
Dosage of Dual-Protein Nutrition Differentially Impacts the Formation of Atherosclerosis in ApoE−/− Mice


 ,
,
Abstract
:
1. Introduction
2. Materials and Methods
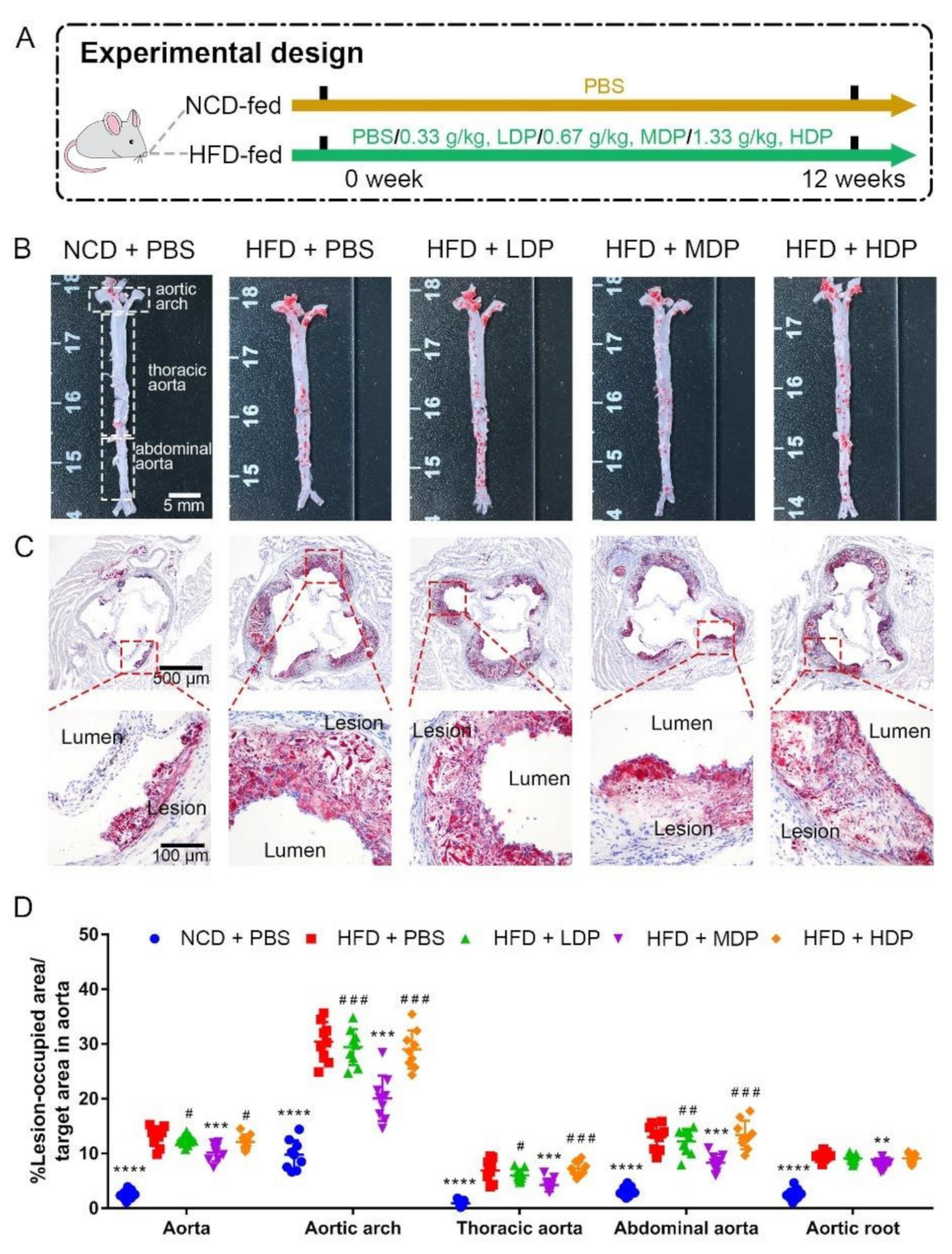
2.1. Food Supplement and Diet
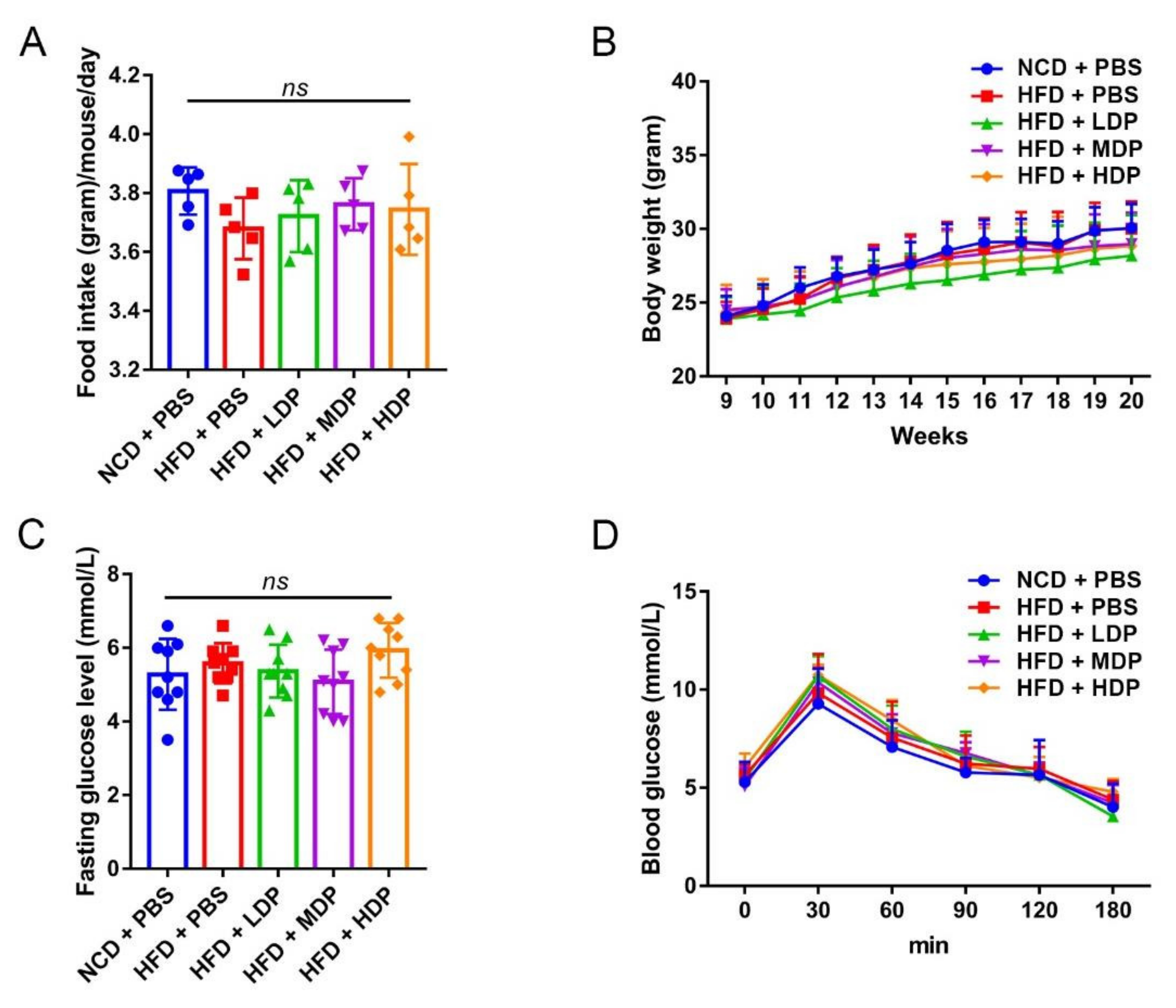
2.2. Animal Raising and Modeling
2.3. Tissue Sampling
2.4. Biochemical and Immunological Assays of Plasma
2.5. Atherosclerotic Lesion Analysis
2.6. Morphologic and Histology Analysis
2.7. Immunofluorescent and Immunohistochemical Assay
2.8. RT-qPCR Analysis of Particular mRNA Levels in Liver
2.9. Glucose Tolerance Test
2.10. 16S rRNA Sequencing of Cecal Contents
2.11. Statistical Analysis
3. Results
3.1. Oral Treatment with MDP Alleviated High-Fat-Diet-Induced Atherosclerotic Lesion of ApoE−/− Mice
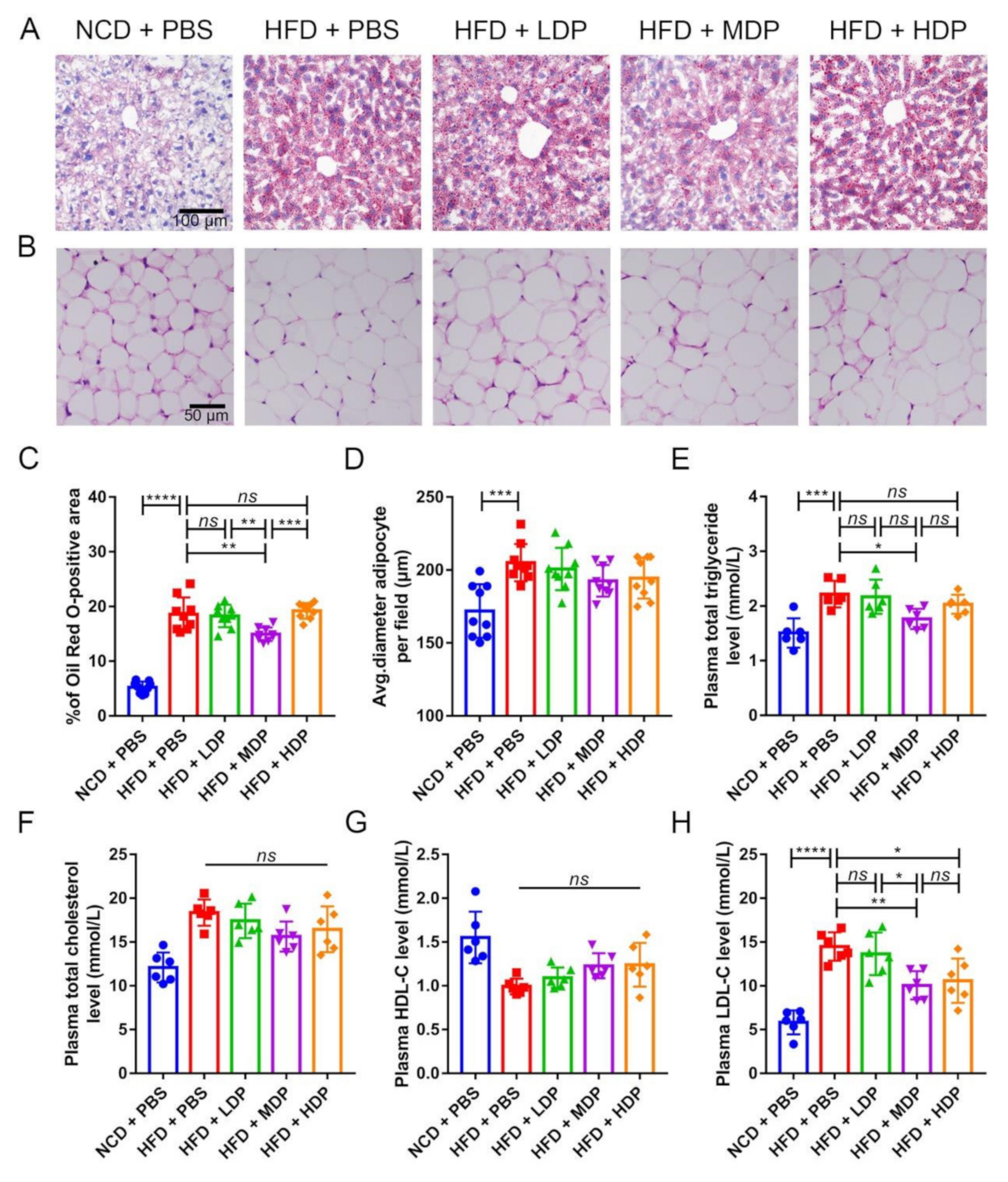
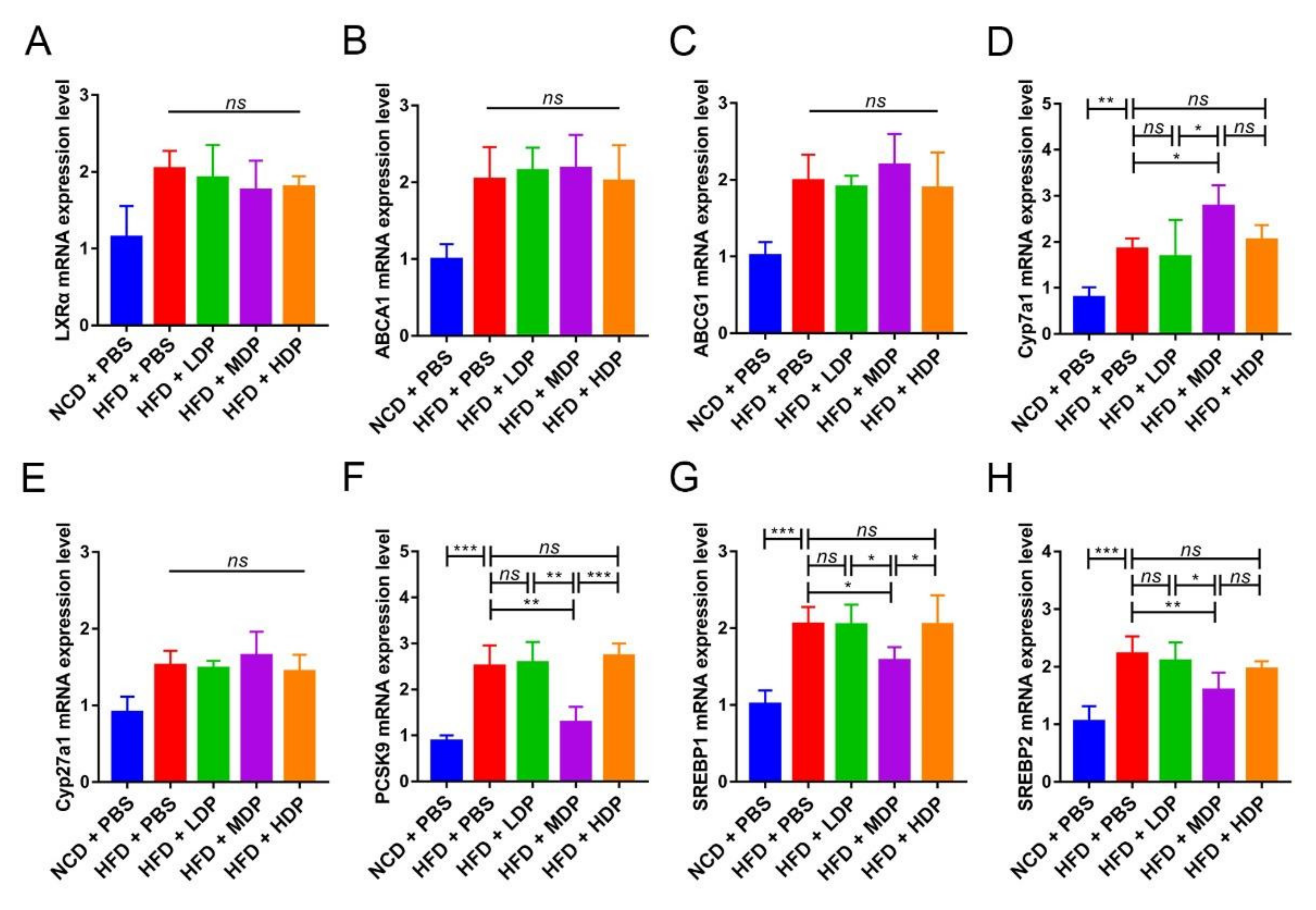
3.2. Oral Gavage with MDP Improved Lipid Metabolism in ApoE−/− Mice
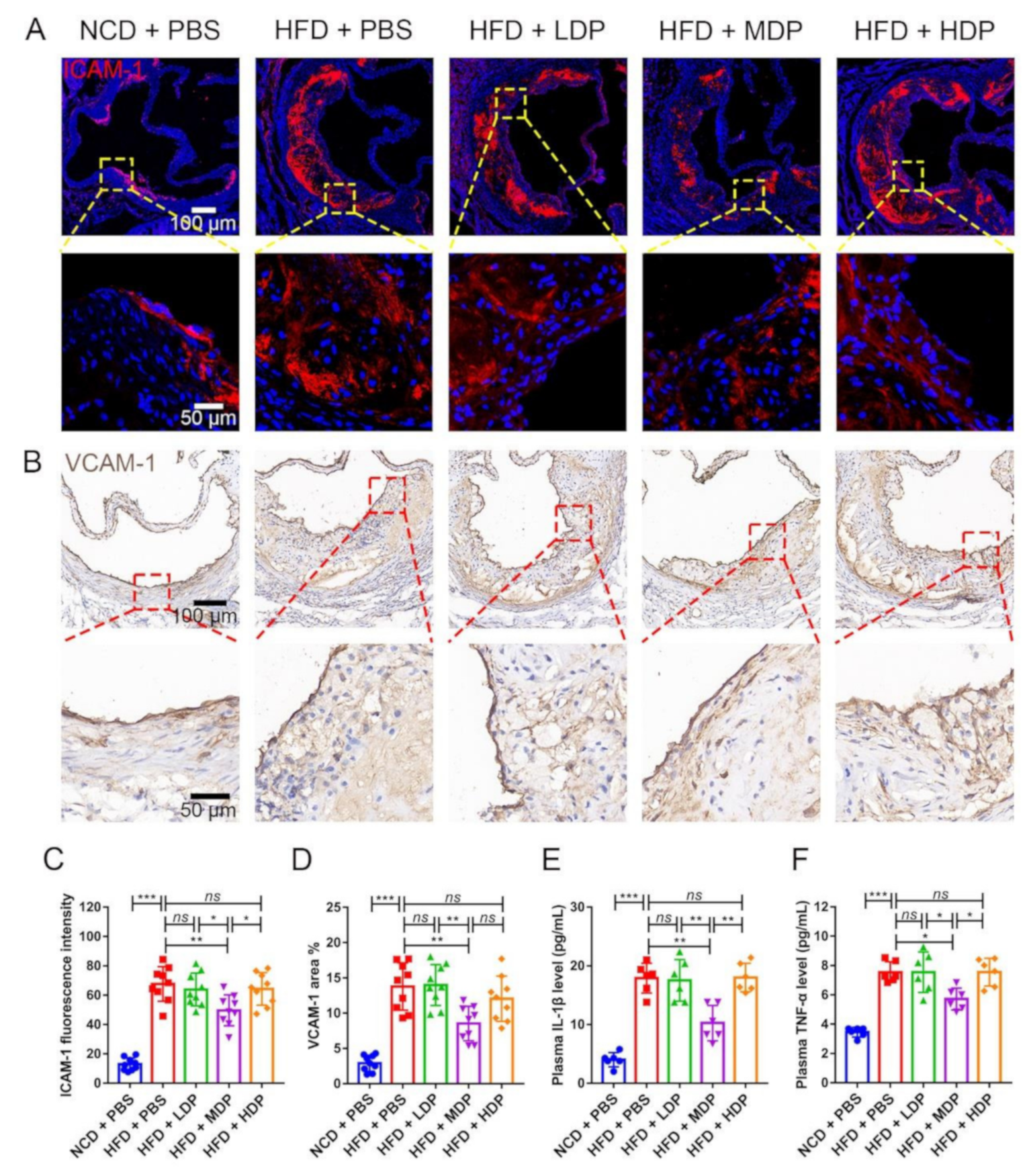
3.3. MDP Administration Mitigated Both Local and Systemic Inflammation in ApoE−/− Mice
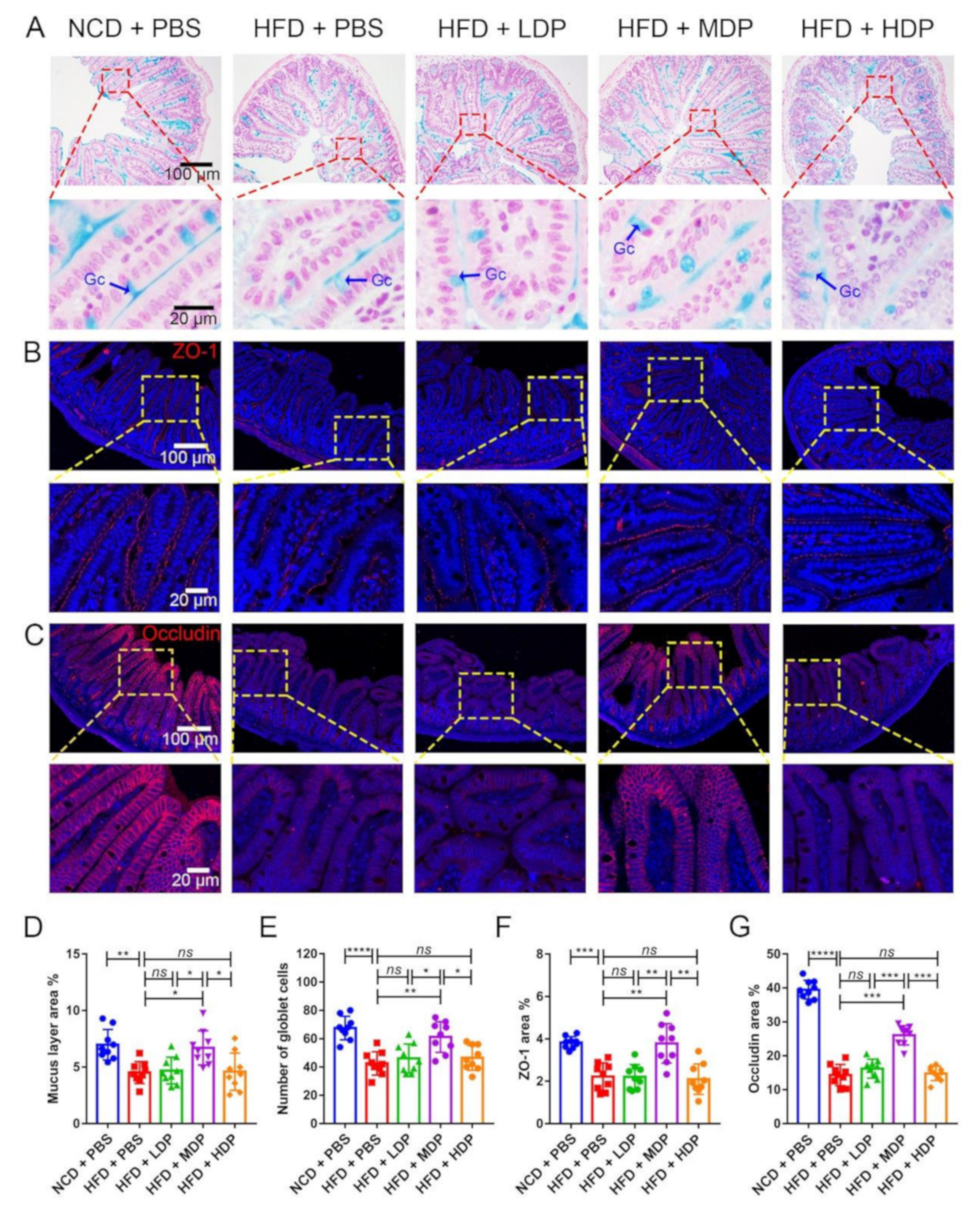
3.4. Oral Gavage with MDP Ameliorated Intestinal Barrier Function in ApoE−/− Mice
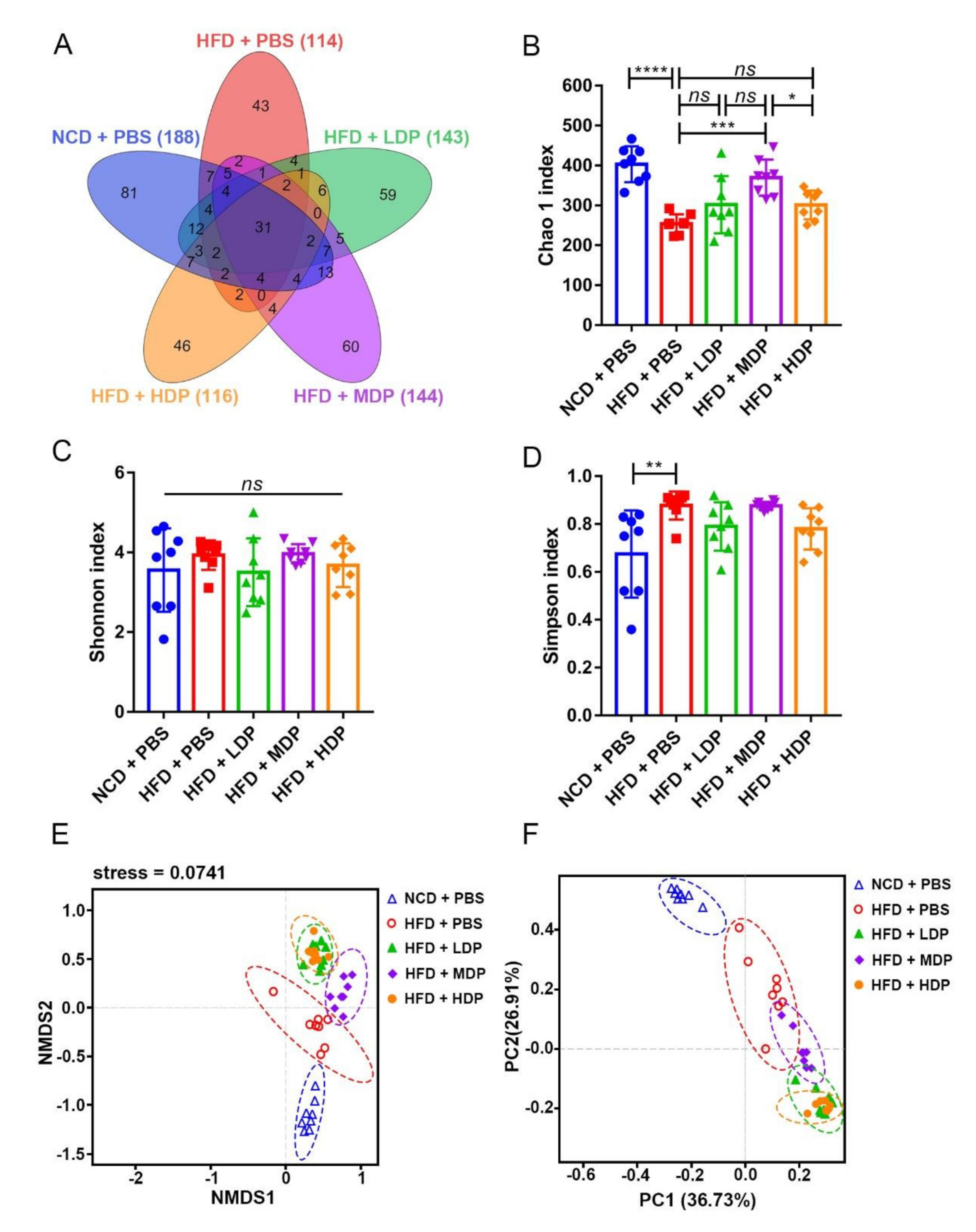
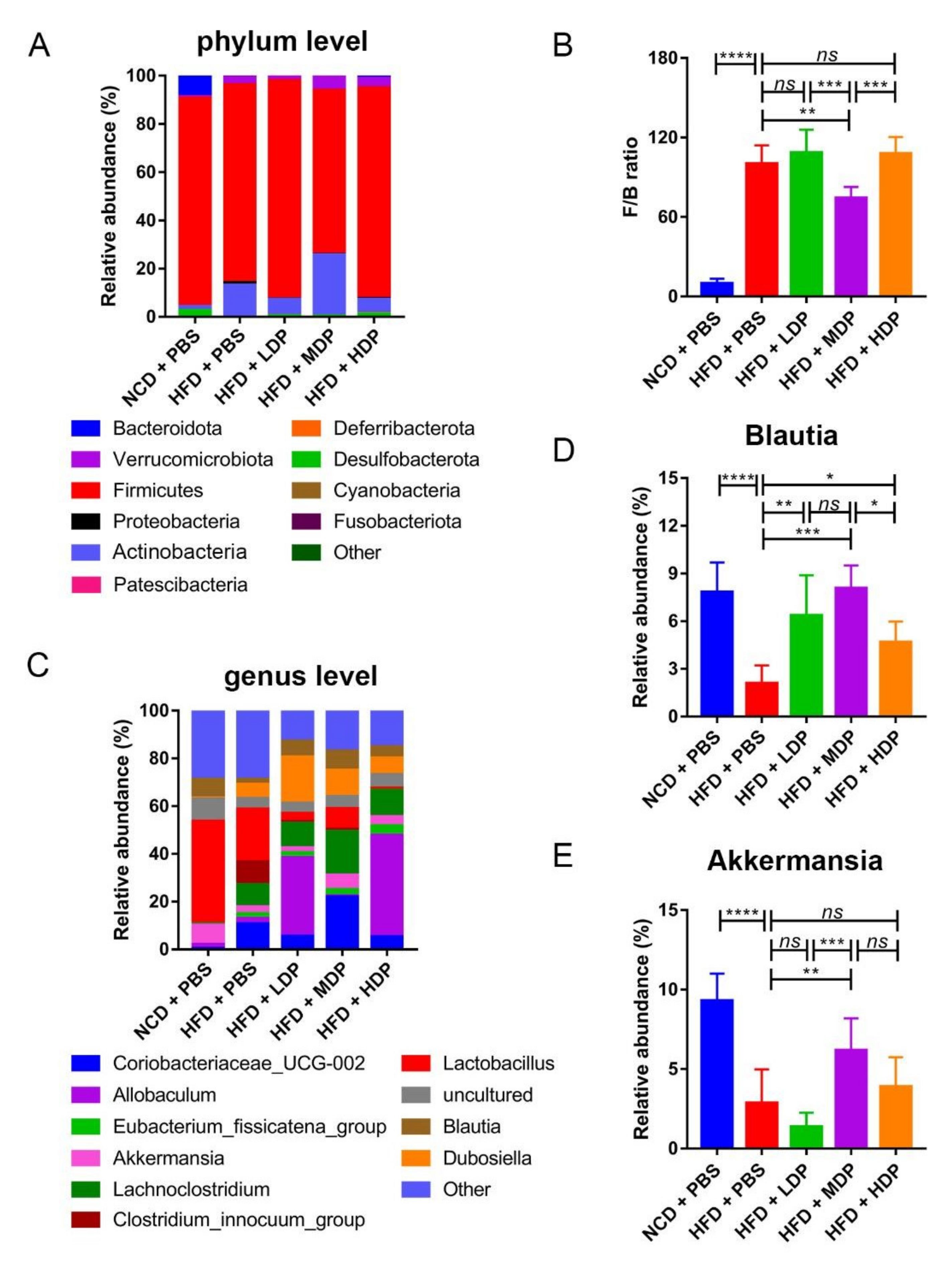
3.5. The Effect of DP Treatment on Gut Microbiota
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and Challenges in Translating the Biology of Atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Maruf, A.; Wang, Y.; Yin, T.; Huang, J.; Wang, N.; Durkan, C.; Tan, Y.; Wu, W.; Wang, G. Atherosclerosis Treatment with Stimuli-Responsive Nanoagents: Recent Advances and Future Perspectives. Adv. Healthc. Mater. 2019, 8, e1900036. [Google Scholar] [CrossRef] [PubMed]
- Tuñón, J.; Bäck, M.; Badimón, L.; Bochaton-Piallat, M.-L.; Cariou, B.; Daemen, M.J.; Egido, J.; Evans, P.C.; Francis, S.E.; Ketelhuth, D.F.; et al. Interplay between Hypercholesterolaemia and Inflammation in Atherosclerosis: Translating Experimental Targets into Clinical Practice. Eur. J. Prev. Cardiol. 2018, 25, 948–955. [Google Scholar] [CrossRef] [PubMed]
- Schwenger, K.J.P.; Arendt, B.M.; Smieja, M.; Ma, D.W.L.; Smaill, F.; Allard, J.P. Relationships between Atherosclerosis and Plasma Antioxidant Micronutrients or Red Blood Cell Polyunsaturated Fatty Acids in People Living with HIV. Nutrients 2019, 11, 1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, L.; Zhang, S.; Su, Q.; Li, S. Effects of Withdrawing an Atherogenic Diet on the Atherosclerotic Plaque in Rabbits. Exp. Ther. Med. 2021, 22, 751. [Google Scholar] [CrossRef]
- Torres, N.; Guevara-Cruz, M.; Velázquez-Villegas, L.A.; Tovar, A.R. Nutrition and Atherosclerosis. Arch. Med. Res. 2015, 46, 408–426. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Myasoedova, V.A.; Revin, V.V.; Orekhov, A.N.; Bobryshev, Y.V. The Phenomenon of Atherosclerosis Reversal and Regression: Lessons from Animal Models. Exp. Mol. Pathol. 2017, 102, 138–145. [Google Scholar] [CrossRef]
- Martínez-González, M.A.; Salas-Salvadó, J.; Estruch, R.; Corella, D.; Fitó, M.; Ros, E.; PREDIMED Investigators. Benefits of the Mediterranean Diet: Insights from the PREDIMED Study. Prog. Cardiovasc. Dis. 2015, 58, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Denysschen, C.A.; Burton, H.W.; Horvath, P.J.; Leddy, J.J.; Browne, R.W. Resistance Training with Soy vs. Whey Protein Supplements in Hyperlipidemic Males. J. Int. Soc. Sports Nutr. 2009, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Sucher, S.; Markova, M.; Hornemann, S.; Pivovarova, O.; Rudovich, N.; Thomann, R.; Schneeweiss, R.; Rohn, S.; Pfeiffer, A.F.H. Comparison of the Effects of Diets High in Animal or Plant Protein on Metabolic and Cardiovascular Markers in Type 2 Diabetes: A Randomized Clinical Trial: SUCHER et al. Diabetes Obes. Metab. 2017, 19, 944–952. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Yi, S.; Zhang, H.; Wang, J. Ingestion of Soy—Whey Blended Protein Augments Sports Performance and Ameliorates Exercise-Induced Fatigue in a Rat Exercise Model. Food Funct. 2017, 8, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhang, R.; Qin, Z.-Z.; Chen, J.-P.; Xu, J.-Y.; Qin, L.-Q. Effects of Chronic Whey Protein Supplementation on Atherosclerosis in ApoE−/− Mice. J. Nutr. Sci. Vitaminol. 2018, 64, 143–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sathyapalan, T.; Aye, M.; Rigby, A.S.; Thatcher, N.J.; Dargham, S.R.; Kilpatrick, E.S.; Atkin, S.L. Soy Isoflavones Improve Cardiovascular Disease Risk Markers in Women during the Early Menopause. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 691–697. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-T.; Luo, Y.; Zhang, Y.; Zhou, Y.; Zhao, T.-T. High Protein Diet Is of Benefit for Patients with Type 2 Diabetes: An Updated Meta-Analysis. Medicine 2018, 97, e13149. [Google Scholar] [CrossRef]
- Paik, J.K.; Park, M.; Shin, J.E.; Jang, S.-Y.; Shin, J.-Y. Dietary Protein to Carbohydrate Ratio and Incidence of Metabolic Syndrome in Korean Adults Based on a Long-Term Prospective Community-Based Cohort. Nutrients 2020, 12, 3274. [Google Scholar] [CrossRef]
- Van Nielen, M.; Feskens, E.J.M.; Mensink, M.; Sluijs, I.; Molina, E.; Amiano, P.; Ardanaz, E.; Balkau, B.; Beulens, J.W.J.; Boeing, H.; et al. Dietary Protein Intake and Incidence of Type 2 Diabetes in Europe: The EPIC-InterAct Case-Cohort Study. Diabetes Care 2014, 37, 1854–1862. [Google Scholar] [CrossRef] [Green Version]
- Kalantar-Zadeh, K.; Joshi, S.; Schlueter, R.; Cooke, J.; Brown-Tortorici, A.; Donnelly, M.; Schulman, S.; Lau, W.-L.; Rhee, C.M.; Streja, E.; et al. Plant-Dominant Low-Protein Diet for Conservative Management of Chronic Kidney Disease. Nutrients 2020, 12, 1931. [Google Scholar] [CrossRef]
- Stefíková, K.; Spustová, V.; Gazdíková, K.; Krivosíková, Z.; Dzúrik, R. Dietary Protein Restriction in Combination with Angiotensin Converting Enzyme Inhibitor Improves Insulin Resistance in Patients with Chronic Renal Disease. Int. Urol. Nephrol. 1997, 29, 497–507. [Google Scholar] [CrossRef]
- Mushenkova, N.V.; Summerhill, V.I.; Silaeva, Y.Y.; Deykin, A.V.; Orekhov, A.N. Modelling of Atherosclerosis in Genetically Modified Animals. Am. J. Transl. Res. 2019, 11, 4614–4633. [Google Scholar]
- Wang, C.; Wang, H.; Zhang, D.; Luo, W.; Liu, R.; Xu, D.; Diao, L.; Liao, L.; Liu, Z. Phosphorylation of ULK1 Affects Autophagosome Fusion and Links Chaperone-Mediated Autophagy to Macroautophagy. Nat. Commun. 2018, 9, 3492. [Google Scholar] [CrossRef]
- Din, A.U.; Hassan, A.; Zhu, Y.; Zhang, K.; Wang, Y.; Li, T.; Wang, Y.; Wang, G. Inhibitory Effect of Bifidobacterium Bifidum ATCC 29521 on Colitis and Its Mechanism. J. Nutr. Biochem. 2020, 79, 108353. [Google Scholar] [CrossRef]
- Hassan, A.; Din, A.U.; Zhu, Y.; Zhang, K.; Li, T.; Wang, Y.; Xu, S.; Lei, H.; Yu, X.; Wang, G. Anti-Atherosclerotic Effects of Lactobacillus Plantarum ATCC 14917 in ApoE−/− Mice through Modulation of Proinflammatory Cytokines and Oxidative Stress. Appl. Microbiol. Biotechnol. 2020, 104, 6337–6350. [Google Scholar] [CrossRef]
- Ulven, S.M.; Dalen, K.T.; Gustafsson, J.-A.; Nebb, H.I. Tissue-Specific Autoregulation of the LXRalpha Gene Facilitates Induction of ApoE in Mouse Adipose Tissue. J. Lipid Res. 2004, 45, 2052–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidah, N.G.; Awan, Z.; Chrétien, M.; Mbikay, M. PCSK9: A Key Modulator of Cardiovascular Health. Circ. Res. 2014, 114, 1022–1036. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, P.; Rengarajan, T.; Thangavel, J.; Nishigaki, Y.; Sakthisekaran, D.; Sethi, G.; Nishigaki, I. The Vascular Endothelium and Human Diseases. Int. J. Biol. Sci. 2013, 9, 1057–1069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Zhang, H.; Qi, W.; Zhang, Y.; Li, J.; Li, Z.; Lin, Y.; Bai, X.; Liu, X.; Chen, X.; et al. Nicotine Promotes Atherosclerosis via ROS-NLRP3-Mediated Endothelial Cell Pyroptosis. Cell Death Dis. 2018, 9, 171. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Cheng, H.; Liu, Y.; Xue, M.; Liang, H. Red Yeast Rice Ameliorates High-Fat Diet-Induced Atherosclerosis in Apoe−/− Mice in Association with Improved Inflammation and Altered Gut Microbiota Composition. Food Funct. 2019, 10, 3880–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia Muciniphila Protects Against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe −/− Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.-M.; et al. Gut Flora Metabolism of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 472, 57–63. [Google Scholar] [CrossRef] [Green Version]
- Lyu, M.; Wang, Y.-F.; Fan, G.-W.; Wang, X.-Y.; Xu, S.-Y.; Zhu, Y. Balancing Herbal Medicine and Functional Food for Prevention and Treatment of Cardiometabolic Diseases through Modulating Gut Microbiota. Front. Microbiol. 2017, 8, 2146. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D.; de Vos, W.M. Next-Generation Beneficial Microbes: The Case of Akkermansia Muciniphila. Front. Microbiol. 2017, 8, 1765. [Google Scholar] [CrossRef] [PubMed]
- Tomayko, E.J.; Kistler, B.M.; Fitschen, P.J.; Wilund, K.R. Intradialytic Protein Supplementation Reduces Inflammation and Improves Physical Function in Maintenance Hemodialysis Patients. J. Ren. Nutr. 2015, 25, 276–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, J.F.; Oya, V.; Coy, C.S.R.; Morcillo, A.M.; Severino, S.D.; Wu, C.; Sgarbieri, V.C.; dos Santos Vilela, M.M. Whey and Soy Protein Supplements Changes Body Composition in Patients with Crohn’s Disease Undergoing Azathioprine and Anti-TNF-Alpha Therapy. Nutr. Hosp. 2015, 31, 1603–1610. [Google Scholar] [CrossRef]
- Aparicio, V.A.; Sánchez, C.; Ortega, F.B.; Nebot, E.; Kapravelou, G.; Porres, J.M.; Aranda, P. Effects of the Dietary Amount and Source of Protein, Resistance Training and Anabolic-Androgenic Steroids on Body Weight and Lipid Profile of Rats. Nutr. Hosp. 2013, 28, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Mohr, R.; Boesecke, C.; Dold, L.; Schierwagen, R.; Schwarze-Zander, C.; Wasmuth, J.-C.; Weisensee, I.; Rockstroh, J.K.; Trebicka, J. Return-to-Health Effect of Modern Combined Antiretroviral Therapy Potentially Predisposes HIV Patients to Hepatic Steatosis. Medicine 2018, 97, e0462. [Google Scholar] [CrossRef]
- Oliveira, L.P.M.; de Jesus, R.P.; Boulhosa, R.S.S.B.; Mendes, C.M.C.; Gnoatto, M.C.; Lemaire, D.C.; Toralles, M.B.P.; Cavalcante, L.N.; Lyra, A.C.; Lyra, L.G.C. Effect of Soy Protein Supplementation in Patients with Chronic Hepatitis C: A Randomized Clinical Trial. World J. Gastroenterol. 2012, 18, 2203–2211. [Google Scholar] [CrossRef]
- Bortolotti, M.; Maiolo, E.; Corazza, M.; Van Dijke, E.; Schneiter, P.; Boss, A.; Carrel, G.; Giusti, V.; Lê, K.-A.; Quo Chong, D.G.; et al. Effects of a Whey Protein Supplementation on Intrahepatocellular Lipids in Obese Female Patients. Clin. Nutr. 2011, 30, 494–498. [Google Scholar] [CrossRef]
- Gregersen, S.; Bystrup, S.; Overgaard, A.; Jeppesen, P.B.; Sønderstgaard Thorup, A.C.; Jensen, E.; Hermansen, K. Effects of Whey Proteins on Glucose Metabolism in Normal Wistar Rats and Zucker Diabetic Fatty (ZDF) Rats. Rev. Diabet. Stud. 2013, 10, 252–269. [Google Scholar] [CrossRef] [Green Version]
- Fekete, Á.A.; Giromini, C.; Chatzidiakou, Y.; Givens, D.I.; Lovegrove, J.A. Whey Protein Lowers Blood Pressure and Improves Endothelial Function and Lipid Biomarkers in Adults with Prehypertension and Mild Hypertension: Results from the Chronic Whey2Go Randomized Controlled Trial. Am. J. Clin. Nutr. 2016, 104, 1534–1544. [Google Scholar] [CrossRef]
- Zhang, H.; Bartley, G.E.; Mitchell, C.R.; Zhang, H.; Yokoyama, W. Lower Weight Gain and Hepatic Lipid Content in Hamsters Fed High Fat Diets Supplemented with White Rice Protein, Brown Rice Protein, Soy Protein, and Their Hydrolysates. J. Agric. Food Chem. 2011, 59, 10927–10933. [Google Scholar] [CrossRef]
- Libby, P. Inflammation in Atherosclerosis. Nature 2002, 420, 868–874. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Jiao, L.; Cao, S.; Song, Z.; Hu, C.; Han, X. Whey Protein Concentrate Enhances Intestinal Integrity and Influences Transforming Growth Factor-Β1 and Mitogen-Activated Protein Kinase Signalling Pathways in Piglets after Lipopolysaccharide Challenge. Br. J. Nutr. 2016, 115, 984–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, M.E.V.; Phillipson, M.; Petersson, J.; Velcich, A.; Holm, L.; Hansson, G.C. The Inner of the Two Muc2 Mucin-Dependent Mucus Layers in Colon Is Devoid of Bacteria. Proc. Natl. Acad. Sci. USA 2008, 105, 15064–15069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, J.R. Intestinal Mucosal Barrier Function in Health and Disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Hassan, A.; Din, A.U.; Zhu, Y.; Zhang, K.; Li, T.; Wang, Y.; Luo, Y.; Wang, G. Updates in Understanding the Hypocholesterolemia Effect of Probiotics on Atherosclerosis. Appl. Microbiol. Biotechnol. 2019, 103, 5993–6006. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Zhang, J.; Li, M.; Tang, Z.; Yang, Z.; Cheng, G.; Wang, J. Gut Microbiota Composition Influences Outcomes of Skeletal Muscle Nutritional Intervention via Blended Protein Supplementation in Posttransplant Patients with Hematological Malignancies. Clin. Nutr. 2021, 40, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Marques, F.Z.; Nelson, E.; Chu, P.-Y.; Horlock, D.; Fiedler, A.; Ziemann, M.; Tan, J.K.; Kuruppu, S.; Rajapakse, N.W.; El-Osta, A.; et al. High-Fiber Diet and Acetate Supplementation Change the Gut Microbiota and Prevent the Development of Hypertension and Heart Failure in Hypertensive Mice. Circulation 2017, 135, 964–977. [Google Scholar] [CrossRef]
- Xie, D.; Jiang, L.; Lin, Y.; Liu, Z. Antioxidant Activity of Selenium-Enriched Chrysomyia Megacephala (Fabricius) Larvae Powder and Its Impact on Intestinal Microflora in D-Galactose Induced Aging Mice. BMC Complement. Med. Ther. 2020, 20, 264. [Google Scholar] [CrossRef]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics Improve Gut Microbiota Dysbiosis in Obese Mice Fed a High-Fat or High-Sucrose Diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef]
- Martignoni, M.; Groothuis, G.M.M.; de Kanter, R. Species Differences between Mouse, Rat, Dog, Monkey and Human CYP-Mediated Drug Metabolism, Inhibition and Induction. Expert Opin. Drug Metab. Toxicol. 2006, 2, 875–894. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (Sense 5′–3′) | Primer Sequence (Antisense 5′–3′) |
|---|---|---|
| LXRα | GCGCTTTGCCCACTTTACTG | TCTTCAGCAAGGCGATCTGG |
| ABCA1 | TGGGCTCCTCCCTGTTTTTG | GTCACTTTCATGGTCGCTGC |
| ABCG1 | CTACGGCTTGGACCGAGAAG | ACCTCTCAGCCCGGATTTTG |
| Cyp7a1 | CTGGGGGATTGCTGTGGTAG | GCACAGCCCAGGTATGGAAT |
| Cyp27a1 | AGGCACAGGAGAGTACGGAG | AAGTCCCAAAGGAGGTTGTCC |
| PCSK9 | ATCACCGACTTCAACAGCGT | GCCCTTCCCTTGACAGTTGA |
| SREBP1 | GCCATCGACTACATCCGCTT | GTCTCCACCACTTCGGGTTT |
| SREBP2 | GGCGATGAGCTGACTCTCGG | CCTCCAGGGAAGGAGCTACAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.; Zhang, K.; Zhang, L.; Qiu, J.; Fu, L.; Yin, T.; Wang, J.; Qin, R.; Zhang, J.; Dong, X.; et al. Dosage of Dual-Protein Nutrition Differentially Impacts the Formation of Atherosclerosis in ApoE−/− Mice. Nutrients 2022, 14, 855. https://doi.org/10.3390/nu14040855
Huang Y, Zhang K, Zhang L, Qiu J, Fu L, Yin T, Wang J, Qin R, Zhang J, Dong X, et al. Dosage of Dual-Protein Nutrition Differentially Impacts the Formation of Atherosclerosis in ApoE−/− Mice. Nutrients. 2022; 14(4):855. https://doi.org/10.3390/nu14040855
Chicago/Turabian StyleHuang, Yingchun, Kun Zhang, Li Zhang, Juhui Qiu, Lin Fu, Tieying Yin, Jing Wang, Rui Qin, Jingjie Zhang, Xianwen Dong, and et al. 2022. "Dosage of Dual-Protein Nutrition Differentially Impacts the Formation of Atherosclerosis in ApoE−/− Mice" Nutrients 14, no. 4: 855. https://doi.org/10.3390/nu14040855
APA StyleHuang, Y., Zhang, K., Zhang, L., Qiu, J., Fu, L., Yin, T., Wang, J., Qin, R., Zhang, J., Dong, X., & Wang, G. (2022). Dosage of Dual-Protein Nutrition Differentially Impacts the Formation of Atherosclerosis in ApoE−/− Mice. Nutrients, 14(4), 855. https://doi.org/10.3390/nu14040855