
Route of Sensitization to Peanut Influences Immune Cell Recruitment at Various Mucosal Sites in Mouse: An Integrative Analysis

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Allergens
2.2. Mice
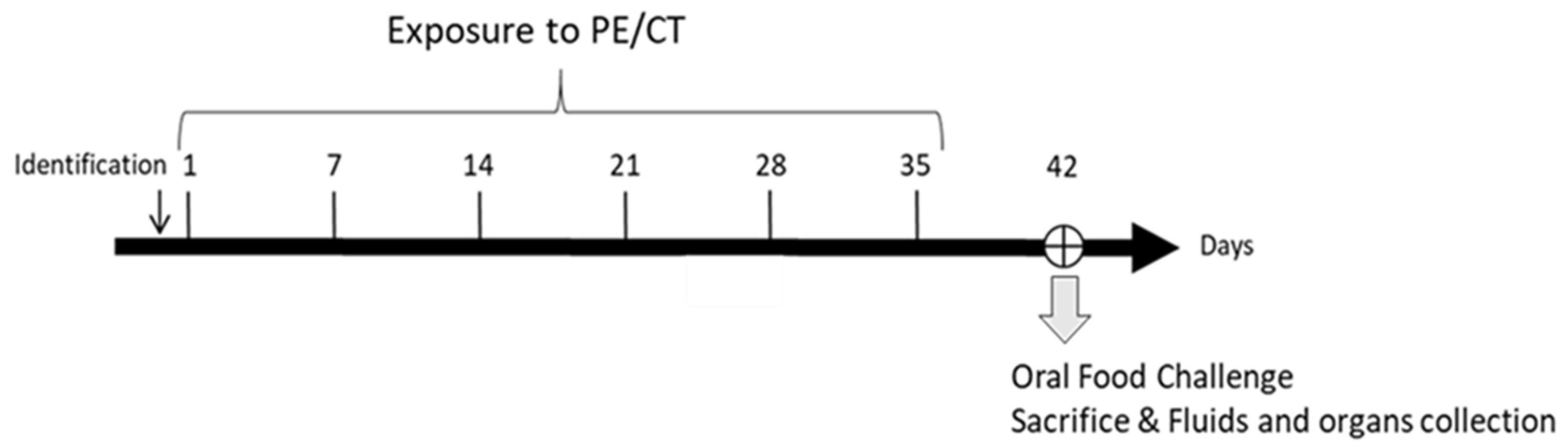
2.3. Sensitization
2.4. Oral Food Challenge (OFC)
2.5. Single Cell Preparations from Collected Organs
2.6. mMCP1 Assay
2.7. Measurement of PE-Specific IgE and IgG1 in Plasma
2.8. Local and Circulating Cytokine Analysis
2.9. Cellular Population Analysis
2.10. Statistical Analysis
3. Results
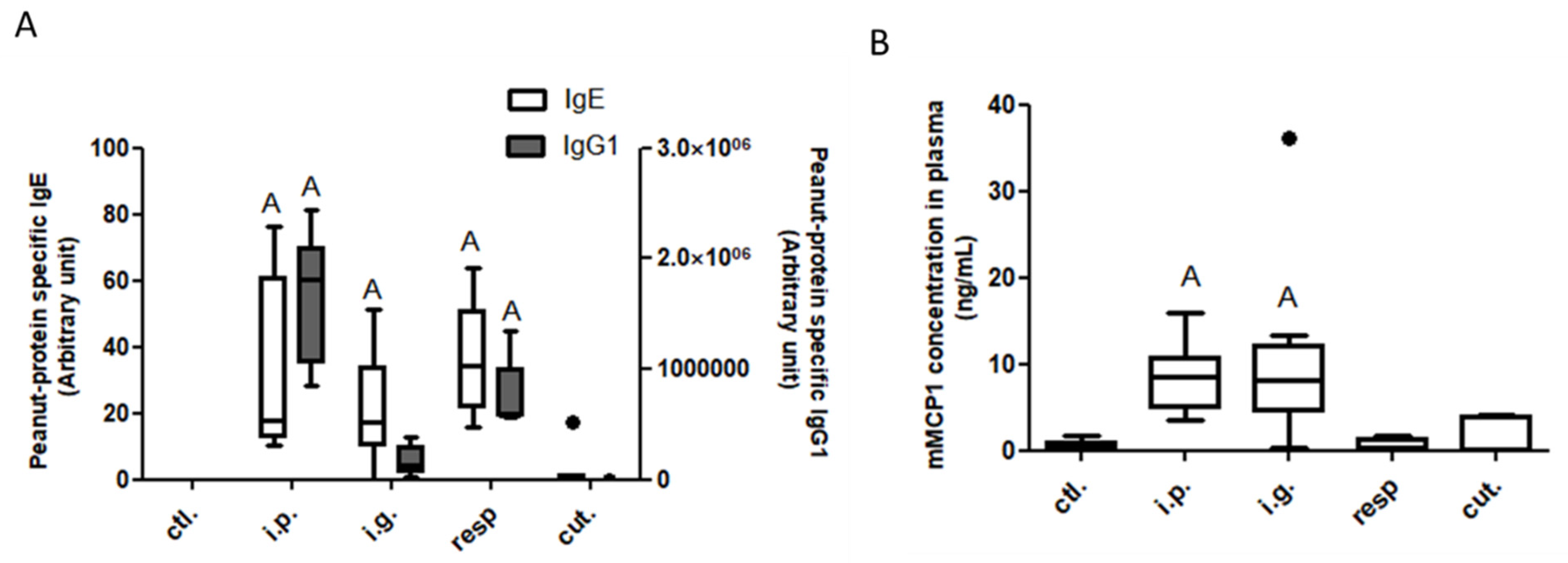
3.1. Assessment of Food Allergy Induction
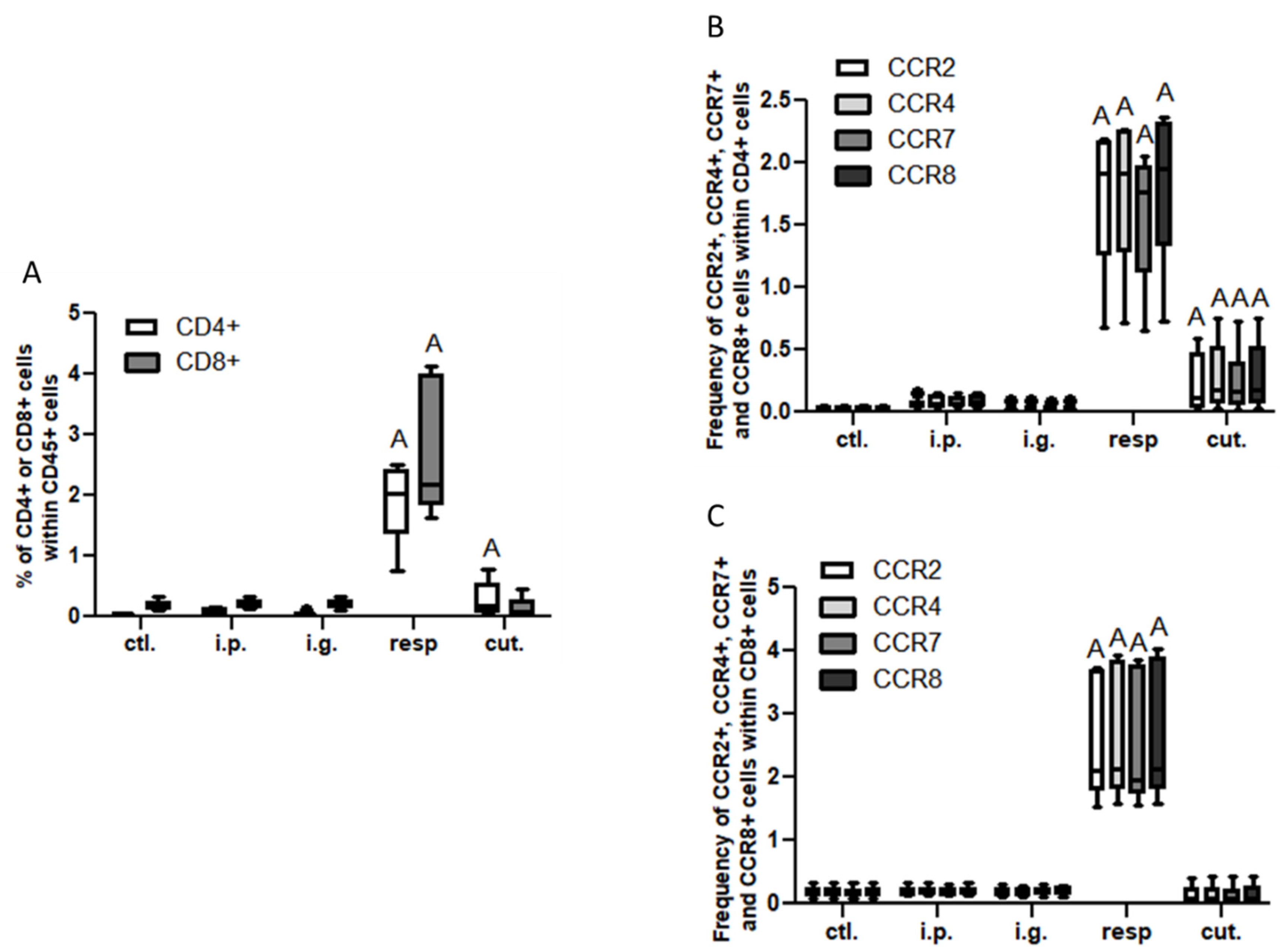
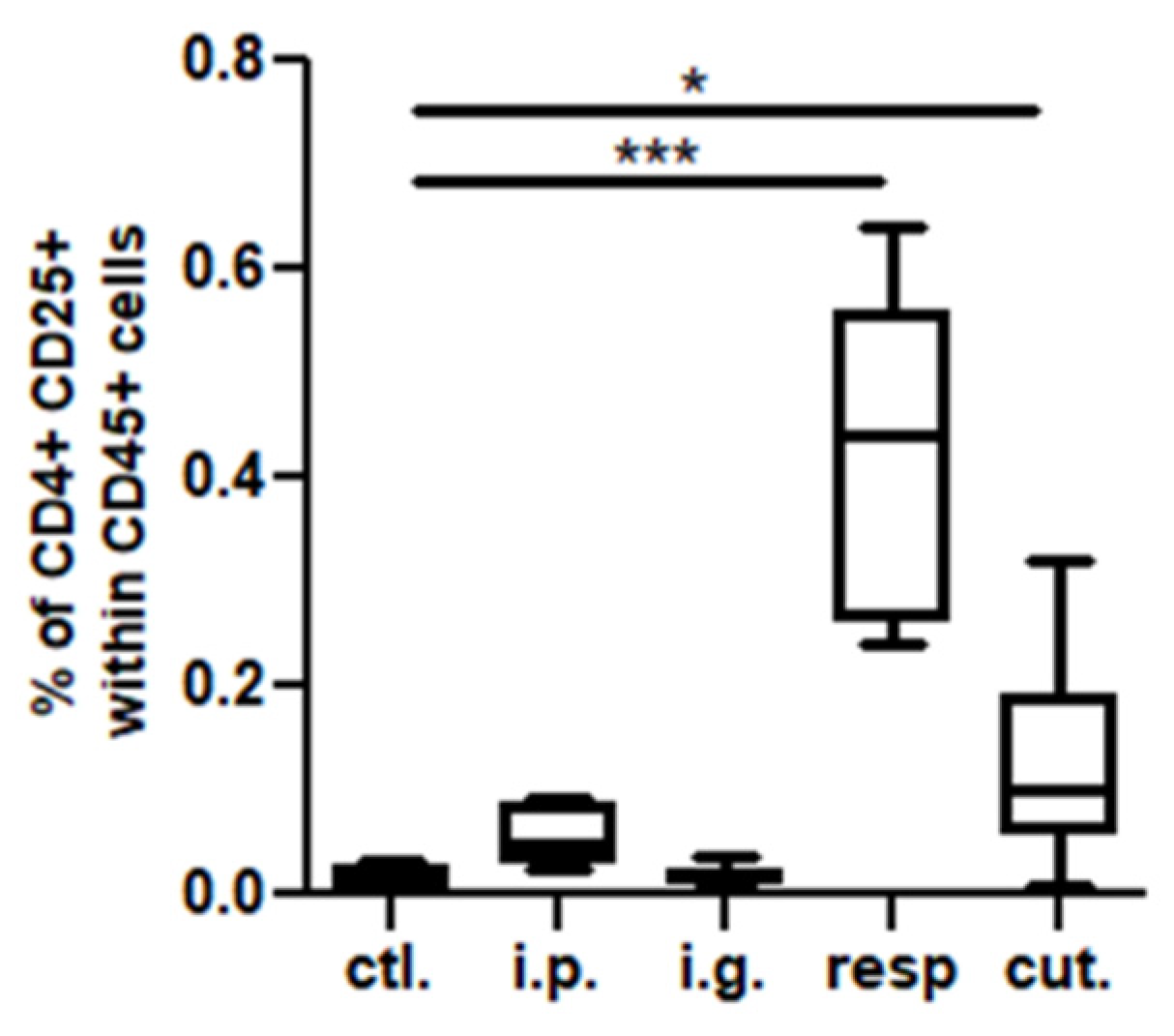
3.2. Analysis of Immune Cellular Components in Tissues, Lymph Nodes, and BAL Fluids
3.2.1. Non-Supervised Analysis
3.2.2. Multivariate and Univariate Supervised Analysis
Data Modelling and Identification of Discriminant and Significant Variables
Analysis in Intestinal Tissue and Associated Lymph Nodes
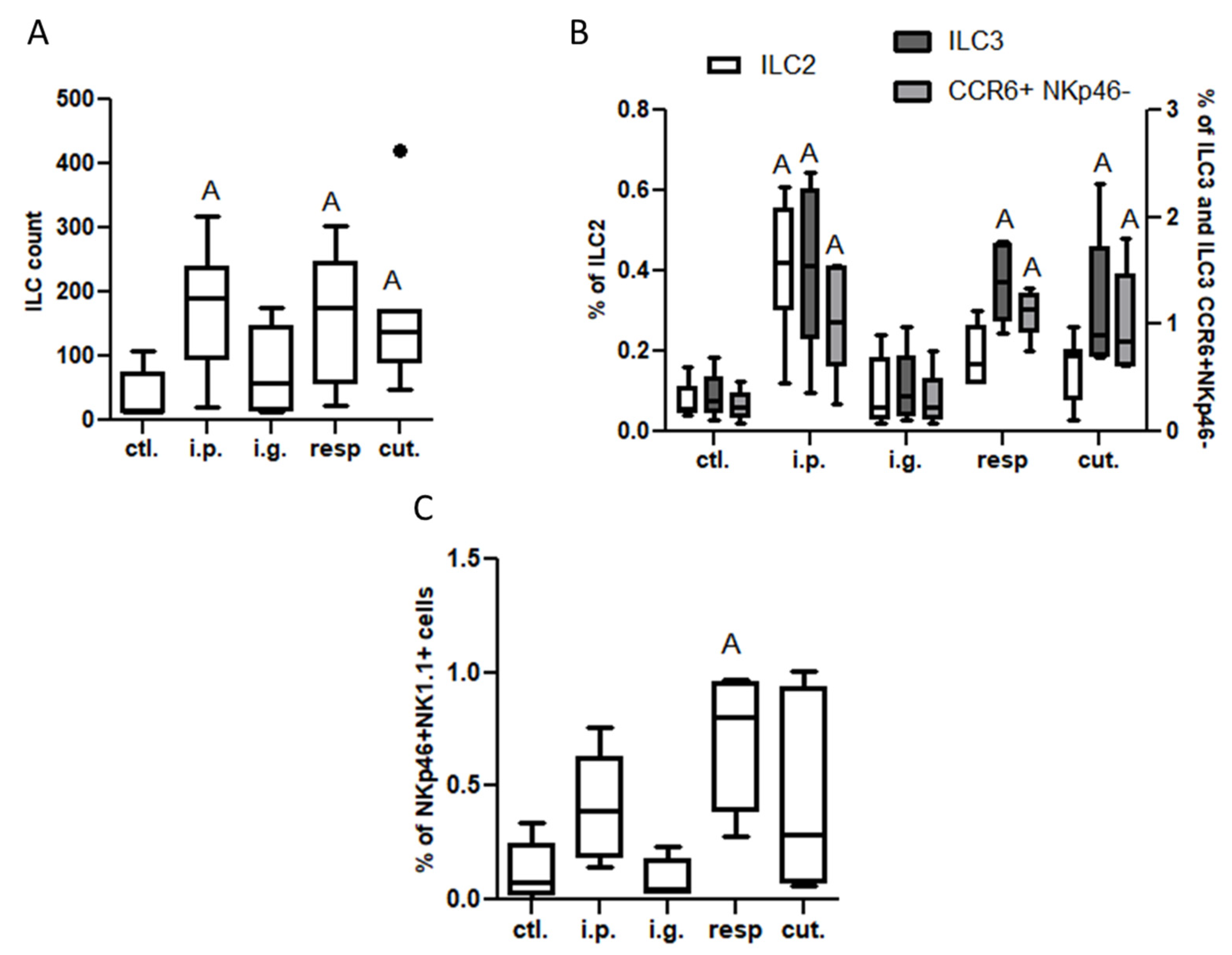
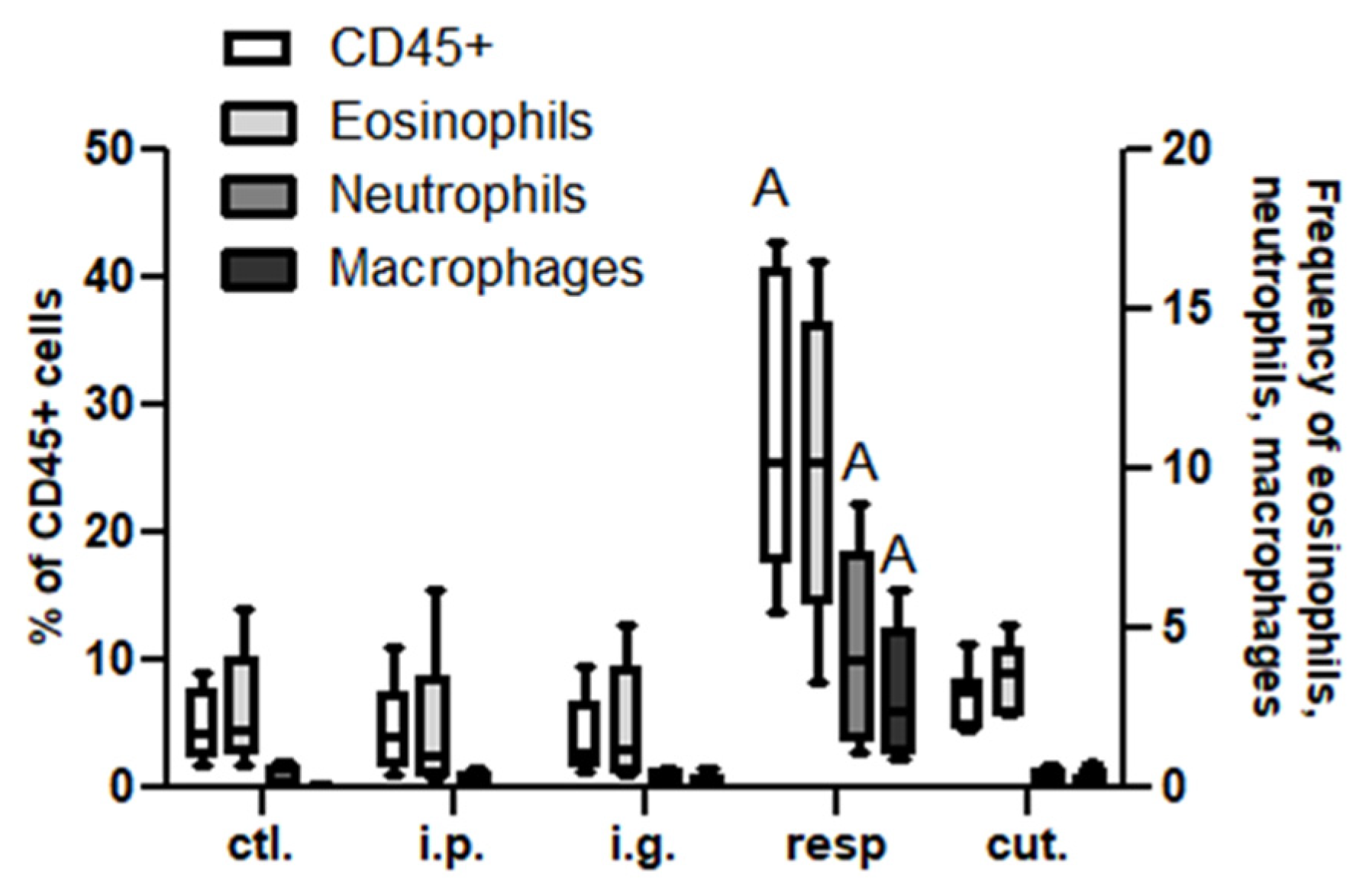
Analysis in the Respiratory Tract (BAL and Lung Tissue) and Associated Lymph Nodes (MedLN)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valenta, R.; Hochwallner, H.; Linhart, B.; Pahr, S. Food Allergies: The Basics. Gastroenterology 2015, 148, 1120–1131.e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wijk, F.; Knippels, L. Initiating Mechanisms of Food Allergy: Oral Tolerance versus Allergic Sensitization. Biomed. Pharmacother. 2007, 61, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Hourihane, J.O.; Kilburn, S.A.; Dean, P.; Warner, J.O. Clinical Characteristics of Peanut Allergy. Clin. Exp. Allergy 1997, 27, 634–639. [Google Scholar] [CrossRef] [PubMed]
- Lack, G. Update on Risk Factors for Food Allergy. J. Allergy Clin. Immunol. 2012, 129, 1187–1197. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, D.A.; Bahna, S.L. Food Hypersensitivity by Inhalation. Clin. Mol. Allergy 2009, 7, 4. [Google Scholar] [CrossRef] [Green Version]
- Allen, K.J.; Dharmage, S.C. The Role of Food Allergy in the Atopic March: Role of Food Allergy in the Atopic March. Clin. Exp. Allergy 2010, 40, 1439–1441. [Google Scholar] [CrossRef]
- Benhamou, A.H.; Eigenmann, P.A. Dermatite atopique et allergies alimentaires. Rev. Med. Suisse 2007, 3, 1038–1043. [Google Scholar]
- Cianferoni, A.; Spergel, J.M. Food Allergy: Review, Classification and Diagnosis. Allergol. Int. 2009, 58, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Cesta, M.F. Normal Structure, Function, and Histology of Mucosa-Associated Lymphoid Tissue. Toxicol. Pathol. 2006, 34, 599–608. [Google Scholar] [CrossRef] [Green Version]
- Wavrin, S.; Bernard, H.; Wal, J.-M.; Adel-Patient, K. Cutaneous or Respiratory Exposures to Peanut Allergens in Mice and Their Impacts on Subsequent Oral Exposure. Int. Arch. Allergy Immunol. 2014, 164, 189–199. [Google Scholar] [CrossRef]
- Jong, D.E.; Van Zijverden; Spanhaak; Koppelman; Pellegrom; Pennink. Identification and partial characterization of multiple major allergens in peanut proteins. Clin. Exp. Allergy 1998, 28, 743–751. [Google Scholar] [CrossRef]
- Adel-Patient, K.; Bernard, H.; Ah-Leung, S.; Creminon, C.; Wal, J.-M. Peanut- and Cow’s Milk-Specific IgE, Th2 Cells and Local Anaphylactic Reaction Are Induced in Balb/c Mice Orally Sensitized with Cholera Toxin. Allergy 2005, 60, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Adel-Patient, K.; Nahori, M.-A.; Proust, B.; Lapa e Silva, J.R.; Créminon, C.; Wal, J.-M.; Vargaftig, B.B. Elicitation of the Allergic Reaction in β-Lactoglobulin-Sensitized Balb/c Mice: Biochemical and Clinical Manifestations Differ According to the Structure of the Allergen Used for Challenge: Induction and Optimization of Anti-BLG Responses in Balb/c Mice. Clin. Exp. Allergy 2003, 33, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Adel-Patient, K.; Guinot, M.; Guillon, B.; Bernard, H.; Chikhi, A.; Hazebrouck, S.; Junot, C. Administration of Extensive Hydrolysates from Caseins and Lactobacillus Rhamnosus GG Probiotic Does Not Prevent Cow’s Milk Proteins Allergy in a Mouse Model. Front. Immunol. 2020, 11, 1700. [Google Scholar] [CrossRef] [PubMed]
- Adel-Patient, K.; Creminon, C.; Bernard, H.; Clement, G.; Negroni, L.; Frobert, Y.; Grassi, J.; Wal, J.-M.; Chatel, J.-M. Evaluation of a High IgE-Responder Mouse Model of Allergy to Bovine b-Lactoglobulin ŽBLG/: Development of Sandwich Immunoassays for Total and Allergen-Specific IgE, IgG1 and IgG2a in BLG-Sensitized Mice. J. Immunol. Methods 2000, 235, 21–32. [Google Scholar] [CrossRef]
- Bernard, H.; Drumare, M.-F.; Guillon, B.; Paty, E.; Scheinmann, P.; Wal, J.-M. Immunochemical Characterisation of Structure and Allergenicity of Peanut 2S Albumins Using Different Formats of Immunoassays. Anal. Bioanal. Chem. 2009, 395, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Brough, H.A.; Makinson, K.; Penagos, M.; Maleki, S.J.; Cheng, H.; Douiri, A.; Stephens, A.C.; Turcanu, V.; Lack, G. Distribution of Peanut Protein in the Home Environment. J. Allergy Clin. Immunol. 2013, 132, 623–629. [Google Scholar] [CrossRef]
- Hamad, A.; Burks, W. Oral Tolerance and Allergy. Semin. Immunol. 2017, 30, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-M.; Serebrisky, D.; Lee, S.-Y.; Huang, C.-K.; Bardina, L.; Schofield, B.H.; Stanley, J.S.; Burks, A.W.; Bannon, G.A.; Sampson, H.A. A Murine Model of Peanut Anaphylaxis: T- and B-Cell Responses to a Major Peanut Allergen Mimic Human Responses. J. Allergy Clin. Immunol. 2000, 106, 150–158. [Google Scholar] [CrossRef]
- Strid, J.; Hourihane, J.; Kimber, I.; Callard, R.; Strobel, S. Epicutaneous Exposure to Peanut Protein Prevents Oral Tolerance and Enhances Allergic Sensitization. Clin. Htmlent Glyphamp Asciiamp Exp. Allergy 2005, 35, 757–766. [Google Scholar] [CrossRef]
- Li, W.; Zhang, Z.; Saxon, A.; Zhang, K. Prevention of Oral Food Allergy Sensitization via Skin Application of Food Allergen in a Mouse Model. Allergy 2012, 67, 622–629. [Google Scholar] [CrossRef] [PubMed]
- Adel-Patient, K.; Wavrin, S.; Bernard, H.; Meziti, N.; Ah-Leung, S.; Wal, J.-M. Oral Tolerance and Treg Cells Are Induced in BALB/c Mice after Gavage with Bovine β-Lactoglobulin: Oral Tolerance and Treg Cells Inductions in BALB/c Mice. Allergy 2011, 66, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Besnard, A.-G.; Togbe, D.; Couillin, I.; Tan, Z.; Zheng, S.G.; Erard, F.; Le Bert, M.; Quesniaux, V.; Ryffel, B. Inflammasome-IL-1-Th17 Response in Allergic Lung Inflammation. J. Mol. Cell Biol. 2012, 4, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-H.; Liu, Y.-J. The IL-17 Cytokine Family and Their Role in Allergic Inflammation. Curr. Opin. Immunol. 2008, 20, 697–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouchaud, G.; Gourbeyre, P.; Bihouée, T.; Aubert, P.; Lair, D.; Cheminant, M.-A.; Denery-Papini, S.; Neunlist, M.; Magnan, A.; Bodinier, M. Consecutive Food and Respiratory Allergies Amplify Systemic and Gut but Not Lung Outcomes in Mice. J. Agric. Food Chem. 2015, 63, 6475–6483. [Google Scholar] [CrossRef]
- Bihouée, T.; Bouchaud, G.; Chesné, J.; Lair, D.; Rolland-Debord, C.; Braza, F.; Cheminant, M.-A.; Aubert, P.; Mahay, G.; Sagan, C.; et al. Food Allergy Enhances Allergic Asthma in Mice. Respir. Res. 2014, 15, 142. [Google Scholar] [CrossRef] [Green Version]
- Castan, L.; Cheminant, M.-A.; Colas, L.; Brouard, S.; Magnan, A.; Bouchaud, G. Food Allergen-Sensitized CCR9+ Lymphocytes Enhance Airways Allergic Inflammation in Mice. Allergy 2018, 73, 1505–1514. [Google Scholar] [CrossRef]
- Dharmage, S.C.; Lowe, A.J.; Matheson, M.C.; Burgess, J.A.; Allen, K.J.; Abramson, M.J. Atopic Dermatitis and the Atopic March Revisited. Allergy 2014, 69, 17–27. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briard, M.; Guinot, M.; Grauso, M.; Guillon, B.; Hazebrouck, S.; Bernard, H.; Bouchaud, G.; Michel, M.-L.; Adel-Patient, K. Route of Sensitization to Peanut Influences Immune Cell Recruitment at Various Mucosal Sites in Mouse: An Integrative Analysis. Nutrients 2022, 14, 790. https://doi.org/10.3390/nu14040790
Briard M, Guinot M, Grauso M, Guillon B, Hazebrouck S, Bernard H, Bouchaud G, Michel M-L, Adel-Patient K. Route of Sensitization to Peanut Influences Immune Cell Recruitment at Various Mucosal Sites in Mouse: An Integrative Analysis. Nutrients. 2022; 14(4):790. https://doi.org/10.3390/nu14040790
Chicago/Turabian StyleBriard, Mélanie, Marine Guinot, Marta Grauso, Blanche Guillon, Stéphane Hazebrouck, Hervé Bernard, Grégory Bouchaud, Marie-Laure Michel, and Karine Adel-Patient. 2022. "Route of Sensitization to Peanut Influences Immune Cell Recruitment at Various Mucosal Sites in Mouse: An Integrative Analysis" Nutrients 14, no. 4: 790. https://doi.org/10.3390/nu14040790
APA StyleBriard, M., Guinot, M., Grauso, M., Guillon, B., Hazebrouck, S., Bernard, H., Bouchaud, G., Michel, M.-L., & Adel-Patient, K. (2022). Route of Sensitization to Peanut Influences Immune Cell Recruitment at Various Mucosal Sites in Mouse: An Integrative Analysis. Nutrients, 14(4), 790. https://doi.org/10.3390/nu14040790