
Dietary Carotenoids in Head and Neck Cancer—Molecular and Clinical Implications


Abstract
1. Introduction
2. Material and Methods
3. Results
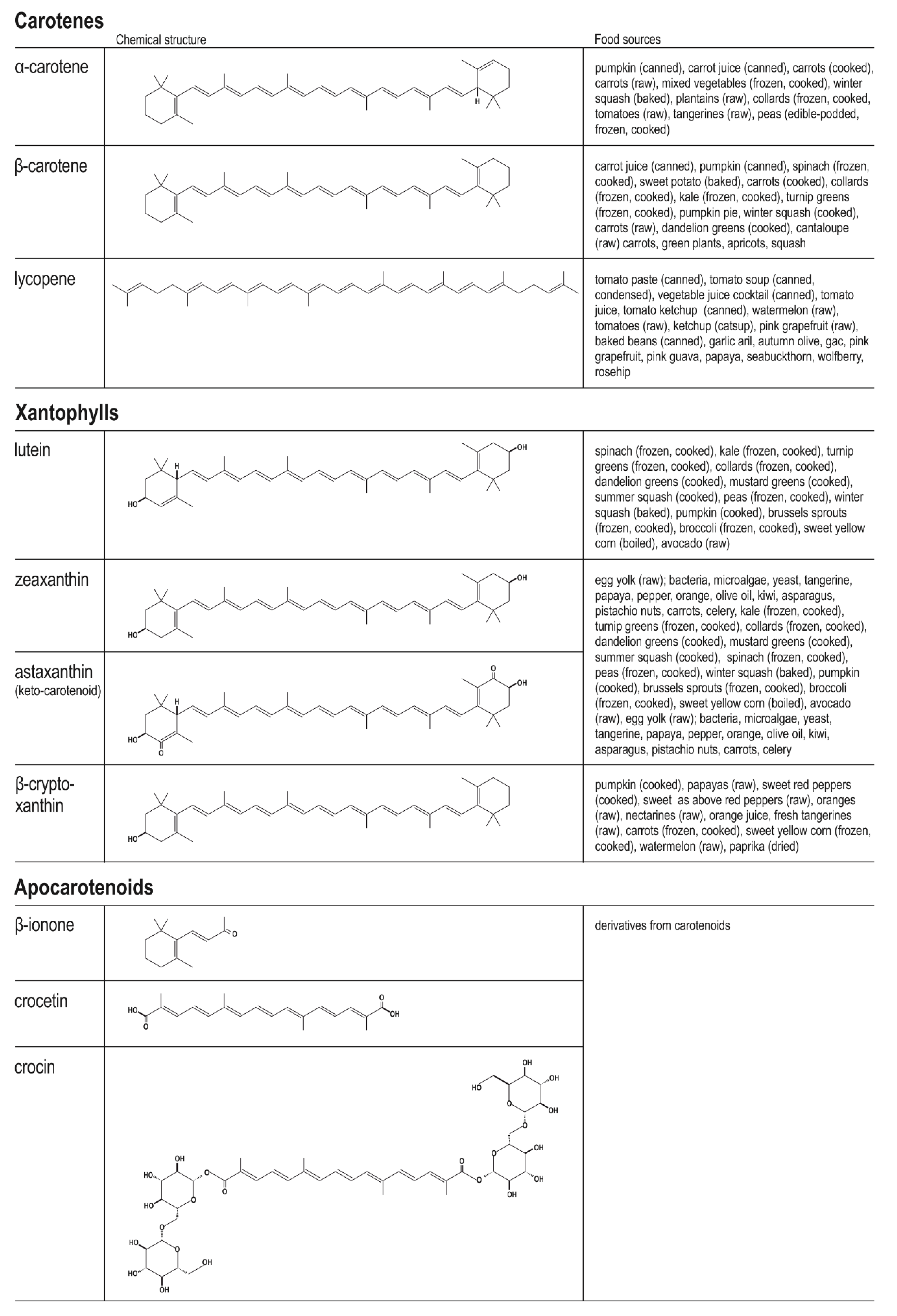
3.1. Chemistry of Dietary Carotenoids and Their Main Food Sources and Bioavailability
3.2. The Molecular Role of Carotenoids in Mechanisms Responsible for Carcinogenesis
3.2.1. Molecular Aspects of Head and Neck Carcinogenesis
3.2.2. Molecular Role of Carotenoids in Major Intracellular Oncogenic Signalling
3.2.3. The Molecular Function of Carotenoids in Apoptotic Signalling and Cell Cycle Progression
3.2.4. The Molecular Role of Carotenoids in Angiogenesis and Cancer Metastasis
3.3. Animal and In Vitro Models of Head and Neck Squamous Cell Cancer
3.3.1. Animal Models of HNSCC
3.3.2. In Vitro Models of HNSCC
3.4. Studies on the Chemopreventive Efficacy of Dietary Carotenoids in Clinical Trials on Healthy Individuals and Those with Premalignant Conditions
3.4.1. Chemopreventive Efficacy of Dietary Carotenoids on Healthy Individuals
3.4.2. Chemopreventive Efficacy of Dietary Carotenoids on Individuals Having Premalignant Conditions
3.5. Studies on the Role of Dietary Carotenoids as Predictors of Cancer Risk, Progression, and Prognosis in Patients with Diagnosed Head and Neck Cancer
3.5.1. Carotenoid Intake and Head and Neck Cancer
3.5.2. Serum Pretreatment and Posttreatment Carotenoid Concentrations in Head and Neck Cancer Risk and as Predictors of Survival and Recurrence
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Charap, A.J.; Enokida, T.; Brody, R.; Sfakianos, J.; Miles, B.; Bhardwaj, N.; Horowitz, A. Landscape of natural killer cell activity in head and neck squamous cell carcinoma. J. Immunother. Cancer 2020, 8, e001523. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, A.K.; Chan, C.J.; Grandis, J.R.; Takata, T.; Slootweg, P.J. WHO Classification of Head and Neck Tumours, 4th ed.; IARC: Lyon, France, 2017; ISBN 9789283224389. [Google Scholar]
- The Cancer Genome Atlas Network. Comprehensive genomic characterization of head and neck squamous cell carcinomas. Nature 2015, 517, 576–582. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2019, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef]
- Bhat, G.R.; Hyole, R.G.; Li, J. Head and neck cancer: Current challenges and future perspectives. Adv. Cancer Res. 2021, 152, 67–102. [Google Scholar] [CrossRef]
- Auperin, A. Epidemiology of head and neck cancers. Curr. Opin. Oncol. 2020, 32, 178–186. [Google Scholar] [CrossRef]
- Di Credico, G.; Edefonti, V.; Polesel, J.; Pauli, F.; Torelli, N.; Serraino, D.; Negri, E.; Luce, D.; Stucker, I.; Matsuo, K.; et al. Joint effects of intensity and duration of cigarette smoking on the risk of head and neck cancer: A bivariate spline model approach. Oral Oncol. 2019, 94, 47–57. [Google Scholar] [CrossRef]
- Sakthivel, P.; Samy, K.; Panda, S.; Singh, C.A. 14 “S” in head and neck cancers. Oral Oncol. 2021, 112, 105070. [Google Scholar] [CrossRef]
- Miranda-Galvis, M.; Loveless, R.; Kowalski, L.P.; Teng, Y. Impacts of Environmental Factors on Head and Neck Cancer Pathogenesis and Progression. Cells 2021, 10, 389. [Google Scholar] [CrossRef]
- Nichols, A.C.; Lang, P.; Prisman, E.; Berthelet, E.; Tran, E.; Hamilton, S.; Wu, J.; Fung, K.; de Almeida, J.R.; Bayley, A.; et al. Treatment de-escalation for HPV-associated oropharyngeal squamous cell carcinoma with radiotherapy vs. trans-oral surgery (ORATOR2): Study protocol for a randomized phase II trial. BMC Cancer 2020, 20, 125. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.B.; Greene, F.L.; Edge, S.B.; Compton, C.C.; Gershenwald, J.E.; Brookland, R.K.; Meyer, L.; Gress, D.M.; Byrd, D.R.; Winchester, D.P. The eighth edition AJCC cancer staging manual: Continuing to build a bridge from a population-based to a more “personalized” approach to cancer staging. CA Cancer J. Clin. 2017, 67, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Bravi, F.; Lee, Y.-C.A.; Hashibe, M.; Boffetta, P.; Conway, D.I.; Ferraroni, M.; La Vecchia, C.; Edefonti, V. Lessons learned from the INHANCE consortium: An overview of recent results on head and neck cancer. Rev. Oral Dis. 2021, 27, 73–93. [Google Scholar] [CrossRef] [PubMed]
- Chuang, S.C.; Jenab, M.; Heck, J.E.; Bosetti, C.; Talamini, R.; Matsuo, K.; Castellsague, X.; Franceschi, S.; Herrero, R.; Winn, D.M.; et al. Diet and the Risk of Head and Neck Cancer: A Pooled Analysis in the INHANCE Consortium. Cancer Causes Control. 2012, 23, 69–88. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, W.T.; Lee, Y.C.; Huang, C.C.; Ou, C.Y.; Lin, Y.H.; Huang, J.S.; Wong, T.Y.; Chen, K.C.; Hsiao, J.R.; et al. Investigating the association between diet and risk of head and neck cancer in Taiwan. Oncotarget 2017, 8, 98865–98875. [Google Scholar] [CrossRef]
- De Podesta, O.P.G.; Peres, S.V.; Salaroli, L.B.; Cattafesta, M.; De Podestá, J.R.V.; von Zeidler, S.L.V.; de Oliveira, J.C.; Kowalski, L.P.; Ikeda, M.K.; Brennan, P.; et al. Consumption of minimally processed foods as protective factors in the genesis of squamous cell carcinoma of the head and neck in Brazil. PLoS ONE 2019, 14, e0220067. [Google Scholar] [CrossRef]
- Lin, J.; Cook, N.R.; Albert, C.; Zaharris, E.; Gaziano, J.M.; Van Denburgh, M.; Buring, J.E.; Manson, J.E. Vitamins C and E and beta carotene supplementation and cancer risk: A randomized controlled trial. J. Natl. Cancer Inst. 2009, 101, 14–23. [Google Scholar] [CrossRef]
- Wright, M.E.; Virtamo, J.; Hartman, A.M.; Pietinen, P.; Edwards, B.K.; Taylor, P.R.; Huttunen, J.K.; Albanes, D. Effects of alpha-tocopherol and beta-carotene supplementation on upper aerodigestive tract cancers in a large, randomized controlled trial. Cancer 2007, 109, 891–898. [Google Scholar] [CrossRef]
- Key, T.J.; Bradbury, K.E.; Perez-Cornago, A.; Sinha, R.; Tsilidis, K.K.; Tsugane, S. Diet, nutrition, and cancer risk: What do we know and what is the way forward? BMJ 2020, 368, m511. [Google Scholar] [CrossRef]
- GBD 2017 Diet Collaborators. Health effects of dietary risks in 195 countries, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet 2019, 393, 1958–1972. [Google Scholar] [CrossRef]
- World Cancer Research Fund/American Institute for Cancer Research. Diet, Nutrition, Physical Activity and Cancer: A Global Perspective; Continuous Update Project Expert Report; American Institute for Cancer Research: Washington, DC, USA, 2018; ISBN 978-1-912259-47-2. [Google Scholar]
- Nosrati, N.; Bakovic, M.; Paliyath, G. Molecular Mechanisms and Pathways as Targets for Cancer Prevention and Progression with Dietary Compounds. Int. J. Mol. Sci. 2017, 18, 2050. [Google Scholar] [CrossRef] [PubMed]
- Khanna, A.; Sturgis, E.M.; Dahlstrom, K.R.; Xu, L.; Wei, Q.; Li, G.; Gross, N.D. Association of pretreatment body mass index with risk of head and neck cancer: A large single-center study. Am. J. Cancer Res. 2021, 11, 2343–2350. [Google Scholar] [PubMed]
- Fattouh, M.; Chang, G.Y.; Ow, T.J.; Shifteh, K.; Rosenblatt, G.; Patel, V.M.; Smith, R.V.; Prystowsky, M.B.; Nicolas, F. Association between pre-treatment obesity, sarcopenia and survival in head and neck cancer patients. Head Neck. 2019, 41, 707–714. [Google Scholar] [CrossRef]
- Nagao, T.; Warnakulasuriya, S.; Nakamura, T.; Kato, S.; Yamamoto, K.; Fukano, H.; Suzuki, K.; Shimozato, K.; Hashimoto, S. Treatment of oral leukoplakia with a low-dose of beta-carotene and vitamin C supplements: A randomized controlled trial. Int. J. Cancer 2015, 136, 1708–1717. [Google Scholar] [CrossRef]
- Lodi, G.; Franchini, R.; Warnakulasuriya, S.; Varoni, E.M.; Sardella, A.; Kerr, A.R.; Carrassi, A.; MacDonald, L.C.; Worthington, H.V. Interventions for treating oral leukoplakia to prevent oral cancer. Cochrane Database Syst. Rev. 2016, 7, CD001829. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wub, X.; Zhuang, W.; Xia, L.; Chen, Y.; Wuc, C.; Rao, Z.; Du, L.; Zhao, R.; Yi, M.; et al. Tomato and lycopene and multiple health outcomes: Umbrella review. Food Chemistry. 2021, 343, 128396. [Google Scholar] [CrossRef]
- Yang, T.; Yang, X.; Wang, X.; Wang, Y.; Song, Z. The role of tomato products and lycopene in the prevention of gastric cancer: A meta-analysis of epidemiologic studies. Med. Hypotheses 2013, 80, 383–388. [Google Scholar] [CrossRef]
- Wang, X.; Yang, H.H.; Liu, Y.; Zhou, Q.; Chen, Z.H. Lycopene Consumption and Risk of Colorectal Cancer: A Meta-Analysis of Observational Studies. Nutr. Cancer 2016, 68, 1083–1096. [Google Scholar] [CrossRef]
- Li, X.; Xu, J. Meta-analysis of the association between dietary lycopene intake and ovarian cancer risk in postmenopausal women. Sci. Rep. 2014, 4, 4885. [Google Scholar] [CrossRef]
- Mittelman, S.D. The Role of Diet in Cancer Prevention and Chemotherapy Efficacy. Annu. Rev. Nutr. 2020, 40, 273–297. [Google Scholar] [CrossRef]
- Shin, J.; Song, M.H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef] [PubMed]
- Focsan, A.L.; Polyakov, N.E.; Kispert, L.D. Carotenoids: Importance in Daily Life–Insight Gained from EPR and ENDOR. Appl. Magn. Reson. 2021, 52, 1093–1112. [Google Scholar] [CrossRef] [PubMed]
- Black, H.S.; Boehm, F.; Edge, R.; Truscott, T.G. The Benefits and Risks of Certain Dietary Carotenoids that Exhibit both Anti- and Pro-Oxidative Mechanisms–A Comprehensive Review. Antioxidants 2020, 9, 264. [Google Scholar] [CrossRef] [PubMed]
- Moody, L.; Crowder, S.L.; Fruge, A.D.; Locher, J.L.; Demark-Wahnefried, W.; Rogers, L.Q.; Delk-Licata, A.; Carroll, W.R.; Spencer, S.A.; Black, M.; et al. Epigenetic stratification of head and neck cancer survivors reveals differences in lycopene levels, alcohol consumption, and methylation of immune regulatory genes. Clin. Epigenetics 2020, 12, 138. [Google Scholar] [CrossRef]
- Crowder, S.L.; Frugé, A.D.; Douglas, K.G.; Chen, Y.T.; Moody, L.; Delk-Licata, A.; Erdman, J.W.; Black, M.; Carroll, W.R.; Spencer, S.A.; et al. Feasibility outcomes of a pilot randomized clinical trial to increase cruciferous and green leafy vegetable intake in Posttreatment head and neck cancer survivors. J. Acad. Nutr. Diet. 2019, 119, 659. [Google Scholar] [CrossRef]
- Nimbalkar, V.K.; Manna, S.; Maru, G.B.; Mahimkar, M.B. Head and Neck Cancer Prevention by Phytochemicals: Current Status and Challenges. Curr. Pharmacol. Rep. 2020, 6, 85–102. [Google Scholar] [CrossRef]
- Murray, M.T.; Capelli, B. 57-Beta-Carotene and Other Carotenoids. In Textbook of Natural Medicine, 5th ed.; Pizzorno, J.E., Murray, M.T., Eds.; Elsevier: St. Louis, MO, USA, 2020; Volume 1, pp. 443–450.e2. ISBN 978-0-323-52342-4. [Google Scholar]
- Aronson, J.K. Vitamin A: Carotenoids. In Meyler’s Side Effects of Drugs, 16th ed.; Aronson, J.K., Ed.; Elsevier Science: Amsterdam, The Netherlands, 2016; pp. 439–451. ISBN 978-0-444-53716-4. [Google Scholar]
- Milani, A.; Basirnejad, M.; Dagliad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef]
- Melendez-Martinez, A.J.; Stinco, C.M.; Mapelli-Brahm, P. Skin carotenoids in public health and nutricosmetics: The emerging roles and applications of the UV radiation-absorbing colourless carotenoids phytoene and phytofluene. Nutrients 2019, 11, 1093. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Saini, R.K.; Sivanesan, I.; Keum, Y.-S. Emerging Roles of Carotenoids in the Survival and Adaptations of Microbes. Indian J. Microbiol. 2019, 59, 125–127. [Google Scholar] [CrossRef]
- Toews, D.P.L.; Hofmeister, N.R.; Taylor, S.A. The evolution and genetics of carotenoid processing in animals. Trends Genet. 2017, 33, 171–182. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef] [PubMed]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.; Widjaja-Adhi, M.A.K.; Rodriguez-Santiago, S.; Hessel, S.; Golczak, M.; Palczewski, K.; von Lintig, J. Two Carotenoid Oxygenases Contribute to Mammalian Provitamin A Metabolis. J. Biol. Chem. 2013, 288, 34081–34096. [Google Scholar] [CrossRef]
- Eroglu, A.; Harrison, E.H. Carotenoid metabolism in mammals, including man: Formation, occurrence, and function of apocarotenoids. J. Lipid Res. 2013, 54, 1719–1730. [Google Scholar] [CrossRef]
- Amengual, J.; Lobo, G.P.; Golczak, M.; Li, H.N.; Klimova, T.; Hoppel, C.L.; Wyss, A.; Palczewski, K.; von Lintig, J. A mitochondrial enzyme degrades carotenoids and protects against oxidative stress. FASEB J. 2011, 25, 948–959. [Google Scholar] [CrossRef]
- Mein, J.R.; Dolnikowski, G.G.; Ernst, H.; Russell, R.M.; Wang, X.D. Enzymatic formation of apo-carotenoids from the xanthophyll carotenoids lutein, zeaxanthin, and β-cryptoxanthin by ferret carotene-9′,10′-monooxygenase. Arch. Biochem. Biophys. 2011, 506, 109–121. [Google Scholar] [CrossRef]
- Arathi, B.P.; Sowmya, P.R.; Kuriakose, G.C.; Vijay, K.; Baskaran, V.; Jayabaskaran, C.; Lakshminarayana, R. Enhanced cytotoxic and apoptosis inducing activity of lycopene oxidation products in different cancer cell lines. Food Chem. Toxicol. 2016, 97, 265–276. [Google Scholar] [CrossRef]
- Kavitha, K.; Kowshik, J.; Kishore, T.K.K.; Baba, A.B.; Nagini, S. Astaxanthin inhibits NF-κB and Wnt/β-catenin signaling pathways via inactivation of Erk/MAPK and PI3K/Akt to induce intrinsic apoptosis in a hamster model of oral cancer. Biochim. Biophys. Acta 2013, 1830, 4433–4444. [Google Scholar] [CrossRef]
- Dos Santos, R.C.; Ombredane, A.S.; Souza, J.M.T.; Vasconcelos, A.G.; Placido, A.; Amorim, A.; Barbosa, E.A.; Lima, F.; Ropke, C.D.; Alves, M.M.M.; et al. Lycopene-rich extract from red guava (Psidium guajava L.) displays cytotoxic effect against human breast adenocarcinoma cell line MCF-7 via an apoptotic-like pathway. Food Res. Int. 2018, 105, 184–196. [Google Scholar] [CrossRef]
- Ganesan, P.; Noda, K.; Manabe, Y.; Ohkubo, T.; Tanaka, Y.; Maoka, T.; Sugawara, T.; Hirata, T. Siphonaxanthin, a marine carotenoid from green algae, effectively induces apoptosis in human leukemia (HL-60) cells. Biochim. Biophys. Acta 2011, 1810, 497–503. [Google Scholar] [CrossRef]
- Sowmya, P.R.-R.; Arathi, B.P.; Vijay, K.; Baskaran, V.; Lakshminarayana, R. Astaxanthin from shrimp efficiently modulates oxidative stress and allied cell death progression in MCF-7 cells treated synergistically with β-carotene and lutein from greens. Food Chem. Toxicol. 2017, 106 Pt A, 58–69. [Google Scholar] [CrossRef]
- Liu, C.-L.; Huang, Y.-S.; Hosokawa, M.; Miyashita, K.; Hu, M.-L. Inhibition of proliferation of a hepatoma cell line by fucoxanthin in relation to cell cycle arrest and enhanced gap junctional intercellular communication. Chem. Biol. Interact. 2009, 182, 165–172. [Google Scholar] [CrossRef]
- Puah, B.-P.; Jalil, J.; Attiq, A.; Kamisah, Y. New Insights into Molecular Mechanism behind Anti-Cancer Activities of Lycopene. Molecules 2021, 26, 3888. [Google Scholar] [CrossRef]
- Eid, S.Y.; El-Readi, M.Z.; Wink, M. Carotenoids reverse multidrug resistance in cancer cells by interfering with ABC-transporters. Phytomedicine 2012, 19, 977–987. [Google Scholar] [CrossRef]
- Eliassen, A.H.; Hendrickson, S.J.; Brinton, L.A.; Buring, J.E.; Campos, H.; Dai, Q.; Dorgan, J.F.; Franke, A.A.; Gao, Y.-T.; Goodman, M.T.; et al. Circulating carotenoids and risk of breast cancer: Pooled analysis of eight prospective studies. J. Natl. Cancer Inst. 2012, 104, 1905–1916. [Google Scholar] [CrossRef]
- Jung, S.; Wu, K.; Giovannucci, E.; Spiegelman, D.; Willett, W.C.; Smith-Warner, S.A. Carotenoid intake and risk of colorectal adenomas in a cohort of male health professionals. Cancer Causes Control 2013, 24, 705–717. [Google Scholar] [CrossRef]
- Takata, Y.; Xiang, Y.-B.; Yang, G.; Li, H.; Gao, J.; Cai, H.; Gao, Y.-T.; Zheng, W.; Shu, X.-O. Intakes of fruits, vegetables, and related vitamins and lung cancer risk: Results from the Shanghai Men’s Health Study (2002–2009). Nutr. Cancer 2013, 65, 51–61. [Google Scholar] [CrossRef]
- Zu, K.; Mucci, L.; Rosner, B.A.; Clinton, S.K.; Loda, M.; Stampfer, M.J.; Giovannucci, E. Dietary lycopene, angiogenesis, and prostate cancer: A prospective study in the prostate-specific antigen era. J. Natl. Cancer Inst. 2014, 106, djt430. [Google Scholar] [CrossRef]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.-S. Microbial platforms to produce commercially vital carotenoids at industrial scale: An updated review of critical issues. J. Ind. Microbiol. Biotechnol. 2018, 46, 657–674. [Google Scholar] [CrossRef]
- Maiani, G.; Caston, M.J.; Catasta, G.; Toti, E.; Cambrodon, I.G.; Bysted, A.; GranadoLorencio, F.; Olmedilla-Alonso, B.; Knuthsen, P.; Valoti, M.; et al. Carotenoids: Actual knowledge on food sources, intakes, stability and bioavailability and their protective role in humans. Mol. Nutr. Food Res. 2009, 53 (Suppl. 2), S194–S218. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Kim, K.-N.; Heo, S.-J.; Kang, S.-M.; Ahn, G.; Jeon, Y.-J. Fucoxanthin induces apoptosis in human leukemia HL-60 cells through a ROS-mediated Bcl-xL pathway. Toxicol. Vitr. 2010, 24, 1648–1654. [Google Scholar] [CrossRef]
- Gansukh, E.; Nile, A.; Sivanesan, I.; Rengasamy, K.R.R.; Kim, D.-H.; Keum, Y.-S.; Saini, R.K. Chemopreventive effect of β-Cryptoxanthin on human cervical carcinoma (HeLa) cells is modulated through oxidative stress-induced apoptosis. Antioxidants 2019, 9, 28. [Google Scholar] [CrossRef]
- Gansukh, E.; Mya, K.K.; Jung, M.; Keum, Y.S.; Kim, D.H.; Saini, R.K. Lutein derived from marigold (Tagetes erecta) petals triggers ROS generation and activates Bax and caspase-3 mediated apoptosis of human cervical carcinoma (HeLa) cells. Food Chem. Toxicol. 2019, 127, 11–18. [Google Scholar] [CrossRef]
- Vijay, K.; Sowmya, P.R.-R.; Arathi, B.P.; Shilpa, S.; Shwetha, H.J.; Raju, M.; Baskaran, V.; Lakshminarayana, R. Low-dose doxorubicin with carotenoids selectively alters redox status and upregulates oxidative stress-mediated apoptosis in breast cancer cells. Food Chem. Toxicol. 2018, 118, 675–690. [Google Scholar] [CrossRef]
- Palozza, P.; Serini, S.; Di Nicuolo, F.; Piccioni, E.; Calviello, G. Prooxidant effects of beta-carotene in cultured cells. Mol. Aspects Med. 2003, 24, 353–362. [Google Scholar] [CrossRef]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef]
- McClements, D.J.; Li, F.; Xiao, H. The nutraceutical bioavailability classification scheme: Classifying nutraceuticals according to factors limiting their oral bioavailability. Annu. Rev. Food Sci. Technol. 2015, 6, 232–299. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Zhai, K.; Abotaleb, M.; Ashrafizadeh, M.; Brockmueller, A.; Shakibaei, M.; Biringer, K.; Bugos, O.; et al. Carotenoids in Cancer Metastasis-Status Quo and Outlook. Biomolecules 2020, 10, 1653. [Google Scholar] [CrossRef]
- Chang, H.P.; Sheen, L.-Y.; Lei, Y.-P. The protective role of carotenoids and polyphenols in patients with head and neck cancer. J. Chin. Med. Assoc. 2015, 78, 89–95. [Google Scholar] [CrossRef]
- Traub, M. 192-Leukoplakia. In Textbook of Natural Medicine, 5th ed.; Pizzorno, J.E., Murray, M.T., Eds.; Elsevier: St. Louis, MO, USA, 2020; Volume 2, pp. 1528–1529. ISBN 978-0-323-52342-4. [Google Scholar]
- Bacanli, M.; Başaran, N.; Başaran, A.A. Lycopene: Is it Beneficial to Human Health as an Antioxidant? Turk. J. Pharm. Sci. 2017, 14, 311–318. [Google Scholar] [CrossRef]
- Anlar, H.A.; Bacanli, M. Chapter 24-Lycopene as an antioxidant in human health and diseases. In Pathology Oxidative Stress and Dietary Antioxidants, 1st ed.; Preedy, V.R., Ed.; Academic Press and Elsevier: London, UK, 2020; pp. 247–254. ISBN 978-0-12-815972-9. [Google Scholar]
- Rowles, J.L., 3rd; Ranard, K.M.; Applegate, C.C.; Jeon, S.; An, R.; Erdman, J.W., Jr. Processed and raw tomato consumption and risk of prostate cancer: A systematic review and dose–response meta-analysis. Prostate Cancer Prostatic Dis. 2018, 21, 319–336. [Google Scholar] [CrossRef]
- Rowles, J.L., 3rd; Ranard, K.M.; Smith, J.W.; An, R.; Erdman, J.W., Jr. Increased dietary and circulating lycopene are associated with reduced prostate cancer risk: A systematic review and meta-analysis. Prostate Cancer Prostatic Dis. 2017, 20, 361–377. [Google Scholar] [CrossRef]
- Sakhi, A.K.; Bøhn, S.K.; Smeland, S.; Thoresen, M.; Smedshaug, G.B.; Tausjø, J.; Svilaas, A.; Karlsen, A.; Russnes, K.M.; Svilaas, T.; et al. Postradiotherapy plasma lutein, alpha-carotene, and beta-carotene are positively associated with survival in patients with head and neck squamous cell carcinoma. Nutr. Cancer 2010, 62, 322–328. [Google Scholar] [CrossRef]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef]
- Shimizu, R.; Ibaragi, S.; Eguchi, T.; Kuwajima, D.; Kodama, S.; Nishioka, T.; Okui, T.; Obata, K.; Takabatake, K.; Kawai, H.; et al. Nicotine promotes lymph node metastasis and cetuximab resistance in head and neck squamous cell carcinoma. Int. J. Oncol. 2019, 54, 283–294. [Google Scholar] [CrossRef]
- Marziliano, A.; Teckie, S.; Diefenbach, M.A. Review Alcohol-related head and neck cancer: Summary of the literature. Head Neck. 2020, 42, 732–738. [Google Scholar] [CrossRef]
- Hanna, G.J.; Kacew, A.; Chau, N.G.; Shivdasani, P.; Lorch, J.H.; Uppaluri, R.; Haddad, R.I.; MacConaill, L.E. Improved outcomes in PI3K-pathway-altered metastatic HPV oropharyngeal cancer. JCI Insight 2018, 3, e122799. [Google Scholar] [CrossRef]
- Leemans, C.R.; Snijders, P.J.F.; Brakenhoff, R.H. The molecular landscape of head and neck cancer. Nat. Rev. Cancer 2018, 18, 269–282. [Google Scholar] [CrossRef]
- Sainia, R.K.; Keumc, Y.-S.; Dagliad, M.; Rengasamya, K.R.R. Dietary carotenoids in cancer chemoprevention and chemotherapy: A review of emerging evidence. Pharmacol. Res. 2020, 157, 104830. [Google Scholar] [CrossRef]
- Zhang, Y.; Ng, P.K.-S.; Kucherlapati, M.; Chen, F.; Liu, Y.; Tsang, Y.H.; de Velasco, G.; Jeong, K.J.; Akbani, R.; Hadjipanayis, A.; et al. A Pan-Cancer Proteogenomic Atlas of PI3K/AKT/mTOR Pathway Alterations. Cancer Cell. 2017, 31, 820–832.e3. [Google Scholar] [CrossRef]
- Duan, Y.; Haybaeck, J.; Yang, Z. Therapeutic Potential of PI3K/AKT/mTOR Pathway in Gastrointestinal Stromal Tumors: Rationale and Progress. Cancers 2020, 12, 2972. [Google Scholar] [CrossRef]
- Griñan-Lison, C.; Blaya-Cánovas, J.L.; López-Tejada, A.; Ávalos-Moreno, M.; Navarro-Ocón, A.; Cara, F.E.; González-González, A.; Lorente, J.A.; Marchal, J.A.; Granados-Principal, S. Antioxidants for the Treatment of Breast Cancer: Are We There Yet? Antioxidants 2021, 10, 205. [Google Scholar] [CrossRef]
- AL-Ishaq, R.K.; Overy, A.J.; Büsselberg, D. Phytochemicals and Gastrointestinal Cancer: Cellular Mechanisms and Effects to Change Cancer Progression. Biomolecules 2020, 10, 105. [Google Scholar] [CrossRef]
- Chen, H.; Liu, H.; Qing, G. Targeting oncogenic Myc as a strategy for cancer treatment. Signal Transduct. Target. Ther. 2018, 3, 5. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Kim, Y.-M.; Hong, S. Astaxanthin suppresses the metastasis of colon cancer by inhibiting the MYC-mediated downregulation of microRNA-29a-3p and microRNA-200a. Sci. Rep. 2019, 9, 9457. [Google Scholar] [CrossRef]
- Shree, G.S.; Prasad, K.Y.; Arpitha, H.S.; Deepika, U.R.; Kumar, K.N.; Mondal, P.; Ganesan, P. β-carotene at physiologically attainable concentration induces apoptosis and down-regulates cell survival and antioxidant markers in human breast cancer (MCF-7) cells. Mol. Cell. Biochem. 2017, 436, 1–12. [Google Scholar] [CrossRef]
- Zou, X.; Gao, J.; Zheng, Y.; Wang, X.; Chen, C.; Cao, K.; Xu, J.; Li, Y.; Lu, W.; Liu, J.; et al. Zeaxanthin induces Nrf2-mediated phase II enzymes in protection of cell death. Cell Death Dis. 2014, 5, e1218. [Google Scholar] [CrossRef]
- Singh, R.; Letai, A.; Sarosiek, K. Regulation of apoptosis in health and disease: The balancing act of BCL-2 family proteins. Nat. Rev. Mol. Cell Biol. 2019, 20, 175–193. [Google Scholar] [CrossRef] [PubMed]
- Rathore, R.; McCallum, J.E.; Varghese, E.; Florea, A.M.; Büsselberg, D. Overcoming chemotherapy drug resistance by targeting inhibitors of apoptosis proteins (IAPs). Apoptosis 2017, 22, 898–919. [Google Scholar] [CrossRef] [PubMed]
- Razeghian, E.; Suksatan, W.; Rahman, H.S.; Bokov, D.O.; Abdelbasset, W.K.; Hassanzadeh, A.; Marofi, F.; Yazdanifar, M.; Jarahian, M. Harnessing TRAIL-Induced Apoptosis Pathway for Cancer Immunotherapy and Associated Challenges. Front. Immunol. 2021, 12, 699746. [Google Scholar] [CrossRef]
- Yun, T.; Yu, K.; Yang, S.; Cui, Y.; Wang, Z.; Ren, H.; Chen, S.; Li, L.; Liu, X.; Fang, M.; et al. Acetylation of p53 Protein at Lysine 120 Up-regulates Apaf-1 Protein and Sensitizes the Mitochondrial Apoptotic Pathway. J. Biol. Chem. 2016, 291, 7386–7395. [Google Scholar] [CrossRef]
- Zhang, X.; Zhao, W.-E.; Hu, L.; Zhao, L.; Huang, J. Carotenoids inhibit proliferation and regulate expression of peroxisome proliferators-activated receptor gamma (PPARγ) in K562 cancer cells. Arch. Biochem. Biophys. 2011, 512, 96–106. [Google Scholar] [CrossRef]
- Kowshik, J.; Baba, A.B.; Giri, H.; Reddy, D.G.; Dixit, M.; Nagini, S. Astaxanthin inhibits JAK/STAT-3 signaling to abrogate cell proliferation, invasion and angiogenesis in a hamster model of oral cancer. PLoS ONE 2014, 9, e109114. [Google Scholar] [CrossRef]
- Palozza, P.; Torelli, C.; Boninsegna, A.; Simone, R.; Catalano, A.; Mele, M.C.; Picci, N. Growth-inhibitory effects of the astaxanthin-rich alga Haematococcus pluvialis in human colon cancer cells. Cancer Lett. 2009, 283, 108–117. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H. Multiple Mechanisms of Anti-Cancer Effects Exerted by Astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef]
- Satomi, Y.; Nishino, H. Implication of mitogen-activated protein kinase in the induction of G1 cell cycle arrest and gadd45 expression by the carotenoid fucoxanthin in human cancer cells. Biochim. Biophys. Acta 2009, 1790, 260–266. [Google Scholar] [CrossRef]
- Nagendraprabhu, P.; Sudhandiran, G. Astaxanthin inhibits tumor invasion by decreasing extracellular matrix production and induces apoptosis in experimental rat colon carcinogenesis by modulating the expressions of ERK-2, NFkB and COX-2. Investig. New Drugs 2011, 29, 207–224. [Google Scholar] [CrossRef]
- Yasui, Y.; Hosokawa, M.; Mikami, N.; Miyashita, K.; Tanaka, T. Dietary astaxanthin inhibits colitis and colitis-associated colon carcinogenesis in mice via modulation of the inflammatory cytokines. Chem. Biol. Interact. 2011, 193, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Bi, M.-C.; Rosen, R.; Zha, R.-Y.; McCormick, S.A.; Song, E.; Hu, D.-N. Zeaxanthin induces apoptosis in human uveal melanoma cells through Bcl-2 family proteins and intrinsic apoptosis pathway. Evid. Complement. Alternat. Med. 2013, 2013, 205082. [Google Scholar] [CrossRef] [PubMed]
- Sheng, Y.-N.; Luo, Y.-H.; Liu, S.-B.; Xu, W.-T.; Zhang, Z.; Zhang, T.; Xue, H.; Zuo, W.-B.; Li, Y.-N.; Wang, C.-Y.; et al. Zeaxanthin Induces Apoptosis via ROS-Regulated MAPK and AKT Signaling Pathway in Human Gastric Cancer Cells. Onco Targets Ther. 2020, 13, 10995–11006. [Google Scholar] [CrossRef] [PubMed]
- Cilla, A.; Attanzio, A.; Barberá, R.; Tesoriere, L.; Livrea, M.A. Anti-proliferative effect of main dietary phytosterols and β-cryptoxanthin alone or combined in humancolon cancer Caco-2 cells through cytosolic Ca+2–and oxidative stress-induced apoptosis. J. Funct. Foods 2015, 12, 282–293. [Google Scholar] [CrossRef]
- Haddad, N.F.; Teodoro, A.J.; de Oliveira, F.L.; Soares, N.; de Mattos, R.M.; Hecht, F.; Dezonne, R.S.; Vairo, L.; Goldenberg, R.C.D.S.; Gomes, F.C.A.; et al. Lycopene and beta-carotene induce growth inhibition and proapoptotic effects on ACTH-secreting pituitary adenoma cells. PLoS ONE 2013, 8, e62773. [Google Scholar] [CrossRef]
- Takeshima, M.; Ono, M.; Higuchi, T.; Chen, C.; Hara, T.; Nakano, S. Anti-proliferative and apoptosis-inducing activity of lycopene against three subtypes of human breast cancer cell lines. Cancer Sci. 2014, 105, 252–257. [Google Scholar] [CrossRef]
- Kim, K.-N.; Ahn, G.; Heo, S.-J.; Kang, S.-M.; Kang, M.-C.; Yang, H.-M.; Kim, D.; Roh, S.W.; Kim, S.-K.; Jeon, B.-T.; et al. Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environ. Toxicol. Pharmacol. 2013, 35, 39–46. [Google Scholar] [CrossRef]
- Yu, R.-X.; Hu, X.-M.; Xu, S.-Q.; Jiang, Z.-J.; Yang, W. Effects of fucoxanthin on proliferation and apoptosis in human gastric adenocarcinoma MGC-803 cells via JAK/STAT signal pathway. Eur. J. Pharmacol. 2011, 657, 10–19. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.; Shen, L. Extracellular vesicle-mediated regulation of tumor angiogenesis—implications for anti-angiogenesis therapy. J. Cell. Mol. Med. 2021, 25, 2776–2785. [Google Scholar] [CrossRef]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportunities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef]
- Muniz-Bongers, L.R.; McClain, C.B.; Saxena, M.; Bongers, G.; Merad, M.; Bhardwaj, N. MMP2 and TLRs modulate immune responses in the tumor microenvironment. JCI Insight 2021, 6, e144913. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, L.; Jayakar, S.K.; Ow, T.J.; Segall, J.E. Mechanisms of Invasion in Head and Neck Cancer. Arch. Pathol. Lab. Med. 2015, 139, 1334–1348. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-S.; Chuang, C.-H.; Lo, T.-F.; Hu, M.-L. Anti-angiogenic effects of lycopene through immunomodualtion of cytokine secretion in human peripheral blood mononuclear cells. J. Nutr. Biochem. 2013, 24, 428–434. [Google Scholar] [CrossRef] [PubMed]
- Quintero-Fabián, S.; Arreola, R.; Becerril-Villanueva, E.; Torres-Romero, J.C.; Arana-Argáez, V.; Lara-Riegos, J.; Ramírez-Camacho, M.A.; Alvarez-Sánchez, M.E. Role of Matrix Metalloproteinases in Angiogenesis and Cancer. Front. Oncol. 2019, 9, 1370. [Google Scholar] [CrossRef]
- Méresse, S.; Fodil, M.; Fleury, F.; Chénais, B. Fucoxanthin, a Marine-Derived Carotenoid from Brown Seaweeds and Microalgae: A Promising Bioactive Compound for Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 9273. [Google Scholar] [CrossRef]
- Koklesova, L.; Liskova, A.; Samec, M.; Buhrmann, C.; Samuel, S.M.; Varghese, E.; Ashrafizadeh, M.; Najafi, M.; Shakibaei, M.; Büsselberg, D.; et al. Carotenoids in Cancer Apoptosis—The Road from Bench to Bedside and Back. Cancers 2020, 12, 2425. [Google Scholar] [CrossRef]
- Mátyási, B.; Farkas, Z.; Kopper, L.; Sebestyén, A.; Boissan, M.; Mehta, A.; Takács-Vellai, K. The Function of NM23-H1/NME1 and Its Homologs in Major Processes Linked to Metastasis. Pathol. Oncol. Res. 2020, 26, 49–61. [Google Scholar] [CrossRef]
- Kim, B.; Lee, K.-J. Activation of Nm23-H1 to suppress breast cancer metastasis via redox regulation. Exp. Mol. Med. 2021, 53, 346–357. [Google Scholar] [CrossRef]
- Wu, D.-M.; Deng, S.H.; Liu, T.; Han, R.; Zhang, T.; Xu, Y. TGF-β-mediated exosomal lnc-MMP2-2 regulates migration and invasion of lung cancer cells to the vasculature by promoting MMP2 expression. Cancer Med. 2018, 7, 5118–5129. [Google Scholar] [CrossRef]
- Chen, H.Y.; Yueh, T.C.; Chen, Y.C.; Huang, C.H.; Yang, C.M.; Hu, M.L. Antimetastatic effects of alpha-carotene and possible mechanisms of action in human hepatocarcinoma SK-Hep-1 cells. J. Agric. Food Chem. 2013, 61, 10368–11037. [Google Scholar] [CrossRef]
- Kotake-Nara, E.; Kushiro, M.; Zhang, H.; Sugawara, T.; Miyashita, K.; Nagao, A. Carotenoids affect proliferation of human prostate cancer cells. J. Nutr. 2001, 131, 3303–3306. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Lee, H.-A.; Lim, J.Y.; Kim, Y.; Jung, C.-H.; Yoo, S.-H.; Kim, Y. β-Carotene inhibits neuroblastoma cell invasion and metastasis in vitro and in vivo by decreasing level of hypoxia-inducible factor-1α. J. Nutr. Biochem. 2014, 25, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Chen, J.; Li, M.; Tang, L.; Wu, R.; Jin, L.; Liang, Z. β-carotene reverses tobacco smoke-induced gastric EMT via Notch pathway in vivo. Oncol. Rep. 2018, 39, 1867–1873. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Kim, Y.; Kim, Y.S.; Shin, J.-H.; Rubin, L.P.; Kim, Y. β-Carotene exerts anti-colon cancer effects by regulating M2 macrophages and activated fibroblasts. J. Nutr. Biochem. 2020, 82, 108402. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-C.; Wang, F.-Y.; Kuo, Y.-H.; Tang, F.-Y. Cancer chemopreventive effects of lycopene: Suppression of MMP-7 expression and cell invasion in human colon cancer cells. J. Agric. Food Chem. 2011, 59, 11304–11318. [Google Scholar] [CrossRef]
- Trejo-Solís, C.; Pedraza-Chaverrí, J.; Torres-Ramos, M.; Jiménez-Farfán, D.; Cruz Salgado, A.; Serrano-García, N.; Osorio-Rico, L.; Sotelo, J. Multiple Molecular and Cellular Mechanisms of Action of Lycopene in Cancer Inhibition. Evid. Based Complement. Altern. Med. 2013, 2013, 705121. [Google Scholar] [CrossRef]
- Marzocco, S.; Singla, R.K.; Capasso, A. Multifaceted Effects of Lycopene: A Boulevard to the Multitarget-Based Treatment for Cancer. Molecules 2021, 26, 5333. [Google Scholar] [CrossRef]
- Bhuvaneswari, V.; Velmurugan, B.; Balasenthil, S.; Ramachandran, C.R.; Nagini, S. Chemopreventive efficacy of lycopene on 7,12-dimethylbenz[a]anthracene-induced hamster buccal pouch carcinogenesis. Fitoterapia 2001, 72, 865–874. [Google Scholar] [CrossRef]
- Yang, P.M.; Chen, H.Z.; Huang, Y.T.; Hsieh, C.W.; Wung, B.S. Lycopene inhibits NF-κB activation and adhesion molecule expression through Nrf2-mediated heme oxygenase-1 in endothelial cells. Int. J. Mol. Med. 2017, 39, 1533–1540. [Google Scholar] [CrossRef]
- Lian, F.; Wang, X.D. Enzymatic metabolites of lycopene induce Nrf2-mediated expression of phase II detoxifying/antioxidant enzymes in human bronchial epithelial cells. Int. J. Cancer 2008, 123, 1262–1268. [Google Scholar] [CrossRef]
- Yang, C.-M.; Hu, T.-Y.; Hu, M.-L. Antimetastatic effects and mechanisms of apo-8′-lycopenal, an enzymatic metabolite of lycopene, against human hepatocarcinoma SK-Hep-1 cells. Nutr. Cancer 2012, 64, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Ip, B.C.; Hu, K.-Q.; Liu, C.; Smith, D.E.; Obin, M.S.; Ausman, L.M.; Wang, X.-D. Lycopene metabolite, apo-10′-lycopenoic acid, inhibits diethylnitrosamine-initiated, high fat diet-promoted hepatic inflammation and tumorigenesis in mice. Cancer Prev. Res. 2013, 6, 1304–1316. [Google Scholar] [CrossRef] [PubMed]
- Jhou, B.-Y.; Song, T.-Y.; Lee, I.; Hu, M.-L.; Yang, N.-C. Lycopene Inhibits Metastasis of Human Liver Adenocarcinoma SK-Hep-1 Cells by Downregulation of NADPH Oxidase 4 Protein Expression. J. Agric. Food Chem. 2017, 65, 6893–6903. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Lu, X.; Yu, R. Lycopene Inhibits Epithelial–Mesenchymal Transition and Promotes Apoptosis in Oral Cancer via PI3K/AKT/m-TOR Signal Pathway. Drug Des. Dev. Ther. 2020, 14, 2461–2471. [Google Scholar] [CrossRef]
- Ming, J.X.; Wang, Z.C.; Huang, Y.; Ohishi, H.; Wu, R.J.; Shao, Y.; Wang, H.; Qin, M.Y.; Wu, Z.L.; Li, Y.Y.; et al. Fucoxanthin extracted from Laminaria Japonica inhibits metastasis and enhances the sensitivity of lung cancer to Gefitinib. J. Ethnopharmacol. 2021, 265, 113302. [Google Scholar] [CrossRef]
- Garg, S.; Afzal, S.; Elwakeel, A.; Sharma, D.; Radhakrishnan, N.; Dhanjal, J.K.; Sundar, D.; Kaul, S.C.; Wadhwa, R. Marine Carotenoid Fucoxanthin Possesses Anti-Metastasis Activity: Molecular Evidence. Mar. Drugs 2019, 17, 338. [Google Scholar] [CrossRef]
- Li, Y.C.; Cheng, A.J.; Lee, L.Y.; Huang, Y.C.; Chang, J.T. Multifaceted mechanisms of areca nuts in oral carcinogenesis: The molecular pathology from precancerous condition to malignant transformation. J. Cancer 2019, 10, 4054–4062. [Google Scholar] [CrossRef]
- Chang, N.W.; Pei, R.J.; Tseng, H.C.; Yeh, K.T.; Chan, H.C.; Lee, M.R.; Lin, C.; Hsieh, W.-T.; Kao, M.-C.; Tsai, M.-H.; et al. Co-treating with arecoline and 4-nitroquinoline 1-oxide to establish a mouse model mimicking oral tumorigenesis. Chem. Biol. Interact. 2010, 183, 231–237. [Google Scholar] [CrossRef]
- Kavitha, K.; Thiyagarajan, P.; Nandhini, J.R.; Mishra, R.; Nagini, S. Chemopreventive effects of diverse dietary phytochemicals against DMBA-induced hamster buccal pouch carcinogenesis via the induction of Nrf2-mediated cytoprotective antioxidant, detoxification, and DNA repair enzymes. Biochimie 2013, 95, 1629–1639. [Google Scholar] [CrossRef]
- Bhuvaneswari, V.; Rao, K.S.; Nagini, S. Altered expression of anti and proapoptotic proteins during chemoprevention of hamster buccal pouch carcinogenesis by tomato and garlic combination. Clin. Chim. Acta 2004, 350, 65–72. [Google Scholar] [CrossRef]
- Supsavhad, W.; Dirksen, W.P.; Martin, C.K.; Rosol, T.J. Animal models of head and neck squamous cell carcinoma. Vet. J. 2016, 210, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Wu, Q.; Zhang, M.; Huang, J. Lycopene inhibits the cell proliferation and invasion of human head and neck squamous cell carcinoma. Mol. Med. Rep. 2016, 14, 2953–2958. [Google Scholar] [CrossRef] [PubMed]
- Steward, W.P.; Brown, K. Cancer chemoprevention: A rapidly evolving field. Br. J. Cancer 2013, 109, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.C.; Kang, H.J.; Kim, Y.S.; Choi, E.C. All-trans-retinoic acid inhibits growth of head and neck cancer stem cells by suppression of Wnt/beta-catenin pathway. Eur. J. Cancer 2012, 48, 3310–3318. [Google Scholar] [CrossRef]
- Bertrand, G.; Maalouf, M.; Boivin, A.; Battiston-Montagne, P.; Beuve, M.; Levy, A.; Jalade, P.; Fournier, C.; Ardail, D.; Magné, N.; et al. Targeting head and neck cancer stem cells to overcome resistance to photon and carbon ion radiation. Stem Cell Rev. 2014, 10, 114–126. [Google Scholar] [CrossRef]
- Chen, D.; Wang, C.-Y. Targeting cancer stem cells in squamous cell carcinoma. Precis. Clin. Med. 2019, 2, 152–165. [Google Scholar] [CrossRef]
- Tao, A.; Wang, X.; Li, C. Effect of Lycopene on Oral Squamous Cell Carcinoma Cell Growth by Inhibiting IGF1 Pathway. Cancer Manag. Res. 2021, 13, 723–732. [Google Scholar] [CrossRef]
- Rahman, M.A.; Amin, A.R.M.; Shin, D.M. Chemopreventive Potential of Natural Compounds in Head and Neck Cancer. Nutr. Cancer 2010, 62, 973–987. [Google Scholar] [CrossRef]
- McCarthy, C.; Fedele, S.; Ottensmeier, C.; Shaw, R.J. Early-Phase Interventional Trials in Oral Cancer Prevention. Cancers 2021, 13, 3845. [Google Scholar] [CrossRef]
- Arduino, P.G.; Bagan, J.; El-Naggar, A.K.; Carrozzo, M. Urban legends series: Oral leukoplakia. Oral Diseases 2013, 19, 642–659. [Google Scholar] [CrossRef]
- Warnakulasuriya, S.; Ariyawardana, A. Malignant transformation of oral leukoplakia: A systematic review of observational studies. J. Oral Pathol. Med. 2016, 45, 155–166. [Google Scholar] [CrossRef] [PubMed]
- van der Waal, I.; Axell, T. Oral leukoplakia: A proposal for uniform reporting. Oral Oncol. 2002, 38, 521–526. [Google Scholar] [CrossRef]
- Marron, M.; Boffetta, P.; Zhang, Z.F.; Zaridze, D.; Wünsch-Filho, V.; Winn, D.M.; Wei, Q.; Talamini, R.; Szeszenia-Dabrowska, N.; Sturgis, E.M.; et al. Cessation of alcohol drinking, tobacco smoking and the reversal of head and neck cancer risk. Int. J. Epidemiol. 2010, 39, 182–196. [Google Scholar] [CrossRef] [PubMed]
- Nagao, T.; Warnakulasuriya, S.; Sakuma, H.; Miyabe, S.; Hasegawa, S.; Machida, J.; Suzuki, K.; Fukano, H.; Shimozato, K.; Hashimoto, S.; et al. p53 and ki67 as biomarkers in determining response to chemoprevention for oral leukoplakia. J. Oral Pathol. Med. 2017, 46, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Papadimitrakopoulou, V.A.; Lee, J.J.; William, W.N., Jr.; Martin, J.W.; Thomas, M.; Kim, E.S.; Khuri, F.R.; Shin, D.M.; Feng, L.; Hong, W.K.; et al. Randomized trial of 13-cis retinoic acid compared with retinyl palmitate with or without beta-carotene in oral premalignancy. J. Clin. Oncol. 2009, 27, 599–604. [Google Scholar] [CrossRef]
- Ribeiro, A.S.; Salles, P.R.; da Silva, T.A.; Mesquita, R.A. A review of the nonsurgical treatment of oral leukoplakia. Int. J. Dent. 2010, 2010, 186018. [Google Scholar] [CrossRef]
- Maheswari, T.N.U. Treatment of oral leukoplakia with antioxidants–A systematic review. Int. J. Pharma Bio Sci. 2013, 4, 33–41. [Google Scholar]
- Singh, M.; Krishanappa, R.; Bagewadi, A.; Keluskar, V. Efficacy of oral lycopene in the treatment of oral leukoplakia. Oral Oncol. 2004, 40, 591–596. [Google Scholar] [CrossRef]
- Aung, W.P. The use of lycopene in oral potentially malignant disorders. Myan. Dent. J. 2013, 20, 58–63. [Google Scholar]
- Zakrzewska, J.M. Oral lycopene-an efficacious treatment for oral leukoplakia? Evid. Based Dent. 2005, 6, 17–18. [Google Scholar] [CrossRef]
- Johny, J.; Bhagvandas, S.C.; Mohan, S.P.; Punathil, S.; Moyin, S.; Bhaskaran, M.K. Comparison of Efficacy of Lycopene and Lycopene-Hyaluronidase Combination in the Treatment of Oral Submucous Fibrosis. J. Pharm. Bioallied. Sci. 2019, 11 (Suppl. 2), S260–S264. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; Dan, H.; Wu, R.; Meng, W.; Liu, N.; Jin, X.; Zhou, M.; Zeng, X.; Zhou, G.; Chen, Q. Lycopene: Features and potential significance in the oral cancer and precancerous lesions. J. Oral Pathol. Med. 2010, 40, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Maasland, D.H.; van den Brandt, P.A.; Kremer, B.; Goldbohm, R.A.; Schouten, L.J. Consumption of vegetables and fruits and risk of subtypes of head-neck cancer in the Netherlands Cohort Study. Int. J. Cancer 2015, 136, E396–E409. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Khor, T.O.; Shu, L.; Su, Z.Y.; Fuentes, F.; Kong, A.N. Dietary phytochemicals and cancer prevention: Nrf2 signaling, epigenetics, and cell death mechanisms in blocking cancer initiation and progression. Pharmacol. Ther. 2013, 137, 153–171. [Google Scholar] [CrossRef] [PubMed]
- Butler, C.; Lee, Y.A.; Li, S.; Li, Q.; Chen, C.J.; Hsu, W.L.; Lou, P.J.; Zhu, C.; Pan, J.; Shen, H.; et al. Diet and the risk of head-and-neck cancer among never-smokers and smokers in a Chinese population. Cancer Epidemiol. 2017, 46, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Bradshaw, P.T.; Siega-Riz, A.M.; Campbell, M.; Weissler, M.C.; Funkhouser, W.K.; Olshan, A.F. Associations between dietary patterns and head and neck cancer: The Carolina head and neck cancer epidemiology study. Am. J. Epidemiol. 2012, 175, 1225–1233. [Google Scholar] [CrossRef]
- Datta, M.; Shaw, E.G.; Lesser, G.J.; Case, L.D.; Vitolins, M.Z.; Schneider, C.; Frizzell, B.; Sullivan, C.; Lively, M.; Franzmann, E.; et al. A Randomized Double-Blind Placebo-Controlled Trial of Fruit and Vegetable Concentrates on Intermediate Biomarkers in Head and Neck Cancer. Integr. Cancer Ther. 2018, 17, 115–123. [Google Scholar] [CrossRef]
- Chainani-Wu, N. Diet and oral, pharyngeal, and esophageal cancer. Nutr. Cancer 2002, 44, 104–126. [Google Scholar] [CrossRef]
- Harvard School of Public Health Nutrition Department’s File Download Site. Available online: https://regepi.bwh.harvard.edu/health/FFQ/files (accessed on 25 May 2021).
- Li, W.-Q.; Park, Y.; Wu, J.W.; Goldstein, A.M.; Taylor, P.R.; Hollenbeck, A.R.; Freedman, N.D.; Abnet, C.C. Index-based dietary patterns and risk of head and neck cancer in a large prospective study. Am. J. Clin. Nutr. 2014, 99, 559–566. [Google Scholar] [CrossRef]
- Crowder, S.L.; Li, Z.; Sarma, K.P.; Arthur, A.E. Chronic Nutrition Impact Symptoms Are Associated with Decreased Functional Status, Quality of Life, and Diet Quality in a Pilot Study of Long-Term Post-Radiation Head and Neck Cancer Survivors. Nutrients 2021, 13, 2886. [Google Scholar] [CrossRef]
- Gnagnarella, P.; Salvini, S.; Parpinel, M. Food Composition Database for Epidemiological Studies in Italy. Version 1. Available online: http://www.bda-ieo.it/ (accessed on 16 April 2018).
- Lee, Y.A.; Al-Temimi, M.; Ying, J.; Muscat, J.; Olshan, A.F.; Zevallos, J.P.; Winn, D.M.; Li, G.; Sturgis, E.M.; Morgenstern, H.; et al. Risk Prediction Models for Head and Neck Cancer in the US Population from the INHANCE Consortium. Am. J. Epidemiol. 2020, 189, 330–342. [Google Scholar] [CrossRef] [PubMed]
- De Vito, R.; Lee, Y.C.A.; Parpinel, M.; Serraino, D.; Olshan, A.F.; Zevallos, J.P.; Levi, F.; Zhang, Z.F.; Morgenstern, H.; Garavello, W.; et al. Shared and study-specific dietary patterns and head and neck cancer risk in an international consortium. Epidemiology 2019, 30, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Galeone, C.; Turati, F.; Zhang, Z.-F.; Guercio, V.; Tavani, A.; Serraino, D.; Brennan, P.; Fabianova, E.; Lissowska, J.; Mates, D.; et al. Relation of allium vegetables intake with head and neck cancers: Evidence from the INHANCE consortium. Mol. Nutr. Food Res. 2015, 59, 1641–1650. [Google Scholar] [CrossRef] [PubMed]
- Leoncini, E.; Edefonti, V.; Hashibe, M.; Parpinel, M.; Cadoni, G.; Ferraroni, M.; Serraino, D.; Matsuo, K.; Olshan, A.F.; Zevallos, J.P.; et al. Carotenoid intake and head and neck cancer: A pooled analysis in the international head and neck cancer epidemiology consortium. Eur. J. Epidemiol. 2016, 31, 369–383. [Google Scholar] [CrossRef]
- Leoncini, E.; Nedovic, D.; Panic, N.; Pastorino, R.; Edefonti, V.; Boccia, S. Carotenoid Intake from Natural Sources and Head and Neck Cancer: A Systematic Review and Meta-analysis of Epidemiological Studies. Cancer Epidemiol. Biomark. Prev. 2015, 24, 1003–1011. [Google Scholar] [CrossRef]
- Edefonti, V.; Hashibe, M.; Ambrogi, F.; Parpinel, M.; Bravi, F.; Talamini, R.; Levi, F.; Yu, G.; Morgenstern, H.; Kelsey, K.; et al. Nutrient-based dietary patterns and the risk of head and neck cancer: A pooled analysis in the International Head and Neck Cancer Epidemiology consortium. Ann. Oncol. 2012, 23, 1869–1880. [Google Scholar] [CrossRef]
- Bidoli, E.; Bosetti, C.; La Vecchia, C.; Levi, F.; Parpinel, M.; Talamini, R.; Negri, E.; Dal Maso, L.; Franceschi, S. Micronutrients and laryngeal cancer risk in Italy and Switzerland: A case–control study. Cancer Causes Control 2003, 14, 477–484. [Google Scholar] [CrossRef]
- Bravi, F.; Bosetti, C.; Filomeno, M.; Levi, F.; Garavello, W.; Galimberti, S.; Negri, E.; La Vecchia, C. Foods, nutrients and the risk of oral and pharyngeal cancer. Br. J. Cancer 2013, 109, 2904–2910. [Google Scholar] [CrossRef]
- Polesel, J.; Negri, E.; Serraino, D.; Parpinel, M.; Barzan, L.; Libra, M.; Bosetti, C.; Garavello, W.; Montella, M.; La Vecchia, C.; et al. Dietary intakes of carotenoids and other nutrients in the risk of nasopharyngeal carcinoma: A case–control study in Italy. Br. J. Cancer 2012, 107, 1580–1583. [Google Scholar] [CrossRef]
- Kasum, C.M.; Jacobs, D.R., Jr.; Nicodemus, K.; Folsom, A.R. Dietary risk factors for upper aerodigestive tract cancers. Int. J. Cancer 2002, 99, 267–272. [Google Scholar] [CrossRef]
- Suzuki, T.; Wakai, K.; Matsuo, K.; Hirose, K.; Ito, H.; Kuriki, K.; Sato, S.; Ueda, R.; Hasegawa, Y.; Tajima, K. Effect of dietary antioxidants and risk of oral, pharyngeal and laryngeal squamous cell carcinoma according to smoking and drinking habits. Cancer Sci. 2006, 97, 760–767. [Google Scholar] [CrossRef] [PubMed]
- Petridou, E.; Zavras, A.I.; Lefatzis, D.; Dessypris, N.; Laskaris, G.; Dokianakis, G.; Segas, J.; Douglas, C.W.; Diehl, S.R.; Trichopoulos, D. The role of diet and specific micronutrients in the etiology of oral carcinoma. Cancer 2002, 94, 2981–2988. [Google Scholar] [CrossRef] [PubMed]
- de Munter, L.; Maasland, D.H.E.; van den Brandt, P.A.; Kremer, B.; Schouten, L.J. Vitamin and carotenoid intake and risk of head-neck cancer subtypes in the Netherlands Cohort Study. Am. J. Clin. Nutr. 2015, 102, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Yan, L.; Lin, L.; Liu, F.; Qiu, Y.; Wang, J.; Wu, J.; Liu, F.; Huang, J.; Cai, L.; et al. Dietary score and the risk of oral cancer: A case-control study in southeast China. Oncotarget 2017, 8, 34610–34616. [Google Scholar] [CrossRef][Green Version]
- Dyer, A.R.; Elliott, P.; Stamler, J.; Chan, Q.; Ueshima, H.; Zhou, B.F.; INTERMAP Research Group. Dietary intake in male and female smokers, ex-smokers, and never smokers: The INTERMAP study. J. Hum. Hypertens. 2003, 17, 641–654. [Google Scholar] [CrossRef]
- Northrop-Clewes, C.A.; Thurnham, D.I. Monitoring micronutrients in cigarette smokers. Clin. Chim. Acta. 2007, 377, 14–38. [Google Scholar] [CrossRef]
- Farshadpour, F.; Hordijk, G.J.; Koole, R.; Slootweg, P.J. Non-smoking and non-drinking patients with head and neck squamous cell carcinoma: A distinct population. Oral Dis. 2007, 13, 239–243. [Google Scholar] [CrossRef]
- Serra-Majem, L. Head and neck cancer: Smoking, drinking, eating and …sexual practices. Eur. J. Epidemiol. 2016, 31, 333–335. [Google Scholar] [CrossRef]
- Meyer, M.S.; Applebaum, K.M.; Furniss, C.S.; Peters, E.S.; Luckett, B.G.; Smith, J.F.; Bryan, J.; McClean, M.D.; Marsit, C.; Kelsey, K.T. Human papillomavirus-16 modifies the association between fruit consumption and head and neck squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 3419–3426. [Google Scholar] [CrossRef]
- Toporcov, T.N.; Znaor, A.; Zhang, Z.F.; Yu, G.P.; Winn, D.M.; Wei, Q.; Vilensky, M.; Vaughan, T.; Thomson, P.; Talamini, R.; et al. Risk factors for head and neck cancer in young adults: A pooled analysis in the INHANCE consortium. Int. J. Epidemiol. 2015, 44, 169–185. [Google Scholar] [CrossRef]
- Lang, S.; Schimansky, S.; Beynon, R.; Penfold, C.; Davies, A.; Waylen, A.; Thomas, S.; Pring, M.; Pawlita, M.; Waterboer, T.; et al. Dietary behaviors and survival in people with head and neck cancer: Results from Head and Neck 5000. Head Neck. 2019, 41, 2074–2084. [Google Scholar] [CrossRef] [PubMed]
- Brewer, M.S. Natural Antioxidants: Sources, compounds, mechanisms of action and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Goodman, G.E.; Thornquist, M.D.; Balmes, J.; Cullen, M.R.; Meyskens, F.L., Jr.; Omenn, G.S.; Valanis, B.; Williams, J.H., Jr. The beta-carotene and retinol efficacy trial: Incidence of lung cancer and cardiovascular disease mortality during 6-year follow-up after stopping beta-carotene and retinol supplements. J. Natl. Cancer Inst. 2004, 96, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Nejatinamini, S.; Debenham, B.J.; Clugston, R.D.; Mawani, A.; Parliament, M.; Wismer, W.V.; Mazurak, V.C. Poor Vitamin Status is Associated with Skeletal Muscle Loss and Mucositis in Head and Neck Cancer Patients. Nutrients 2018, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Mason, H.; DeRubeis, M.B.; Burke, N.; Shannon, M.; Karsies, D.; Wolf, G.; Eisbruchl, A.; Worden, F. Symptom management during and after treatment with concurrent chemoradiotherapy for oropharyngeal cancer: A review of the literature and areas for future research. World J. Clin. Oncol. 2016, 7, 220–226. [Google Scholar] [CrossRef]
- Crowder, S.L.; Douglas, K.G.; Pepino, M.Y.; Sarma, K.P.; Arthur, A.E. Nutrition impact symptoms and associated outcomes in post-chemoradiotherapy head and neck cancer survivors: A systematic review. J. Cancer Surviv. 2018, 12, 479–494. [Google Scholar] [CrossRef]
- Payakachat, N.; Ounpraseuth, S.; Suen, J.Y. Late complications and long-term quality of life for survivors (>5 years) with history of head and neck cancer. Head Neck. 2013, 35, 819–825. [Google Scholar] [CrossRef]
- Ganzer, H.; Rothpletz-Puglia, P.; Byham-Gray, L.; Murphy, B.A.; Touger-Decker, R. The eating experience in long-term survivors of head and neck cancer: A mixed-methods study. Support Care Cancer 2015, 23, 3257–3268. [Google Scholar] [CrossRef]
- Arvin, C.G.; Kiraly, L.; McCarthy, M.S.; Martindale, R. Nutrition Support and Therapy in Patients with Head and Neck Squamous Cell Carcinoma. Curr. Gastroenterol. Rep. 2012, 14, 349–355. [Google Scholar] [CrossRef]
- Andic, F.; Garipagaoglu, M.; Yurdakonar, E.; Tuncel, N.; Kucuk, O. Lycopene in the prevention of gastrointestinal toxicity of radiotherapy. Nutr. Cancer 2009, 61, 748–784. [Google Scholar] [CrossRef]
- Cartmill, B.; Cornwell, P.; Ward, E.; Davidson, W.; Porceddu, S. Long-term functional outcomes and patient perspective following altered fractionation radiotherapy with concomitant boost for oropharyngeal cancer. Dysphagia 2012, 27, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Crowder, S.L.; Najam, N.; Sarma, K.P.; Fiese, B.H.; Arthur, A.E. Head and Neck Cancer Survivors’ Experiences with Chronic Nutrition Impact Symptom Burden after Radiation: A Qualitative Study. J. Acad. Nutr. Diet. 2020, 120, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Filomeno, M.; Bosetti, C.; Garavello, W.; Levi, F.; Galeone, C.; Negri, E.; La Vecchia, C. The role of a Mediterranean diet on the risk of oral and pharyngeal cancer. Br. J. Cancer 2014, 111, 981–986. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Lin, X.-L.; Fan, Y.-Y.; Liu, Y.-T.; Zhang, X.-L.; Lu, Y.-K.; Xu, C.-H.; Chen, Y.-M. Diet Quality Scores and Risk of Nasopharyngeal Carcinoma in Chinese Adults: A Case-Control Study. Nutrients 2016, 8, 112. [Google Scholar] [CrossRef]
- Freedman, N.D.; Park, Y.; Subar, A.F.; Hollenbeck, A.R.; Leitzmann, M.F.; Schatzkin, A.; Abnet, C.C. Fruit and vegetable intake and head and neck cancer risk in a large United States prospective cohort study. Int. J. Cancer 2008, 122, 2330–2336. [Google Scholar] [CrossRef]
- Al-Delaimy, W.K.; Ferrari, P.; Slimani, N.; Pala, V.; Johansson, I.; Nilsson, S.; Mattisson, I.; Wirfalt, E.; Galasso, R.; Palli, D.; et al. Plasma carotenoids as biomarkers of intake of fruits and vegetables: Individual-level correlations in the European Prospective Investigation into Cancer and Nutrition (EPIC). Eur. J. Clin. Nutr. 2005, 59, 1387–1396. [Google Scholar] [CrossRef]
- Böhm, V.; Lietz, G.; Dagliad, M.; Olmedilla-Alonso, B.; Phelan, D.; Reboul, E.; Bánati, D.; Borel, P.; Corte-Real, J.; de Lera, A.R.; et al. From carotenoid intake to carotenoid blood and tissue concentrations – implications for dietary intake recommendations. Nutr. Rev. 2021, 79, 544–573. [Google Scholar] [CrossRef]
- Lee, T.Y.; Tseng, H.H. The Potential of Phytochemicals in Oral Cancer Prevention and Therapy: A Review of the Evidence. Biomolecules. 2020, 10, 1150. [Google Scholar] [CrossRef]
- Nagao, T.; Ikeda, N.; Warnakulasuriya, S.; Fukano, H.; Yuasa, H.; Yano, M.; Miyazaki, H.; Ito, Y. Serum antioxidant micronutrients and the risk of oral leukoplakia among Japanese. Oral Oncol. 2000, 36, 466–470. [Google Scholar] [CrossRef]
- Sakhi, A.K.; Russnes, K.M.; Thoresen, M.; Bastani, N.E.; Karlsen, A.; Smeland, S.; Blomhoff, R. Pre-radiotherapy plasma carotenoids and markers of oxidative stress are associated with survival in head and neck squamous cell carcinoma patients: A prospective study. BMC Cancer 2009, 9, 458. [Google Scholar] [CrossRef]
- Meyer, F.; Bairati, I.; Jobin, E.; Gelinas, M.; Fortin, A.; Nabid, A.; Têtu, B. Acute adverse effects of radiation therapy and local recurrence in relation to dietary and plasma beta carotene and alpha tocopherol in head and neck cancer patients. Nutr. Cancer 2007, 59, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Arthur, A.E.; Peterson, K.E.; Shen, J.; Djuric, Z.; Taylor, J.M.G.; Hebert, J.R.; Duffy, S.A.; Peterson, L.A.; Bellile, E.L.; Whitfield, J.R.; et al. Diet and proinflammatory cytokine levels in head and neck squamous cell carcinoma. Cancer 2014, 120, 2704–2712. [Google Scholar] [CrossRef] [PubMed]
- Galland, L. Diet and inflammation. Nutr. Clin. Pract. 2010, 25, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.A.; Teknos, T.; Taylor, J.M.; Fowler, K.E.; Islam, M.; Wolf, G.T.; McLean, S.; Ghanem, T.A.; Terrell, J.E. Health behaviors predict higher interleukin-6 levels among patients newly diagnosed with head and neck squamous cell carcinoma. Cancer Epidemiol. Biomark. Prev. 2013, 22, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Kim, J.H.; Kannappan, R.; Reuter, S.; Dougherty, P.M.; Aggarwal, B.B. Role of nuclear factor κB-mediated inflammatory pathways in cancer-related symptoms and their regulation by nutritional agents. Exp. Biol. Med. 2011, 236, 658–671. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Sundaram, C.; Reuter, S.; Aggarwal, B.B. Inhibiting NF-κB activation by small molecules as a therapeutic strategy. Biochim. Biophys. Acta 2010, 1799, 775–787. [Google Scholar] [CrossRef]
- Hua, R.; Liang, G.; Yang, F. Meta-Analysis of the Association between Dietary Inflammatory Index (DII) and Upper Aerodigestive Tract Cancer Risk. Medicine 2020, 99, e19879. [Google Scholar] [CrossRef]
- Liu, C.; Marioni, R.E.; Hedman, Å.K.; Pfeiffer, L.; Tsai, P.C.; Reynolds, L.M.; Just, A.C.; Duan, Q.; Boer, C.G.; Tanaka, T.; et al. A DNA methylation biomarker of alcohol consumption. Mol. Psychiatry 2018, 23, 422–433. [Google Scholar] [CrossRef]
- Perrier, F.; Viallon, V.; Ambatipudi, S.; Ghantous, A.; Cuenin, C.; Hernandez-Vargas, H.; Chajès, V.; Baglietto, L.; Matejcic, M.; Moreno-Macias, H.; et al. Association of leukocyte DNA methylation changes with dietary folate and alcohol intake in the EPIC study. Clin. Epigenetics. 2019, 11, 57. [Google Scholar] [CrossRef]
- Smith, I.M.; Mydlarz, W.K.; Mithani, S.K.; Califano, J.A. DNA global hypomethylation in squamous cell head and neck cancer associated with smoking, alcohol consumption and stage. Int. J. Cancer 2007, 121, 1724–1728. [Google Scholar] [CrossRef]
- Kim, H.J.; Maiti, P.; Barrientos, A. Mitochondrial ribosomes in cancer. Semin. Cancer Biol. 2017, 47, 67–81. [Google Scholar] [CrossRef] [PubMed]
- Arthur, A.E.; Bellile, E.L.; Rozek, L.S.; Peterson, K.E.; Ren, J.; Harris, E.; Mueller, C.; Jolly, S.; Peterson, L.A.; Wolf, G.T.; et al. Pretreatment serum xanthophyll concentrations as predictors of head and neck cancer recurrence and survival. Head Neck. 2016, 38 (Suppl. 1), E1591–E1597. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.Y.; Lee, B.J.; Chang, P.S.; Hsiao, H.Y.; Hsu, L.P.; Chang, C.H.; Lin, P.T. The risks of ubiquinone and beta-carotene deficiency and metabolic disorders in patients with oral cancer. BMC Cancer 2020, 20, 310. [Google Scholar] [CrossRef] [PubMed]
- Mayne, S.T.; Cartmel, B.; Lin, H.; Zheng, T.; Goodwin, W.J. Low Plasma Lycopene Concentration is Associated with Increased Mortality in a Cohort of Patients with Prior Oral, Pharynx or Larynx Cancers. J. Am. Coll. Nutr. 2004, 23, 34–42. [Google Scholar] [CrossRef]
- Meyer, F.; Bairati, I.; Fortin, A.; Gélinas, M.; Nabid, A.; Brochet, F.; Têtu, B. Interaction between antioxidant vitamin supplementation and cigarette smoking during radiation therapy in relation to long-term effects on recurrence and mortality: A randomized trial among head and neck cancer patients. Int. J. Cancer 2008, 122, 1679–1683. [Google Scholar] [CrossRef]
- Fakhry, C.; Westra, W.H.; Li, S.; Cmelak, A.; Ridge, J.A.; Pinto, H.; Forastiere, A.; Gillison, M.L. Improved survival of patients with human papillomavirus-positive head and neck squamous cell carcinoma in a prospective clinical trial. J. Natl. Cancer Inst. 2008, 100, 261–269. [Google Scholar] [CrossRef]
- Arthur, A.E.; Duffy, S.A.; Sanchez, G.I.; Gruber, S.B.; Terrell, J.E.; Hebert, J.R.; Light, E.; Bradford, C.R.; D’Silva, N.J.; Carey, T.E.; et al. Higher micronutrient intake is associated with human papillomavirus-positive head and neck cancer: A case-only analysis. Nutr. Cancer 2011, 63, 734–742. [Google Scholar] [CrossRef]
- Cartmel, B.; Bowen, D.; Ross, D.; Johnson, E.; Mayne, S.T. A randomized trial of an intervention to increase fruit and vegetable intake in curatively treated patients with early-stage head and neck cancer. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2848–2854. [Google Scholar] [CrossRef]
- Silveira, B.K.S.; Oliveira, T.M.S.; Andrade, P.A.; Hermsdorff, H.H.M.; Rosa, C.O.B.; Franceschini, S.D.C.C. Dietary Pattern and Macronutrients Profile on the Variation of Inflammatory Biomarkers: Scientific Update. Cardiol. Res. Pract. 2018, 2018, m4762575. [Google Scholar] [CrossRef]
- Smidowicz, A.; Regula, J. Effect of nutritional status and dietary patterns on human serum C-reactive protein and interleukin-6 concentrations. Adv. Nutr. 2015, 6, 738–747. [Google Scholar] [CrossRef]
- Masuda, M.; Toh, S.; Koike, K.; Kuratomi, J.; Suzui, M.; Deguchi, A.; Komiyama, S.; Weinstein, I.B. The roles of JNK1 and Stat3 in the response of head and neck cancer cell lines to combined treatment with all-trans-retinoic acid and 5-fluorouracil. Jpn. J. Cancer Res. 2002, 93, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Masuda, M.; Wakasaki, T.; Suzui, M.; Toh, S.; Joe, A.K.; Weinstein, I.B. Stat3 orchestrates tumor development and progression: The Achilles’ heel of head and neck cancers? Curr. Cancer Drug Targets 2010, 10, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, M.; Suzui, M.; Deguchi, A.; Lim, J.T.; Weinstein, I.B. Effects of acyclic retinoid on growth, cell cycle control, epidermal growth factor receptor signaling, and gene expression in human squamous cell carcinoma cells. Clin. Cancer Res. 2004, 10, 1130–1140. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, E.; Mielicki, W. All-trans retinoic acid (ATRA) in prevention and cancer therapy. Postepy Hig. Med. Dosw. 2010, 64, 284–290. [Google Scholar]
- Soprano, D.R.; Qin, P.; Soprano, K.J. Retinoid acid receptors and cancers. Annu. Rev. Nutr. 2004, 24, 201–221. [Google Scholar] [CrossRef]
- Rashid, A.; Duan, X.; Gao, F.; Yang, M.; Yen, A. Roscovitine enhances all-trans retinoic acid (ATRA)-induced nuclear enrichment of an ensemble of activated signaling molecules and augments ATRA-induced myeloid cell differentiation. Oncotarget 2020, 11, 1017–1036. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Starska-Kowarska, K. Dietary Carotenoids in Head and Neck Cancer—Molecular and Clinical Implications. Nutrients 2022, 14, 531. https://doi.org/10.3390/nu14030531
Starska-Kowarska K. Dietary Carotenoids in Head and Neck Cancer—Molecular and Clinical Implications. Nutrients. 2022; 14(3):531. https://doi.org/10.3390/nu14030531
Chicago/Turabian StyleStarska-Kowarska, Katarzyna. 2022. "Dietary Carotenoids in Head and Neck Cancer—Molecular and Clinical Implications" Nutrients 14, no. 3: 531. https://doi.org/10.3390/nu14030531
APA StyleStarska-Kowarska, K. (2022). Dietary Carotenoids in Head and Neck Cancer—Molecular and Clinical Implications. Nutrients, 14(3), 531. https://doi.org/10.3390/nu14030531