1. Introduction
Vaginal microbiota, although characterized by few microbial species, is a dynamic ecosystem subjected to changes during a woman’s lifetime, from birth to pre- and postmenopause [
1,
2,
3]. In addition, intrinsic (e.g., race, immune system imbalance, genetic susceptibility, etc.) and extrinsic factors (such as a diet rich in vitamins and folic acid, use of oral contraceptives, sexual behaviors, hygiene habits, antibiotics, or immunosuppressor therapies) can affect the balance of the vaginal bacterial biota. It is well known that the reduction in lactobacilli and the increase in facultative or obligate anaerobic microorganisms, along with different
Candida species, are associated with vaginal microbiota dysbiosis [
4,
5]. Although most lactobacilli have a GRAS (generally recognized as safe) status and some of them are included in the Qualified Presumption of Safety (QPS) list [
6,
7], both safety and probiotic features are strain-specific and cannot be generalized to the species. Several studies demonstrated the ability of probiotic lactobacilli to protect vaginal health by different mechanisms of action, including antagonistic activity against pathogens through competitive exclusion, competition for nutrients, and production of metabolites with antimicrobial properties. In particular, the antagonistic activity against pathogens, one of the key features of probiotics, could be linked to the ability of lactobacilli to acidify the vaginal environment, producing mainly lactic acid thus preventing the colonization and proliferation of pathogens [
4,
5]. In addition, the ability to adhere to epithelial cells and to form biofilm on vaginal mucosa as well as modulation of the immune system is included among the health-promoting properties of lactobacilli [
7,
8,
9,
10,
11,
12,
13,
14,
15]. In fact, the ability to adhere to the vaginal epithelium and compete for adhesion sites is involved in the inhibition of colonization by pathogens. In this context, the formation of a biofilm is a strategy that allows organisms to persist under stressful conditions allowing resistance to harsh environments. In addition, the development of a biofilm, by promoting mucosal colonization, interferes with the growth and adhesion of pathogens [
16].
The beneficial effect of lactobacilli to balance vaginal microbiota has recently gained a lot of interest. This assumption is corroborated by several clinical studies reporting their usefulness for both the prevention and treatment of urogenital tract infections in women under different physiological conditions [
2]. According to that, the use of lactobacilli is considered a valuable approach for the prevention and treatment of urogenital tract infections [
2]. Probiotic strains, ascribed to the
Lacticaseibacillus rhamnosus species, were extensively studied to evaluate their potential in the treatment of vaginal dysbiosis. Based on clinical applications, the
L. rhamnosus GR-1 strain was firstly studied for the ability to balance the vaginal microbiota reducing the recurrence of urinary tract infections (UTIs) [
9]. As reported by De Alberti and coworkers [
17],
L. rhamnosus HN001 and
L. acidophilus La-14, orally administrated, significantly increased the vaginal
L. rhamnosus and
L. acidophilus abundance. Similarly, when administrated in association with
L. acidophilus GLA-14, the
L. rhamnosus HN001 strain significantly inhibited the cell density of
Gardnerella vaginalis,
Atopobium vaginae,
Staphylococcus aureus, and
Escherichia coli [
18]. Recently, Pino and coworkers [
19], demonstrated, by conducting an in vivo pilot study, the ability of the
L. rhamnosus TOM 22.8 strain to restore normal vaginal microbiota in patients with bacterial vaginosis (BV).
Although commercial probiotic products are now available, there is great interest from nutraceutical companies in selecting new probiotic candidates to benefit the vaginal environment. In the present study, the probiotic potential, as well as the safety properties of the Lacticaseibacillus rhamnosus CA15 (DSM 33960) strain isolated from a healthy woman of reproductive age, was in vitro investigated.
4. Discussion
It is well known that, during reproductive age, a balanced vaginal microbiota is dominated by lactobacilli, while their reduction, and the increase in pathogens, is often associated with a dysbiotic state [
42]. Although antimicrobial drugs have been traditionally used to treat dysbiosis, several side effects, such as short-term relapse and the promotion of antibiotic resistance, are frequently reported. For these reasons, alternative therapeutic strategies, able to modulate the microbiota and correct the imbalance, from an ecological approach, have been investigated. In this context, the probiotic supplementation, alone or complementary to antibiotic therapy, has gained increased attention to restore vaginal health avoiding the recurrence of bacterial vaginosis (BV) and vulvovaginal candidiasis (VVC) [
43,
44]. Although several probiotic strains have been characterized, only few are host-specific. It has been suggested that probiotic vaginal isolates, when administered, have increased colonization and adaptation abilities to the vaginal niche. The workflow of the present study was designed to evaluate the potential probiotic properties of the vaginal isolate
L. rhamnosus CA15 (DSM 33960) strain as a potential candidate for women’s health.
Noteworthy, probiotics must satisfy safety requirements. In this context, the inability to produce hemolysins and gelatinases, as well as to exert mucin degradation and DNase activities, suggest that the
L. rhamnosus CA15 (DSM 33960) strain can be safely used. In addition, regarding the horizontal transfer of genes encoding antibiotic resistance [
45,
46], no transferable antibiotic resistance should be carried by promising probiotic strains. In the present study, the
L. rhamnosus CA15 (DSM 33960) strain did not show resistance to the antibiotics suggested by the EFSA, whereas resistance to metronidazole, clotrimazole, and boric acid was detected. The high resistance to antibiotics routinely used to treat BV and VVC suggests the possible use of the CA15 (DSM 33960) strain as an adjuvant to antibiotic treatment, ensuring survival during antibiotic administration.
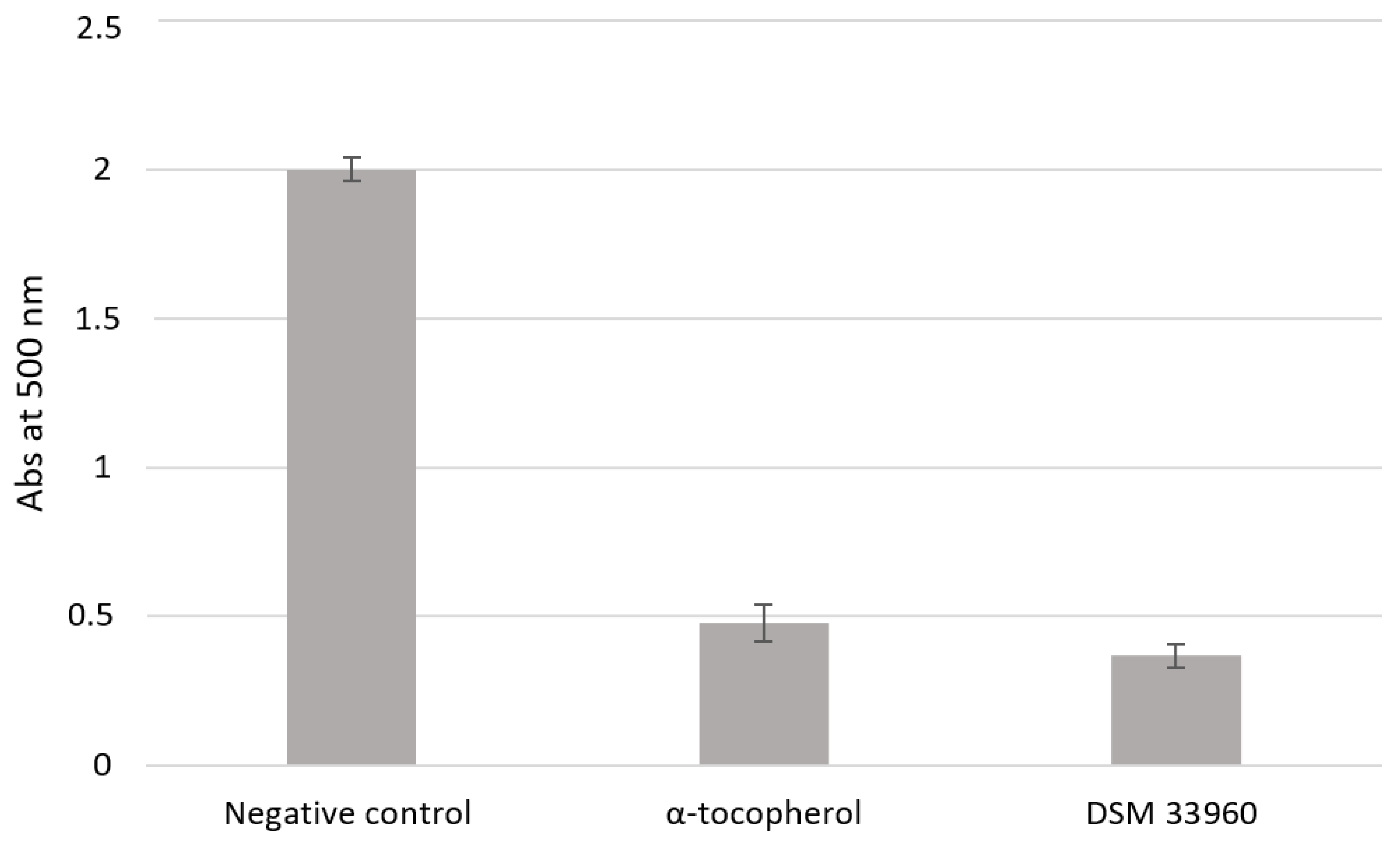
The ability of probiotics to counteract pathogen invasion and colonization is a prerequisite to modulate the vaginal microbiota in a homeostatic way. Antimicrobial substances, such as hydrogen peroxide (H
2O
2) and organic acids, produced by probiotic bacteria, play a key role in the maintenance of a healthy vaginal ecosystem. Although hydrogen peroxide is able to exert a microbicide effect, there is no consensus about its activity against pathogens due to both a low physiological concentration in the vagina and the neutralizing effects of semen and vaginal fluid [
47]. It is well established that lactic acid, by acidifying the vagina, acts as a virucide and microbicide along with an immunomodulatory agent [
48]. In the present study, we observed that the tested strain was able to produce both H
2O
2 and lactic acid. In particular, the amount of lactic acid produced could be related to the broad antagonistic effect displayed by the CA15 (DSM 33960) strain against the tested pathogens [
48]. The absence of antimicrobial activity after CFS neutralization suggests that the acid environment had a substantial action against the growth of the tested pathogens. However, other components, not yet identified, may occur in CFS. Further study based on the metabolomic approach will be carried out to characterize in depth the components of CFS and to evaluate their synergistic action to inhibit pathogens [
49,
50].
Along with the production of metabolites with antimicrobial properties, probiotic strains should be able to colonize and survive in harsh environments. The ability to adhere to the intestinal mucosa is considered a prerequisite for probiotics, allowing the colonization, albeit transient, of the human intestinal tract [
9,
14]. In vitro experiments showed the ability of the CA15 (DSM 33960) strain to adhere to intestinal mucus as well as to both Caco-2 and VK2/E6E7 cell lines with adhesion percentages higher than those previously obtained by
L. rhamnosus GG [
15]. In addition, the tested strain showed hydrophobicity and the ability to autoaggregate and coaggregate with pathogens. A higher hydrophobicity and autoaggregation ability was displayed by the CA15 strain compared to
L. rhamnosus GG [
40]. It is well known that the ability to adhere is related to the predisposition to self-aggregate, which, in turn, can exert a protective role by inhibiting the growth and adhesion of pathogens [
14]. Moreover, the exclusion/competition behavior of probiotics is also related to coaggregation with pathogens [
37]. According to that, the
L. rhamnosus CA15 (DSM 33960) strain showed a significant capability to coaggregate with all the tested pathogens as well as to displace and inhibit their adhesion.
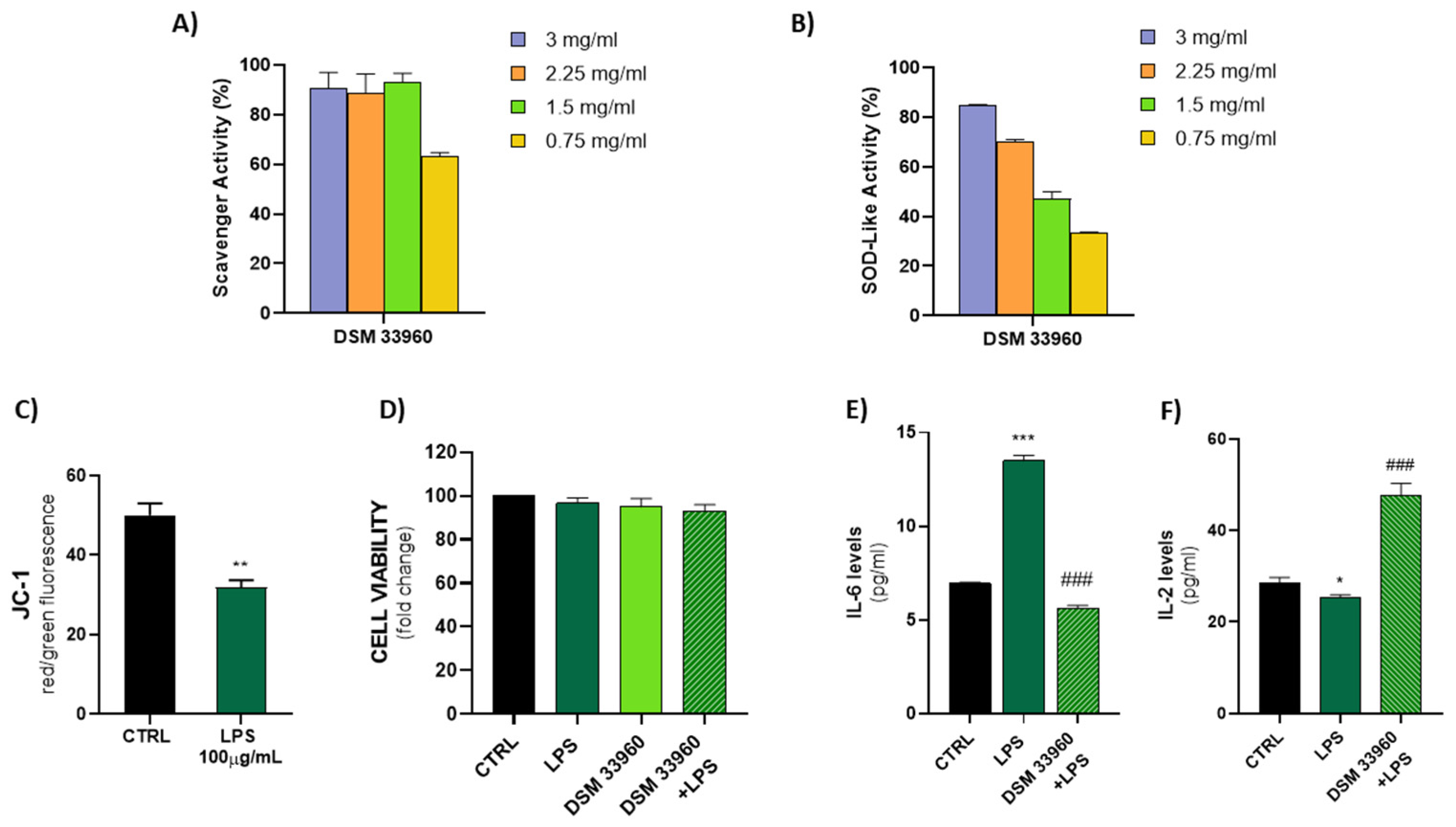
Recently, several health benefits (e.g., immunomodulation, antioxidant, antiviral, and antiyeast properties) have been associated with exopolysaccharide (EPS) produced by lactobacilli [
51]. According to that, the ability of the CA15 (DSM 33960) strain to produce EPS could be useful to maintain vaginal health. Other interesting properties of the
L. rhamnosus CA15 (DSM 33960) strain are the antioxidant and anti-inflammatory profiles. The ability to counteract the lipid peroxidation, and in turn oxidative stress, is considered central for the prevention of tissue damage and the development of human pathologies. In particular, the superoxide radical scavenging activity is an important enzymatic antioxidant defense mechanism catalyzed by the enzyme superoxide dismutase (SOD), which catalyzes the dismutation of the superoxide anion into oxygen and hydrogen peroxide to protect the body from oxygen toxicity. In this study, we measured the scavenging activity of the superoxide radical generated by the self-oxidation of pyrogallol. In addition, the anti-inflammatory profile suggests a potential role of the CA15 (DSM 33960) strain to decrease the levels of proinflammatory cytokines and maintain the inflammatory homeostasis by increasing the levels of the anti-inflammatory molecules. In particular, the tested strain was able to induce a decrease in the IL-6 and, in turn, determine an increase in the IL-2 in LPS-treated Caco-2 cells. It is well known that IL-6 is an important proinflammatory cytokine involved in the development of inflammatory bowel diseases (IBD) [
52,
53], whereas considerable evidence suggest the important role played by IL-2 in maintaining a healthy immune response in the gut. In fact, a high rate of colitis, with striking clinical and morphological similarities to ulcerative colitis, was reported in a IL-2-deficient mouse model [
54]. In addition, low doses of IL-2 are used for the treatment of moderate to severe ulcerative colitis [
55,
56].
Recently, the ascending colonization hypothesis was proposed suggesting that probiotics orally delivered can ascend to the vagina and colonize them after excretion from the rectum [
2]. To make this possible, probiotic strains must survive through the GI tract. Interestingly, the CA15 (DSM 33960) strain, tested in the present study, exhibited a high survival rate at a low pH, in the presence of lysozymes and bile salts as well as during the in vitro-simulated GI digestion. In particular, the survival percentages detected after exposure to SGJ and SIF were higher than those displayed by the
L. rhamnosus GG [
40].
Hence, the probiotic potential of the L. rhamnosus CA15 (DSM 33960) strain supports its use as a therapeutic strategy for women’s health. Further studies will be designed to set up an in vitro coculture model using Caco-2 cell lines and U937 cells to investigate the crosstalk between intestinal cells and the cells of the immune system treated with the L. rhamnosus CA15 (DSM 33960) strain. In addition, the in vitro therapeutic potential of the L. rhamnosus CA15 (DSM 33960) strain will be validated in a cohort of patients with vaginal dysbiosis.


 ,
, 










{kind=link}
{kind=link}
{kind=link}