
Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins

Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Animals
2.3. Experimental Design
2.4. Oral Glucose Tolerance Test (OGTT) and Insulin Tolerance Test (ITT)
2.5. Analysis of Serum Lipid Levels
2.6. Determination of Blood Glucose and Insulin Levels
2.7. Determination of Serum Lipopolysaccharide (LPS) and Glycosylated Hemoglobin (HbA1c) Levels
2.8. Determination of Liver Glycogen Levels
2.9. Western Blot
2.10. Plate Colony Counting
2.11. Statistical Analysis
3. Results
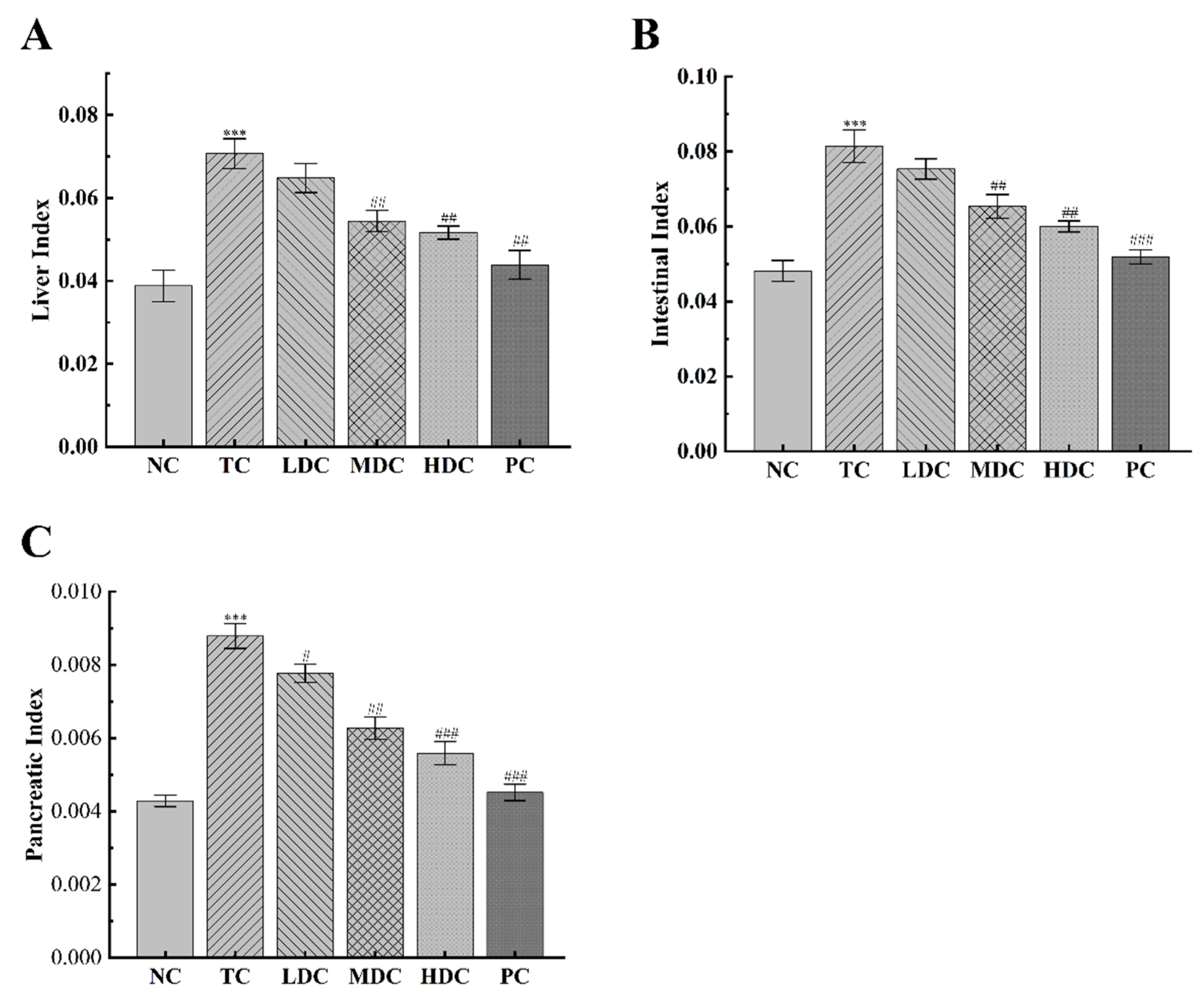
3.1. Effects of PSS on General Physiological Indices
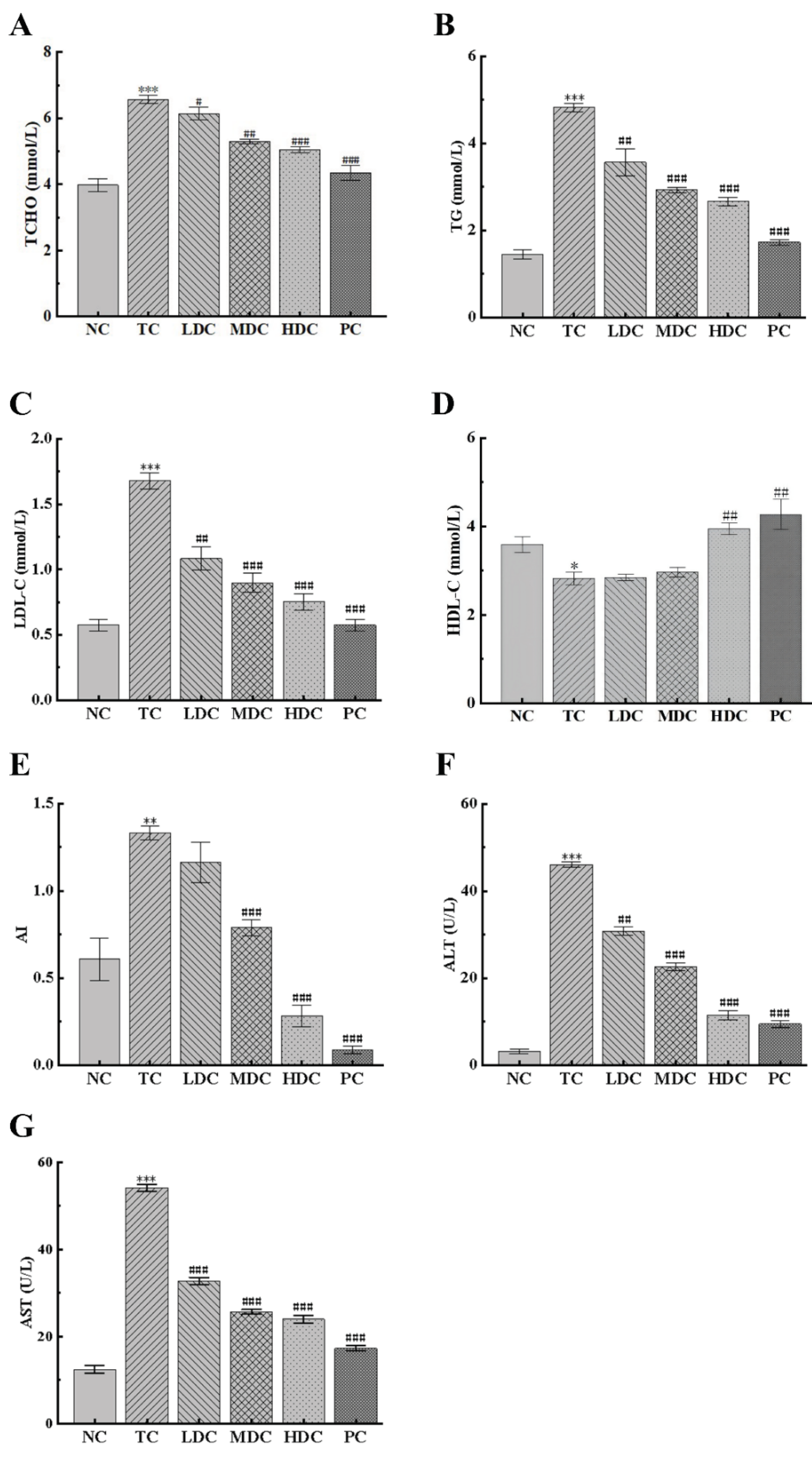
3.2. Effects of PSS on Serum Lipids and Biochemical Indices of Hepatocytes
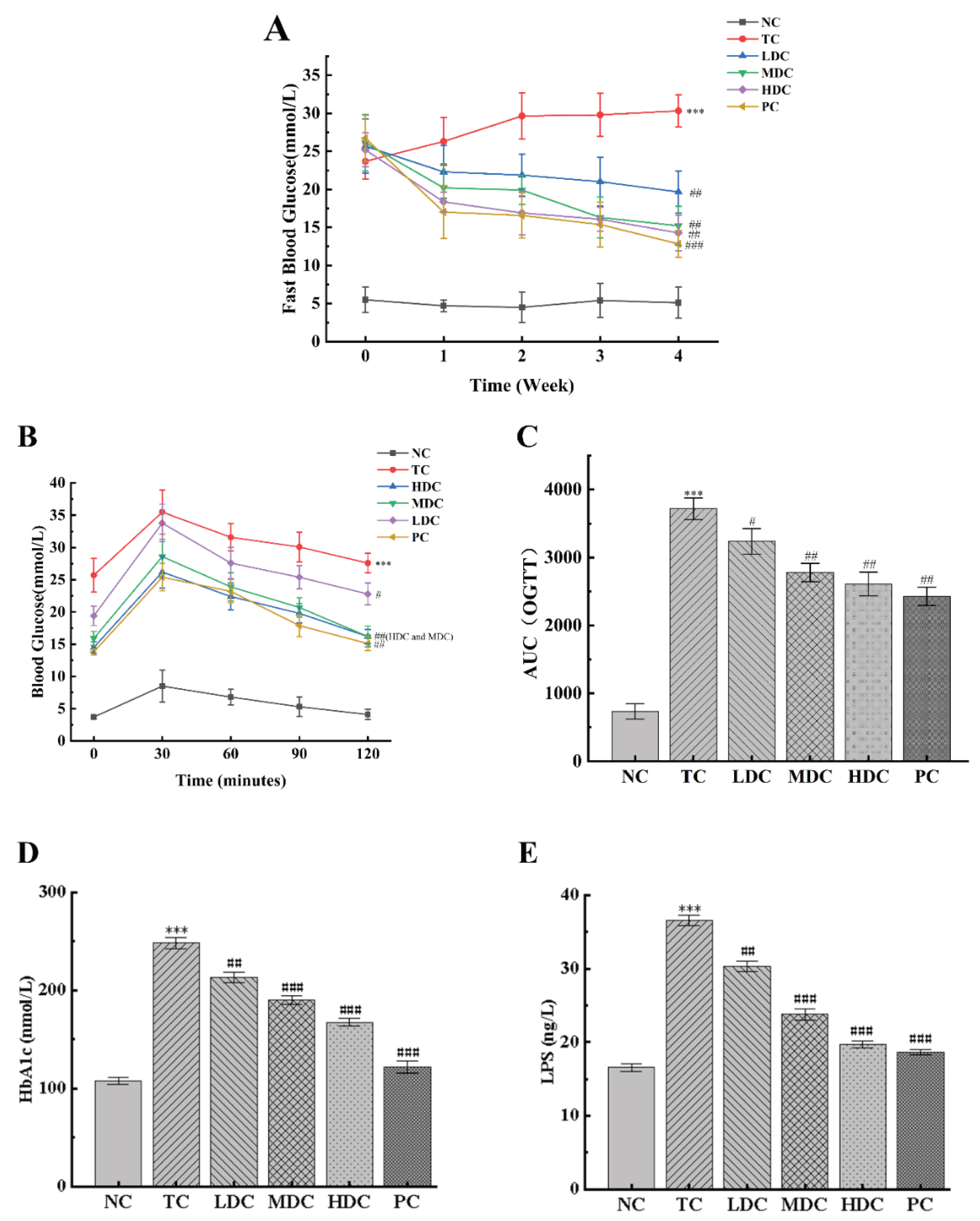
3.3. Effect of PSS on Glucose Tolerance in Mice
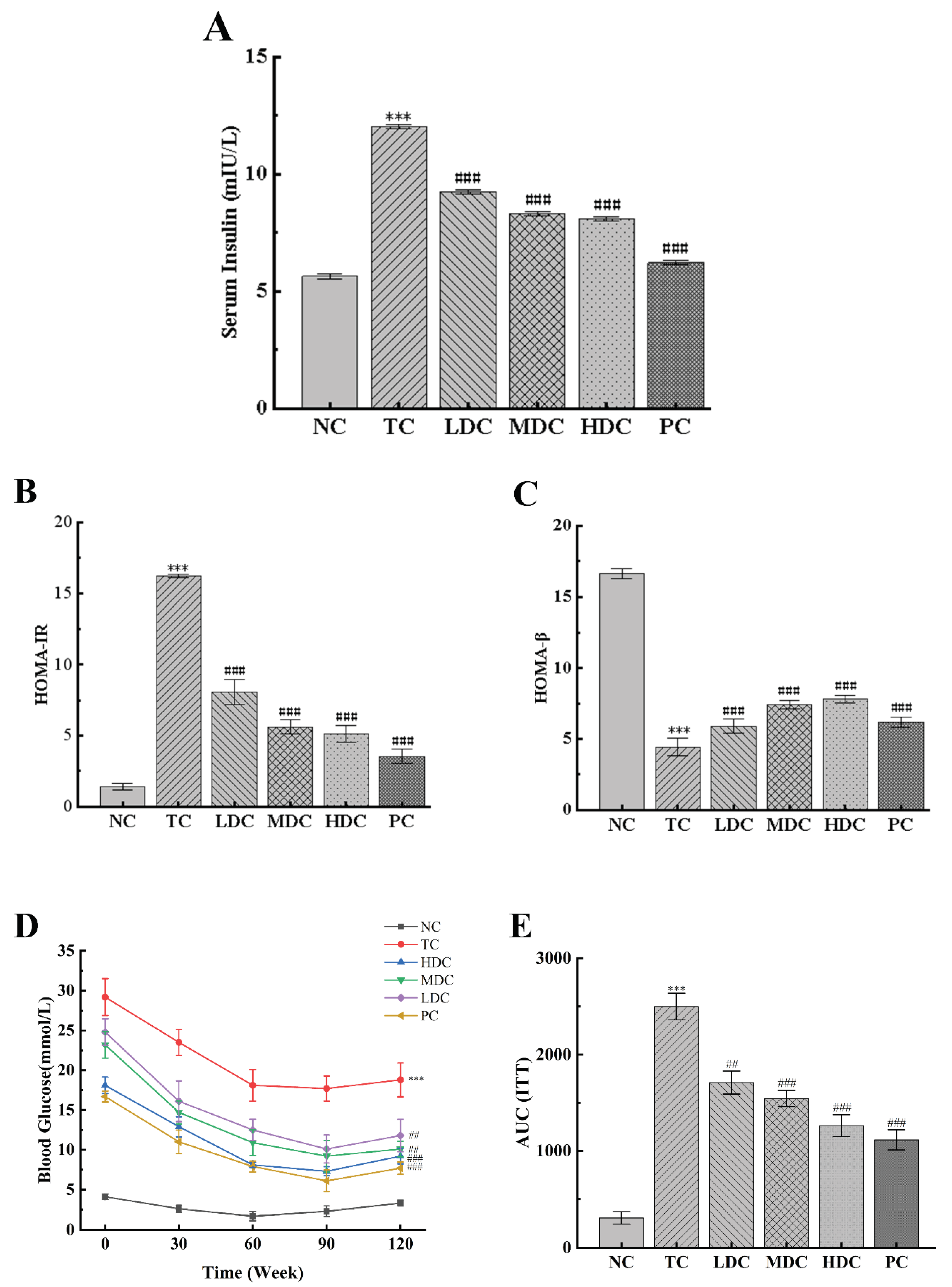
3.4. PSS Increases Insulin Sensitivity in Diabetic Mice
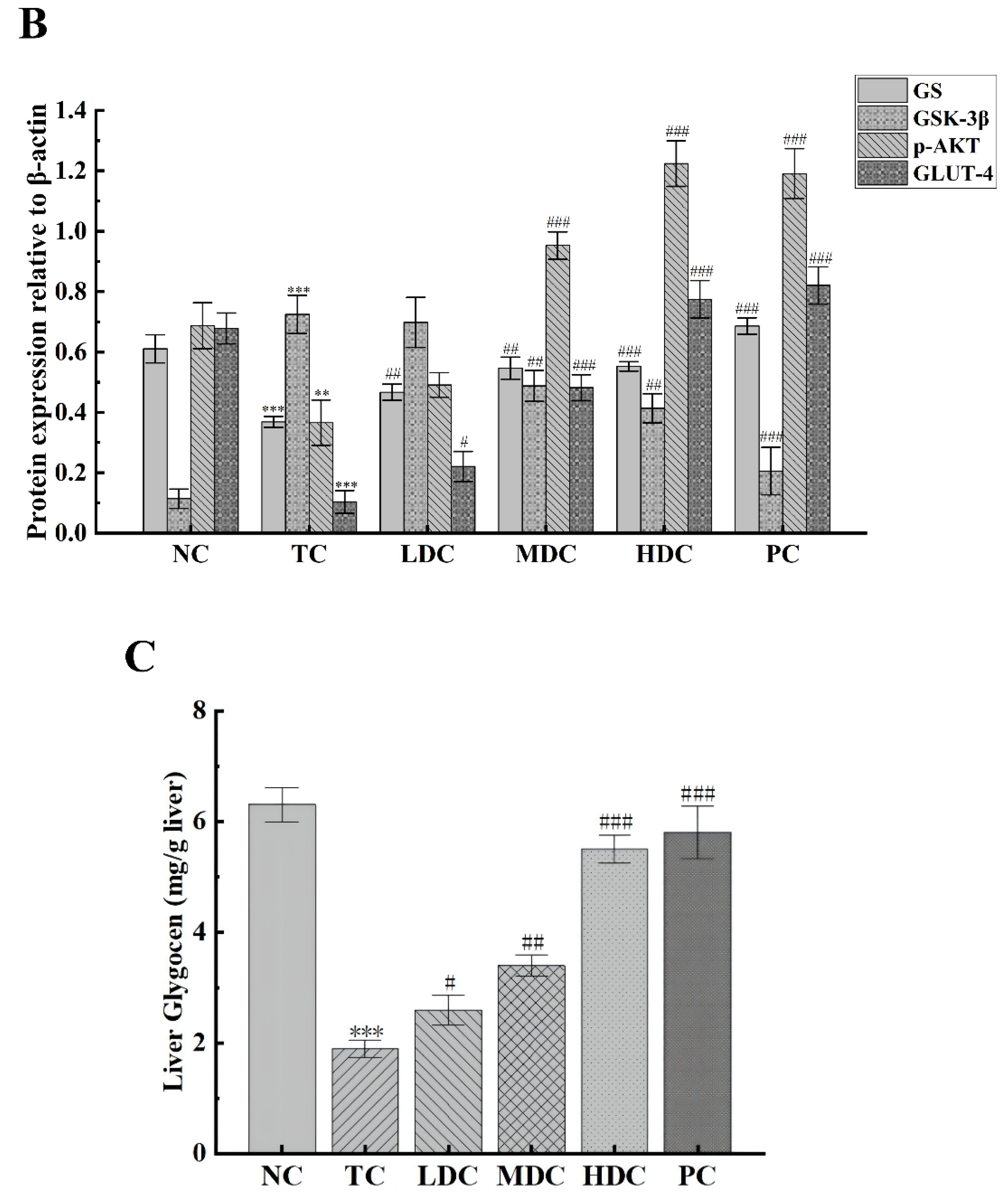
3.5. Effects of PSS on Insulin Pathway-Related Proteins and Hepatic Glycogen Synthesis-Related Proteins
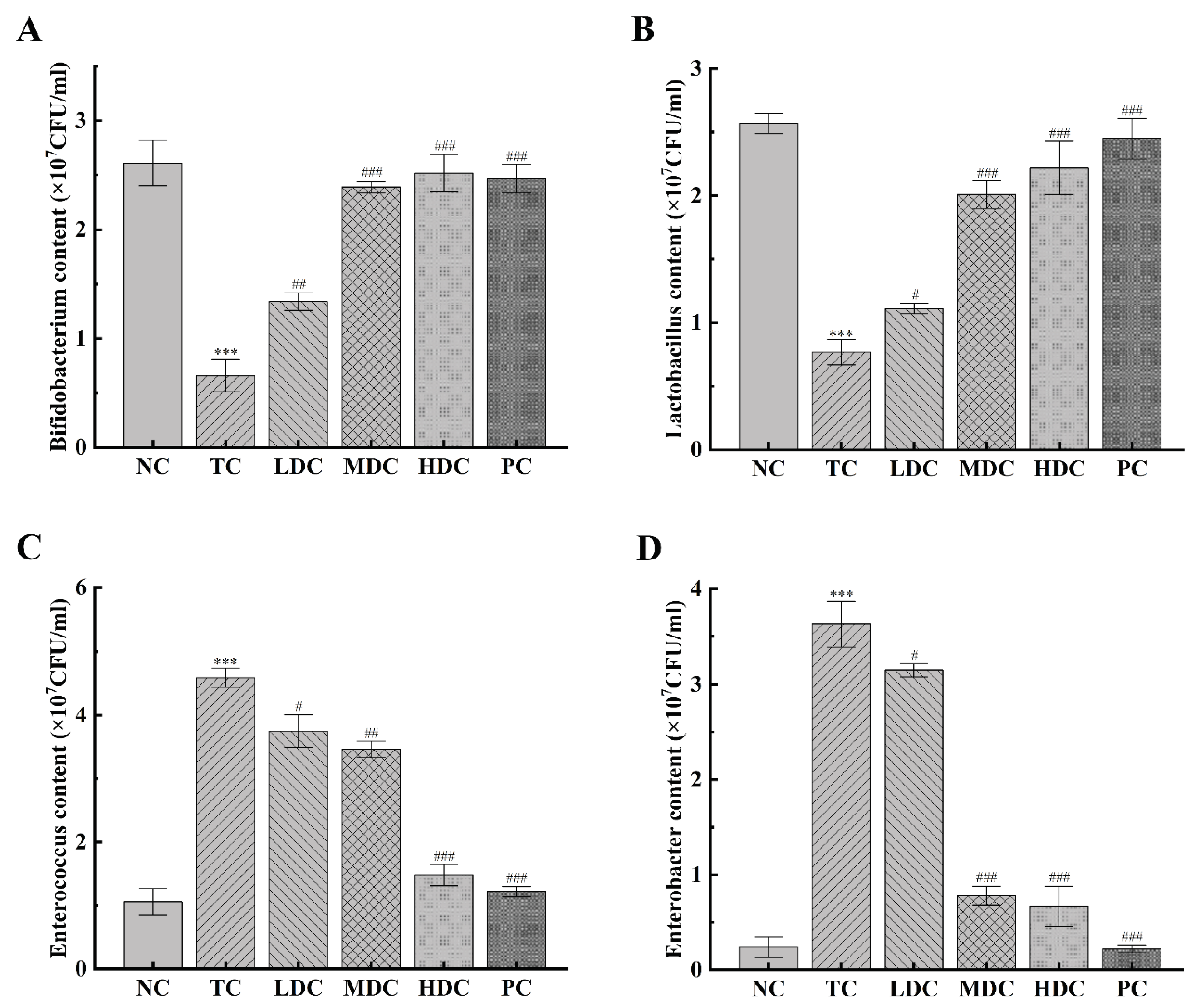
3.6. Effect of PSS on Important Intestinal Bacteria
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.; Pan, M.; Pan, S.; Li, W.; Zhong, Y.; Hu, J.; Nie, S. Effects of insoluble and soluble fibers isolated from barley on blood glucose, serum lipids, liver function and caecal short-chain fatty acids in type 2 diabetic and normal rats. Food Chem. Toxicol. 2020, 135, 110937. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.E.B.F.; Moreira, N.C.S.; Sakamoto-Hojo, E.T. Mechanisms underlying the pathophysiology of type 2 diabetes: From risk factors to oxidative stress, metabolic dysfunction, and hyperglycemia. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2022, 874–875, 503437. [Google Scholar] [CrossRef] [PubMed]
- Yuming, W.; Huiqin, L.; Hui, G.; Xiaohua, X.; Tingting, C.; Huiying, W.; Yunting, Z.; Rong, H.; Xiaofei, S.; Jiahuan, M. Effect of chiglitazar and sitagliptin on glucose variations, insulin resistance and inflammatory-related biomarkers in untreated patients with type 2 diabetes. Diabetes Res. Clin. Pract. 2021, 183, 109171. [Google Scholar]
- Zhou, B.; Lu, Y.; Hajifathalian, K.; Bentham, J.; Cesare, M.D.; Danaei, G.; Bixby, H.; Cowan, M.; Ali, M.; Taddei, C.; et al. Worldwide trends in diabetes since 1980: A pooled analysis of 751 population-based studies with 4·4 million participants. Lancet 2016, 387, 1513–1530. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.Y.Y.; Feig, D.S.; Ho, J.; Siemens, S.; Josephine, H.; Rick, S. Blood Glucose Monitoring in Adults and Children with Diabetes: Update 2021. Can. J. Diabetes 2021, 45, 580–587. [Google Scholar] [CrossRef]
- Saeedi, P.; Petersohn, P.I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2021, 183, 109119. [Google Scholar] [CrossRef]
- Gomes, M.B.; Rathmann, W.; Charbonnel, B.; Khunti, K.; Kosiborod, M.; Nicolucci, A.; Pocock, S.J.; Shestakova, M.V.; Shimomura, I.; Tang, F. Treatment of type 2 diabetes mellitus worldwide: Baseline patient characteristics in the global DISCOVER study. Diabetes Res. Clin. Pract. 2019, 151, 20–32. [Google Scholar] [CrossRef]
- Polfus, L.M.; Darst, B.F.; Highland, H.; Sheng, X.; Ng, M.; Below, J.E.; Petty, L.; Bien, S.; Sim, X.; Wang, W. Genetic Discovery and Risk Characterization in Type 2 Diabetes across Diverse Populations. HGG Adv. 2021, 2, 100029. [Google Scholar] [CrossRef]
- Alberto, R.C.; Luigi, G. Kidney disease in diabetes: From mechanisms to clinical presentation and treatment strategies. Metab. Clin. Exp. 2021, 124, 154890. [Google Scholar]
- Stefano, C.; Rosa, C.; Simone, M.; Andrea, M.; Gianluca, P. Twenty-year trends in heart failure among U.S. adults, 1999-2018: The growing impact of obesity and diabetes. Int. J. Cardiol. 2022, 362, 104–109. [Google Scholar]
- Karis, L.; María, L.; Sarah, S.; Cristina, H.; Olga, S.; Angel, D.M.; Esmeralda, B.; Maria, V.; Jose, C.M.; Tine, V.B.; et al. Common pathways in dementia and diabetic retinopathy: Understanding the mechanisms of diabetes-related cognitive decline. Trends Endocrinol. Metab. TEM 2021, 33, 50–71. [Google Scholar]
- Chao, J.; Nau, D.P.; Aikens, J.E. Patient-reported perceptions of side effects of antihyperglycemic medication and adherence to medication regimens in persons with diabetes mellitus. Clin. Ther. 2007, 29, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Ha, A.; Ba, B.; Ayn, C.; Es, E.; Sq, B.; Ks, D.; Msd, A.; Lh, D.; Ya, D.; Ms, A. The safety and efficacy of the use of oral anticoagulant medications in patients with diabetes mellitus: A systematic review—ScienceDirect. Saudi Pharm. J. 2021, 29, 1374–1382. [Google Scholar]
- Rampersad, C.; Kraut, E.; Whitlock, R.H.; Komenda, P.; Woo, V.; Rigatto, C.; Tangri, N. Acute Kidney Injury Events in Patients With Type 2 Diabetes Using SGLT2 Inhibitors Versus Other Glucose-Lowering Drugs: A Retrospective Cohort Study. Am. J. Kidney Dis. 2020, 76, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Muche, B.Y.; Teshome, M.; Solomon, A.; Demelash, K.Z. A retrospective study of drug related problems and contributing factors among type 2 diabetes mellitus patients on follow up at public health institutions of kemisse town, north east Ethiopia. Metab. Open 2021, 11, 100098. [Google Scholar]
- Wang, H.Y.; Kan, W.C.; Cheng, T.J.; Yu, S.H.; Chang, L.H.; Chuu, J.J. Differential anti-diabetic effects and mechanism of action of charantin-rich extract of Taiwanese Momordica charantia between type 1 and type 2 diabetic mice. Food Chem. Toxicol. 2014, 69, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Choi, E.-H.; Chun, Y.-S.; Kim, J.; Ku, S.-K.; Jeon, S.; Park, T.-S.; Shim, S.-M. Modulating lipid and glucose metabolism by glycosylated kaempferol rich roasted leaves of Lycium chinense via upregulating adiponectin and AMPK activation in obese mice-induced type 2 diabetes. J. Funct. Foods 2020, 72, 104072. [Google Scholar] [CrossRef]
- Kuiniu, Z.; Zhaoqing, M.; Yushan, T.; Rui, G.; Zhongkun, X.; Hui, F.; Wenjun, L.; Wenzhe, H.; Gang, D.; Wei, X. Hypoglycemic and hypolipidemic effects of total glycosides of Cistanche tubulosa in diet/streptozotocin-induced diabetic rats. J. Ethnopharmacol. 2021, 276, 113991. [Google Scholar]
- Guo, W.-L.; Deng, J.-C.; Pan, Y.-Y.; Xu, J.-X.; Hong, J.-L.; Shi, F.-F.; Liu, G.-L.; Qian, M.; Bai, W.-D.; Zhang, W.; et al. Hypoglycemic and hypolipidemic activities of Grifola frondosa polysaccharides and their relationships with the modulation of intestinal microflora in diabetic mice induced by high-fat diet and streptozotocin. Int. J. Biol. Macromol. 2020, 153, 1231–1240. [Google Scholar] [CrossRef] [PubMed]
- Ru, Y.; Liu, K.; Kong, X.; Li, X.; Shi, X.; Chen, H. Synthesis of selenylated polysaccharides from Momordica charantia L. and its hypoglycemic activity in streptozotocin-induced diabetic mice. Int. J. Biol. Macromol. 2020, 152, 295–304. [Google Scholar] [CrossRef]
- Yanyan, Z.; Haixia, Y.; Jianjun, D.; Daidi, F. Ginsenoside Rg5 Improves Insulin Resistance and Mitochondrial Biogenesis of Liver via Regulation of the Sirt1/PGC-1α Signaling Pathway in db/db Mice. J. Agric. Food Chem. 2021, 69, 8428–8439. [Google Scholar]
- Shruti, S.; Robin, J.; Dinesh, K. Metabolomics insights and bioprospection of Polygonatum verticillatum: An important dietary medicinal herb of alpine Himalaya. Food Res. Int. 2021, 148, 110619. [Google Scholar]
- Huan, Z.; Qiao-Lian, W.; Shu-Bing, H.; Guang, C. Chemical constituents from the rhizomes of Polygonatum sibiricum Red. and anti-inflammatory activity in RAW264.7 macrophage cells. Nat. Prod. Res. 2019, 33, 2359–2362. [Google Scholar]
- Long, T.; Liu, Z.; Shang, J.; Zhou, X.; Yu, S.; Tian, H.; Bao, Y. Corrigendum to “ Polygonatum sibiricum polysaccharides play anti-cancer effect through TLR4-MAPK/NF-κB signaling pathways” [Int. J. Biol. Macromol. 111 (2018):813–821]. Int. J. Biol. Macromol. 2018, 127, 703. [Google Scholar] [CrossRef] [PubMed]
- Fengming, S.; Zhujin, S.; Pan, X.; Lei, L.; Bin, W.; Daiyin, P.; Guoqi, Z. Polygonatum sibiricum polysaccharide prevents depression-like behaviors by reducing oxidative stress, inflammation, and cellular and synaptic damage. J. Ethnopharmacol. 2021, 275, 114164. [Google Scholar]
- Han, C.; Sun, T.; Liu, Y.; Fan, G.; Zhang, W.; Liu, C. Protective effect of Polygonatum sibiricum polysaccharides on gentamicin-induced acute kidney injury in rats via inhibiting p38 MAPK/ATF2 pathway. Int. J. Biol. Macromol. 2020, 151, 595–601. [Google Scholar] [CrossRef]
- Yelithao, K.; Surayot, U.; Park, W.; Lee, S.; Lee, D.-H.; You, S. Effect of sulfation and partial hydrolysis of polysaccharides from Polygonatum sibiricum on immune-enhancement. Int. J. Biol. Macromol. 2018, 122, 10–18. [Google Scholar] [CrossRef]
- Bingjin, M.; Youzhong, W.; Yiwen, B.; Mao, W.; Mengyang, H. Saponins from the rhizomes of Polygonatum nodosum Hua and their chemotaxonomic significance. Biochem. Syst. Ecol. 2021, 98, 104308. [Google Scholar]
- Cheng, T.; Ya-Ming, Y.; Qing-Ling, Q.; Xiao-Dan, W.; Jia, W.; Sheng-An, T. Steroidal saponins from the rhizome of Polygonatum sibiricum. J. Asian Nat. Prod. Res. 2019, 21, 360–364. [Google Scholar]
- Ma, Y.-L.; Zhang, Y.-S.; Zhang, F.; Zhang, Y.-Y.; Thakur, K.; Zhang, J.-G.; Wei, Z.-J. Methyl protodioscin from Polygonatum sibiricum inhibits cervical cancer through cell cycle arrest and apoptosis induction. Food Chem. Toxicol. 2019, 132, 110655. [Google Scholar] [CrossRef] [PubMed]
- Elekofehinti, O.O.; Kamdem, J.P.; Kade, I.J.; Rocha, J.; Adanlawo, I.G. Hypoglycemic, antiperoxidative and antihyperlipidemic effects of saponins from Solanum anguivi Lam. fruits in alloxan-induced diabetic rats. South Afr. J. Bot. 2013, 88, 56–61. [Google Scholar] [CrossRef]
- Luo, J.; Chai, Y.; Zhao, M.; Guo, Q.; Bao, Y. Hypoglycemic effects and modulation of gut microbiota of diabetic mice by saponin from Polygonatum sibiricum. Food Funct. 2020, 11, 4327–4338. [Google Scholar] [CrossRef] [PubMed]
- Muskan, S.; Arunima, W.; Kumar, S.R. Recent developments in probiotics: An emphasis on Bifidobacterium. Food Biosci. 2021, 41, 100993. [Google Scholar]
- Jéssica, G.V.d.S.; Angélica, T.V.; Sousa, T.J.; Marcus, V.C.V.; Doglas, P.; Bruna, S.; Alessandra, L.d.S.; Luís, C.L.d.J.; Pedro, K.R.M.L.d.C.; Letícia, d.C.O.; et al. Comparative genomics and in silico gene evaluation involved in the probiotic potential of Bifidobacterium longum 51A. Gene 2021, 795, 145781. [Google Scholar]
- Songkang, Q.; Zonghao, H.; Yingli, W.; Lulu, P.; Yaoqin, S. Probiotic potential of Lactobacillus isolated from horses and its therapeutic effect on DSS-induced colitis in mice. Microb. Pathog. 2021, 165, 105216. [Google Scholar]
- Srujana, M.; Sourin, B. Pleural empyema due to Enterococcus species: An uncommon etiology. Respir. Med. Case Rep. 2021, 32, 101375. [Google Scholar]
- Kazushi, Y.; Koichi, Y.; Waki, I.; Gaku, K.; Wataru, S.; Hiroki, N.; Kazuhiro, O.; Kiyotaka, N.; Makoto, N.; Yukihiro, K.; et al. Risk factor analysis for piperacillin-tazobactam-resistant Enterobacter spp. bacteremia at a tertiary hospital. Diagn. Microbiol. Infect. Dis. 2021, 100, 115342. [Google Scholar]
- Lvarez-Marín, R.; Lepe, J.A.; Gasch-Blasi, O.; Rodríguez-Martínez, J.; Jiménez-Mejías, M. Clinical characteristics and outcome of bacteremia caused by Enterobacter cloacae and Klebsiella aerogenes: More similarities than differences. J. Glob. Antimicrob. Resist. 2021, 25, 351–358. [Google Scholar] [CrossRef]
- Zhou, B.; Li, Q.; Wang, J.; Chen, P.; Jiang, S. Ellagic acid attenuates streptozocin induced diabetic nephropathy via the regulation of oxidative stress and inflammatory signaling. Food Chem. Toxicol. 2018, 123, 16–27. [Google Scholar] [CrossRef]
- Yooni, B.; Hunter, W.; Rodica, P.-B.; Lynn, A.; Sarma, A.V. Urologic complications in diabetes. J. Diabetes Its Complicat. 2022, 36, 177–185. [Google Scholar]
- Entezari, M.; Hashemi, D.; Taheriazam, A.; Zabolian, A.; Samarghandian, S. AMPK signaling in diabetes mellitus, insulin resistance and diabetic complications: A pre-clinical and clinical investigation. Biomed. Pharmacother. 2022, 146, 112563. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Yang, H.; Zhu, C.; Deng, J.; Fan, D. Ginsenoside Rg5 relieves type 2 diabetes by improving hepatic insulin resistance in db/db mice. J. Funct. Foods 2020, 71, 104014. [Google Scholar] [CrossRef]
- Parkman, J.K.; Kristiana, S.L.; Kalhara, R.M.; Logan, F.; Naima, M.M.; Han, K.J. Effects of high fat diets and supplemental tart cherry and fish oil on obesity and type 2 diabetes in male and female C57BL/6J and TALLYHO/Jng mice. J. Nutr. Biochem. 2021, 94, 108644. [Google Scholar] [CrossRef] [PubMed]
- Jie, D.; Jing, Z.; Jin, L.C.; Xza, B.; Da, W.; Ysa, B.; Kai, Z.D.; Yla, B.; Rha, B.; Xwa, B. Hypoglycemic effects and mechanism of different molecular weights of konjac glucomannans in type 2 diabetic rats—ScienceDirect. Int. J. Biol. Macromol. 2020, 165, 2231–2243. [Google Scholar]
- Limin, Y.; Lin, Z.; Xiaohui, W.; Wang, W.; Jin, W. Inhibition of HMGB1 involved in the protective of salidroside on liver injury in diabetes mice. Int. Immunopharmacol. 2020, 89, 106987. [Google Scholar]
- Yuan, Y.; Zhou, J.; Zheng, Y.; Xu, Z.; Li, Y.; Zhou, S.; Zhang, C. Beneficial effects of polysaccharide-rich extracts from Apocynum venetum leaves on hypoglycemic and gut microbiota in type 2 diabetic mice. Biomed. Pharmacother. 2020, 127, 110182. [Google Scholar] [CrossRef]
- Elsayed, R.H.; Kamel, E.M.; Mahmoud, A.M.; El-Bassuony, A.A.; Bin-Jumah, M.; Lamsabhi, A.M.; Ahmed, S.A. Rumex dentatus L. phenolics ameliorate hyperglycemia by modulating hepatic key enzymes of carbohydrate metabolism, oxidative stress and PPARγ in diabetic rats. Food Chem. Toxicol. 2020, 138, 111202. [Google Scholar] [CrossRef]
- Li, S.; Chen, H.; Wang, J.; Wang, X.; Hu, B.; Lv, F. Involvement of the PI3K/Akt signal pathway in the hypoglycemic effects of tea polysaccharides on diabetic mice. Int. J. Biol. Macromol. 2015, 81, 967–974. [Google Scholar] [CrossRef]
- Li, X.; Zhang, X.; Zhang, R.; Ni, Z.; Elam, E.; Thakur, K.; Cespedes-Acuña, C.L.; Zhang, J.; Wei, Z. Gut modulation based anti-diabetic effects of carboxymethylated wheat bran dietary fiber in high-fat diet/streptozotocin-induced diabetic mice and their potential mechanisms. Food Chem. Toxicol. 2021, 152, 112235. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Z.; Xu, X.; Jiang, H.; Yu, G. Odd-numbered agaro-oligosaccharides alleviate type 2 diabetes mellitus and related colonic microbiota dysbiosis in mice. Carbohydr. Polym. 2020, 240, 116261. [Google Scholar] [CrossRef] [PubMed]
- Chai, Y.; Luo, J.; Bao, Y. Effects of Polygonatum sibiricum saponin on hyperglycemia, gut microbiota composition and metabolic profiles in type 2 diabetes mice. Biomed. Pharmacother. 2021, 143, 112155. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Julia, P.J.; Jose, M.-M.; van Douwe, S. A comprehensive review on the impact of β-glucan metabolism by Bacteroides and Bifidobacterium species as members of the gut microbiota. Int. J. Biol. Macromol. 2021, 181, 877–889. [Google Scholar] [CrossRef] [PubMed]
- Alcon-Giner, C.; Dalby, M.J.; Caim, S.; Ketskemety, J.; Shaw, A.; Sim, K.; Lawson, M.A.E.; Kiu, R.; Leclaire, C.; Chalklen, L.; et al. Microbiota Supplementation with Bifidobacterium and Lactobacillus Modifies the Preterm Infant Gut Microbiota and Metabolome: An Observational Study. Cell Rep. Med. 2020, 1, 100077. [Google Scholar] [CrossRef]
- Paula, M.A.; Eichelberger, G.C. Mapping the Worldwide Knowledge of Antimicrobial Substances Produced by Lactobacillus spp.: A Bibliometric Analysis. Biochem. Eng. J. 2022, 180, 108343. [Google Scholar]
- Haicong, Z.; Shibo, Z.; Xueqi, Z.; Hong, Z.; Tieqiao, W.; Jiao, W. Depression-like symptoms due to Dcf1 deficiency are alleviated by intestinal transplantation of Lactobacillus murine and Lactobacillus reuteri. Biochem. Biophys. Res. Commun. 2022, 593, 137–143. [Google Scholar]
- Anshu, Y.; Yaru, L.; Jierui, Z.; Jie, Z.; Zhihua, W.; Xin, L.; Ping, T.; Hongbing, C.; Shuixing, W.; Zhiyong, L. Screening of anti-allergy Lactobacillus and its effect on allergic reactions in BALB/c mice sensitized by soybean protein. J. Funct. Foods 2021, 87, 104858. [Google Scholar]
- Ll, A.; So, B.; Lc, A.; Ob, A.; Rf, A.; Tc, A.; Lv, A.; Bdv, A.; Pdl, A. Risk factors and outcomes associated with vancomycin-resistant Enterococcus faecium and ampicillin-resistant Enterococcus faecalis bacteraemia: A 10-year study in a tertiary-care centre in Mexico City. J. Glob. Antimicrob. Resist. 2021, 24, 198–204. [Google Scholar]
- Karl, O.; Magnus, R. Enterococcus faecalis in blood cultures—A prospective study on the role of persistent bacteremia. Diagn. Microbiol. Infect. Dis. 2021, 101, 115433. [Google Scholar]
- Zhiyong, Z.; Yu, F.; Alan, M. Carbapenem and Colistin Resistance in Enterobacter: Determinants and Clones. Trends Microbiol. 2021, 29, 473–476. [Google Scholar] [CrossRef]
- Negin, B.; Giske, C.G.; Shoeib, N.; Amir, M.; Seyyedi, A.S.; Hamid, S.; Farzad, B. Comparative resistome and virulome analysis of clinical NDM-1 producing carbapenem-resistant Enterobacter cloacae complex. J. Glob. Antimicrob. Resist. 2022, 28, 254–263. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | 1 Week | 2 Week | 3 Week | 4 Week |
|---|---|---|---|---|
| NC | 5.53 ± 0.27 | 5.39 ± 0.87 | 6.01 ± 0.68 | 5.18 ± 0.36 |
| TC | 11.71 ± 0.67 *** | 11.97 ± 1.58 *** | 12.59 ± 1.66 *** | 13.12 ± 0.93 *** |
| HDC | 10.93 ± 1.71 | 11.11 ± 0.79 | 10.62 ± 1.58 # | 9.83 ± 1.27 # |
| MDC | 11.64 ± 0.43 | 11.72 ± 0.23 | 10.98 ± 1.12 | 9.92 ± 1.01 # |
| LDC | 12.03 ± 0.94 | 12.27 ± 0.86 | 11.85 ± 0.40 | 11.62 ± 0.71 |
| PC | 11.50 ± 0.51 | 10.93 ± 1.31 | 11.12 ± 0.27 | 10.14 ± 0.43 # |
| Group | 1 Week | 2 Week | 3 Week | 4 Week |
|---|---|---|---|---|
| NC | 5.82 ± 0.71 | 5.29 ± 0.80 | 6.39 ± 1.11 | 5.65 ± 1.03 |
| TC | 38.72 ± 2.36 *** | 40.88 ± 1.99 *** | 42.48 ± 3.32 *** | 46.19 ± 2.85 *** |
| HDC | 38.07 ± 1.61 | 33.93 ± 2.55 # | 30.93 ± 1.78 ## | 29.36 ± 1.13 ## |
| MDC | 39.60 ± 1.42 | 37.01 ± 1.98 | 35.82 ± 1.23 # | 32.79 ± 0.78 ## |
| LDC | 40.06 ± 2.03 | 39.81 ± 1.59 | 38.87 ± 0.41 | 36.71 ± 2.24 # |
| PC | 36.78 ± 0.51 | 31.07 ± 1.41 # | 28.11 ± 1.04 ## | 23.68 ± 1.35 ## |
| Group | 0 Week | 1 Week | 2 Week | 3 Week | 4 Week |
|---|---|---|---|---|---|
| NC | 32.98 ± 3.54 | 33.82 ± 2.99 | 35.01 ± 2.46 | 35.62 ± 3.13 | 36.68 ± 3.29 |
| TC | 38.90 ± 4.67 | 40.08 ± 5.23 | 44.45 ± 4.62 * | 47.18 ± 2.91 * | 49.08 ± 3.04 * |
| HDC | 36.47 ± 3.07 | 37.41 ± 2.41 | 38.26 ± 1.70 | 39.13 ± 2.76 | 39.49 ± 3.35 # |
| MDC | 37.71 ± 2.39 | 38.22 ± 1.40 | 39.60 ± 2.26 | 40.13 ± 3.22 | 40.56 ± 2.73 # |
| LDC | 37.77 ± 2.76 | 38.93 ± 4.21 | 40.01 ± 2.73 | 40.51 ± 3.44 | 40.88 ± 3.85 # |
| PC | 36.84 ± 3.17 | 37.53 ± 3.24 | 38.28 ± 3.76 | 38.92 ± 4.77 # | 39.51 ± 3.69 # |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.; Luo, J.; Jia, M.; Chai, Y.; Bao, Y. Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins. Nutrients 2022, 14, 5222. https://doi.org/10.3390/nu14245222
Chen Z, Luo J, Jia M, Chai Y, Bao Y. Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins. Nutrients. 2022; 14(24):5222. https://doi.org/10.3390/nu14245222
Chicago/Turabian StyleChen, Zefu, Jiayuan Luo, Mingjie Jia, Yangyang Chai, and Yihong Bao. 2022. "Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins" Nutrients 14, no. 24: 5222. https://doi.org/10.3390/nu14245222
APA StyleChen, Z., Luo, J., Jia, M., Chai, Y., & Bao, Y. (2022). Polygonatum sibiricum saponin Exerts Beneficial Hypoglycemic Effects in Type 2 Diabetes Mice by Improving Hepatic Insulin Resistance and Glycogen Synthesis-Related Proteins. Nutrients, 14(24), 5222. https://doi.org/10.3390/nu14245222