
EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and TNBC Cells Secretome Collection
2.3. Total RNA Isolation, cDNA Synthesis, and Real-Time Quantitative PCR
2.4. Total RNA Library Preparation and Sequencing
2.5. Reads Alignment and Differential Expression Analysis
2.6. Gene Ontology Pathway Enrichment Analysis
2.7. Human Cancer Inflammation and Immunity Crosstalk PCR Array
2.8. RNA Interference
2.9. Western Blot
2.10. Multiplex Cytokine ELISA Array
2.11. Immunoprecipitation Procedures
2.12. Chemotactic Cell Migration Assay
2.13. Statistical Data Analysis
3. Results
3.1. Transcriptomic Analysis of Human Adipocyte-Derived Mesenchymal Stem/Stromal Cells Response to Variations of the TNBC Secretome
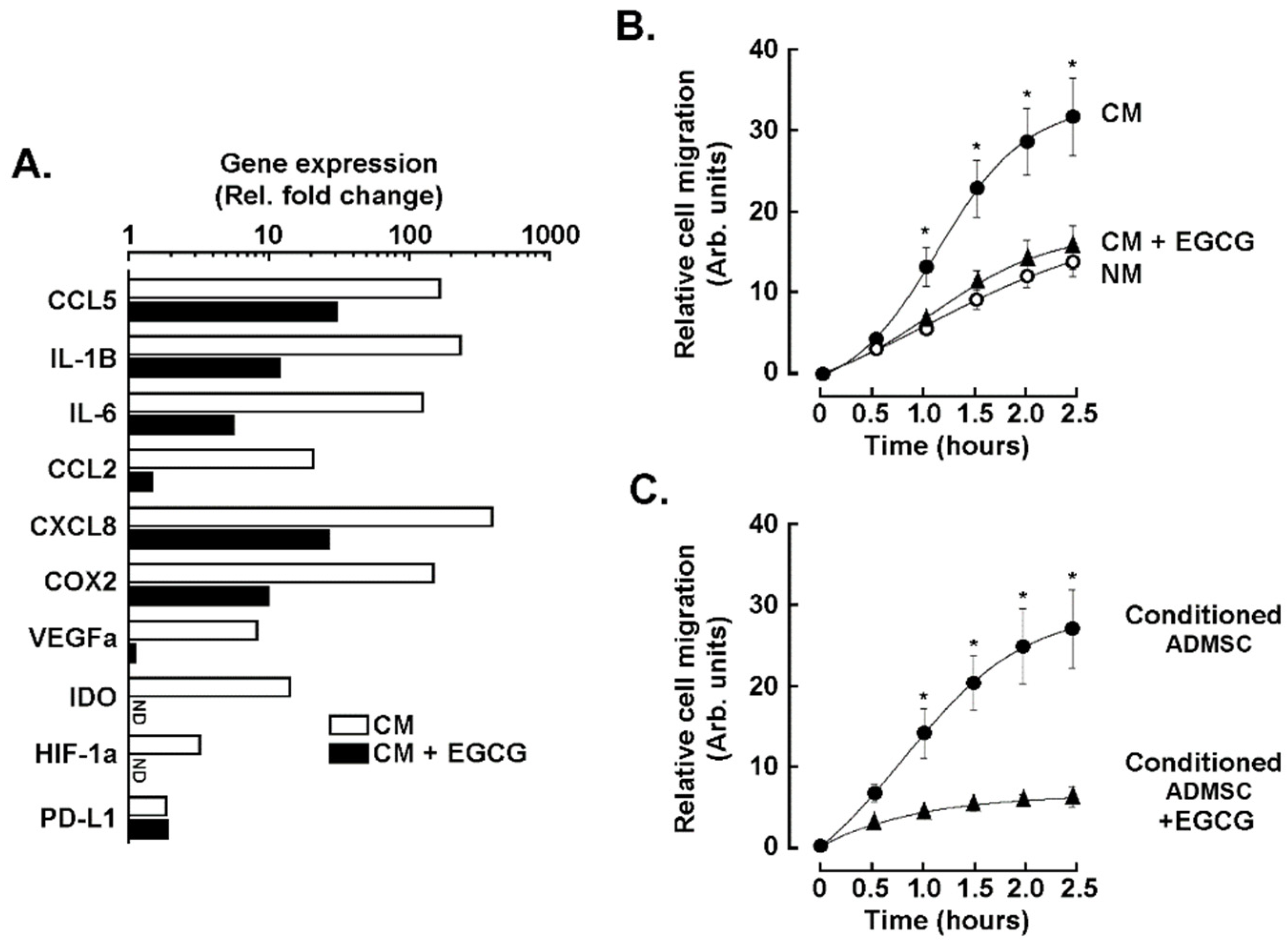
3.2. EGCG Inhibits the Expression of Biomarkers Associated with the CAA Phenotype, Epithelial-to-Mesenchymal Transition, and Inflammatory Signaling Pathways Induced by the TNBC Cell Secretome
3.3. The Epithelial-to-Mesenchymal Transition (EMT) Contributes to the CAA-Induced Phenotype in ADMSC
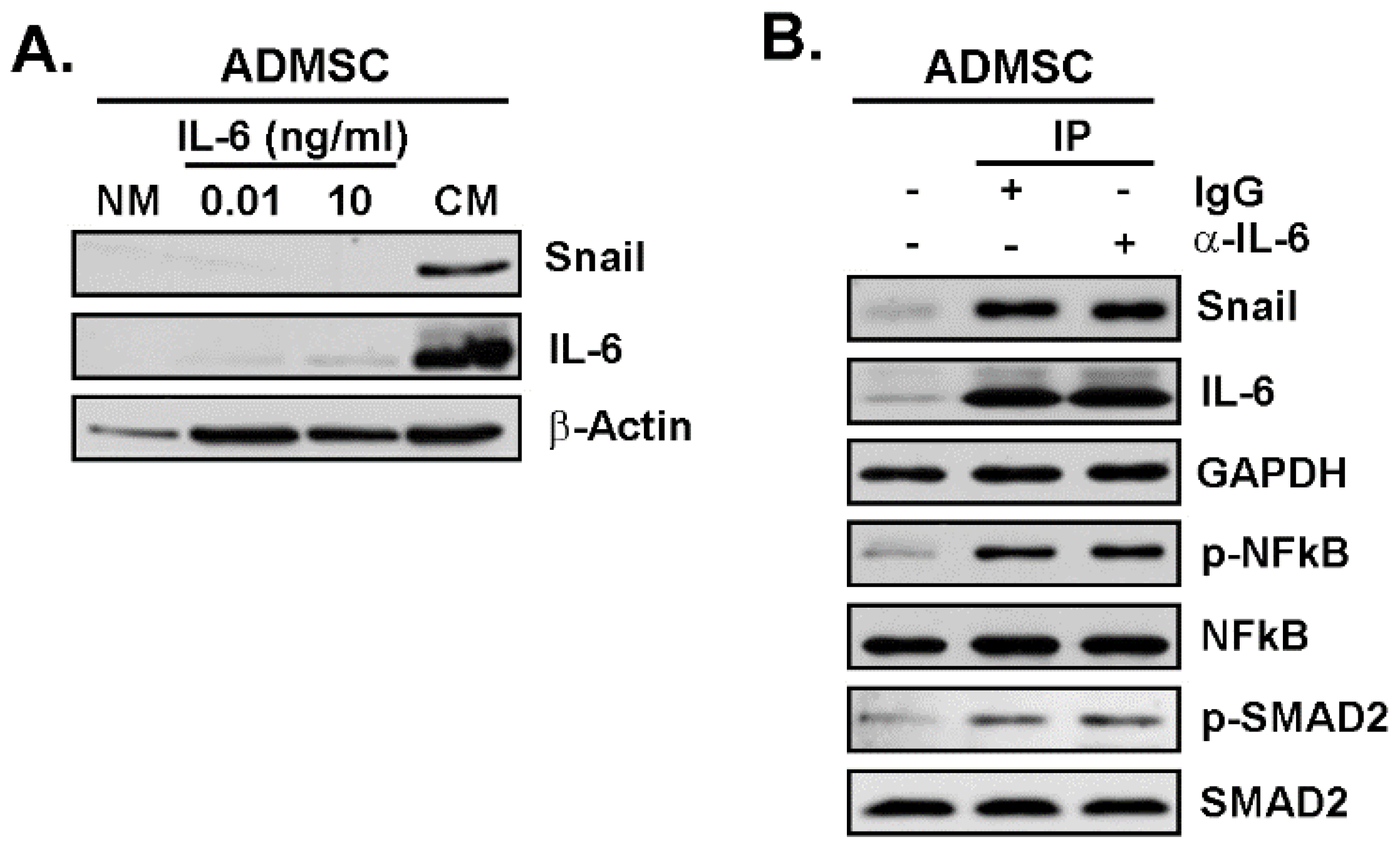
3.4. Snail as a Crucial Intermediate in the Upregulation of CAA Genes in Response to TNBC Cells Secretome
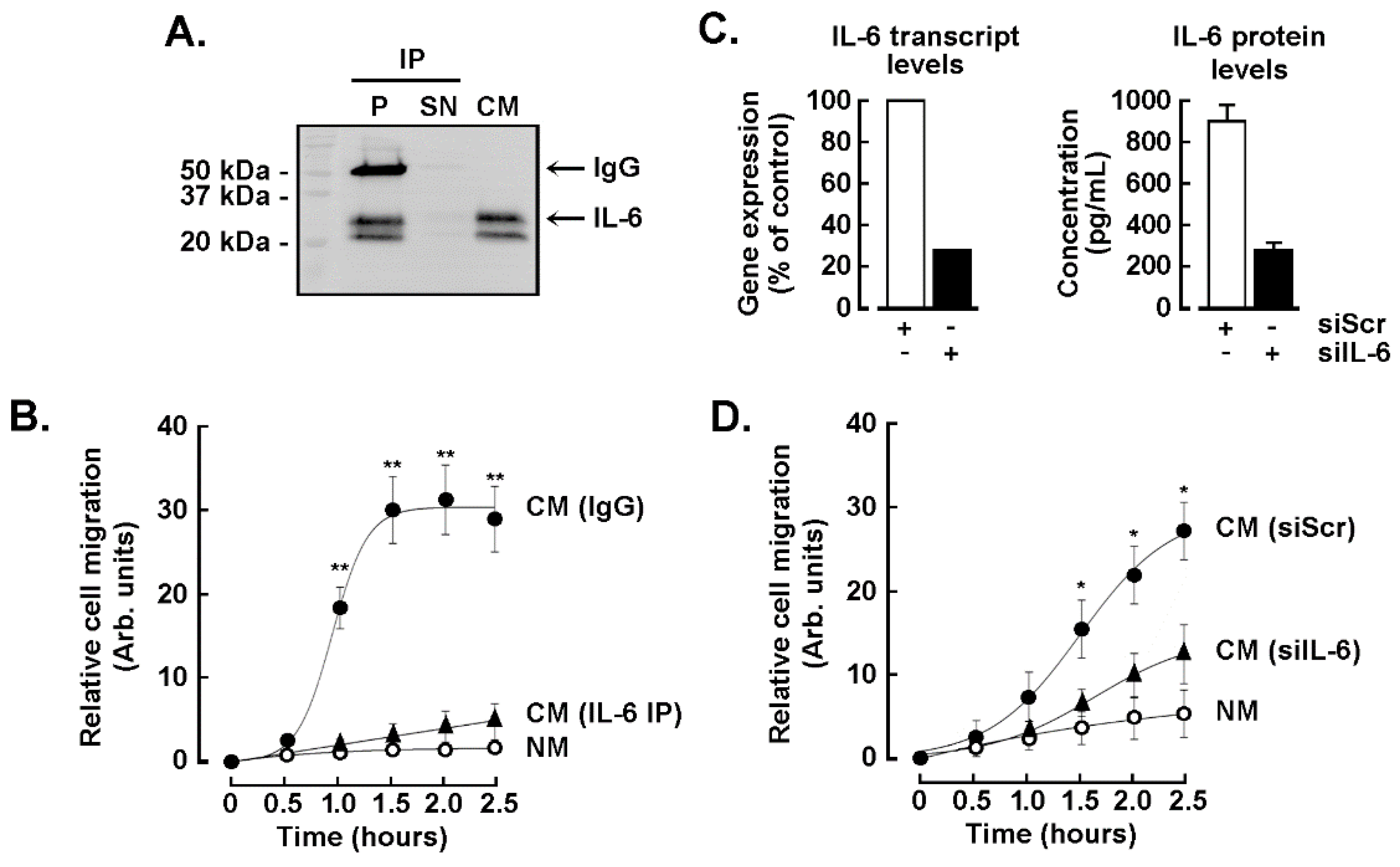
3.5. The TNBC-Derived MDA-MB-231 Secrete High Levels of IL-6
3.6. The IL-6 Secreted by the TNBC-Derived MDA-MB-231 Is Required for the Chemotactic Response of the ADMSC but Does Not Trigger the CAA Phenotype
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Zhao, C.; Wu, M.; Zeng, N.; Xiong, M.; Hu, W.; Lv, W.; Yi, Y.; Zhang, Q.; Wu, Y. Cancer-associated adipocytes: Emerging supporters in breast cancer. J. Exp. Clin. Cancer Res. 2020, 39, 156. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Hong, B.S.; Ryu, H.S.; Lee, H.B.; Lee, M.; Park, I.A.; Kim, J.; Han, W.; Noh, D.Y.; Moon, H.G. Transition into inflammatory cancer-associated adipocytes in breast cancer microenvironment requires microRNA regulatory mechanism. PLoS ONE 2017, 12, e0174126. [Google Scholar] [CrossRef] [Green Version]
- Pu, X.; Chen, D. Targeting adipokines in obesity-related tumors. Front. Oncol. 2021, 11, 685923. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Liang, Y.; Xing, C.; Wang, H.; Hu, P.; Li, J.; Huang, H.; Wang, W.; Jiang, C. Cancer-associated adipocytes exhibit distinct phenotypes and facilitate tumor progression in pancreatic cancer. Oncol. Rep. 2019, 42, 2537–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, Q.; Li, B.; Li, Z.; Li, J.; Sun, S.; Sun, S. Cancer-associated adipocytes: Key players in breast cancer progression. J. Hematol. Oncol. 2019, 12, 95. [Google Scholar] [CrossRef]
- Dirat, B.; Bochet, L.; Dabek, M.; Daviaud, D.; Dauvillier, S.; Majed, B.; Wang, Y.Y.; Meulle, A.; Salles, B.; Le Gonidec, S.; et al. Cancer-associated adipocytes exhibit an activated phenotype and contribute to breast cancer invasion. Cancer Res. 2011, 71, 2455–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybinska, I.; Mangano, N.; Tagliabue, E.; Triulzi, T. Cancer-associated adipocytes in breast cancer: Causes and consequences. Int. J. Mol. Sci. 2021, 22, 3775. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Nájera, L.E.; Chanona-Pérez, J.J.; Valdivia-Flores, A.; Marrero-Rodríguez, D.; Salcedo-Vargas, M.; García-Ruiz, D.I.; Castro-Reyes, M.A. Morphometric study of adipocytes on breast cancer by means of photonic microscopy and image analysis. Microsc. Res. Tech. 2018, 81, 240–249. [Google Scholar] [CrossRef]
- Attané, C.; Muller, C. Drilling for oil: Tumor-surrounding adipocytes fueling cancer. Trends Cancer 2020, 6, 593–604. [Google Scholar] [CrossRef]
- Wang, Z.H.; Peng, W.B.; Zhang, P.; Yang, X.P.; Zhou, Q. Lactate in the tumour microenvironment: From immune modulation to therapy. EBioMedicine 2021, 73, 103627. [Google Scholar] [CrossRef]
- Brown, T.P.; Ganapathy, V. Lactate/GPR81 signaling and proton motive force in cancer: Role in angiogenesis, immune escape, nutrition, and Warburg phenomenon. Pharmacol. Ther. 2020, 206, 107451. [Google Scholar] [CrossRef] [PubMed]
- Masoud, G.N.; Li, W. HIF-1α pathway: Role, regulation and intervention for cancer therapy. Acta Pharm. Sin. B 2015, 5, 378–389. [Google Scholar] [CrossRef] [Green Version]
- Andarawewa, K.L.; Motrescu, E.R.; Chenard, M.P.; Gansmuller, A.; Stoll, I.; Tomasetto, C.; Rio, M.C. Stromelysin-3 is a potent negative regulator of adipogenesis participating to cancer cell-adipocyte interaction/crosstalk at the tumor invasive front. Cancer Res. 2005, 65, 10862–10871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochet, L.; Lehuédé, C.; Dauvillier, S.; Wang, Y.Y.; Dirat, B.; Laurent, V.; Dray, C.; Guiet, R.; Maridonneau-Parini, I.; Le Gonidec, S.; et al. Adipocyte-derived fibroblasts promote tumor progression and contribute to the desmoplastic reaction in breast cancer. Cancer Res. 2013, 3, 5657–5668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Protani, M.; Coory, M.; Martin, J.H. Effect of obesity on survival of women with breast cancer: Systematic review and meta-analysis. Breast Cancer Res. Treat. 2010, 123, 627–635. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Ramos-Nino, M.E. The role of chronic inflammation in obesity-associated cancers. ISRN Oncol. 2013, 2013, 697521. [Google Scholar] [CrossRef] [Green Version]
- Lengyel, E.; Makowski, L.; DiGiovanni, J.; Kolonin, M.G. Cancer as a matter of fat: The crosstalk between adipose tissue and tumors. Trends Cancer 2018, 4, 374–384. [Google Scholar] [CrossRef]
- Pascut, D.; Pratama, M.Y.; Vo, N.V.T.; Masadah, R.; Tiribelli, C. The crosstalk between tumor cells and the microenvironment in hepatocellular carcinoma: The role of exosomal microRNAs and their clinical implications. Cancers 2020, 12, 823. [Google Scholar] [CrossRef] [Green Version]
- World Cancer Research Fund/American Institute for Cancer Research. Diet, Nutrition, Physical Activity and Cancer: A Global Perspective. Continuous Update Project Expert Report 2018. Available online: https://www.wcrf.org/dietandcancer (accessed on 13 January 2022).
- Andersen, C.; Rayalam, S.; Della-Fera, M.A.; Baile, C.A. Phytochemicals and adipogenesis. Biofactors 2010, 36, 415–422. [Google Scholar] [CrossRef]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells—Current trends and future prospective. Biosci. Rep. 2015, 35, e00191. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Suarez, N.; Rodriguez Torres, S.; Ouanouki, A.; El Cheikh-Hussein, L.; Annabi, B. EGCG inhibits adipose-derived mesenchymal stem cells differentiation into adipocytes and prevents a STAT3-mediated paracrine oncogenic control of triple-negative breast cancer cell invasive phenotype. Molecules 2021, 26, 1506. [Google Scholar] [CrossRef] [PubMed]
- D′Esposito, V.; Liguoro, D.; Ambrosio, M.R.; Collina, F.; Cantile, M.; Spinelli, R.; Raciti, G.A.; Miele, C.; Valentino, R.; Campiglia, P.; et al. Adipose microenvironment promotes triple negative breast cancer cell invasiveness and dissemination by producing CCL5. Oncotarget 2016, 7, 24495–24509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamat, P.; Schweizer, R.; Kaenel, P.; Salemi, S.; Calcagni, M.; Giovanoli, P.; Gorantla, V.S.; Eberli, D.; Andres, A.C.; Plock, J.A. Human adipose-derived mesenchymal stromal cells may promote breast cancer progression and metastatic spread. Plast Reconstr. Surg. 2015, 136, 76–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rybinska, I.; Agresti, R.; Trapani, A.; Tagliabue, E.; Triulzi, T. Adipocytes in breast cancer, the thick and the thin. Cells 2020, 9, 560. [Google Scholar] [CrossRef] [Green Version]
- Robert, A.W.; Marcon, B.H.; Dallagiovanna, B.; Shigunov, P. Adipogenesis, osteogenesis, and chondrogenesis of human mesenchymal stem/stromal cells: A comparative transcriptome approach. Front. Cell Dev. Biol. 2020, 8, 561. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef]
- The R Core Team. R: A Language and Environment for Statistical Computing; Version 3.4.1; R Foundation for Statistical Computing: Viena, Austria, 2017. [Google Scholar]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [Green Version]
- Ulgen, E.; Ozisik, O.; Sezerman, O.U. pathfindR: An R Package for Comprehensive Identification of Enriched Pathways in Omics Data through Active Subnetworks. Front Genet. 2019, 10, 858. [Google Scholar] [CrossRef] [PubMed]
- Gu, Z.; Eils, R.; Schlesner, M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics 2016, 32, 2847–2849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catalán, V.; Gómez-Ambrosi, J.; Rodríguez, A.; Frühbeck, G. Adipose tissue immunity and cancer. Front. Physiol. 2013, 4, 275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cozzo, A.J.; Fuller, A.M.; Makowski, L. Contribution of adipose tissue to development of cancer. Compr. Physiol. 2017, 8, 237–282. [Google Scholar]
- Labani-Motlagh, A.; Ashja-Mahdavi, M.; Loskog, A. The tumor microenvironment: A milieu hindering and obstructing antitumor immune responses. Front. Immunol. 2020, 11, 940. [Google Scholar] [CrossRef]
- Picon-Ruiz, M.; Pan, C.; Drews-Elger, K.; Jang, K.; Besser, A.H.; Zhao, D.; Morata-Tarifa, C.; Kim, M.; Ince, T.A.; Azzam, D.J.; et al. Interactions between adipocytes and breast cancer cells stimulate cytokine production and drive Src/Sox2/miR-302b-mediated malignant progression. Cancer Res. 2016, 76, 491–504. [Google Scholar] [CrossRef] [Green Version]
- Picon-Ruiz, M.; Morata-Tarifa, C.; Valle-Goffin, J.J.; Friedman, E.R.; Slingerland, J.M. Obesity and adverse breast cancer risk and outcome: Mechanistic insights and strategies for intervention. CA Cancer J. Clin. 2017, 67, 378–397. [Google Scholar] [CrossRef]
- Plava, J.; Cihova, M.; Burikova, M.; Bohac, M.; Adamkov, M.; Drahosova, S.; Rusnakova, D.; Pindak, D.; Karaba, M.; Simo, J.; et al. Permanent pro-tumorigenic shift in adipose tissue-derived mesenchymal stromal cells induced by breast malignancy. Cells 2020, 9, 480. [Google Scholar] [CrossRef] [Green Version]
- Trayhurn, P. Hypoxia and adipose tissue function and dysfunction in obesity. Physiol. Rev. 2013, 93, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Trabold, O.; Wagner, S.; Wicke, C.; Scheuenstuhl, H.; Hussain, M.Z.; Rosen, N.; Seremetiev, A.; Becker, H.D.; Hunt, T.K. Lactate and oxygen constitute a fundamental regulatory mechanism in wound healing. Wound Repair Regen. 2003, 11, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Tornatore, L.; Thotakura, A.K.; Bennett, J.; Moretti, M.; Franzoso, G. The nuclear factor kappa B signaling pathway: Integrating metabolism with inflammation. Trends Cell Biol. 2012, 22, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Asadipooya, K.; Uy, E.M. Advanced glycation end products (AGEs), receptor for AGEs, diabetes, and bone: Review of the literature. J. Endocr. Soc. 2019, 3, 1799–1818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, D. Role of advanced glycation end products (AGEs) and its receptor (RAGE)-mediated diabetic vascular complications. Integr. Food Nutr. Metab. 2019, 6, 1–6. [Google Scholar] [CrossRef]
- Trayhurn, P.; Wood, I.S. Adipokines: Inflammation and the pleiotropic role of white adipose tissue. Br. J. Nutr. 2004, 92, 347–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trayhurn, P.; Wang, B.; Wood, I.S. Hypoxia in adipose tissue: A basis for the dysregulation of tissue function in obesity? Br. J. Nutr. 2008, 100, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.Y.; Liao, Y.W.; Lu, M.Y.; Yang, C.M.; Hsieh, P.L.; Yu, C.C. Positive feedback loop of SNAIL-IL-6 mediates myofibroblastic differentiation activity in precancerous oral submucous fibrosis. Cancers 2020, 12, 1611. [Google Scholar] [CrossRef]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef]
- Djediai, S.; Gonzalez Suarez, N.; El Cheikh-Hussein, L.; Rodriguez Torres, S.; Gresseau, L.; Dhayne, S.; Joly-Lopez, Z.; Annabi, B. MT1-MMP cooperates with TGF-β receptor-mediated signaling to trigger SNAIL and induce epithelial-to-mesenchymal-like transition in U87 glioblastoma cells. Int. J. Mol. Sci. 2021, 22, 13006. [Google Scholar] [CrossRef]
- Sicard, A.A.; Dao, T.; Suarez, N.G.; Annabi, B. Diet-derived gallated catechins prevent TGF-β-mediated epithelial-mesenchymal transition, cell migration and vasculogenic mimicry in chemosensitive ES-2 ovarian cancer cells. Nutr. Cancer 2021, 73, 169–180. [Google Scholar] [CrossRef]
- Chokor, R.; Lamy, S.; Annabi, B. Transcriptional targeting of sphingosine-1-phosphate receptor S1P2 by epigallocatechin-3-gallate prevents sphingosine-1-phosphate-mediated signaling in macrophage-differentiated HL-60 promyelomonocytic leukemia cells. Onco Targets Ther. 2014, 7, 667–677. [Google Scholar] [PubMed] [Green Version]
- Zgheib, A.; Lamy, S.; Annabi, B. Epigallocatechin gallate targeting of membrane type 1 matrix metalloproteinase-mediated Src and Janus kinase/signal transducers and activators of transcription 3 signaling inhibits transcription of colony-stimulating factors 2 and 3 in mesenchymal stromal cells. J. Biol. Chem. 2013, 288, 13378–13386. [Google Scholar] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ENSEMBL | Gene | Fold Change Vehicle vs. Control | Fold Change EGCG vs. Control | Gene Description | Enriched Terms by KEGG Analysis |
|---|---|---|---|---|---|
| ENSG00000122641 | INHBA | 5.36 | −2.51 | Follicle-Stimulating Hormone-Releasing Protein/secreted |
|
| ENSG00000152952 | PLOD2 | 4.51 | −4.00 | 2 procollagen-lysine/cisternae of the RER. |
|
| ENSG00000170961 | HAS2 | 4.25 | −2.99 | Hyaluronan synthase 2 Polysaccharide/extracellular matrix |
|
| ENSG00000105835 | NAMPT | 3.94 | −1.54 | Nicotinamide phosphoribosyltransferase; enzime |
|
| ENSG00000104321 | TRPA1 | 3.87 | −4.76 | Transient receptor potential cation channel, subfamily a/transmembrane proteins |
|
| ENSG00000112972 | HMGCS1 | 3.73 | −2.37 | 3-α-hydroxy-3-methylglutaryl-coa synthase 1/ |
|
| ENSG00000196611 | MMP1 | 3.68 | −1.80 | Matrix metalloproteinase 1/interstitial collagenase |
|
| ENSG00000067064 | IDI1 | 3.56 | −2.33 | Isopentenyl-diphosphate delta isomerase 1/peroxisomally-localized enzyme |
|
| ENSG00000041353 | RAB27B | 3.40 | −6.85 | Member RAS oncogene family/membrane-bound proteins involved in vesicular fusion and trafficking |
|
| ENSG00000104415 | CCN4 | 3.39 | −2.87 | Wnt1-inducible signaling pathway protein 1/ |
|
| ENSG00000157214 | STEAP2 | 3.39 | −2.92 | Six-transmembrane epithelial antigen of prostate/metalloreductase localized in Golgi complex, plasma membrane, and in the cytosol. |
|
| ENSG00000164211 | STARD4 | 3.14 | −2.38 | Start domain-containing protein 4 |
|
| ENSG00000119927 | GPAM | 3.13 | −4.02 | Glycerol-3-phosphate acyltransferase/Mitochondrial |
|
| ENSG00000120437 | ACAT2 | 3.11 | −2.60 | Acetyl-coa acetyltransferase 2/cytosolic |
|
| ENSG00000171208 | NETO2 | 3.09 | −1.57 | Neuropilin- and tolloid-like 2/transmembrane protein |
|
| ENSG00000153823 | PID1 | 3.00 | −1.64 | Phosphotyrosine interaction domain-containing 1 |
|
| ENSG00000153162 | BMP6 | 2.89 | −2.05 | Bone morphogenetic protein 6 |
|
| ENSG00000164647 | STEAP1 | 2.80 | −2.36 | Six-transmembrane epithelial antigen of prostate 1/cell surface antigen significantly expressed at cell-cell junctions. |
|
| ENSG00000135048 | CEMIP2 | 2.76 | −2.24 | Cell migration inducing hyaluronidase 2/transmembrane protein |
|
| ENSG00000113161 | HMGCR | 2.70 | −2.13 | 3-@hydroxy-3-methylglutaryl-coa reductase/rate-limiting enzyme for cholesterol synthesis |
|
| ENSG00000114251 | WNT5A | 2.70 | −2.10 | Wingless-type mmtv integration site family/secreted signaling proteins |
|
| ENSG00000182752 | PAPPA | 2.67 | −1.72 | Pregnancy-associated plasma protein/secreted metalloproteinase |
|
| ENSG00000125845 | BMP2 | 2.61 | −2.26 | Bone morphogenetic protein 2/regulatory element: cis-acting enhancer |
|
| ENSG00000144810 | COL8A1 | 2.60 | −2.67 | Collagen, type VIII |
|
| ENSG00000140416 | TPM1 | 2.40 | −3.17 | Tropomyosin 1/actin-binding proteins involved in the contractile system of muscles |
|
| ENSG00000068366 | ACSL4 | 2.34 | −5.44 | acyl-coa synthetase long chain family |
|
| ENSG00000140285 | FGF7 | 2.30 | −5.16 | Fibroblast growth factor 7 |
|
| ENSG00000072274 | TFRC | 2.07 | −2.52 | Transferrin receptor/ cell surface receptor |
|
| ENSG00000100644 | HIF1A | 2.05 | −5.07 | Hypoxia-inducible factor 1, alpha subunit |
|
| ENSG00000148848 | ADAM12 | 2.02 | −3.54 | A disintegrin and metalloproteinase domain 12 |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonzalez Suarez, N.; Fernandez-Marrero, Y.; Torabidastgerdooei, S.; Annabi, B. EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome. Nutrients 2022, 14, 1099. https://doi.org/10.3390/nu14051099
Gonzalez Suarez N, Fernandez-Marrero Y, Torabidastgerdooei S, Annabi B. EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome. Nutrients. 2022; 14(5):1099. https://doi.org/10.3390/nu14051099
Chicago/Turabian StyleGonzalez Suarez, Narjara, Yuniel Fernandez-Marrero, Sima Torabidastgerdooei, and Borhane Annabi. 2022. "EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome" Nutrients 14, no. 5: 1099. https://doi.org/10.3390/nu14051099
APA StyleGonzalez Suarez, N., Fernandez-Marrero, Y., Torabidastgerdooei, S., & Annabi, B. (2022). EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome. Nutrients, 14(5), 1099. https://doi.org/10.3390/nu14051099