
Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway


Abstract
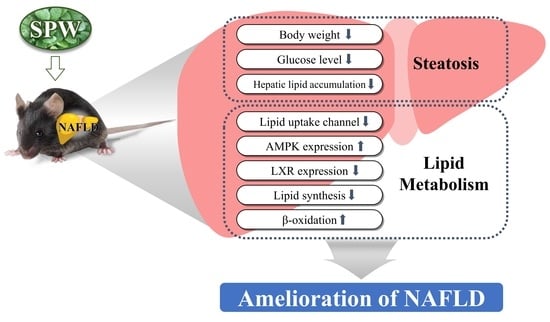
1. Introduction
2. Materials and Methods
2.1. Preparation of SPW
2.2. Measurement of Total Phenolic Compounds and Flavonoid Content
2.3. Cell Culture and Cell Viability Assay
2.4. Oil Red O Staining
2.5. AdipoRed Assay
2.6. Animals and Experimental Design
2.7. Oral Glucose Tolerance Test (OGTT)
2.8. Histological Analysis of the Liver
2.9. Biochemical Assays
2.10. Total RNA Isolation and Real-Time Polymerase Chain Reaction
2.11. Statistical Analysis
3. Results
3.1. Total Phenolic Compounds and Flavonoid Content of SPW was Determinated
3.2. SPW Ameliorates FFA-Induced Lipid Accumulation
3.3. SPW Regulates Lipid Metabolism via the AMPK Pathway in FFA-Induced Hepatic Steatosis
3.4. SPW Improves HFD-Induced Body Weight Gain
3.5. SPW Improves HFD-Induced Glucose Level
3.6. SPW Ameliorates HFD-Induced Hepatic Steatosis and Liver Injury
3.7. SPW Regulates Lipid Metabolism via AMPK Pathway in HFD-Induced Hepatic Steatosis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Im, H.J.; Ahn, Y.C.; Wang, J.-H.; Lee, M.M.; Son, C.G. Systematic review on the prevalence of nonalcoholic fatty liver disease in South Korea. Clin. Res. Hepatol. Gastroenterol. 2021, 45, 101526. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; De, A.; Chowdhury, A. Epidemiology of non-alcoholic and alcoholic fatty liver diseases. Transl. Gastroenterol. Hepatol. 2020, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metabolism 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Karim, M.; Al-Mahtab, M.; Rahman, S.; Debnath, C. Non-alcoholic Fatty Liver Disease (NAFLD)—A Review. Mymensingh Med. J. MMJ 2015, 24, 873–880. [Google Scholar]
- He, J.; Lee, J.H.; Febbraio, M.; Xie, W. The emerging roles of fatty acid translocase/CD36 and the aryl hydrocarbon receptor in fatty liver disease. Exp. Biol. Med. 2011, 236, 1116–1121. [Google Scholar] [CrossRef]
- Enooku, K.; Tsutsumi, T.; Kondo, M.; Fujiwara, N.; Sasako, T.; Shibahara, J.; Kado, A.; Okushin, K.; Fujinaga, H.; Nakagomi, R. Hepatic FATP5 expression is associated with histological progression and loss of hepatic fat in NAFLD patients. J. Gastroenterol. 2020, 55, 227–243. [Google Scholar] [CrossRef]
- Mun, J.; Kim, S.; Yoon, H.G.; You, Y.; Kim, O.K.; Choi, K.C.; Lee, Y.H.; Lee, J.; Park, J.; Jun, W. Water Extract of Curcuma longa L. Ameliorates Non-Alcoholic Fatty Liver Disease. Nutrients 2019, 11, 2536. [Google Scholar] [CrossRef]
- Oliveras-Ferraros, C.; Vazquez-Martin, A.; Fernández-Real, J.M.; Menendez, J.A. AMPK-sensed cellular energy state regulates the release of extracellular Fatty Acid Synthase. Biochem. Biophys. Res. Commun. 2009, 378, 488–493. [Google Scholar] [CrossRef]
- Hu, Y.; Yin, F.; Liu, Z.; Xie, H.; Xu, Y.; Zhou, D.; Zhu, B. Acerola polysaccharides ameliorate high-fat diet-induced non-alcoholic fatty liver disease through reduction of lipogenesis and improvement of mitochondrial functions in mice. Food Funct. 2020, 11, 1037–1048. [Google Scholar] [CrossRef]
- Lund, E.G.; Peterson, L.B.; Adams, A.D.; Lam, M.H.; Burton, C.A.; Chin, J.; Guo, Q.; Huang, S.; Latham, M.; Lopez, J.C.; et al. Different roles of liver X receptor alpha and beta in lipid metabolism: Effects of an alpha-selective and a dual agonist in mice deficient in each subtype. Biochem. Pharmacol. 2006, 71, 453–463. [Google Scholar] [CrossRef]
- Liang, Y.-Y.; Wan, X.-H.; Niu, F.-J.; Xie, S.-M.; Guo, H.; Yang, Y.-Y.; Guo, L.-Y.; Zhou, C.-Z. Salvia plebeia R. Br.: An overview about its traditional uses, chemical constituents, pharmacology and modern applications. Biomed. Pharmacother. 2020, 121, 109589. [Google Scholar] [CrossRef] [PubMed]
- Seo, H.W.; Suh, J.H.; Kyung, J.-S.; Jang, K.H.; So, S.-H. Subacute oral toxicity and bacterial mutagenicity study of a mixture of korean red ginseng (Panax ginseng CA Meyer) and Salvia plebeia R. Br. Extracts. Toxicol. Res. 2019, 35, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.; Ha, T.K.Q.; Lee, C.; Li, W.; Oh, W.-K.; Shim, S.H. Antiviral activities of compounds from aerial parts of Salvia plebeia R. Br. J. Ethnopharmacol. 2016, 192, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Qu, X.J.; Xia, X.; Wang, Y.S.; Song, M.J.; Liu, L.L.; Xie, Y.Y.; Cheng, Y.N.; Liu, X.J.; Qiu, L.L.; Xiang, L.; et al. Protective effects of Salvia plebeia compound homoplantaginin on hepatocyte injury. Food Chem. Toxicol. 2009, 47, 1710–1715. [Google Scholar] [CrossRef]
- Lee, G.T.; Duan, C.H.; Lee, J.-N.; Lee, K.-S.; Hong, J.-T.; Lee, K.-K. Phytochemical constituents from Salvia plebeia. Nat. Prod. Sci. 2010, 16, 207–210. [Google Scholar]
- Weng, X.; Wang, W. Antioxidant activity of compounds isolated from Salvia plebeia. Food Chem. 2000, 71, 489–493. [Google Scholar] [CrossRef]
- Choi, S.I.; Cho, I.H.; Han, S.H.; Jeon, Y.J.; Choi, J.G.; Kim, J.S.; Lee, J.H. Antiobesity Effects of Salvia plebeia R. Br. Extract in High-Fat Diet-Induced Obese Mice. J. Med. Food 2016, 19, 1048–1056. [Google Scholar] [CrossRef]
- Won, H.R. Effect of Salvia plebeia water extract on antioxidant activity and lipid composition of rats fed a high fat-high cholesterol diet. Korean J. Community Living Sci. 2016, 27, 233–243. [Google Scholar] [CrossRef]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Atanassova, M.; Georgieva, S.; Ivancheva, K. Total phenolic and total flavonoid contents, antioxidant capacity and biological contaminants in medicinal herbs. J. Univ. Chem. Technol. Metall. 2011, 46, 81–88. [Google Scholar]
- Roehm, N.W.; Rodgers, G.H.; Hatfield, S.M.; Glasebrook, A.L. An improved colorimetric assay for cell proliferation and viability utilizing the tetrazolium salt XTT. J. Immunol. Methods 1991, 142, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Iverson, S.J.; Lang, S.L.; Cooper, M.H. Comparison of the Bligh and Dyer and Folch methods for total lipid determination in a broad range of marine tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Mu, H.-N.; Zhou, Q.; Yang, R.-Y.; Tang, W.-Q.; Li, H.-X.; Wang, S.-M.; Li, J.; Chen, W.-X.; Dong, J. Caffeic acid prevents non-alcoholic fatty liver disease induced by a high-fat diet through gut microbiota modulation in mice. Food Res. Int. 2021, 143, 110240. [Google Scholar] [CrossRef]
- Balachander, G.J.; Subramanian, S.; Ilango, K. Rosmarinic acid attenuates hepatic steatosis by modulating ER stress and autophagy in oleic acid-induced HepG2 cells. RSC Adv. 2018, 8, 26656–26663. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, R.; Shen, Z.; Cai, G. Combination of luteolin and lycopene effectively protect against the “two-hit” in NAFLD through Sirt1/AMPK signal pathway. Life Sci. 2020, 256, 117990. [Google Scholar] [CrossRef] [PubMed]
- Han, L.P.; Sun, B.; Li, C.J.; Xie, Y.; Chen, L.M. Effect of celastrol on toll-like receptor 4-mediated inflammatory response in free fatty acid-induced HepG2 cells. Int. J. Mol. Med. 2018, 42, 2053–2061. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-C.; Chen, H.-J.; Huang, B.-M.; Chen, Y.-C.; Chang, C.-F. Polyphenol-Rich Extracts from Toona sinensis Bark and Fruit Ameliorate Free Fatty Acid-Induced Lipogenesis through AMPK and LC3 Pathways. J. Clin. Med. 2019, 8, 1664. [Google Scholar] [CrossRef] [PubMed]
- Im, A.-R.; Kim, Y.H.; Lee, H.W.; Song, K.H. Water extract of Dolichos lablab attenuates hepatic lipid accumulation in a cellular nonalcoholic fatty liver disease model. J. Med. Food 2016, 19, 495–503. [Google Scholar] [CrossRef]
- Mitmesser, S.H.; Carr, T.P. Trans fatty acids alter the lipid composition and size of apoB-100-containing lipoproteins secreted by HepG2 cells. J. Nutr. Biochem. 2005, 16, 178–183. [Google Scholar] [CrossRef]
- Gaggini, M.; Morelli, M.; Buzzigoli, E.; DeFronzo, R.A.; Bugianesi, E.; Gastaldelli, A. Non-alcoholic fatty liver disease (NAFLD) and its connection with insulin resistance, dyslipidemia, atherosclerosis and coronary heart disease. Nutrients 2013, 5, 1544–1560. [Google Scholar] [CrossRef]
- Goorden, S.; Buffart, T.E.; Bakker, A.; Buijs, M.M. Liver disorders in adults: ALT and AST. Ned. Tijdschr. Voor Geneeskd. 2013, 157, A6443. [Google Scholar]
- Chiang, J.Y.; Ferrell, J.M. Bile acid metabolism in liver pathobiology. Gene Expr. 2018, 18, 71. [Google Scholar] [CrossRef] [PubMed]
- Rui, L. Energy metabolism in the liver. Compr. Physiol. 2014, 4, 177–179. [Google Scholar] [PubMed]
- Chen, L.; Chen, X.-W.; Huang, X.; Song, B.-L.; Wang, Y.; Wang, Y. Regulation of glucose and lipid metabolism in health and disease. Sci. China Life Sci. 2019, 62, 1420–1458. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Shen, L.; Tan, Z.; Zhang, P.; Zhao, X.; Xu, Y.; Gan, M.; Yang, Q.; Ma, J.; Jiang, A.A. Betaine supplementation enhances lipid metabolism and improves insulin resistance in mice fed a high-fat diet. Nutrients 2018, 10, 131. [Google Scholar] [CrossRef]
- Sekizkardes, H.; Chung, S.T.; Chacko, S.; Haymond, M.W.; Startzell, M.; Walter, M.; Walter, P.J.; Lightbourne, M.; Brown, R.J. Free fatty acid processing diverges in human pathologic insulin resistance conditions. J. Clin. Investig. 2020, 130, 3592–3602. [Google Scholar] [CrossRef]
- Li, H.; Yoo, W.; Park, H.M.; Lim, S.Y.; Shin, D.H.; Kim, S.; Park, H.Y.; Jeong, T.S. Arazyme Suppresses Hepatic Steatosis and Steatohepatitis in Diet-Induced Non-Alcoholic Fatty Liver Disease-Like Mouse Model. Int. J. Mol. Sci. 2019, 20, 2325. [Google Scholar] [CrossRef]
- Oligschlaeger, Y.; Shiri-Sverdlov, R. NAFLD Preclinical Models: More than a Handful, Less of a Concern? Biomedicines 2020, 8, 28. [Google Scholar] [CrossRef]
- Castro, R.E.; Diehl, A.M. Towards a definite mouse model of NAFLD. J. Hepatol. 2018, 69, 272–274. [Google Scholar] [CrossRef]
- Nakamura, A.; Terauchi, Y. Lessons from mouse models of high-fat diet-induced NAFLD. Int. J. Mol. Sci. 2013, 14, 21240–21257. [Google Scholar] [CrossRef]
- Prill, S.; Caddeo, A.; Baselli, G.; Jamialahmadi, O.; Dongiovanni, P.; Rametta, R.; Kanebratt, K.P.; Pujia, A.; Pingitore, P.; Mancina, R.M. The TM6SF2 E167K genetic variant induces lipid biosynthesis and reduces apolipoprotein B secretion in human hepatic 3D spheroids. Sci. Rep. 2019, 9, 11585. [Google Scholar] [CrossRef] [PubMed]
- Mundi, M.S.; Velapati, S.; Patel, J.; Kellogg, T.A.; Abu Dayyeh, B.K.; Hurt, R.T. Evolution of NAFLD and its management. Nutr. Clin. Pract. 2020, 35, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Utzschneider, K.M.; Kahn, S.E.; Polidori, D.C. Hepatic insulin extraction in NAFLD is related to insulin resistance rather than liver fat content. J. Clin. Endocrinol. Metab. 2019, 104, 1855–1865. [Google Scholar] [CrossRef]
- Wilson, C.G.; Tran, J.L.; Erion, D.M.; Vera, N.B.; Febbraio, M.; Weiss, E.J. Hepatocyte-Specific Disruption of CD36 Attenuates Fatty Liver and Improves Insulin Sensitivity in HFD-Fed Mice. Endocrinology 2016, 157, 570–585. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Wang, H.; Zhang, M.; Zhang, M.; Peng, R.; Zhao, Q.; Liu, J. FATP2-targeted therapies—A role beyond fatty liver disease. Pharmacol. Res. 2020, 161, 105228. [Google Scholar] [CrossRef] [PubMed]
- Kumari, A.; Pathak, D.P.; Asthana, S. Bile acids mediated potential functional interaction between FXR and FATP5 in the regulation of Lipid Metabolism. Int. J. Biol. Sci. 2020, 16, 2308. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, J.P.; Eckman, K.; Lee, Y.K.; Abdelmegeed, M.A.; Esterle, A.; Chilian, W.M.; Chiang, J.Y.; Song, B.-J. Chapter Five—Eicosanoids in Metabolic Syndrome. In Advances in Pharmacology; Webb, D.R., Ed.; Academic Press: Cambridge, MA, USA, 2013; Volume 66, pp. 157–266. [Google Scholar]
- Jeon, S.-M. Regulation and function of AMPK in physiology and diseases. Exp. Mol. Med. 2016, 48, e245. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.J.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef]
- Foretz, M.; Viollet, B. Regulation of hepatic metabolism by AMPK. J. Hepatol. 2011, 54, 827–829. [Google Scholar] [CrossRef]
- Calkin, A.C.; Tontonoz, P. Transcriptional integration of metabolism by the nuclear sterol-activated receptors LXR and FXR. Nat. Rev. Mol. Cell Biol. 2012, 13, 213–224. [Google Scholar] [CrossRef]
- Komati, R.; Spadoni, D.; Zheng, S.; Sridhar, J.; Riley, K.E.; Wang, G. Ligands of Therapeutic Utility for the Liver X Receptors. Molecules 2017, 22, 88. [Google Scholar] [CrossRef] [PubMed]
- Kratzer, A.; Buchebner, M.; Pfeifer, T.; Becker, T.M.; Uray, G.; Miyazaki, M.; Miyazaki-Anzai, S.; Ebner, B.; Chandak, P.G.; Kadam, R.S. Synthetic LXR agonist attenuates plaque formation in apoE-/-mice without inducing liver steatosis and hypertriglyceridemias. J. Lipid Res. 2009, 50, 312–326. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′ to 3′) | |
|---|---|---|
| hCD36 | Forward Reverse | 5′-TGGAACAGAGGCTGACAACT-3′ 5′-TTGATTTTTGATAGATATGGG-3′ |
| hFATP2 | Forward Reverse | 5′-CTTTCAGCACATTGCTGATTACCT-3′ 5′-CAGTGATCTCAATGGTGTCCTGTAT-3′ |
| hFATP5 | Forward Reverse | 5′-AGCTCCTGCGGTACTTGTGT-3′ 5′-AAGGTCTCCCACACATCAGC-3′ |
| hAMPK | Forward Reverse | 5′-GGCACCCTCCCATTTGATG-3′ 5′-ACACCCCCTCGGATCTTCTT-3′ |
| hAMPK | Forward Reverse | 5′-GGCACCCTCCCATTTGATG-3′ 5′-ACACCCCCTCGGATCTTCTT-3′ |
| hCPT-1 | Forward Reverse | 5′-TGTTGGGTATGCTGTTCATGACA-3′ 5′-GCGGCCTGGGTAGGAAGA-3′ |
| hPPARα | Forward Reverse | 5′-AACATCCAAGAGATTTCGCAATC-3′ 5′-CCGTAAAGCCAAAGCTTCCA-3′ |
| hSREBP-1c | Forward Reverse | 5′-CGGAACCATCTTGGCAACA-3′ 5′-GCCGGTTGATAGGCAGCTT-3′ |
| hFAS | Forward Reverse | 5′-CGCTCGGCATGGCTATCT-3′ 5′-CTCGTTGAAGAACGCATCCA-3′ |
| hACC | Forward Reverse | 5′-TGCAGATCTTAGCGGACCAA-3′ 5′-GCCTGCGTTGTACAGAGCAA-3′ |
| hβ-Actin | Forward Reverse | 5′-ACGGCCAGGTCATCACTATTG-3′ 5′-CAAGAAGGAAGGCTGGAAAAGA-3′ |
| mCD36 | Forward Reverse | 5′-TTGAAGGCATTCCCACGTATC-3′ 5′-CGGACCCGTTGGCAAA-3′ |
| mFATP2 | Forward Reverse | 5′-GGAACCACAGGTCTTCCAAA-3′ 5′-TAAAGTAGCCCCAACCACGA-3′ |
| mFATP5 | Forward Reverse | 5′-GGAACCACAGGTCTTCCAAA-3′ 5′-TAAAGTAGCCCCAACCACGA-3′ |
| mAMPK | Forward Reverse | 5′-TTCGTGCCGCCCCTTT-3′ 5′-GGTCAGCATGCCCACAAAA-3′ |
| mLXRα | Forward Reverse | 5′-CCTCTGGCTTCCATTACAAC-3′ 5′-CTTCTGACAGCACACACTC-3′ |
| mLXRβ | Forward Reverse | 5′-CACCATTGAGATCATGTTGC-3′ 5′-TTGATCCTCGTGTAGGAGAG-3′ |
| mCPT-1 | Forward Reverse | 5′-GTGACTGGTGGGAGGAATAC-3′ 5′-GAGCATCTCCATGGCGTAG-3′ |
| mPPARα | Forward Reverse | 5′-TGGCAAAAGGCAAGGAGAAG-3′ 5′-CCCTCTACATAGAACTGCAA-3′ |
| mSREBP-1c | Forward Reverse | 5′-TGGCTTGGTGATGCTATGTTG-3′ 5′-GACCATCAAGGCCCCTCAA-3′ |
| mFAS | Forward Reverse | 5′-GAAGTGTCTGGACTGTGTCATTTTTAC-3′ 5′-TTAATTGTGGGATCAGGAGAGCAT-3′ |
| mACC | Forward Reverse | 5′-TCCCCAAGTTCTTCACGTTCA-3′ 5′-CAGGCTCCAAGTGGCGATAA-3′ |
| mβ-Actin | Forward Reverse | 5′-AGCCATGTACGTAGCCATCC-3′ 5′-CTCTCAGCTGTGGTGGTGAA-3′ |
| Total Phenolic Compounds Content (mg GAE/100 g SPW) | Flavonoid Content (mg CE/100 g SPW) | |
|---|---|---|
| SPW | 6984.69 ± 138.25 | 5334.44 ± 172.78 |
| Groups | CON | HFD | HFD + SPW |
|---|---|---|---|
| Body mass (g) | |||
| Initial (A) | 22.78 ± 0.37 ns | 22.79 ± 0.33 | 22.76 ± 0.28 |
| Final (B) | 35.45 ± 0.66 c | 45.81 ± 0.67 a | 43.36 ± 0.61 b |
| Weight gain (B–A) | 12.67 ± 0.72 c | 23.10 ± 0.55 a | 20.61 ± 0.58 b |
| Food intake (g/day) | 3.23 ± 0.03 a | 2.52 ± 0.04 b | 2.66 ± 0.05 b |
| Energy intake (kcal/day) | 12.60 ± 0.13 ns | 13.19 ± 0.19 | 13.93 ± 0.25 |
| FER (%) | 4.62 ± 0.27 c | 10.61 ± 0.26 a | 9.19 ± 0.31 b |
| Liver weight (g) | 1.44 ± 0.02 b | 1.94 ± 0.05 a | 1.55 ± 0.12 b |
| Liver index (%) | 3.68 ± 0.04 b | 4.19 ± 0.10 a | 3.09 ± 0.21 c |
| Relative fat weight (g) | |||
| Perirenal fat | 0.75 ± 0.03 c | 1.39 ± 0.07 a | 1.08 ± 0.06 b |
| Epididymal fat | 1.49 ± 0.03 b | 2.12 ± 0.12 a | 1.62 ± 0.05 b |
| Groups | CON | HFD | HFD + SPW |
|---|---|---|---|
| Serum | |||
| TG (mg/dL) | 101.79 ± 1.05 c | 134.23 ± 3.51 a | 118.16 ± 1.91 b |
| TC (mg/dL) | 239.44 ± 1.27 b | 278.38 ± 5.56 a | 237.44 ± 3.22 b |
| HDL (mg/dL) | 122.12 ± 1.72 b | 95.75 ± 0.72 a | 119.55 ± 2.40 b |
| LDL (mg/dL) | 96.96 ± 2.06 b | 155.78 ± 5.13 a | 96.25 ± 4.79 b |
| NEFA (mEq/L) | 80.33 ± 2.66 b | 120.33 ± 5.61 a | 81.00 ± 3.22 b |
| Liver tissue | |||
| TG (mg/g liver) | 5.77 ± 0.35 c | 10.73 ± 0.07 a | 8.93 ± 0.05 b |
| TC (mg/g liver) | 2.80 ± 0.06 b | 4.16 ± 0.12 a | 2.75 ± 0.09 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bae, S.; Lee, Y.-H.; Lee, J.; Park, J.; Jun, W. Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway. Nutrients 2022, 14, 5379. https://doi.org/10.3390/nu14245379
Bae S, Lee Y-H, Lee J, Park J, Jun W. Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway. Nutrients. 2022; 14(24):5379. https://doi.org/10.3390/nu14245379
Chicago/Turabian StyleBae, Subin, Yoo-Hyun Lee, Jeongmin Lee, Jeongjin Park, and Woojin Jun. 2022. "Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway" Nutrients 14, no. 24: 5379. https://doi.org/10.3390/nu14245379
APA StyleBae, S., Lee, Y.-H., Lee, J., Park, J., & Jun, W. (2022). Salvia plebeia R. Br. Water Extract Ameliorates Hepatic Steatosis in a Non-Alcoholic Fatty Liver Disease Model by Regulating the AMPK Pathway. Nutrients, 14(24), 5379. https://doi.org/10.3390/nu14245379