
Gut Microbiota Modulation of Moderate Undernutrition in Infants through Gummy Lactobacillus plantarum Dad-13 Consumption: A Randomized Double-Blind Controlled Trial

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Sample Size Calculation
2.3. Research Subjects and Randomization
2.4. Research Product
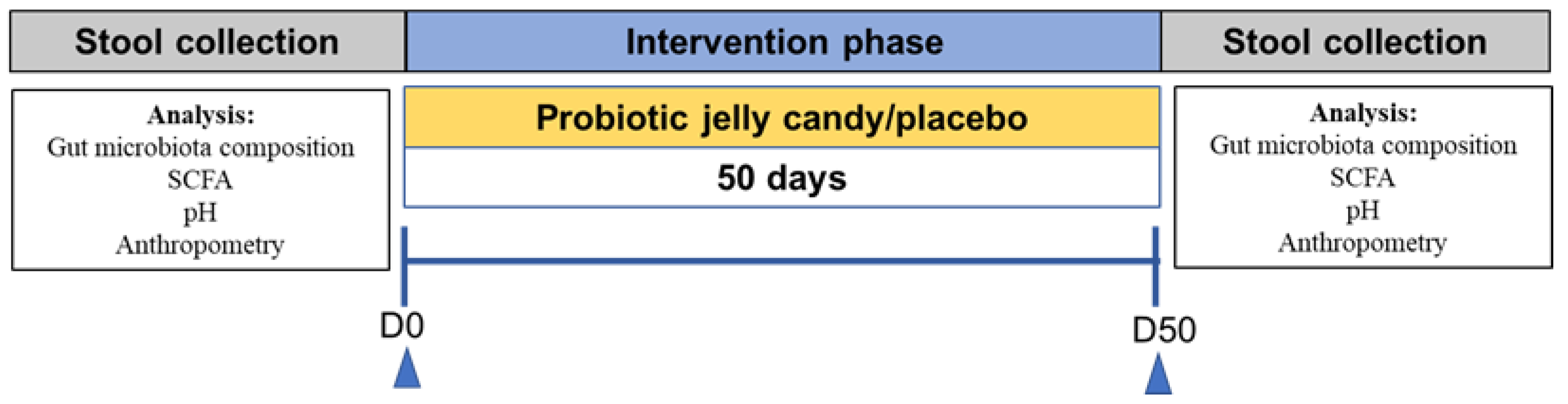
2.5. Research Design
2.6. Research Outcome
2.6.1. Anthropometric Measurement
2.6.2. Dietary Intake Analysis
2.6.3. Stool Sample Collection and DNA Extraction
2.6.4. DNA Quality Control and Purification for 16S rRNA Sequencing
2.6.5. 16S rRNA Sequencing
2.6.6. Sequencing Data Processing
2.6.7. qPCR Analysis
2.6.8. Stool pH and SCFA Analysis
2.7. Statistical Analysis
3. Results
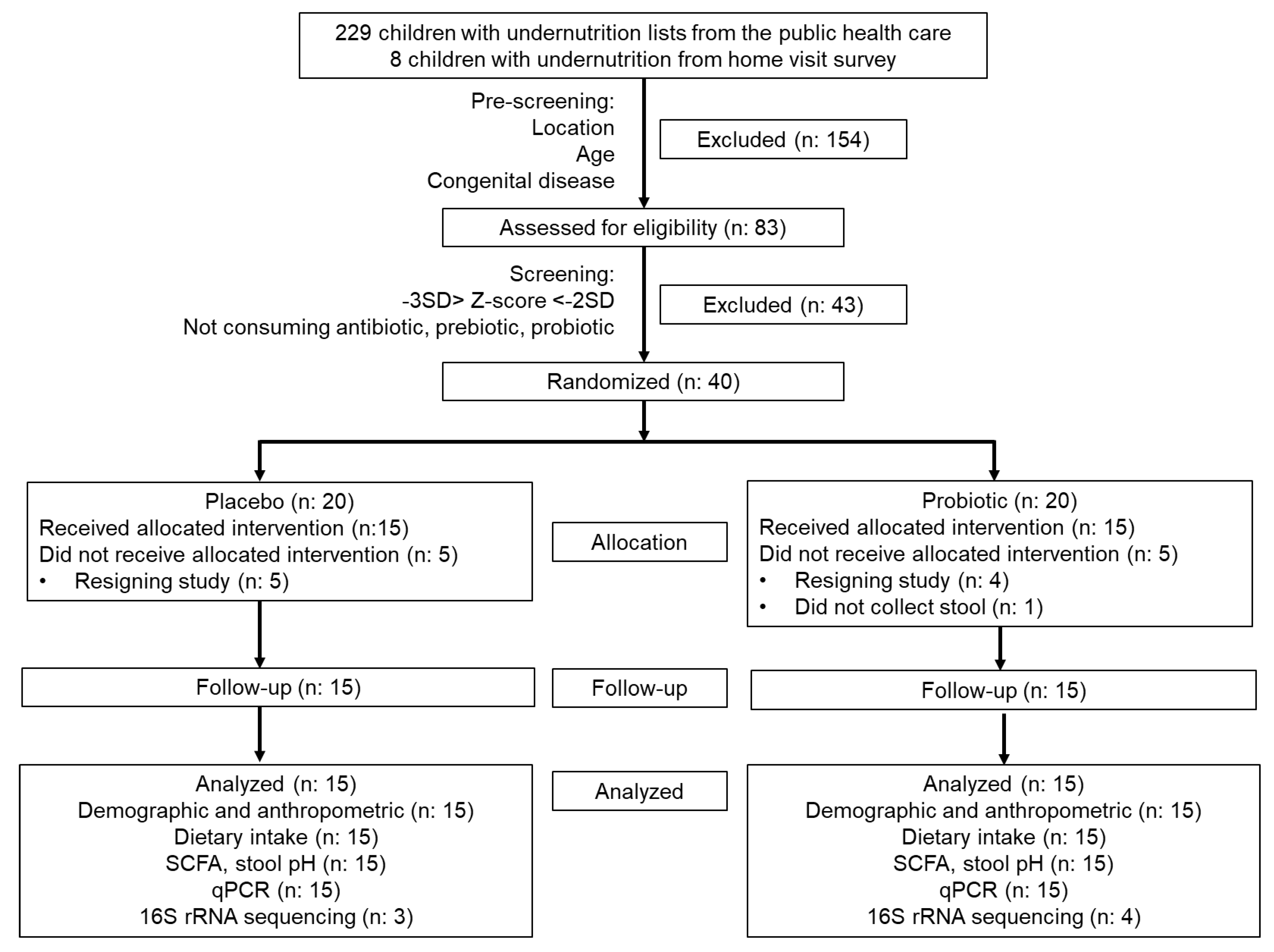
3.1. Demographic Data and Participant Flowchart
3.2. Dietary Intake
3.3. The Changes in Anthropometry and Nutritional Status
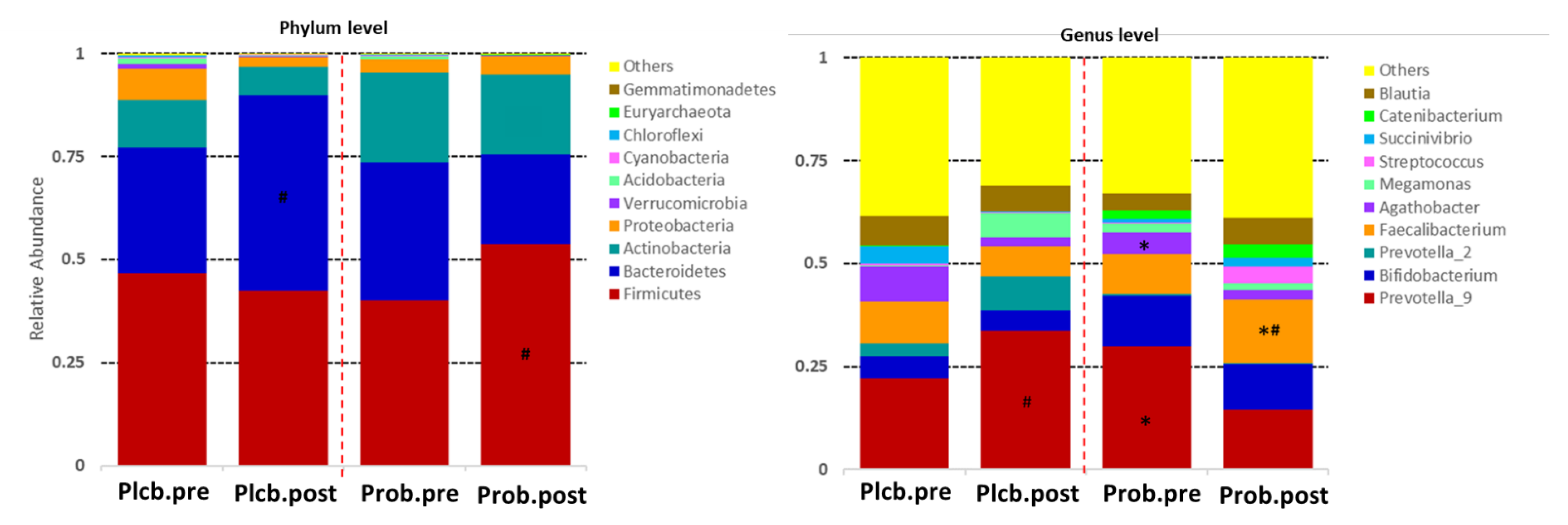
3.4. The Changes of Gut Microbiota Taxonomic between Groups
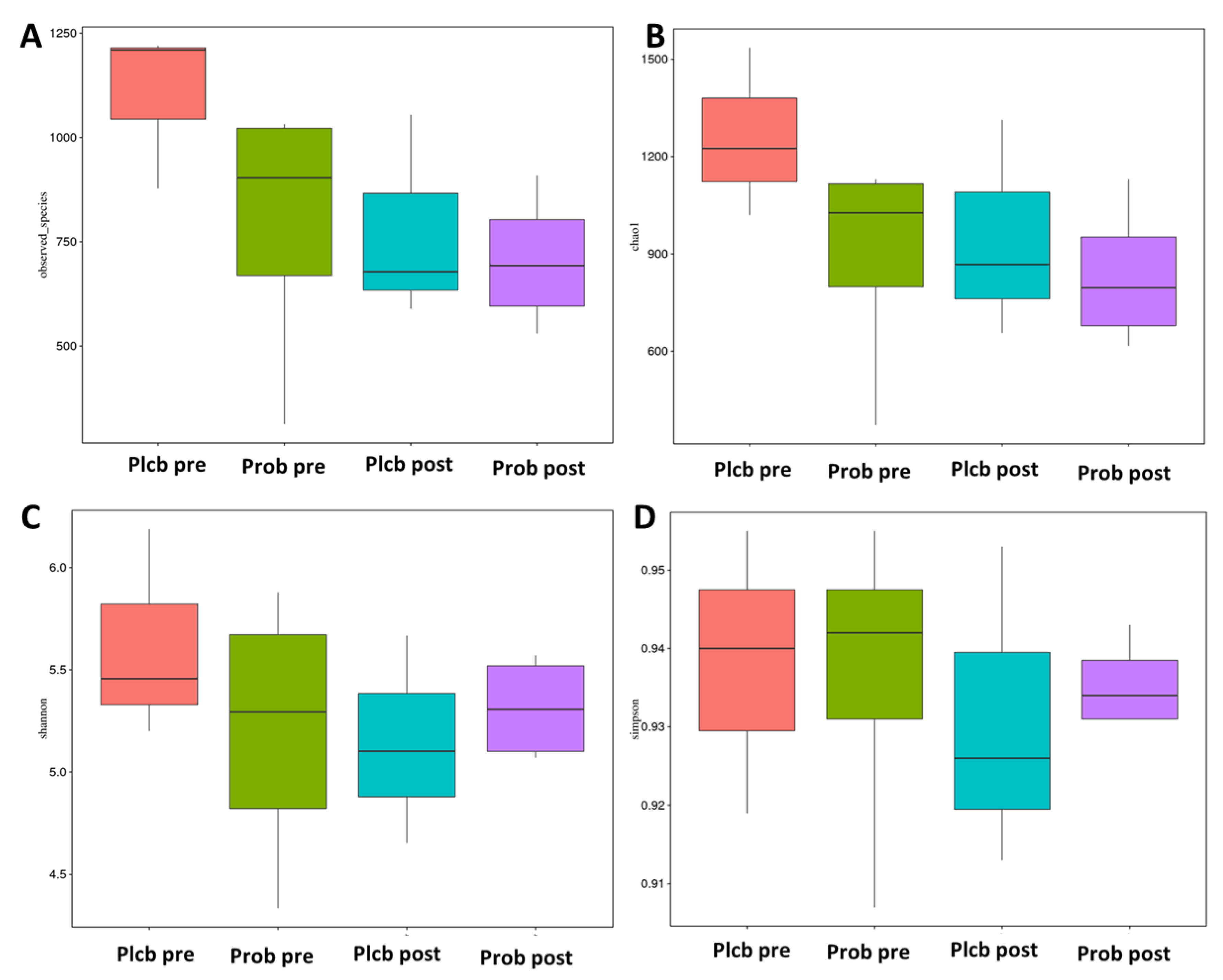
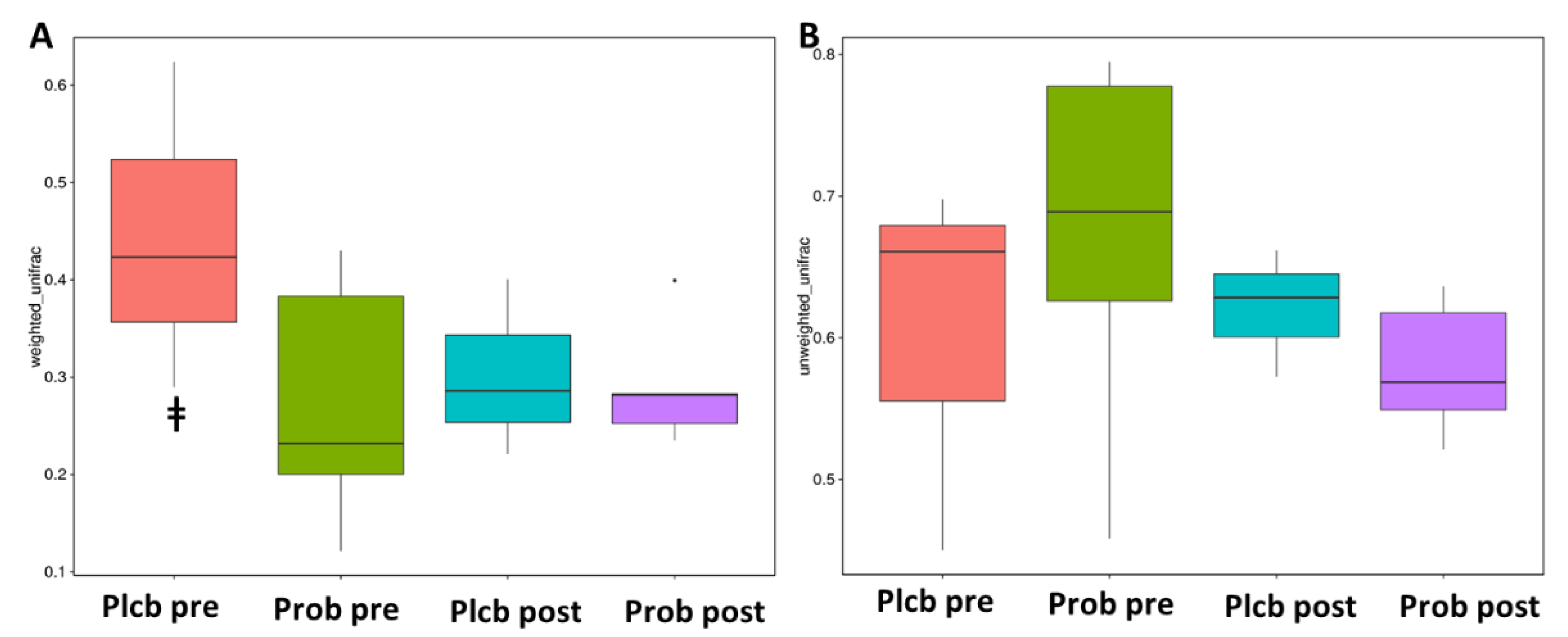
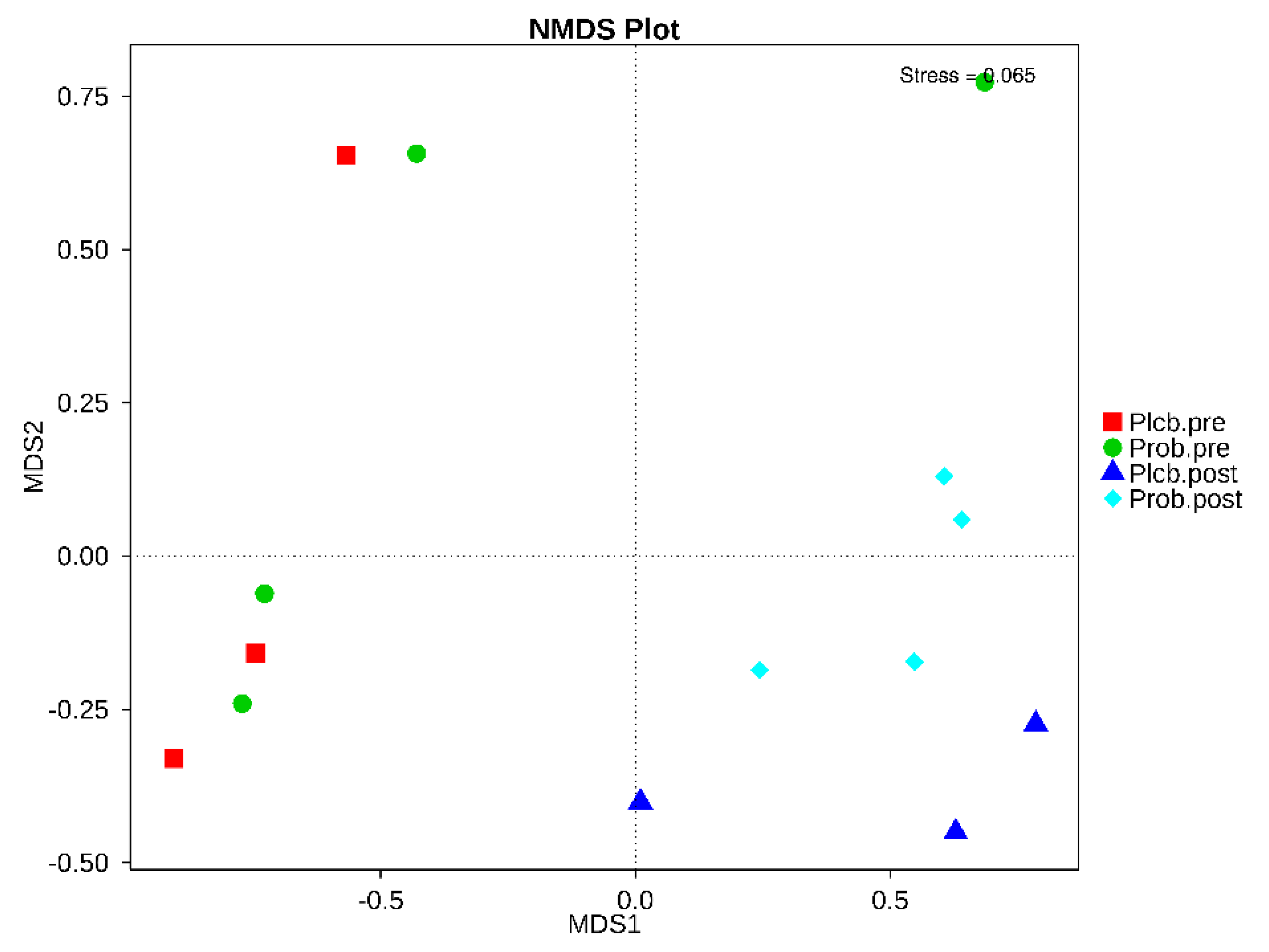
3.5. The Changes in Gut Microbiota Diversity and Composition
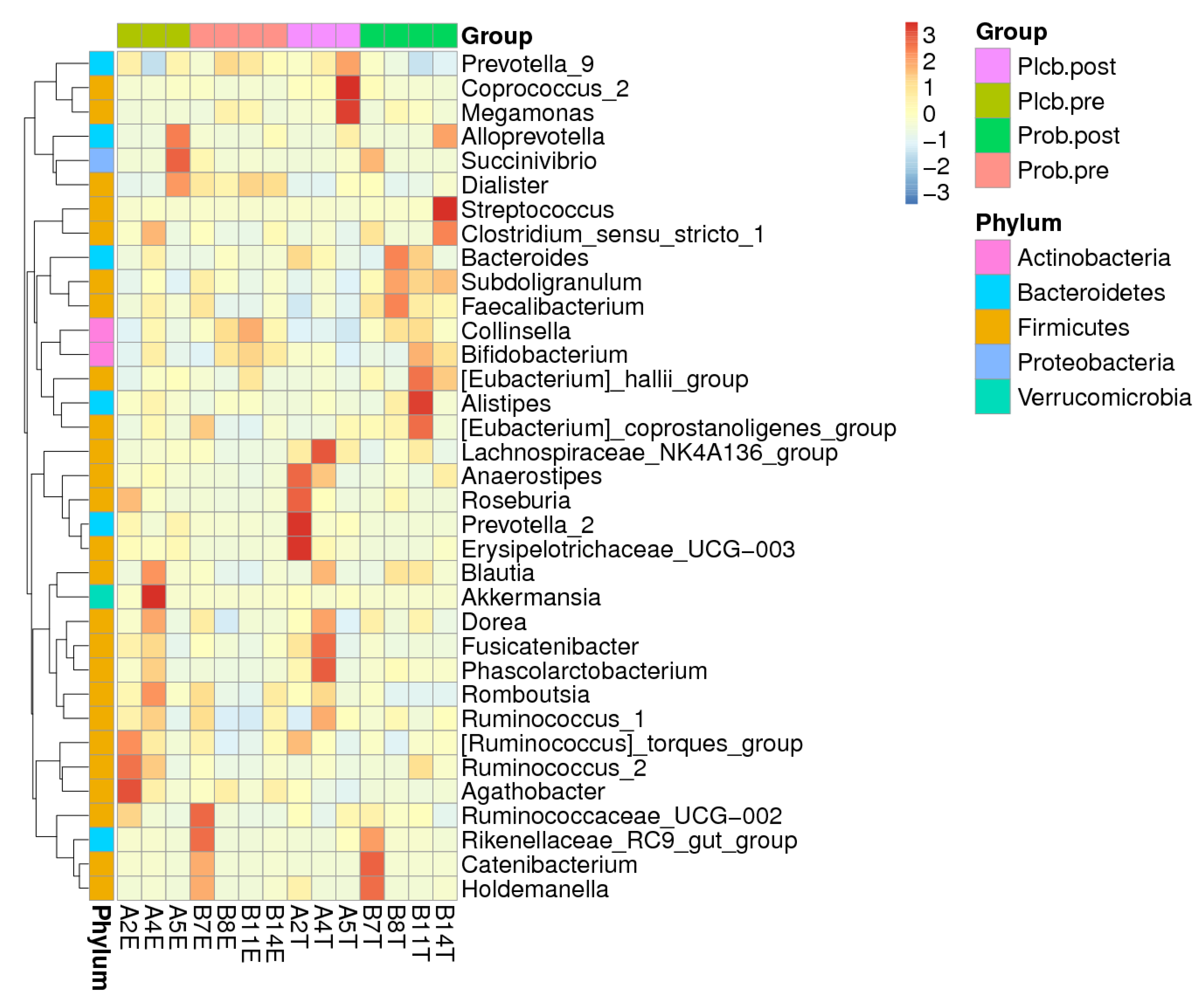
3.6. Gut Microbiota Biomarker Identification
3.7. Specific Bacterial Quantification
3.8. SCFA Concentration and Stool pH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kementerian Kesehatan Badan Penelitian dan Kesehatan. Hasil utama riskesdas 2018; Lembaga Penerbit Badan Penelitian dan Pengembangan Kesehatan: Jakarta, Indonesia, 2018; Available online: http://labdata.litbang.kemkes.go.id/ccount/click.php?id=19 (accessed on 26 November 2021).
- WHO/UNICEF. WHO Child Growth Standards and the Identification of Severe Acute Malnutrition in Infants and Children (World Health Organization United Nations Childrens Fund); WHO: Geneva, Switzerland; UNICEF: New York, NY, USA, 2009; ISBN 978-92-4-159816-3. [Google Scholar]
- Monira, S.; Nakamura, S.; Gotoh, K.; Izutsu, K.; Watanabe, H.; Alam, N.H.; Endtz, H.P.; Cravioto, A.; Ali, S.I.; Nakaya, T.; et al. Gut Microbiota of Healthy and Malnourished Children in Bangladesh. Front. Microbiol. 2011, 2, 1–7. [Google Scholar] [CrossRef]
- Alou, M.T.; Million, M.; Traore, S.I.; Mouelhi, D.; Khelaifia, S.; Bachar, D.; Caputo, A.; Delerce, J.; Brah, S.; Alhousseini, D.; et al. Gut Bacteria Missing in Severe Acute Malnutrition, Can We Identify Potential Probiotics by Culturomics? Front. Microbiol. 2017, 8, 1–17. [Google Scholar] [CrossRef]
- Subramanian, S.; Huq, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; Destefano, J.; Meier, M.F.; Muegge, B.D.; et al. Persistent Gut Microbiota Immaturity in Malnourished Bangladeshi Children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.I.; Yatsunenko, T.; Manary, M.J.; Trehan, I.; Mkakosya, R.; Cheng, J.; Kau, A.L.; Rich, S.S.; Concannon, P.; Mychaleckyj, J.C.; et al. Gut Microbiomes of Malawian Twin Pairs Discordant for Kwashiorkor. Science 2013, 339, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Ward, H.; Ghosh, S.; Rogers, B.; Rosenberg, I. Biomarkers of Environmental Enteric DDsfunction (EED) Predict Growth and Recovery Among Children with Moderate Acute Malnutrition (MAM) in Sierra Leone. Curr. Dev. Nutr. 2020, 4, 1081. [Google Scholar] [CrossRef]
- Keusch, G.T.; Denno, D.M.; Black, R.E.; Duggan, C.; Guerrant, R.L.; Lavery, J.V.; Nataro, J.P.; Rosenberg, I.H.; Ryan, E.T.; Tarr, P.I.; et al. Environmental Enteric Dysfunction: Pathogenesis, Diagnosis, and Clinical Consequences. Clin. Infect. Dis. 2014, 59, 207–212. [Google Scholar] [CrossRef]
- Kamil, R.Z.; Murdiati, A.; Juffrie, M.; Nakayama, J.; Rahayu, E.S. Gut Microbiota and Short-Chain Fatty Acid Profile between Normal and Moderate Malnutrition Children in Yogyakarta, Indonesia. Microorganisms 2021, 9, 1–15. [Google Scholar] [CrossRef]
- Velly, H.; Britton, R.A.; Preidis, G.A. Mechanisms of Cross-Talk between the Diet, the Intestinal Microbiome, and the Undernourished Host. Gut Microbes 2017, 8, 98–112. [Google Scholar] [CrossRef]
- Pérez-Cobas, A.E.; Artacho, A.; Knecht, H.; Ferrús, M.L.; Friedrichs, A.; Ott, S.J.; Moya, A.; Latorre, A.; Gosalbes, M.J. Differential Effects of Antibiotic Therapy on the Structure and Function of Human Gut Microbiota. PLoS ONE 2013, 8, e80201. [Google Scholar] [CrossRef]
- Food and Agriculture Organization/World Health Organization. Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food; Food and Agriculture Organization/World Health Organization: London, ON, Canada, 2002. [Google Scholar]
- Rahayu, E.S.; Yogeswara, A.; Maryatun, M.; Windiarti, L.; Utami, T.; Watanabe, K. Molecular Characteristics of Indigenous Probiotic Strains from Indonesia. Int. J. Probiotics Prebiotics 2015, 10, 109–116. [Google Scholar]
- Rahayu, E.S.; Rusdan, I.H.; Athennia, A.; Kamil, R.Z.; Pramesi, P.C.; Marsono, Y.; Utami, T.; Widada, J. Safety Assessment of Indigenous Probiotic Strain Lactobacillus Plantarum Dad-13 Isolated from Dadih Using Sprague Dawley Rats as a Model. Am. J. Pharmacol. Toxicol. 2019, 14, 38–47. [Google Scholar] [CrossRef]
- Sazawal, S.; Dhingra, U.; Hiremath, G.; Sarkar, A.; Dhingra, P.; Dutta, A.; Menon, V.P.; Black, R.E. Effects of Bifidobacterium Lactis HN019 and Prebiotic Oligosaccharide Added to Milk on Iron Status, Anemia, and Growth among Children 1 to 4 Years Old. J. Pediatric Gastroenterol. Nutr. 2010, 51, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Saran, S.; Gopalan, S.; Krishna, T.P. Use of Fermented Foods to Combat Stunting and Failure to Thrive. Nutrition 2002, 18, 393–396. [Google Scholar] [CrossRef]
- Surono, I.S.; Koestomo, F.P.; Novitasari, N.; Zakaria, F.R.; Koesnandar, Y. Novel Probiotic Enterococcus Faecium IS-27526 Supplementation Increased Total Salivary SIgA Level and Bodyweight of Pre-School Children: A Pilot Study. Anaerobe 2011, 17, 496–500. [Google Scholar] [CrossRef]
- Kamil, R.Z.; Fadhila, F.H.; Rachmasari, A.D.; Murdiati, A.; Juffrie, M.; Rahayu, E.S. Development of Probiotic Gummy Candy Using the Indigenous Lactobacillus Plantarum Dad-13 Strain: Evaluation of Its Gastrointestinal Resistance and Shelf-Life Prediction. Food Res. 2021, 5, 265–273. [Google Scholar] [CrossRef]
- Lwanga, S.K.; Lemeshow, S. Sample Size Determination in Health Studies: A Practical Manual; World Health Organization: Geneva, Switzerland, 1991; ISBN 9241544058. [Google Scholar]
- Kamil, R.Z.; Yanti, R.; Murdiati, A.; Jurffrie, M.; Rahayu, E.S. Microencapsulation of Indigenous Probiotic Lactobacillus Plantarum Dad-13 by Spray and Freeze-Drying: Strain-Dependent Effect and Its Antibacterial Property. Food Res. 2020, 4, 2181–2189. [Google Scholar] [CrossRef]
- Nakayama, J. Pyrosequence-Based 16S RRNA Profiling of Gastro-Intestinal Microbiota. Biosci. Microflora 2010, 29, 83–96. [Google Scholar] [CrossRef][Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-Filtering Vastly Improves Diversity Estimates from Illumina Amplicon Sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME Improves Sensitivity and Speed of Chimera Detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Matsuki, T.; Watanabe, K.; Fujimoto, J.; Miyamoto, Y.; Takada, T.; Matsumoto, K.; Oyaizu, H.; Tanaka, R. Development of 16S RRNA-Gene-Targeted Group-Specific Primers for the Detection and Identification of Predominant Bacteria in Human Feces. Appl. Environ. Microbiol. 2002, 68, 5445–5451. [Google Scholar] [CrossRef]
- Matsuda, K.; Tsuji, H.; Asahara, T.; Matsumoto, K.; Takada, T.; Nomoto, K. Establishment of an Analytical System for the Human Fecal Microbiota, Based on Reverse Transcription-Quantitative PCR Targeting of Multicopy RRNA Molecules. Appl. Environ. Microbiol. 2009, 75, 1961–1969. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, K.; Tsuji, H.; Asahara, T.; Kado, Y.; Nomoto, K. Sensitive Quantitative Detection of Commensal Bacteria by RRNA-Targeted Reverse Transcription-PCR. Appl. Environ. Microbiol. 2007, 73, 32–39. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic Biomarker Discovery and Explanation. Genome Biol. 2011, 12, 1–18. [Google Scholar] [CrossRef]
- Anukam, K.C.; Osazuwa, E.O.; Reid, G. Improved Appetite of Pregnant Rats and Increased Birth Weight of Newborns Following Feeding with Probiotic Lactobacillus Rhamnosus GR-1 and Lactobacillus Fermentum RC-14. J. Appl. Res. 2005, 5, 46–52. [Google Scholar]
- Kazemi, A.; Noorbala, A.A.; Djafarian, K. Effect of Probiotic and Prebiotic versus Placebo on Appetite in Patients with Major Depressive Disorder: Post Hoc Analysis of a Randomised Clinical Trial. J. Hum. Nutr. Diet. 2020, 33, 56–65. [Google Scholar] [CrossRef]
- Harahap, I.A.; Suliburska, J. Probiotics and Isoflavones as a Promising Therapeutic for Calcium Status and Bone Health: A Narrative Review. Foods 2021, 10, 2685. [Google Scholar] [CrossRef]
- Whitfield, K.C.; Smith, G.; Chamnan, C.; Karakochuk, C.D.; Sophonneary, P.; Kuong, K.; Dijkhuizen, M.A.; Hong, R.; Berger, J.; Green, T.J.; et al. High Prevalence of Thiamine (Vitamin B1) Deficiency in Early Childhood among a Nationally Representative Sample of Cambodian Women of Childbearing Age and Their Children. PLoS Negl. Trop. Dis. 2017, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Michaelsen, K.F.; Hoppe, C.; Roos, N.; Kaestel, P.; Stougaard, M.; Lauritzen, L.; Molgaard, C.; Girma, T.; Friis, H. Choice of Foods and Ingredients for Moderately Malnourished Children 6 Months to 5 Years of Age. Food Nutr. Bull. 1992, 30, 343–405. [Google Scholar] [CrossRef] [PubMed]
- Reid, G.; Younes, J.A.; van der Mei, H.C.; Gloor, G.B.; Knight, R.; Busscher, H.J. Microbiota Restoration: Natural and Supplemented Recovery of Human Microbial Communities. Nat. Rev. Microbiol. 2011, 9, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Gargari, G.; Taverniti, V.; Balzaretti, S.; Ferrario, C.; Gardana, C.; Simonetti, P.; Guglielmetti, S. Consumption of a Bifidobacterium Bifidum Strain for 4 Weeks Modulates Dominant Intestinal Bacterial Taxa and Fecal Butyrate in Healthy Adults. Appl. Environ. Microbiol. 2016, 82, 5850–5859. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.F.; Zhu, C.R.; Gong, X.L.; Li, H.L.; Xiong, L.K.; Wang, K.J.; Liu, G.S. Beneficial Effects of Probiotic Treatment on Gut Microbiota in Very Low Birth Weight Infants. Gastroenterol. Res. Pract. 2019, 2019, 1–7. [Google Scholar] [CrossRef]
- Castro-Mejía, J.L.; O’Ferrall, S.; Krych, Ł.; O’Mahony, E.; Namusoke, H.; Lanyero, B.; Kot, W.; Nabukeera-Barungi, N.; Michaelsen, K.F.; Mølgaard, C.; et al. Restitution of Gut Microbiota in Ugandan Children Administered with Probiotics (Lactobacillus Rhamnosus GG and Bifidobacterium Animalis Subsp. Lactis BB-12) during Treatment for Severe Acute Malnutrition. Gut Microbes 2020, 11, 855–867. [Google Scholar] [CrossRef]
- Fluitman, K.S.; de Clercq, N.C.; Keijser, B.J.F.; Visser, M.; Nieuwdorp, M.; Ijzerman, R.G. The Intestinal Microbiota, Energy Balance, and Malnutrition: Emphasis on the Role of Short-Chain Fatty Acids. Expert Rev. Endocrinol. Metab. 2017, 12, 215–226. [Google Scholar] [CrossRef]
- Kang, S.; You, H.J.; Lee, Y.G.; Jeong, Y.; Johnston, T.v.; Baek, N.I.; Ku, S.; Ji, G.E. Production, Structural Characterization, and in Vitro Assessment of the Prebiotic Potential of Butyl-Fructooligosaccharides. Int. J. Mol. Sci. 2020, 21, 445. [Google Scholar] [CrossRef]
- Yan, H.; Potu, R.; Lu, H.; Vezzoni de Almeida, V.; Stewart, T.; Ragland, D.; Armstrong, A.; Adeola, O.; Nakatsu, C.H.; Ajuwon, K.M. Dietary Fat Content and Fiber Type Modulate Hind Gut Microbial Community and Metabolic Markers in the Pig. PLoS ONE 2013, 8, e59581. [Google Scholar] [CrossRef]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I.; Anaerostipes, L.; Faecalibacterium, F. Important Metabolic Pathways and Biological Processes Expressed by Chicken Cecal Microbiota. Appl. Environ. Microbiol. 2016, 82, 1569–1576. [Google Scholar] [CrossRef]
- Shankar, V.; Gouda, M.; Moncivaiz, J.; Gordon, A.; Reo, N.V.; Hussein, L.; Paliy, O. Differences in Gut Metabolites and Microbial Composition and Functions. mSystems 2017, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Leylabadlo, H.E.; Ghotaslou, R.; Feizabadi, M.M.; Farajnia, S.; Moaddab, S.Y.; Ganbarov, K.; Khodadadi, E.; Tanomand, A.; Sheykhsaran, E.; Yousefi, B.; et al. The Critical Role of Faecalibacterium Prausnitzii in Human Health: An Overview. Microb. Pathog. 2020, 149, 104344. [Google Scholar] [CrossRef] [PubMed]
- Thomas, L.V.; Ockhuizen, T.; Suzuki, K. Exploring the Influence of the Gut Microbiota and Probiotics on Health: A Symposium Report. Br. J. Nutr. 2014, 112, S1–S18. [Google Scholar] [CrossRef] [PubMed]
- Vitetta, L.; Llewellyn, H.; Oldfield, D. Gut Dysbiosis and the Intestinal Microbiome: Streptococcus Thermophilus a Key Probiotic for Reducing Uremia. Microorganisms 2019, 7, 228. [Google Scholar] [CrossRef]
- van den Bogert, B.; Meijerink, M.; Zoetendal, E.G.; Wells, J.M.; Kleerebezem, M. Immunomodulatory Properties of Streptococcus and Veillonella Isolates from the Human Small Intestine Microbiota. PLoS ONE 2014, 9, 1–20. [Google Scholar] [CrossRef]
- Nakayama, J.; Watanabe, K.; Jiang, J.; Matsuda, K.; Chao, S.H.; Haryono, P.; La-Ongkham, O.; Sarwoko, M.A.; Sujaya, I.N.; Zhao, L.; et al. Diversity in Gut Bacterial Community of School-Age Children in Asia. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef]
- Rahayu, E.S.; Utami, T.; Mariyatun, M.; Hasan, P.N.; Kamil, R.Z.; Setyawan, R.H.; Pamungkaningtyas, F.H.; Harahap, I.A.; Wiryohanjoyo, D.V.; Pramesi, P.C.; et al. Gut Microbiota Profile in Healthy Indonesians. World J. Gastroenterol. 2019, 9327, 1478–1491. [Google Scholar] [CrossRef]
- Dinh, D.M.; Ramadass, B.; Kattula, D.; Sarkar, R.; Braunstein, P.; Tai, A.; Wanke, C.A.; Hassoun, S.; Kane, A.v.; Naumova, E.N.; et al. Longitudinal Analysis of the Intestinal Microbiota in Persistently Stunted Young Children in South India. PLoS ONE 2016, 11, 1–17. [Google Scholar] [CrossRef]
- Banin, M.M.; Utami, T.; Cahyanto, M.N.; Widada, J.; Rahayu, E.S. Effects of Consumption of Probiotic Powder Containing Lactobacillus Plantarum Dad-13 on Fecal Bacterial Population in School-Age Children in Indonesia. Int. J. Probiotics Prebiotics 2019, 14, 1–8. [Google Scholar] [CrossRef]
- Rahayu, E.S.; Cahyanto, M.N.; Windiarti, L.; Sutriyanto, J.; Kandarina, T.; Utami, T. Effects of Consumption of Fermented Milk Containing Indigenous Probiotic Lactobacillus Plantarum Dad-13 on the Fecal Microbiota of Healthy Indonesian Volunteers. Int. J. Probiotics Prebiotics 2016, 11, 91–98. [Google Scholar]
- Pekmez, C.T.; Dragsted, L.O.; Brahe, L.K. Gut Microbiota Alterations and Dietary Modulation in Childhood Malnutrition—The Role of Short Chain Fatty Acids. Clin. Nutr. 2019, 38, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Faden, H.S.; Zhu, L. The Response of the Gut Microbiota to Dietary Changes in the First Two Years of Life. Front. Pharmacol. 2020, 11, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, P.O.; Bindels, L.B.; Saulnier, D.M.; Reid, G.; Nova, E.; Holmgren, K.; O’Toole, P.W.; Bunn, J.; Delzenne, N.; Scott, K.P. Can Prebiotics and Probiotics Improve Therapeutic Outcomes for Undernourished Individuals? Gut Microbes 2014, 5, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; Jocken, J.W.; Blaak, E.E. Short-Chain Fatty Acids in Control of Body Weight and Insulin Sensitivity. Nat. Rev. Endocrinol. 2015, 11, 577–591. [Google Scholar] [CrossRef]
- Soty, M.; Gautier-Stein, A.; Rajas, F.; Mithieux, G. Gut-Brain Glucose Signaling in Energy Homeostasis. Cell Metab. 2017, 25, 1231–1242. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte Metabolism Shapes the Gut Microbiota Single Sentence Summary. Science 2018, 362, 6418. [Google Scholar] [CrossRef]
- Yan, H.; Ajuwon, K.M. Mechanism of Butyrate Stimulation of Triglyceride Storage and Adipokine Expression during Adipogenic Differentiation of Porcine Stromovascular Cells. PLoS ONE 2015, 10, 1–20. [Google Scholar] [CrossRef]
- Purwandhani, S.N.; Utami, T.; Millati, R.; Rahayu, E.S. Potency of Lactobacillus Plantarum Isolated from Dadih to Increase the Folate Levels in Fermented Milk. Agritech 2018, 37, 395. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | 5′–3′ | Annealing (°C) | Ref |
|---|---|---|---|
| Bifidobacterium | g-Bifid-F CTCCTGGAAACGGGTGG g-Bifid-R GGTGTTCTTCCCGATATCTACA | 58.8 | [28] |
| L. plantarum | sg-Lpla-F CTCTGGTATTGATTGGTGCTTGCAT sg-Lpla-R GTTCGCCACTCACTCAAATGTAAA | 60 | [29] |
| Enterobacteriaceae | En-lsu-3F TGCCGTAACTTCGGGAGAAGGCA En-lsu-3’R TCAAGGACCAGTGTTCAGTGTC | 60 | [30] |
| Placebo (n: 15) | Probiotic (n: 15) | p | |
|---|---|---|---|
| Male | 10 (66.67%) | 9 (60.00%) | |
| Female | 5 (33.33%) | 6 (40.00%) | |
| Age (months) | 37.80 ± 11.78 | 37.93 ± 12.98 | 0.977 |
| Weight (kg) | 11.20 ± 1.96 | 10.84 ± 1.43 | 0.563 |
| Height (cm) | 88.88 ± 8.00 | 87.06 ± 6.84 | 0.509 |
| WHZ | −1.40 ± 0.61 | −1.19 ± 0.87 | 0.436 |
| WAZ | −2.22 ± 0.74 | −2.28 ± 0.94 | 0.838 |
| HAZ | −2.21 ± 0.79 | −2.55 ± 1.03 | 0.512 |
| Unit | Placebo | p | Probiotic | p | |||
|---|---|---|---|---|---|---|---|
| Before | After | Before | After | ||||
| Energy | kcal | 677.13 ± 189.46 | 653.84 ± 185.74 | 0.733 | 681.99 ± 262.33 | 747.42 ± 263.18 | 0.427 |
| Protein | g | 27.58 ± 6.82 | 26.63 ± 7.79 | 0.570 | 27.39 ± 8.21 | 29.56 ± 10.57 | 0.670 |
| Fat | g | 26.13 ± 8.07 | 25.21 ± 8.38 | 0.638 | 26.74 ± 11.97 | 29.37 ± 12.26 | 0.320 |
| Carbohydrate | g | 83.15 ± 28.38 | 80.05 ± 25.79 | 0.776 | 82.93 ± 34.05 | 91.49 ± 33.36 | 0.363 |
| Fiber | g | 3.56 ± 1.73 | 3.17 ± 1.53 | 0.197 | 3.09 ± 1.37 | 3.13 ± 1.33 | 0.861 |
| Vit. A | µg | 386.71 ± 207.88 | 376.87 ± 140.36 | 0.955 | 584.82 ± 401.09 | 442.17 ± 240.49 | 0.532 |
| Vit. E | mg | 2.75 ± 1.40 | 3.33 ± 1.19 | 0.094 | 2.83 ± 1.33 | 3.43 ± 1.84 | 0.207 |
| Vit. D | µg | 2.83 ± 2.26 | 3.82 ± 2.09 | 0.152 | 3.17 ± 2.16 | 4.15 ± 3.06 | 0.147 |
| Vit. B1 | mg | 0.28 ± 0.10 | 0.28 ± 0.10 | 0.971 | 0.25 ± 0.10 | 0.31 ± 0.14 | 0.058 |
| Vit. B2 | mg | 0.50 ± 0.20 | 0.53 ± 0.18 | 0.558 | 0.57 ± 0.23 | 0.58 ± 0.25 | 0.969 |
| Vit. B6 | mg | 0.43 ± 0.14 | 0.40 ± 0.12 | 0.371 | 0.41 ± 0.16 | 0.43 ± 0.14 | 0.587 |
| Vit. K | µg | 5.47 ± 3.76 | 3.22 ± 3.14 | 0.050 | 3.53 ± 1.48 | 2.71 ± 2.19 | 0.686 |
| Folic acid | µg | 75.93 ± 30.71 | 64.83 ± 20.56 | 0.211 | 89.82 ± 36.89 | 79.85 ± 36.92 | 0.649 |
| Vit. C | mg | 19.18 ± 16.25 | 33.08 ± 19.75 | 0.009 | 24.91 ± 19.07 | 35.46 ± 22.79 | 0.078 |
| Na | mg | 245.35 ± 124.63 | 298.76 ± 165.15 | 0.363 | 256.49 ± 110.42 | 323.35 ± 147.23 | 0.281 |
| K | mg | 681.55 ± 341.62 | 754.07 ± 274.48 | 0.363 | 747.73 ± 360.18 | 859.49 ± 411.01 | 0.460 |
| Ca | mg | 291.37 ± 251.24 | 379.77 ± 212.47 | 0.140 | 340.08 ± 246.67 | 443.14 ± 315.65 | 0.281 |
| Mg | mg | 96.25 ± 32.86 | 92.79 ± 30.55 | 0.460 | 93.69 ± 36.68 | 99.03 ± 37.24 | 0.460 |
| P | mg | 432.19 ± 180.76 | 471.27 ± 169.25 | 0.427 | 437.97 ± 188.58 | 515.21 ± 254.79 | 0.307 |
| Fe | mg | 5.29 ± 3.00 | 5.61 ± 2.54 | 0.670 | 6.52 ± 3.65 | 6.24 ± 3.49 | 0.615 |
| Zn | mg | 3.47 ± 1.08 | 3.35 ± 1.07 | 0.801 | 3.39 ± 1.17 | 3.77 ± 1.49 | 0.460 |
| Parameter | Group | Before | After | p | Increment | p |
|---|---|---|---|---|---|---|
| Weight (kg) | Placebo | 11.20 ± 1.96 | 11.59 ± 1.96 | 0.000 | 0.39 ± 0.30 | 0.109 |
| Probiotic | 10.84 ± 1.43 | 11.43 ± 1.38 | 0.000 | 0.59 ± 0.36 | ||
| Height (cm) | Placebo | 88.88 ± 8.00 | 90.17 ± 8.25 | 0.000 | 1.29 ± 0.68 | 0.980 |
| Probiotic | 87.06 ± 6.84 | 88.35 ± 6.67 | 0.000 | 1.29 ± 0.75 | ||
| WHZ | Placebo | −1.40 ± 0.61 | −1.30 ± 0.74 | 0.140 | 0.11 ± 0.39 | 0.187 |
| Probiotic | −1.19 ± 0.87 | −0.90 ± 0.76 | 0.022 | 0.30 ± 0.48 | ||
| WAZ | Placebo | −2.22 ± 0.74 | −2.04 ± 0.78 | 0.012 | 0.18 ± 0.24 | 0.187 |
| Probiotic | −2.28 ± 0.94 | −2.01 ± 0.76 | 0.080 | 0.27 ± 0.30 | ||
| HAZ | Placebo | −2.21 ± 0.79 | −2.04 ± 0.74 | 0.256 | 0.17 ± 0.27 | 0.806 |
| Probiotic | −2.55 ± 1.03 | −2.35 ± 1.03 | 0.015 | 0.20 ± 0.26 |
| vs. Group | R2 | p |
|---|---|---|
| PlcbPre–ProbPre | 0.1519 | 0.646 |
| PlcbPost–ProbPost | 0.33456 | 0.001 |
| PlcbPre–PlcbPost | 0.21162 | 0.500 |
| ProbPre–ProbPost | 0.19063 | 0.265 |
| Group | Log 10 Bacterial Cells/g Feces | p | ||
|---|---|---|---|---|
| Before | After | |||
| L. plantarum | Placebo | 4.89 ± 0.32 | 4.89 ± 0.54 | 0.887 |
| Probiotic | 4.85 ± 0.30 | 5.53 ± 0.79 | 0.027 | |
| Bifidobacterium | Placebo | 6.24 ± 1.54 | 6.07 ± 0.84 | 0.087 |
| Probiotic | 6.24 ± 1.21 | 6.50 ± 0.93 | 0.776 | |
| Enterobacteriaceae | Placebo | 6.55 ± 0.68 | 6.28 ± 0.56 | 0.221 |
| Probiotic | 6.27 ± 0.67 | 5.80 ± 0.76 | 0.027 | |
| SCFA (mmol/g Feces) | ||||
|---|---|---|---|---|
| Group | Before | After | p | |
| Total SCFA | Placebo | 35.83 ± 17.22 | 29.28 ± 15.26 | 0.185 |
| Probiotic | 23.55 ± 9.03 | 33.78 ± 14.16 | 0.024 | |
| Acetic acid | Placebo | 21.77 ± 12.07 | 17.41 ± 9.79 | 0.194 |
| Probiotic | 15.28 ± 7.61 | 19.40 ± 7.63 | 0.156 | |
| Propionic acid | Placebo | 6.57 ± 3.75 | 6.92 ± 4.70 | 0.930 |
| Probiotic | 4.43 ± 2.46 | 6.89 ± 3.95 | 0.053 | |
| Butyric acid | Placebo | 5.04 ± 2.64 | 3.56 ± 2.32 | 0.023 |
| Probiotic | 2.62 ± 1.59 | 4.67 ± 2.95 | 0.017 | |
| Stool pH | ||||
| Group | Before | After | p | |
| pH | Placebo | 6.23 ± 0.29 | 6.29 ± 0.35 | 0.607 |
| Probiotic | 6.28 ± 0.28 | 6.10 ± 0.46 | 0.185 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamil, R.Z.; Murdiati, A.; Juffrie, M.; Rahayu, E.S. Gut Microbiota Modulation of Moderate Undernutrition in Infants through Gummy Lactobacillus plantarum Dad-13 Consumption: A Randomized Double-Blind Controlled Trial. Nutrients 2022, 14, 1049. https://doi.org/10.3390/nu14051049
Kamil RZ, Murdiati A, Juffrie M, Rahayu ES. Gut Microbiota Modulation of Moderate Undernutrition in Infants through Gummy Lactobacillus plantarum Dad-13 Consumption: A Randomized Double-Blind Controlled Trial. Nutrients. 2022; 14(5):1049. https://doi.org/10.3390/nu14051049
Chicago/Turabian StyleKamil, Rafli Zulfa, Agnes Murdiati, Mohammad Juffrie, and Endang Sutriswati Rahayu. 2022. "Gut Microbiota Modulation of Moderate Undernutrition in Infants through Gummy Lactobacillus plantarum Dad-13 Consumption: A Randomized Double-Blind Controlled Trial" Nutrients 14, no. 5: 1049. https://doi.org/10.3390/nu14051049
APA StyleKamil, R. Z., Murdiati, A., Juffrie, M., & Rahayu, E. S. (2022). Gut Microbiota Modulation of Moderate Undernutrition in Infants through Gummy Lactobacillus plantarum Dad-13 Consumption: A Randomized Double-Blind Controlled Trial. Nutrients, 14(5), 1049. https://doi.org/10.3390/nu14051049