
Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression


Abstract
1. Introduction
2. Materials and Methods
2.1. In vitro Digestion
2.2. Cell Culture
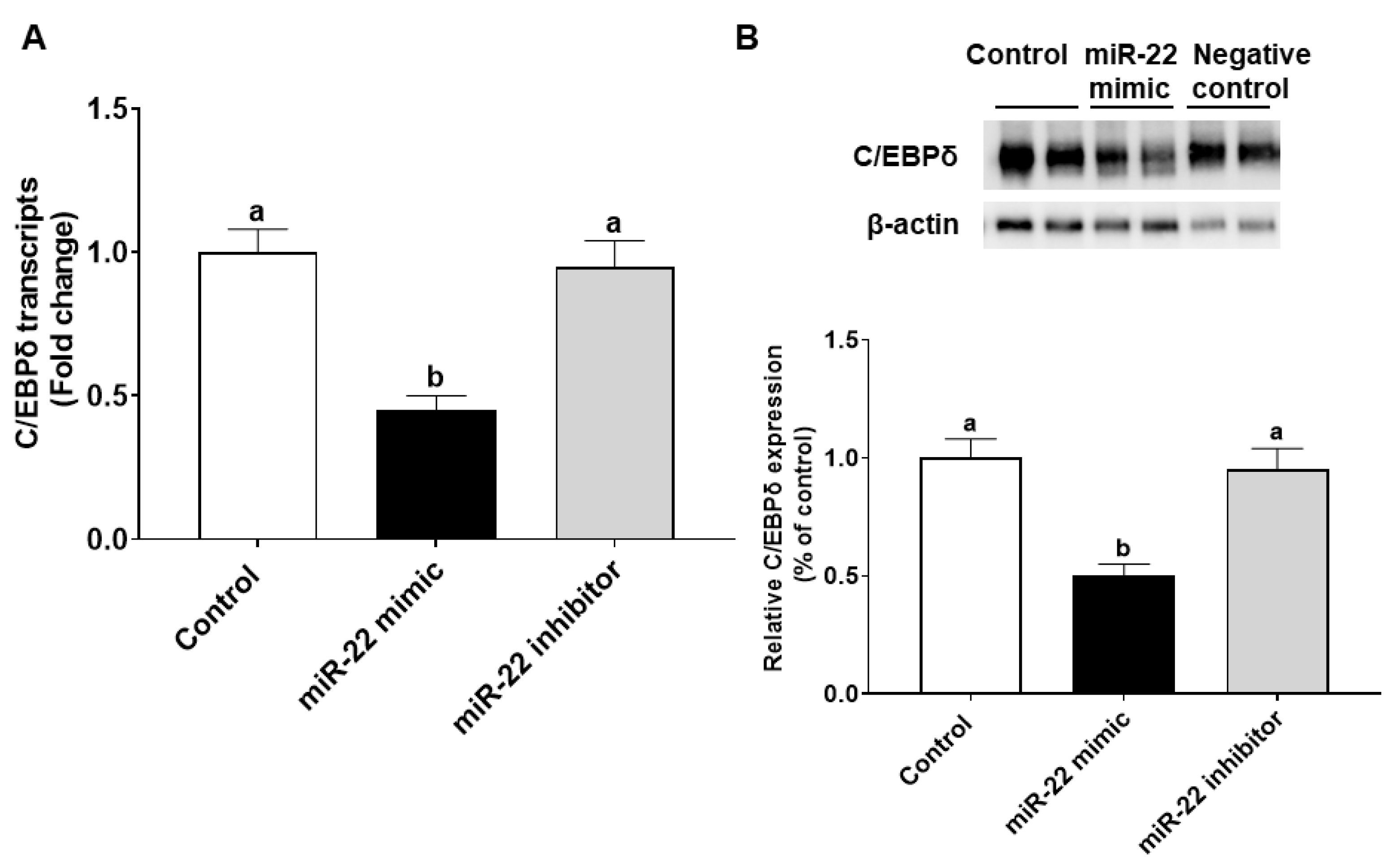
2.3. miR-22 Transfection and Verification
2.4. Microarray and qRT-PCR Verification
2.5. Microarray Data Analysis
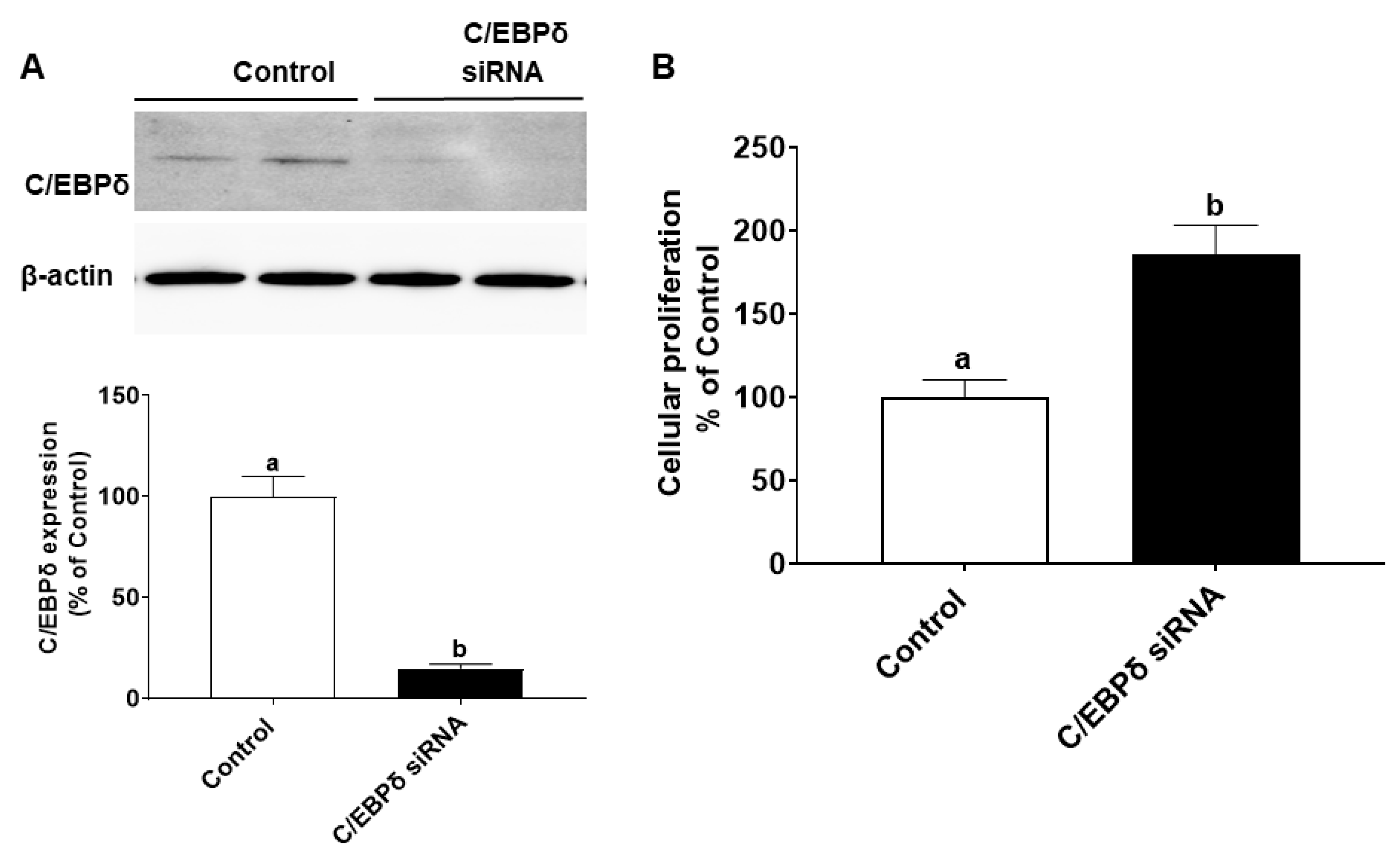
2.6. Proliferation Assay
2.7. Immunoblotting
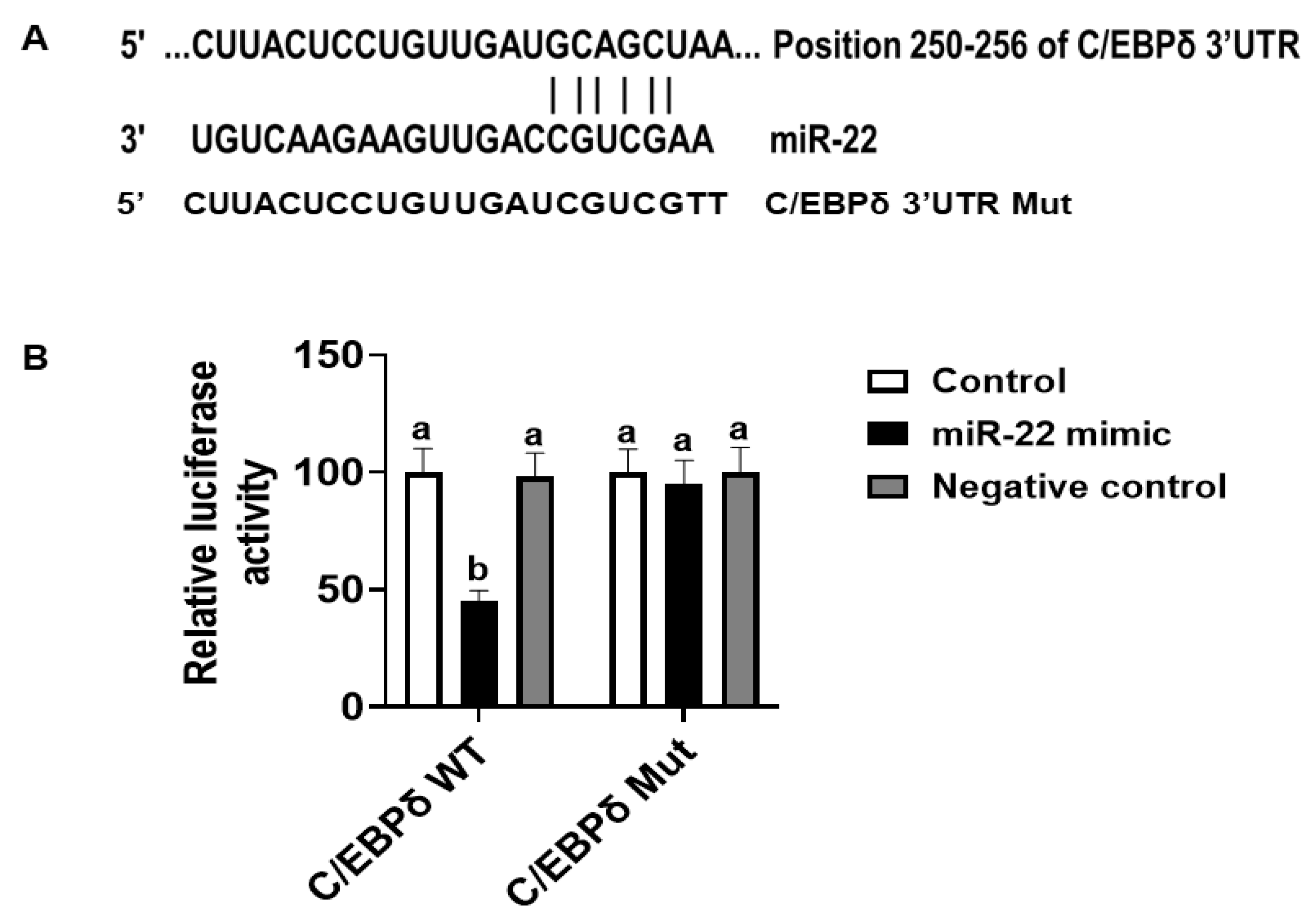
2.8. Construction of C/EBPδ 3′-UTR Reporter Vector and Luciferase Assay
2.9. RNA Interference
2.10. Statistical Analysis
3. Results
3.1. miR-22 Survives in vitro Digestion
3.2. Whole Genome Transcriptome Analysis Reveals That miR-22 Significantly Regulates mRNA Levels in HIECs
3.3. miR-22 Promotes Intestinal Proliferation
3.4. miR-22 May Promote Proliferation of HIECs by Directly Targeting the C/EBPδ Gene
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lönnerdal, B. Human Milk MicroRNAs/Exosomes: Composition and Biological Effects. In Nestlé Nutrition Institute Workshop Series; Donovan, S.M., German, J.B., Lönnerdal, B., Lucas, A., Eds.; S. Karger AG: Basel, Switzerland, 2019; Volume 90, pp. 83–92. ISBN 978-3-318-06340-0. [Google Scholar]
- Lönnerdal, B. Bioactive Proteins in Human Milk: Health, Nutrition, and Implications for Infant Formulas. J. Pediatr. 2016, 173, S4–S9. [Google Scholar] [CrossRef]
- Krol, J.; Loedige, I.; Filipowicz, W. The Widespread Regulation of MicroRNA Biogenesis, Function and Decay. Nat. Rev. Genet. 2010, 11, 597–610. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Friedman, R.C.; Farh, K.K.-H.; Burge, C.B.; Bartel, D.P. Most Mammalian MRNAs Are Conserved Targets of MicroRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef]
- Garcia, C.; Duan, R.D.; Brévaut-Malaty, V.; Gire, C.; Millet, V.; Simeoni, U.; Bernard, M.; Armand, M. Bioactive Compounds in Human Milk and Intestinal Health and Maturity in Preterm Newborn: An Overview. Cell. Mol. Biol. 2013, 59, 108–131. [Google Scholar] [PubMed]
- Liao, Y.; Du, X.; Li, J.; Lönnerdal, B. Human Milk Exosomes and Their MicroRNAs Survive Digestion in Vitro and Are Taken up by Human Intestinal Cells. Mol. Nutr. Food Res. 2017, 61, 1700082. [Google Scholar] [CrossRef]
- Kahn, S.; Liao, Y.; Du, X.; Xu, W.; Li, J.; Lönnerdal, B. Exosomal MicroRNAs in Milk from Mothers Delivering Preterm Infants Survive in Vitro Digestion and Are Taken Up by Human Intestinal Cells. Mol. Nutr. Food Res. 2018, 62, 1701050. [Google Scholar] [CrossRef]
- Raymond, F.; Lefebvre, G.; Texari, L.; Pruvost, S.; Metairon, S.; Cottenet, G.; Zollinger, A.; Mateescu, B.; Billeaud, C.; Picaud, J.-C.; et al. Longitudinal Human Milk MiRNA Composition over the First 3 Mo of Lactation in a Cohort of Healthy Mothers Delivering Term Infants. J. Nutr. 2022, 152, 94–106. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Zhao, H.; Han, X.; Zhao, T.; Qu, P.; Li, G.; Wang, W. Extracellular Vesicle-Encapsulated MiR-22-3p from Bone Marrow Mesenchymal Stem Cell Promotes Osteogenic Differentiation via FTO Inhibition. Stem Cell. Res. Ther. 2020, 11, 227. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Liu, Y.; Zhu, Z. MiR-22-3p Targeting Alpha-Enolase 1 Regulates the Proliferation of Retinoblastoma Cells. Biomed. Pharmacother. 2018, 105, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Takeshita, F.; Hino, Y.; Fukunaga, S.; Kudo, Y.; Tamaki, A.; Matsunaga, J.; Takahashi, R.; Takata, T.; Shimamoto, A.; et al. MiR-22 Represses Cancer Progression by Inducing Cellular Senescence. J. Cell Biol. 2011, 193, 409–424. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tang, J.; Li, C.; Kong, J.; Wang, J.; Wu, Y.; Xu, E.; Lai, M. MiR-22 Regulates 5-FU Sensitivity by Inhibiting Autophagy and Promoting Apoptosis in Colorectal Cancer Cells. Cancer Lett. 2015, 356, 781–790. [Google Scholar] [CrossRef]
- Xia, P.; Chen, J.; Liu, Y.; Cui, X.; Wang, C.; Zong, S.; Wang, L.; Lu, Z. MicroRNA-22-3p Ameliorates Alzheimer’s Disease by Targeting SOX9 through the NF-ΚB Signaling Pathway in the Hippocampus. J. Neuroinflamm. 2022, 19, 180. [Google Scholar] [CrossRef] [PubMed]
- Brusselle, G.G.; Bracke, K.R. MicroRNA MiR-22 Drives TH17 Responses in Emphysema. Nat. Immunol. 2015, 16, 1109–1110. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Liu, J.; Chen, P.; Lin, L.; Luo, Y.; Ma, X.; Lin, J.; Shen, Y.; Zhang, L. Exosomal MiR-22-3p from Human Umbilical Cord Blood-Derived Mesenchymal Stem Cells Protects against Lipopolysaccharid-Induced Acute Lung Injury. Life Sci. 2021, 269, 119004. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J. Emerging Roles of MicroRNA-22 in Human Disease and Normal Physiology. CMM 2012, 12, 247–258. [Google Scholar] [CrossRef] [PubMed]
- Balamurugan, K.; Sterneck, E. The Many Faces of C/EBPδ and Their Relevance for Inflammation and Cancer. Int. J. Biol. Sci. 2013, 9, 917–933. [Google Scholar] [CrossRef]
- Pulido-Salgado, M.; Vidal-Taboada, J.M.; Saura, J. C/EBPβ and C/EBPδ Transcription Factors: Basic Biology and Roles in the CNS. Prog. Neurobiol. 2015, 132, 1–33. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Fu, Q.; Shah, S.K.; Melnyk, S.B.; Sterneck, E.; Hauer-Jensen, M.; Pawar, S.A. C/EBPδ Protects from Radiation-Induced Intestinal Injury and Sepsis by Suppression of Inflammatory and Nitrosative Stress. Sci. Rep. 2019, 9, 13953. [Google Scholar] [CrossRef]
- Hartl, L.; Duitman, J.; Aberson, H.L.; Chen, K.; Dijk, F.; Roelofs, J.J.T.H.; Dings, M.P.G.; Hooijer, G.K.J.; Hernanda, P.Y.; Pan, Q.; et al. CCAAT/Enhancer-Binding Protein Delta (C/EBPδ): A Previously Unrecognized Tumor Suppressor That Limits the Oncogenic Potential of Pancreatic Ductal Adenocarcinoma Cells. Cancers 2020, 12, 2546. [Google Scholar] [CrossRef] [PubMed]
- Perreault, N.; Beaulieu, J.-F. Use of the Dissociating Enzyme Thermolysin to Generate Viable Human Normal Intestinal Epithelial Cell Cultures. Exp. Cell Res. 1996, 224, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Lönnerdal, B.; Jiang, R.; Du, X. Bovine Lactoferrin Can Be Taken Up by the Human Intestinal Lactoferrin Receptor and Exert Bioactivities. J. Pediatric Gastroenterol. Nutr. 2011, 53, 606–614. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma Powers Differential Expression Analyses for RNA-Sequencing and Microarray Studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Drozdowski, L.A.; Clandinin, T.; Thomson, A.B.R. Ontogeny, Growth and Development of the Small Intestine: Understanding Pediatric Gastroenterology. World J. Gastroenterol. 2010, 16, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Abbas, T.; Dutta, A. P21 in Cancer: Intricate Networks and Multiple Activities. Nat. Rev. Cancer 2009, 9, 400–414. [Google Scholar] [CrossRef] [PubMed]
- Timchenko, N.A.; Wilde, M.; Nakanishi, M.; Smith, J.R.; Darlington, G.J. CCAAT/Enhancer-Binding Protein Alpha (C/EBP Alpha) Inhibits Cell Proliferation through the P21 (WAF-1/CIP-1/SDI-1) Protein. Genes Dev. 1996, 10, 804–815. [Google Scholar] [CrossRef] [PubMed]
- Matys, V. TRANSFAC(R) and Its Module TRANSCompel(R): Transcriptional Gene Regulation in Eukaryotes. Nucleic Acids Research 2006, 34, D108–D110. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Hannon, G.J.; Zhang, H.; Casso, D.; Kobayashi, R.; Beach, D. P21 Is a Universal Inhibitor of Cyclin Kinases. Nature 1993, 366, 701–704. [Google Scholar] [CrossRef]
- Melnik, B.C.; Kakulas, F.; Geddes, D.T.; Hartmann, P.E.; John, S.M.; Carrera-Bastos, P.; Cordain, L.; Schmitz, G. Milk MiRNAs: Simple Nutrients or Systemic Functional Regulators? Nutr. Metab. 2016, 13, 42. [Google Scholar] [CrossRef]
- López de las Hazas, M.-C.; del Pozo-Acebo, L.; Hansen, M.S.; Gil-Zamorano, J.; Mantilla-Escalante, D.C.; Gómez-Coronado, D.; Marín, F.; Garcia-Ruiz, A.; Rasmussen, J.T.; Dávalos, A. Dietary Bovine Milk MiRNAs Transported in Extracellular Vesicles Are Partially Stable during GI Digestion, Are Bioavailable and Reach Target Tissues but Need a Minimum Dose to Impact on Gene Expression. Eur. J. Nutr. 2022, 61, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Stremmel, W.; Weiskirchen, R.; Melnik, B.C. Milk Exosomes Prevent Intestinal Inflammation in a Genetic Mouse Model of Ulcerative Colitis: A Pilot Experiment. Inflamm. Intest. Dis. 2020, 5, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Benmoussa, A.; Lee, C.H.C.; Laffont, B.; Savard, P.; Laugier, J.; Boilard, E.; Gilbert, C.; Fliss, I.; Provost, P. Commercial Dairy Cow Milk MicroRNAs Resist Digestion under Simulated Gastrointestinal Tract Conditions. J. Nutr. 2016, 146, 2206–2215. [Google Scholar] [CrossRef] [PubMed]
- Hamosh, M. A Review. Fat Digestion in the Newborn: Role of Lingual Lipase and Preduodenal Digestion. Pediatr. Res. 1979, 13, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C.; Stremmel, W.; Weiskirchen, R.; John, S.M.; Schmitz, G. Exosome-Derived MicroRNAs of Human Milk and Their Effects on Infant Health and Development. Biomolecules 2021, 11, 851. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liu, G.; Zhu, X. Human Breast Milk-Derived Exosomes May Help Maintain Intestinal Epithelial Barrier Integrity. Pediatr. Res. 2021, 90, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Miyake, H.; Lee, C.; Chusilp, S.; Bhalla, M.; Li, B.; Pitino, M.; Seo, S.; O’Connor, D.L.; Pierro, A. Human Breast Milk Exosomes Attenuate Intestinal Damage. Pediatr. Surg. Int. 2020, 36, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Van Herwijnen, M.J.C.; Driedonks, T.A.P.; Snoek, B.L.; Kroon, A.M.T.; Kleinjan, M.; Jorritsma, R.; Pieterse, C.M.J.; Hoen, E.N.M.N.-‘t.; Wauben, M.H.M. Abundantly Present MiRNAs in Milk-Derived Extracellular Vesicles Are Conserved Between Mammals. Front. Nutr. 2018, 5, 81. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Gao, C.; Li, H.; Huang, L.; Sun, Q.; Dong, Y.; Tian, C.; Gao, S.; Dong, H.; Guan, D.; et al. Identification and Characterization of MicroRNAs in Raw Milk during Different Periods of Lactation, Commercial Fluid, and Powdered Milk Products. Cell. Res. 2010, 20, 1128–1137. [Google Scholar] [CrossRef]
- Wijndaele, K.; Lakshman, R.; Landsbaugh, J.R.; Ong, K.K.; Ogilvie, D. Determinants of Early Weaning and Use of Unmodified Cow’s Milk in Infants: A Systematic Review. J. Am. Diet. Assoc. 2009, 109, 2017–2028. [Google Scholar] [CrossRef]
- McCarthy, N.A.; Magan, J.B.; Kelleher, C.M.; Kelly, A.L.; O’Mahony, J.A.; Murphy, E.G. Heat Treatment of Milk: Effect on Concentrate Viscosity, Powder Manufacture and End-Product Functionality. Int. Dairy J. 2022, 128, 105289. [Google Scholar] [CrossRef]
- Yuan, S.; Zhang, P.; Wen, L.; Jia, S.; Wu, Y.; Zhang, Z.; Guan, L.; Yu, Z.; Zhao, L. MiR-22 Promotes Stem Cell Traits via Activating Wnt/β-Catenin Signaling in Cutaneous Squamous Cell Carcinoma. Oncogene 2021, 40, 5799–5813. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, C. Exosomes MiR-22-3p Derived from Mesenchymal Stem Cells Suppress Colorectal Cancer Cell Proliferation and Invasion by Regulating RAP2B and PI3K/AKT Pathway. J. Oncol. 2021, 2021, 3874478. [Google Scholar] [CrossRef]
- Yang, X.; Su, W.; Li, Y.; Zhou, Z.; Zhou, Y.; Shan, H.; Han, X.; Zhang, M.; Zhang, Q.; Bai, Y.; et al. MiR-22-3p Suppresses Cell Growth via MET/STAT3 Signaling in Lung Cancer. Am. J. Transl. Res. 2021, 13, 1221–1232. [Google Scholar] [PubMed]
- Chen, H.; Lu, Q.; Fei, X.; Shen, L.; Jiang, D.; Dai, D. MiR-22 Inhibits the Proliferation, Motility, and Invasion of Human Glioblastoma Cells by Directly Targeting SIRT1. Tumor Biol. 2016, 37, 6761–6768. [Google Scholar] [CrossRef] [PubMed]
- Naakka, E.; Barros-Filho, M.C.; Adnan-Awad, S.; Al-Samadi, A.; Marchi, F.A.; Kuasne, H.; Korelin, K.; Suleymanova, I.; Brown, A.L.; Scapulatempo-Neto, C.; et al. MiR-22 and MiR-205 Drive Tumor Aggressiveness of Mucoepidermoid Carcinomas of Salivary Glands. Front. Oncol. 2022, 11, 786150. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Jiang, Y.; Zhang, H.; Greenlee, A.R.; Yu, R.; Yang, Q. MiR-22 Functions as a Micro-Oncogene in Transformed Human Bronchial Epithelial Cells Induced by Anti-Benzo[a]Pyrene-7,8-Diol-9,10-Epoxide. Toxicol. Vitr. 2010, 24, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Hock, A.; Miyake, H.; Li, B.; Lee, C.; Ermini, L.; Koike, Y.; Chen, Y.; Määttänen, P.; Zani, A.; Pierro, A. Breast Milk-Derived Exosomes Promote Intestinal Epithelial Cell Growth. J. Pediatric Surg. 2017, 52, 755–759. [Google Scholar] [CrossRef]
- Gao, H.N.; Guo, H.Y.; Zhang, H.; Xie, X.L.; Wen, P.C.; Ren, F.Z. Yak-Milk-Derived Exosomes Promote Proliferation of Intestinal Epithelial Cells in an Hypoxic Environment. J. Dairy Sci. 2019, 102, 985–996. [Google Scholar] [CrossRef]
- Reif, S.; Elbaum Shiff, Y.; Golan-Gerstl, R. Milk-Derived Exosomes (MDEs) Have a Different Biological Effect on Normal Fetal Colon Epithelial Cells Compared to Colon Tumor Cells in a MiRNA-Dependent Manner. J. Transl. Med. 2019, 17, 325. [Google Scholar] [CrossRef] [PubMed]
- Tsukada, J.; Yoshida, Y.; Kominato, Y.; Auron, P.E. The CCAAT/Enhancer (C/EBP) Family of Basic-Leucine Zipper (BZIP) Transcription Factors Is a Multifaceted Highly-Regulated System for Gene Regulation. Cytokine 2011, 54, 6–19. [Google Scholar] [CrossRef]
- Gery, S.; Tanosaki, S.; Hofmann, W.-K.; Koppel, A.; Koeffler, H.P. C/EBPδ Expression in a BCR-ABL-Positive Cell Line Induces Growth Arrest and Myeloid Differentiation. Oncogene 2005, 24, 1589–1597. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, T.; Yan, P.; Huang, T.; DeWille, J. Identification and Characterization of CCAAT/Enhancer Binding Proteindelta (C/EBPdelta) Target Genes in G0 Growth Arrested Mammary Epithelial Cells. BMC Mol. Biol. 2008, 9, 83. [Google Scholar] [CrossRef] [PubMed]
- Shamloo; Usluer P21 in Cancer Research. Cancers 2019, 11, 1178. [CrossRef] [PubMed]
- Wang, H.; Zhang, Q.; Wang, B.; Wu, W.; Wei, J.; Li, P.; Huang, R. MiR-22 Regulates C2C12 Myoblast Proliferation and Differentiation by Targeting TGFBR1. Eur. J. Cell Biol. 2018, 97, 257–268. [Google Scholar] [CrossRef]
- Sun, R.; Liu, Z.; Han, L.; Yang, Y.; Wu, F.; Jiang, Q.; Zhang, H.; Ma, R.; Miao, J.; He, K.; et al. MiR-22 and MiR-214 Targeting BCL9L Inhibit Proliferation, Metastasis, and Epithelial-mesenchymal Transition by Down-regulating Wnt Signaling in Colon Cancer. FASEB J. 2019, 33, 5411–5424. [Google Scholar] [CrossRef] [PubMed]
- Ji, D.; Li, B.; Shao, Q.; Li, F.; Li, Z.; Chen, G. MiR-22 Suppresses BMP7 in the Development of Cirrhosis. Cell. Physiol. Biochem. 2015, 36, 1026–1036. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; You, R.; Yuan, X.; Yang, T.; Samuel, E.L.G.; Marcano, D.C.; Sikkema, W.K.A.; Tour, J.M.; Rodriguez, A.; Kheradmand, F.; et al. The MicroRNA MiR-22 Inhibits the Histone Deacetylase HDAC4 to Promote TH17 Cell–Dependent Emphysema. Nat. Immunol. 2015, 16, 1185–1194. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Sequence, | 5′-3′ | ||
|---|---|---|---|
| Gene | Forward | Reverse | Accession Number |
| C/EBPδ | GCCATGTACGACGACGAGAG | GAGTCGATGTAGGCGCTGAA | NM_005195.4 |
| TP53INP | CACCCGTGGGACTGATGAAT | GAGCTTCCACTCTGGGACTAC | NM_033285.2 |
| IFIT3 | TTTGAAGCAGGCCATTGAGC | CAGGCCCAAGAGAACCTTGA | NM_001549.2 |
| IFIT1 | AACCTCGTCATTGTCAGGCA | TCCGCTGCCCACTTAGAGAA | NM_022168.2 |
| CXCL10 | CCTTAAAACCAGAGGGGAGCA | TGTGGTCCATCCTTGGAAGC | NM_001565.1 |
| p21 | ACTTTGGAGTCCCCTCACCT | CCCTAGGCTGTGCTCACTTC | NM_000389.5 |
| GAPDH | GCTGAGTACGTCGTGGAGTC | AAATGAGCCCCAGCCTTCTC | NM_001289746.2 |
| Term | Annotated | p Value |
|---|---|---|
| G1/S transition of mitotic cell cycle | 200 | 0.00019 |
| Regulation of mitotic cell cycle phase transition | 218 | 0.0008 |
| Positive regulation of cell proliferation | 505 | 0.0019 |
| G2/M transition of mitotic cell cycle | 151 | 0.00264 |
| Positive regulation of cell cycle process | 109 | 0.00012 |
| Transmembrane receptor protein tyrosine kinase signaling pathway | 668 | 0.00224 |
| Cell division | 276 | 0.0072 |
| Positive regulation of gene expression | 630 | 0.0087 |
| Defense response to virus | 128 | 7.1 × 10-5 |
| Negative regulation of viral genome replication | 25 | 7.2 × 10-5 |
| Type I interferon signaling pathway | 44 | 0.00026 |
| Positive regulation of type I interferon production | 37 | 0.00429 |
| NIK/NF-kappa B signaling | 59 | 0.01305 |
| Positive regulation of interleukin-6 production | 15 | 0.0199 |
| Term | Annotated | p Value |
|---|---|---|
| Positive regulation of cell death | 228 | 0.00021 |
| Regulation of epithelial to mesenchymal transition | 22 | 0.00161 |
| Positive regulation of apoptotic process | 218 | 0.00214 |
| Regulation of release of cytochrome c from mitochondria | 16 | 0.00216 |
| Positive regulation of protein insertion into mitochondrial membrane involved in apoptotic signaling pathway | 13 | 0.00237 |
| Negative regulation of cytoskeleton organization | 55 | 0.00487 |
| Regulation of reactive oxygen species metabolic process | 53 | 0.00806 |
| Negative regulation of protein phosphorylation | 120 | 0.022 |
| Negative regulation of cell communication | 410 | 0.0288 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, R.; Lönnerdal, B. Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression. Nutrients 2022, 14, 4901. https://doi.org/10.3390/nu14224901
Jiang R, Lönnerdal B. Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression. Nutrients. 2022; 14(22):4901. https://doi.org/10.3390/nu14224901
Chicago/Turabian StyleJiang, Rulan, and Bo Lönnerdal. 2022. "Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression" Nutrients 14, no. 22: 4901. https://doi.org/10.3390/nu14224901
APA StyleJiang, R., & Lönnerdal, B. (2022). Milk-Derived miR-22-3p Promotes Proliferation of Human Intestinal Epithelial Cells (HIECs) by Regulating Gene Expression. Nutrients, 14(22), 4901. https://doi.org/10.3390/nu14224901