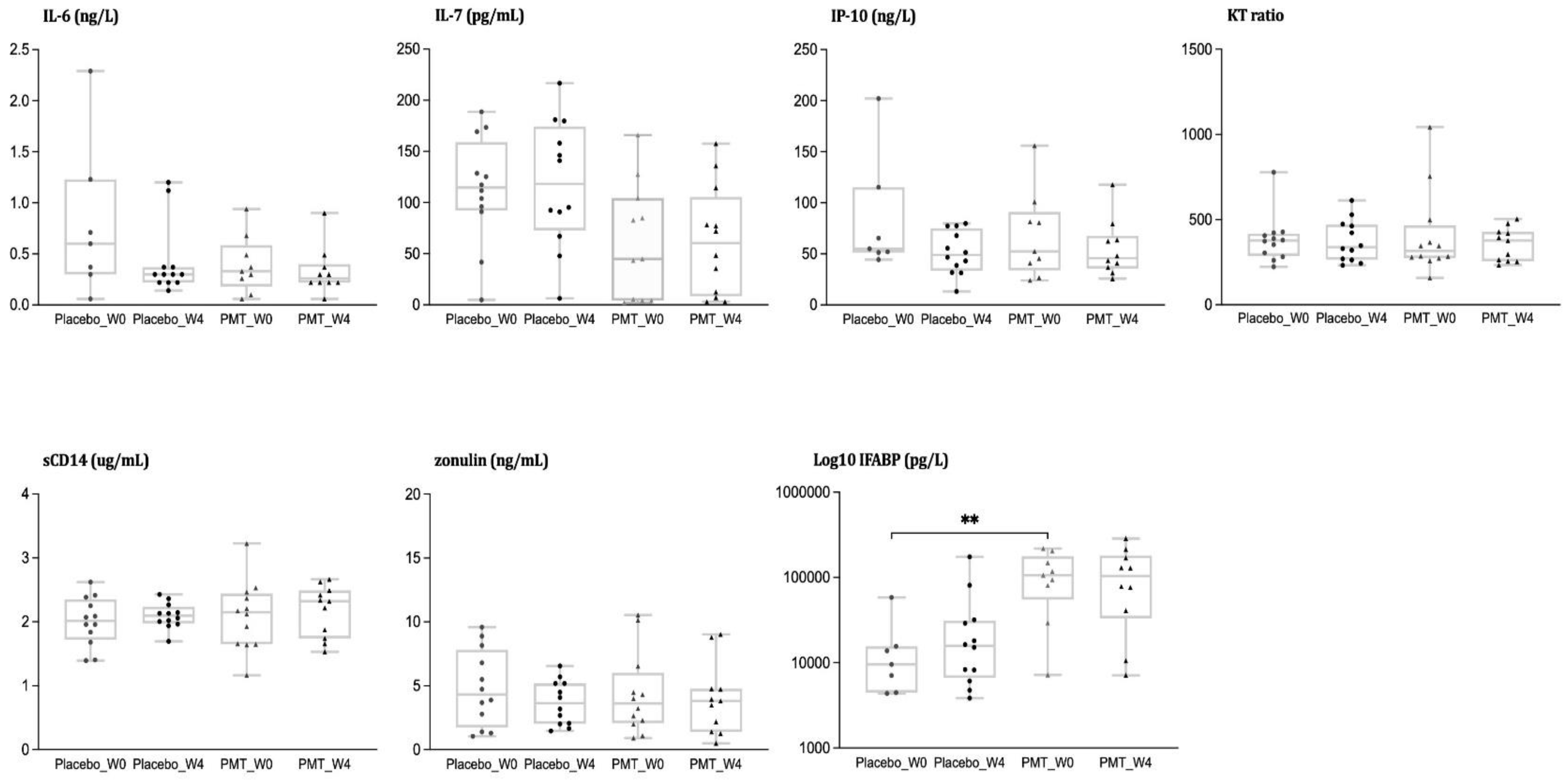
3.2. Analysis of Inflammatory Biomarkers
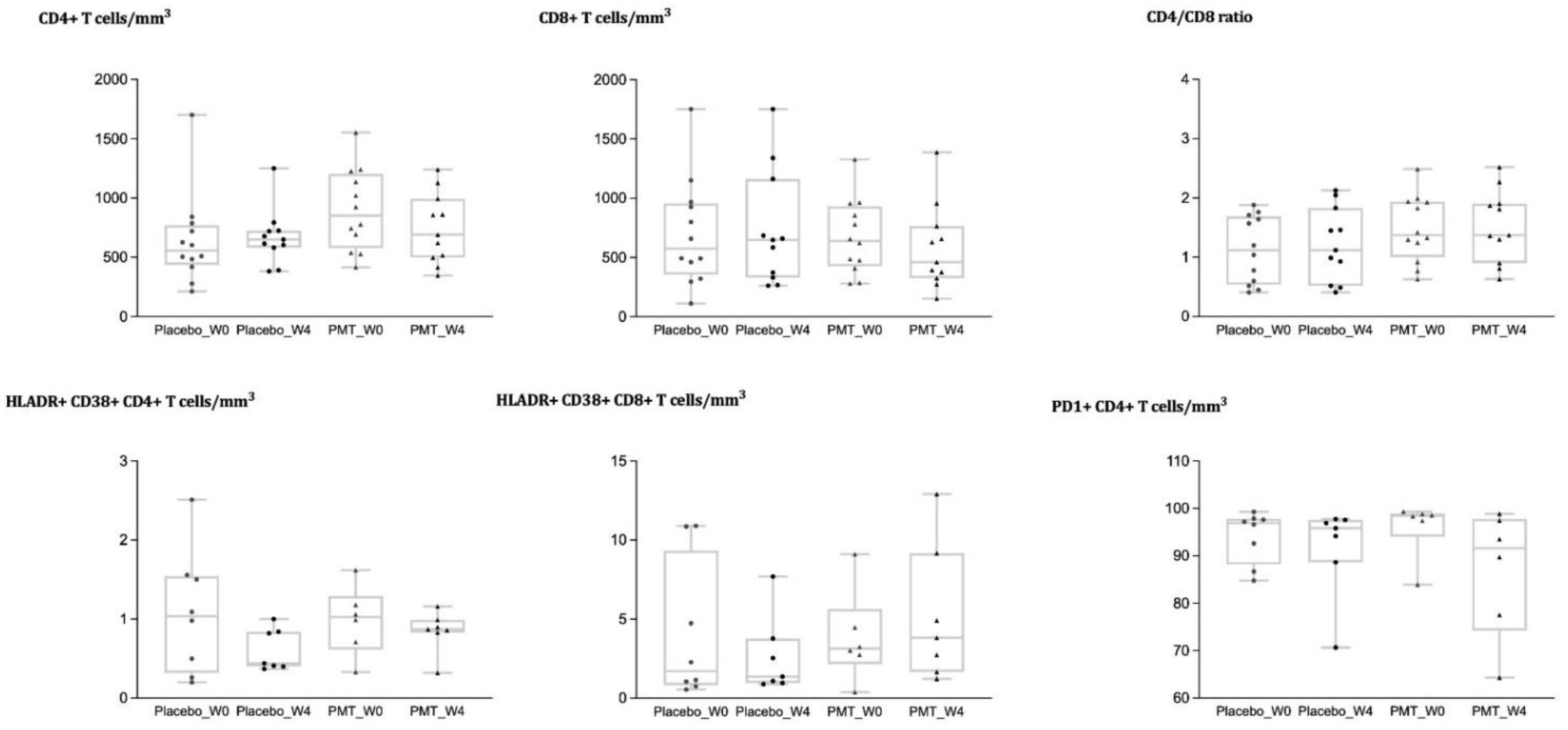
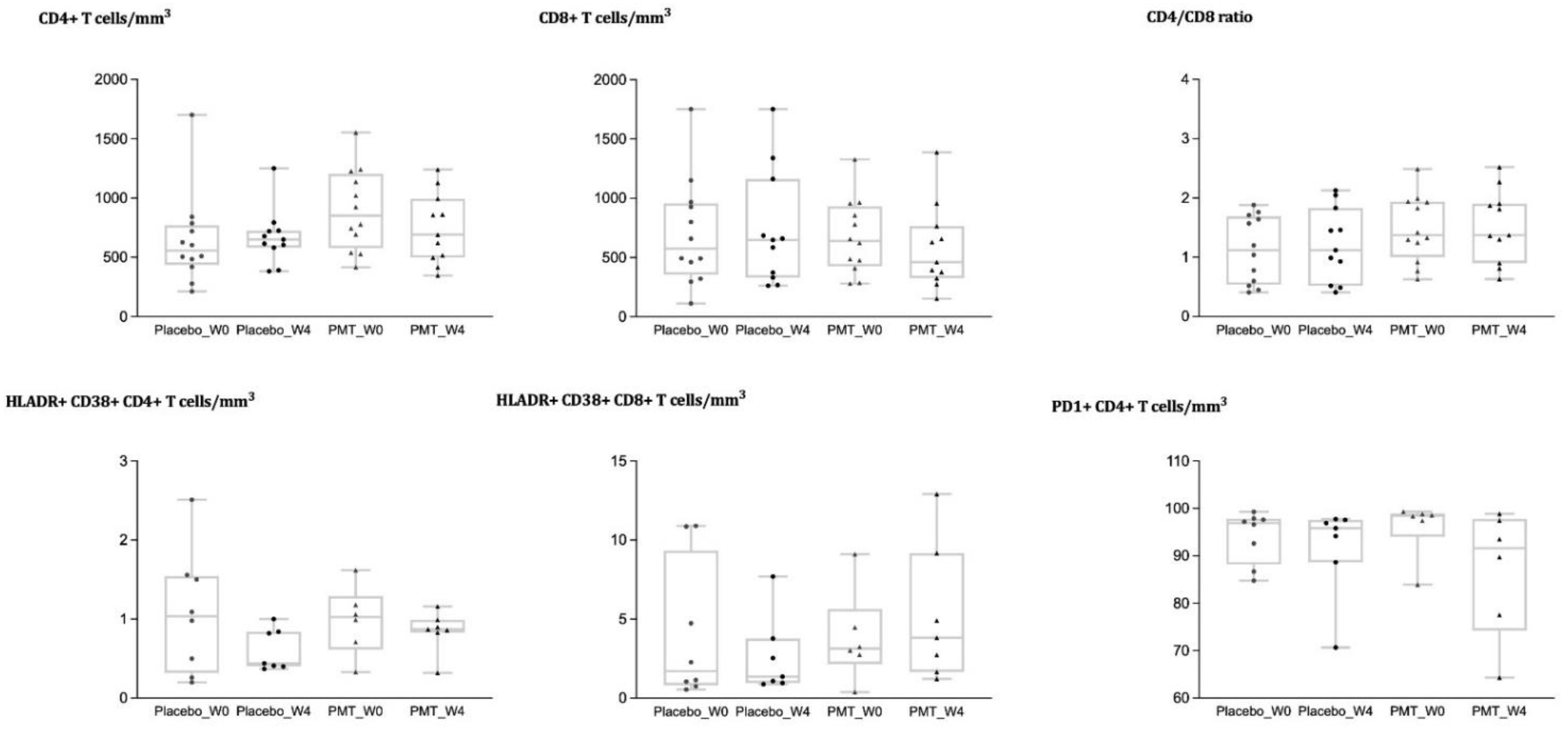
3.3. Analysis of Immune Cell Phenotypes
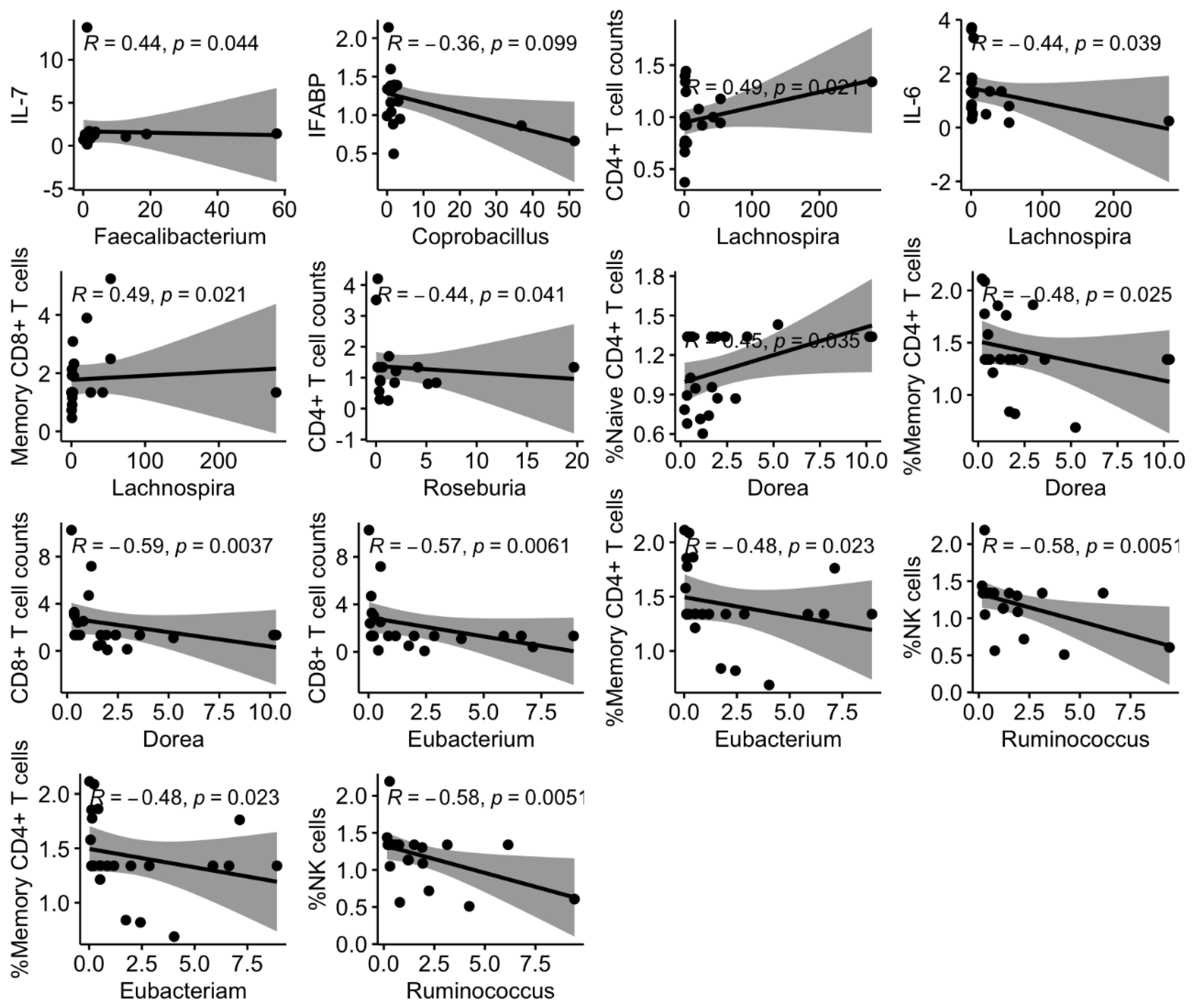
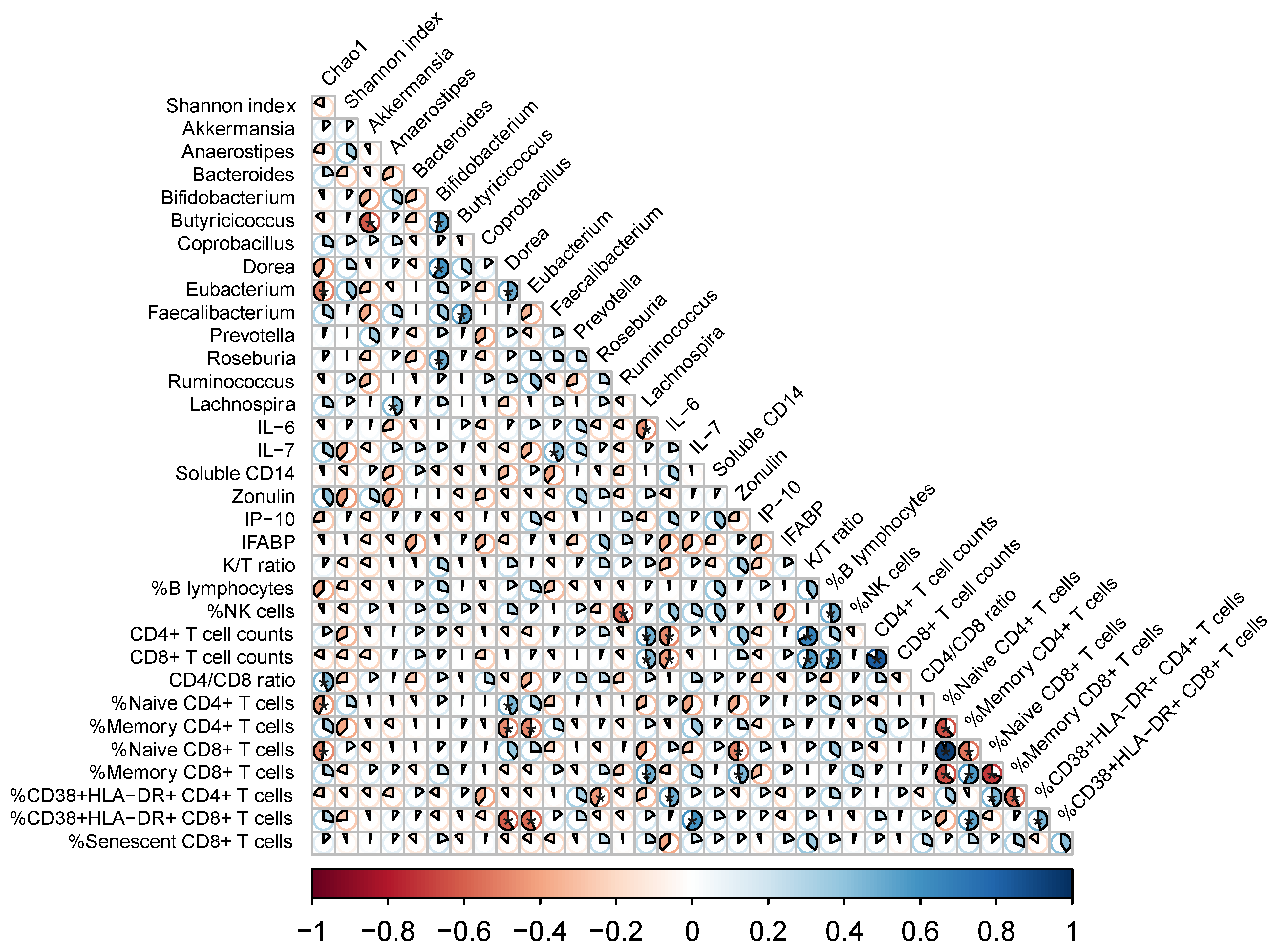
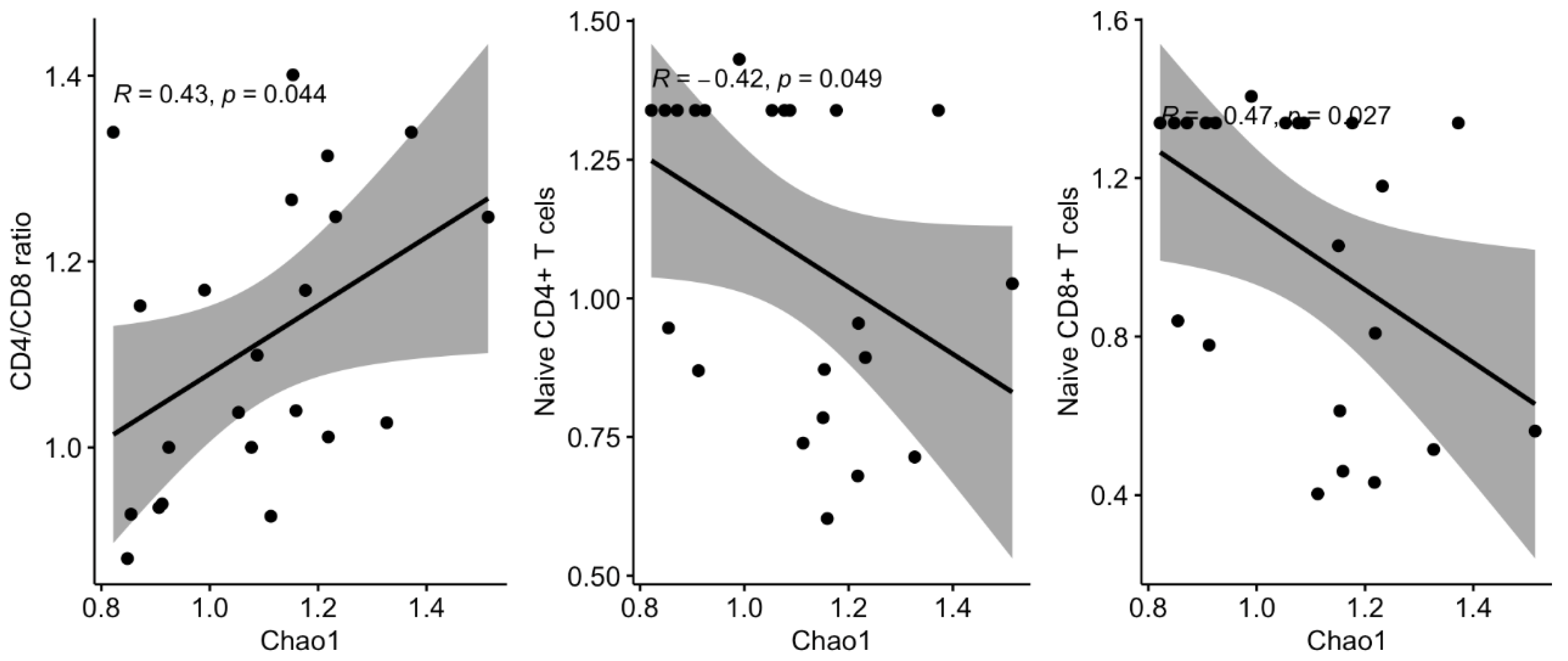
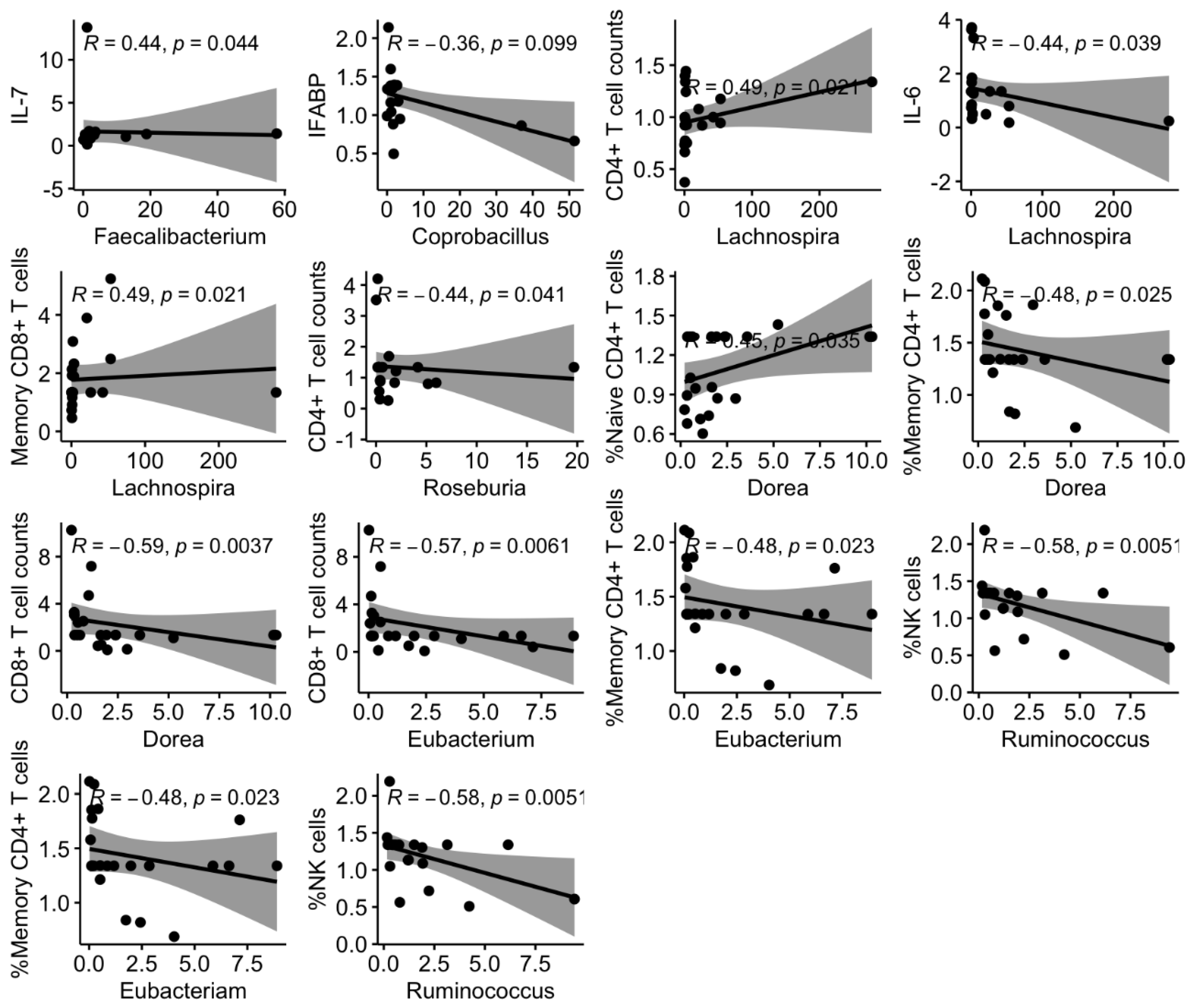
3.4. Links between Shifts in the Microbiota and Changes in Systemic Markers
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carrasco, I.; Tarancon-Diez, L.; Vázquez-Alejo, E.; Jiménez dPe Ory, S.; Sainz, T.; Apilanez, M.; Epalza, C.; Guillen, S.; Ramos, J.T.; Diez, C.; et al. Innate and adaptive abnormalities in youth with vertically acquired HIV through a multicentre cohort in Spain integrated in the Translational Research Network in Pediatric Infectious Diseases (RITIP). J. Int. AIDS Soc. 2021, 24, e25804. [Google Scholar]
- Adetokunboh, O.O.; Ndwandwe, D.; Awotiwon, A.; Uthman, O.A.; Wiysonge, C.S. Vaccination among HIV-infected, HIV-exposed uninfected and HIV-uninfected children: A systematic review and meta-analysis of evidence related to vaccine efficacy and effectiveness. Hum. Vaccines Immunother. 2019, 15, 2578–2589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niehues, T.; Horneff, G.; Knipp, S.; Adams, O.; Wahn, V. Treatment-Resistant Expansion of CD8+CD28−Cells in Pediatric HIV Infection. Pediatr. Res. 2000, 47, 418–421. [Google Scholar] [CrossRef]
- Sainz, T.; Serrano-Villar, S.; Díaz, L.; Tomé, M.I.G.; Gurbindo, M.D.; de José, M.I.; Mellado, M.J.; Ramos, J.T.; Zamora, J.; Moreno, S.; et al. The CD4/CD8 ratio as a marker T-cell activation, senescence and activation/exhaustion in treated HIV-infected children and young adults. AIDS 2013, 27, 1513–1516. [Google Scholar] [CrossRef] [PubMed]
- Vujkovic-Cvijin, I.; Dunham, R.M.; Iwai, S.; Maher, M.C.; Albright, R.G.; Broadhurst, M.J.; Hernandez, R.D.; Lederman, M.M.; Huang, Y.; Somsouk, M.; et al. Dysbiosis of the Gut Microbiota Is Associated with H.I.V. Disease Progression and Tryptophan Catabolism. Sci. Transl. Med. 2013, 5, 193ra91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dillon, S.M.; Lee, E.J.; Kotter, C.V.; Austin, G.; Dong, Z.; Hecht, D.K.; Gianella, S.; Siewe, B.; Smith, D.; Landay, A.L.; et al. An altered intestinal mucosal microbiome in HIV-1 infection is associated with mucosal and systemic immune activation and endotoxemia. Mucosal Immunol. 2014, 7, 983–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.X.; Sen, S.; Schneider, J.M.; Xiong, K.N.; Nusbacher, N.M.; Moreno-Huizar, N.; Shaffer, M.; Armstrong, A.J.S.; Severs, E.; Kuhn, K.; et al. Gut microbiota from high-risk men who have sex with men drive immune activation in gnotobiotic mice and in vitro H.I.V. infection. PLoS Pathog. 2019, 15, e1007611. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Villar, S.; Rojo, D.; Martinez-Martinez, M.; Deusch, S.; Castellanos, J.F.V.; Bargiela, R.; Sainz, T.; Vera, M.; Moreno, S.; Estrada, V.; et al. Gut Bacteria Metabolism Impacts Immune Recovery in HIV-infected Individuals. EBioMedicine 2016, 8, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Cahn, P.; Ruxrungtham, K.; Gori, A.; Vriesema, A.; Georgiou, N.A.; Garssen, J.; Clerici, M.; Lange, J.M.A.; Gazzard, B.; Diaz, R.; et al. The immunomodulatory nutritional intervention NR100157 reduced CD4+ T-cell decline and immune activation: A 1-year multicenter randomized controlled double-blind trial in HIV-infected persons not receiving antiretroviral therapy (The BITE Study). Clin. Infect. Dis. 2013, 57, 139–146. [Google Scholar] [CrossRef] [Green Version]
- Serrano-Villar, S.; Vázquez-Castellanos, J.F.; Vallejo, A.; Latorre, A.; Sainz, T.; Ferrando-Martínez, S.; Rojo, D.; Martínez-Botas, J.; Del Romero, J.; Madrid, N.; et al. The effects of prebiotics on microbial dysbiosis, butyrate production and immunity in HIV-infected subjects. Mucosal Immunol. 2017, 10, 1279–1293. [Google Scholar] [CrossRef] [Green Version]
- Vázquez-Castellanos, J.F.; Serrano-Villar, S.; Latorre, A.; Artacho, A.; Ferrus, M.L.; Madrid, N.M.; Vallejo, A.; Sainz, T.; Martinez-Botas, J.; Ferrandomartinez, S.; et al. Altered metabolism of gut microbiota contributes to chronic immune activation in HIV-infected individuals. Mucosal immunol. 2015, 8, 760–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguera-Julian, M.; Rocafort, M.; Guillén, Y.; Rivera, J.; Casadellà, M.; Nowak, P.; Hildebrand, F.; Zeller, G.; Parera, M.; Bellido, R.; et al. Gut Microbiota Linked to Sexual Preference and HIV Infection. EBioMedicine 2016, 5, 135–146. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Li, M.; Campbell, T.B.; Flores, S.C.; Linderman, D.; Gebert, M.J.; Knight, R.; Fontenot, A.P.; Palmer, B.E. Alterations in the gut microbiota associated with HIV-1 infection. Cell Host Microbe 2013, 14, 329–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deusch, S.; Serrano-Villar, S.; Rojo, D.; Martinez-Martinez, M.; Bargiela, R.; Vázquez-Castellanos, J.F.; Sainz, T.; Barbas, C.; Moya, A.; Moreno, S.; et al. Effects of HIV; antiretroviral therapy and prebiotics on the active fraction of the gut microbiota. AIDS 2018, 32, 1229–1237. [Google Scholar] [CrossRef] [PubMed]
- Sessa, L.; Reddel, S.; Manno, E.; Quagliariello, A.; Cotugno, N.; Del Chierico, F.; Amodio, D.; Capponi, C.; Leone, F.; Bernardi, S.; et al. Distinct gut microbiota profile in ART-treated perinatally HIV-infected patients associated with cardiac and inflammatory biomarkers. AIDS 2019, 33, 1001–1011. [Google Scholar] [CrossRef]
- Kaur, U.S.; Shet, A.; Rajnala, N.; Gopalan, B.P.; Moar, P.; D, H.; Singh, B.P.; Chaturvedi, R.; Tandon, R. High Abundance of genus Prevotella in the gut of perinatally HIV-infected children is associated with IP-10 levels despite therapy. Sci. Rep. 2018, 8, 17679. [Google Scholar] [CrossRef]
- Ruiz-Mateos, E.; Rubio, A.; Vallejo, A.; De la Rosa, R.; Sanchez-Quijano, A.; Lissen, E.; Leal, M. Thymic volume is associated independently with the magnitude of short- and long-term repopulation of CD4+ T cells in HIV-infected adults after highly active antiretroviral therapy (HAART). Clin. Exp. Immunol. 2004, 136, 501–506. [Google Scholar] [CrossRef]
- Mesa, M.D.; Loureiro, B.; Iglesia, I.; Fernandez Gonzalez, S.; Llurba Olive, E.; Garcia Algar, O.; Solana, M.J.; Cabero Perez, M.J.; Sainz, T.; Martinez, L.; et al. The Evolving Microbiome from Pregnancy to Early Infancy: A Comprehensive Review. Nutrients 2020, 12, 133. [Google Scholar] [CrossRef] [Green Version]
- Ishizaki, A.; Bi, X.; van Nguyen, L.; Matsuda, K.; Pham, H.V.; Phan, C.T.T.; Khu, D.T.K.; Ichimura, H. Effects of Short-Term Probiotic Ingestion on Immune Profiles and Microbial Translocation among HIV-1-Infected Vietnamese Children. Int. J. Mol. Sci. 2017, 18, 2185. [Google Scholar] [CrossRef] [Green Version]
- Sainz, T.; Gosalbes, M.J.; Talavera, A.; Jimenez-Hernandez, N.; Prieto, L.; Escosa, L.; Guillén, S.; Ramos, J.T.; Muñoz-Fernández, M.; Moya, A.; et al. Effect of a Nutritional Intervention on the Intestinal Microbiota of Vertically HIV-Infected Children: The Pediabiota Study. Nutrients 2020, 12, 2112. [Google Scholar] [CrossRef]
- Perneger, T.V. What’s wrong with Bonferroni adjustments. BMJ 1998, 316, 1236–1238. [Google Scholar] [CrossRef]
- Serrano-Villar, S.; Moreno, S.; Ferrer, M. The functional consequences of the microbiome in HIV: Insights from metabolomic studies. Curr. Opin. HIV AIDS 2018, 13, 88–94. [Google Scholar] [CrossRef]
- Tenorio, A.R.; Zheng, Y.; Bosch, R.J.; Krishnan, S.; Rodriguez, B.; Hunt, P.W.; Plants, J.; Seth, A.; Wilson, C.C.; Deeks, S.G.; et al. Soluble Markers of Inflammation & Coagulation, but not T-Cell Activation, Predict Non-AIDS- Defining Events during Suppressive Antiretroviral Therapy (ART). In Proceedings of the Conference of Retrovirus and Opportunistic Infections, Boston, MA, USA, 3–6 March 2014. [Google Scholar]
- Serrano-Villar, S.; de Lagarde, M.; Vázquez-Castellanos, J.; Vallejo, A.; Bernadino, J.I.; Madrid, N.; Matarranz, M.; Díaz-Santiago, A.; Gutierrez, C.; Cabello, A.; et al. Effects of Immunonutrition in Advanced Human Immunodeficiency Virus Disease: A Randomized Placebo-controlled Clinical Trial (Promaltia Study). Clin. Infect. Dis. 2019, 68, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Overton, E.T.; Yeh, E.; Presti, R.; Jacobson, J.; Williams, B.; Wilson, C.; Landay, A.; Brenchley, J.; Dube, M.; Fichtenbaum, C.; et al. Andrade A for the ACTG A5350 Study Team. Assessing the probiotic effect in treated HIV: Results of ACTG A5350. In Proceedings of the Conference on Retroviruses and Opportunistic Infections (CROI), Seattle, WA, USA, 4–7 March 2019. [Google Scholar]
- Reikvam, D.H.; Meyer-Myklestad, M.H.; Trøseid, M.; Stiksrud, B. Probiotics to manage inflammation in HIV infection. Curr. Opin. Infect. Dis. 2020, 33, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Villar, S.; Talavera-Rodríguez, A.; Gosalbes, M.J.; Madrid, N.; Pérez-Molina, J.A.; Elliott, R.J.; Navia, B.; Lanza, V.F.; Vallejo, A.; Osman, M.; et al. Fecal microbiota transplantation in HIV: A pilot placebo-controlled study. Nat. Commun. 2021, 12, 1139. [Google Scholar] [CrossRef] [PubMed]
- Trois, L.; Cardoso, E.M.; Miura, E. Use of probiotics in HIV-infected children: A randomized double-blind controlled study. J. Trop. Pediatr. 2008, 54, 19–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.L.; Chen, M.H.; Geng, S.T.; Yu, J.; Kuang, Y.-Q.; Luo, H.-Y.; Wang, K.-H. Effects of Probiotics on Diarrhea and CD4 Cell Count in People Living with HIV: A Systematic Review and Meta-Analysis. Front. Pharmacol. 2021, 12, 570520. [Google Scholar] [CrossRef] [PubMed]
- Athiyyah, A.F.; Brahmantya, H.; Dwiastuti, S.; Darma, A.; Puspitasari, D.; Husada, D.; Ranuh, R.; Endaryanto, A.; Surono, I.; Sudarmo, S.M. Effect of Lactobacillus plantarum IS-10506 on blood lipopolysaccharide level and immune response in HIV-infected children. Iran. J. Microbiol. 2019, 11, 137. [Google Scholar]
- Tuddenham, S.A.; Koay, W.L.A.; Zhao, N.; White, J.R.; Ghanem, K.G.; Sears, C.L.; HIV Microbiome Re-analysis Consortium; Dillon, S.; Wilson, C.; Lozupone, C.; et al. The Impact of Human Immunodeficiency Virus Infection on Gut Microbiota α-Diversity: An Individual-level Meta-analysis. Clin. Infect. Dis. 2020, 70, 615–627. [Google Scholar] [CrossRef]
- Vujkovic-Cvijin, I.; Somsouk, M. HIV and the Gut Microbiota: Composition, Consequences, and Avenues for Amelioration. Curr. HIV/AIDS Rep. 2019, 16, 204–213. [Google Scholar] [CrossRef]
- Parbie, P.K.; Mizutani, T.; Ishizaka, A.; Kawana-Tachikawa, A.; Runtuwene, L.R.; Seki, S.; Abana, C.Z.; Kushitor, D.; Bonney, E.Y.; Ofori, S.B.; et al. Dysbiotic Fecal Microbiome in HIV-1 Infected Individuals in Ghana. Front. Cell. Infect. Microbiol. 2021, 11, 379. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Villar, S.; Sainz, T.; Lee, S.A.; Hunt, P.W.; Sinclair, E.; Shacklett, B.L.; Ferre, A.L.; Hayes, T.L.; Somsouk, M.; Hsue, P.Y.; et al. HIV-Infected Individuals with Low CD4/CD8 Ratio despite Effective Antiretroviral Therapy Exhibit Altered T Cell Subsets, Heightened CD8+ T Cell Activation, and Increased Risk of Non-AIDS Morbidity and Mortality. PLoS Pathog. 2014, 10, e1004078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grund, B.; Baker, J.V.; Deeks, S.G.; Wolfson, J.; Wentworth, D.; Cozzi-Lepri, A.; Cohen, C.J.; Phillips, A.; Lundgren, J.D.; Neaton, J.D.; et al. Relevance of Interleukin-6 and D-Dimer for Serious Non-AIDS Morbidity and Death among HIV-Positive Adults on Suppressive Antiretroviral Therapy. PLoS ONE 2016, 11, e0155100. [Google Scholar] [CrossRef]
- Vacca, M.; Celano, G.; Calabrese, F.M.; Portincasa, P.; Gobbetti, M.; de Angelis, M. The controversial role of human gut lachnospiraceae. Microorganisms 2020, 8, 573. [Google Scholar] [CrossRef] [PubMed]
- Bik, E.M. Composition and function of the human-associated microbiota. Nutr. Rev. 2009, 67 (Suppl. 2), S164–S171. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.V.; Hao, L.; Offermanns, S.; Medzhitov, R. The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 2014, 111, 2247–2252. [Google Scholar] [CrossRef] [Green Version]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placebo N = 12 | Nutritional Intervention N = 12 | p | |
|---|---|---|---|
| Female (N,%) | 7 (58.3) | 8 (66.7) | 1.000 |
| Age (years), mean (SD) | 13.8 (3.6) | 10 (3.4) | 0.064 |
| Caucasian (N, %) | 6 (50) | 7 (58.3) | 1.000 |
| CD4 count (cells/mm3) | 556 (453–754) | 852 (617–1182) | 0.0496 |
| CD4/CD8 ratio | 1.1 (0.56–1.67) | 1.4 (1.09–1.94) | 0.106 |
| CD4 Nadir (cells/mm3) | 333 (169–382) | 519 (384–979) | 0.006 |
| PI based ART (N, %) | 8 (66.7) | 9 (75) | 0.136 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sainz, T.; Diaz, L.; Rojo, D.; Clemente, M.I.; Barbas, C.; Gosalbes, M.J.; Jimenez-Hernandez, N.; Escosa, L.; Guillen, S.; Ramos, J.T.; et al. Targeting the Gut Microbiota of Vertically HIV-Infected Children to Decrease Inflammation and Immunoactivation: A Pilot Clinical Trial. Nutrients 2022, 14, 992. https://doi.org/10.3390/nu14050992
Sainz T, Diaz L, Rojo D, Clemente MI, Barbas C, Gosalbes MJ, Jimenez-Hernandez N, Escosa L, Guillen S, Ramos JT, et al. Targeting the Gut Microbiota of Vertically HIV-Infected Children to Decrease Inflammation and Immunoactivation: A Pilot Clinical Trial. Nutrients. 2022; 14(5):992. https://doi.org/10.3390/nu14050992
Chicago/Turabian StyleSainz, Talía, Laura Diaz, David Rojo, María Isabel Clemente, Coral Barbas, María José Gosalbes, Nuria Jimenez-Hernandez, Luis Escosa, Sara Guillen, José Tomás Ramos, and et al. 2022. "Targeting the Gut Microbiota of Vertically HIV-Infected Children to Decrease Inflammation and Immunoactivation: A Pilot Clinical Trial" Nutrients 14, no. 5: 992. https://doi.org/10.3390/nu14050992
APA StyleSainz, T., Diaz, L., Rojo, D., Clemente, M. I., Barbas, C., Gosalbes, M. J., Jimenez-Hernandez, N., Escosa, L., Guillen, S., Ramos, J. T., Muñoz-Fernández, M. Á., Navarro, M. L., Mellado, M. J., & Serrano-Villar, S. (2022). Targeting the Gut Microbiota of Vertically HIV-Infected Children to Decrease Inflammation and Immunoactivation: A Pilot Clinical Trial. Nutrients, 14(5), 992. https://doi.org/10.3390/nu14050992