
Mushroom Polysaccharides as Potential Candidates for Alleviating Neurodegenerative Diseases

Abstract
1. Introduction
2. Retrieval Strategy
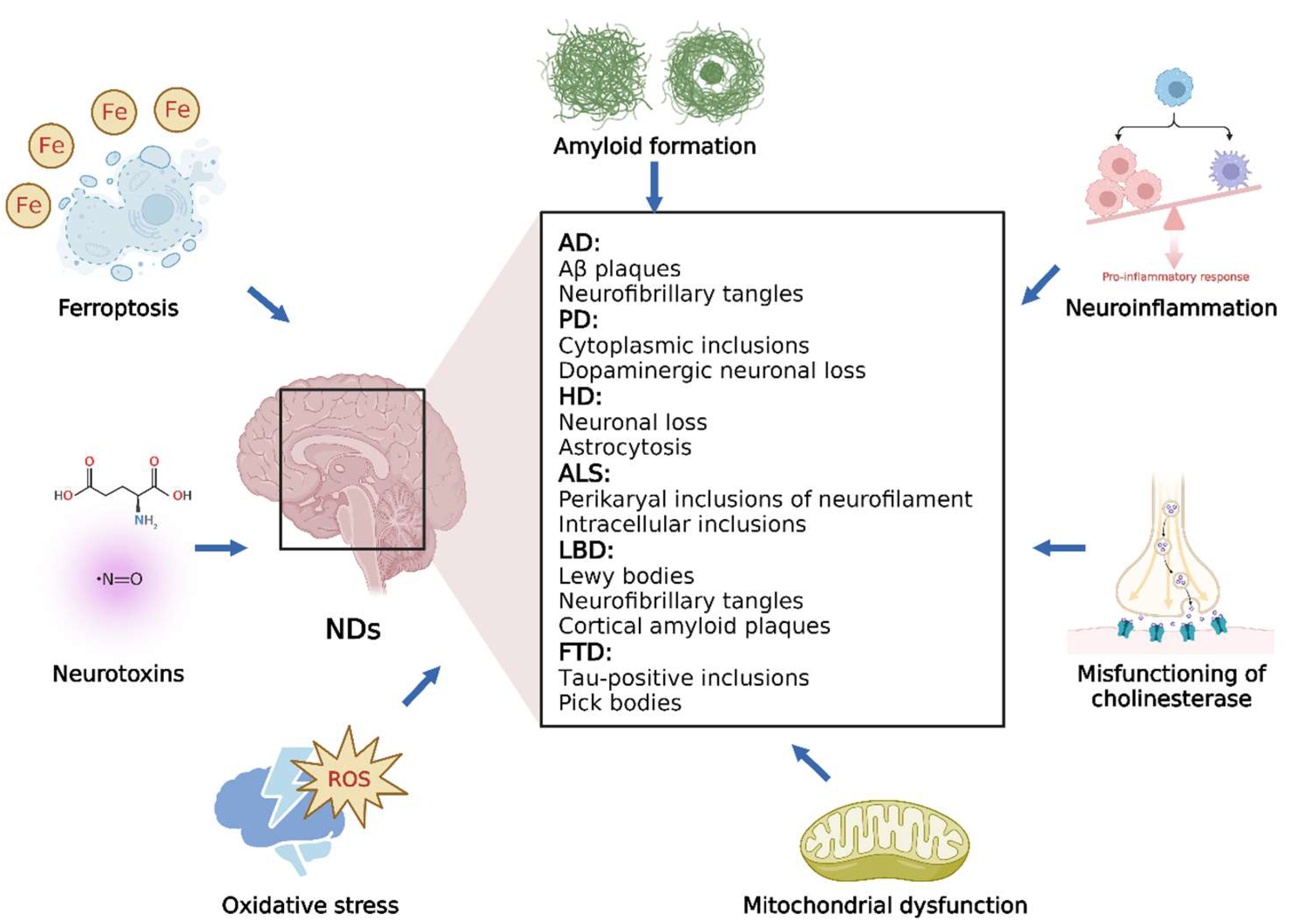
3. Mechanisms of NDs
3.1. Overview of NDs
3.2. Different Processes Involved in NDs
3.2.1. OS
3.2.2. Amyloid Formation
3.2.3. Neuroinflammation
3.2.4. Misfunctioning of Cholinesterase
3.2.5. Mitochondrial Dysfunction
3.2.6. Neurotoxins
3.2.7. Ferroptosis
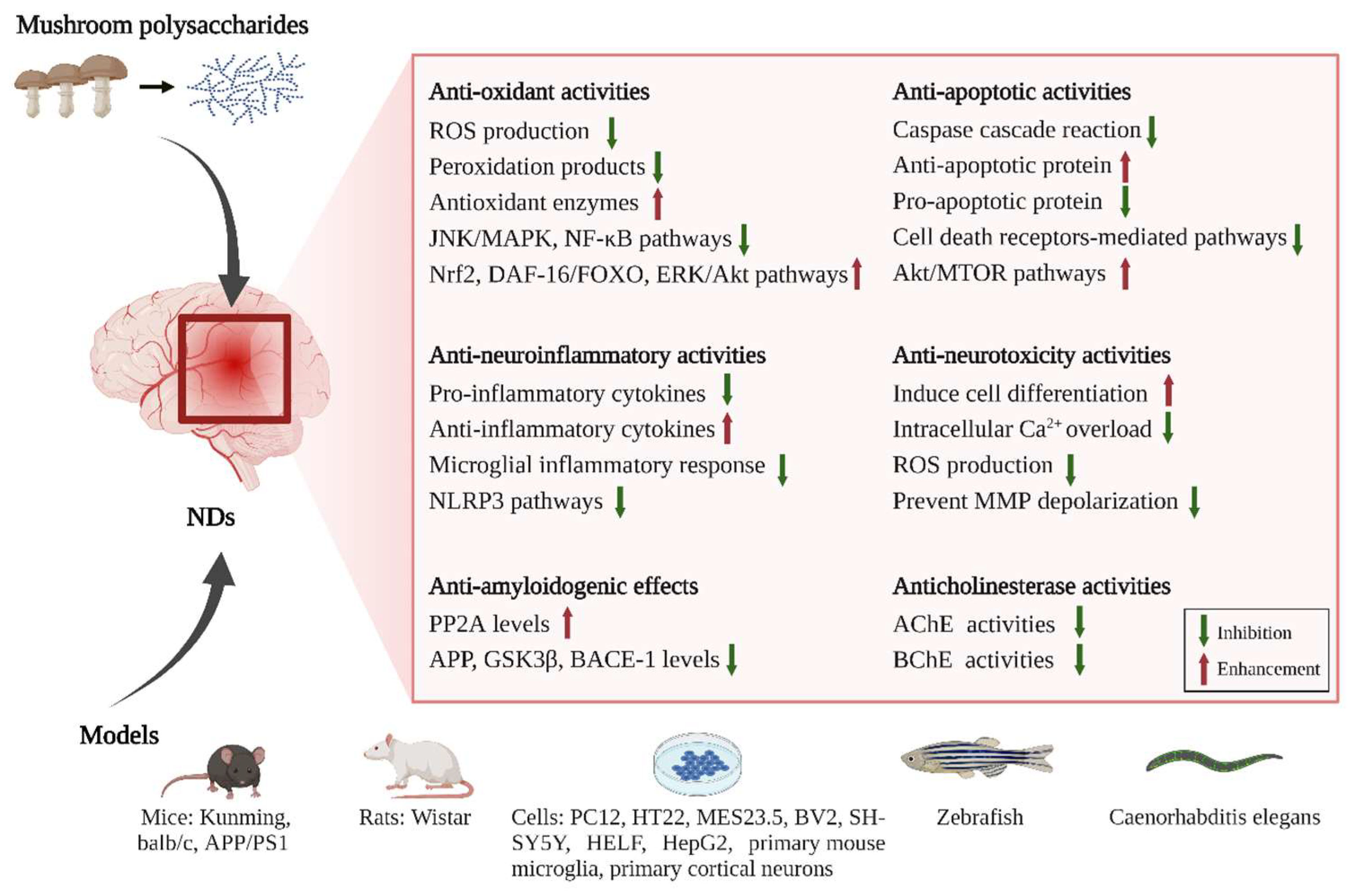
4. Neuroprotective and Therapeutic Effects of Mushroom Polysaccharides
4.1. Antioxidant Activities of Mushroom Polysaccharides
4.2. Anti-Amyloidogenic Effects of Mushroom Polysaccharides
4.3. Anti-Neuroinflammatory Activities of Mushroom Polysaccharides
4.4. Anticholinesterase Activities of Mushroom Polysaccharides
4.5. Anti-Apoptotic Activities of Mushroom Polysaccharides
4.6. Anti-Neurotoxic Activities of Mushroom Polysaccharides
4.7. Anti-Ferroptosis Activities of Mushroom Polysaccharides

{kind=link}
{kind=link}
| Mechanism | Source (Latin Name) | Polysaccharide | Molecular Weight (kDa) | Monosaccharide Composition | Cell Lines/Model | Type of NDs | Potential Mechanism | Year/References |
|---|---|---|---|---|---|---|---|---|
| Anti-oxidant activities | Inonotus obliquus | IOPS | 111.9 | / | L-Glu damaged HT22 cells; APP/PS1 mice | AD | Regulating Nrf2 signaling and exerting antioxidative and antiapoptotic effects | 2019 [61] |
| IOP | / | / | Tacrine-induced HepG2 cells | AD | Inhibition of ROS generation, 8-OHdG formation in mitochondrial DNA, and loss of the mitochondrial transmembrane potential, decrease in the cytochrome c release and activation of caspase-3 | 2019 [90] | ||
| Ganoderma lucidum | GLP | 15.0 | / | APP/PS1 mice | AD | Reduce Aβ deposits, increase protein levels of p-FGFR1, p-ERK and p-Akt, potentiate FGFR pathways | 2017 [63] | |
| GLA | / | Gal and Glc (1.0:8.3) | Aβ25-35- or Aβ42-exposed rat primary cortical neurons | AD | Antagonize Aβ peptide neurotoxicity, inhibit JNK, ERK and p38 MAPK pathways | 2008 [64] | ||
| GLP | / | / | Kainic acid-treated Wistar rats | Epilepsy | Inhibit calcium overloading and ERK1/2 and NF-κB expression; stimulate CaMK II α and Cav-1 expression | 2015 [65] | ||
| Ganoderma atrum | PSG-1 | 1013 | Man, Gal and Glc (1:1.28:4.91) | D-Gal-treated Kunming mice | Aging | Decrease MDA and GSSG levels, increase SOD, CAT, GPx and GSH activities in liver, brain and spleen | 2012 [56] | |
| Dictyophora indusiata | DiPS | / | Man, Fuc, Glc, Gal, Rha, Glc-UA and Xyl (86.8:4.5:3.9:1.6:1.2:1.1:0.9) | Caenorhabditis elegans | NDs | Decrease ROS and MDA levels, increase SOD activity, restore MMP and ATP content, regulate DAF-16/FOXO pathways | 2016 [26] | |
| Pleurotus ostreatus | POP | / | / | D-Gal and AlCl3-treated Wistar rats | AD | Decrease MDA content, increase SOD, GPx and CAT activities in hippocampus, liver and serum | 2016 [57] | |
| Tricholoma lobayense | TLH-3 | 4.23 | Rha, Man, Glc-UA, Gal-UA, Glc, Gal and Ara (0.07:0.23:0.02:0.02:1.57:1:0.11) | t-BHP-exposed HELF cells; D-Gal-treated Kunming mice | Aging | Decrease ROS level and inhibit oxidative damage induced by tert-butylhydroperoxide in HELF cells; decrease MDA content and increase SOD and CAT activities in mouse liver and serum | 2016 [55] | |
| Auricularia auricula-judae | APP1-a | 206 | Rha, Ara, Xyl, Man, Glc and Gal (0.2:2.6:0.4:3.6:1.0:0.4) | D-Gal-treated Kunming mice | Aging | Decrease MDA content, increase SOD and GPx activities in liver, serum and heart | 2011 [58] | |
| Amanita caesarea | ACPS | 18.62 | Xyl, Man, Gal and Glc | L-Glu exposed HT22 cells; D-Gal and AlCl3-treated balb/c mice | AD | Modulate Nrf2 pathways | 2019 [60] | |
| Flammulina velutipes | FVP | / | Man, Rib, Glc, Gal and Xyl (4.07:4.54:3.07:1:2.21) | D-Gal induced Wistar rats | AD | Increase SOD, CAT and GPx levels, decrease MDA levels; anti-apoptosis | 2018 [59] | |
| Cantharellus cibarius | CC2a CC3 | / | CC2a (Fuc, GlcN, Gal, Glc, Man) CC3 (Glc, Man, Rib) | Human undifferentiated neuroblastoma cell line SH-SY5Y | NDs | Shown antioxidant capacity, effectively neutralize the negative changes induced by activators of glutamatergic system (glutamate, NMDA, AMPA) | 2018 [32] | |
| Anti-amyloidogenic effects | Pleurotus ostreatus | POP | / | / | D-Gal and Al-treated Wistar rats | AD | Decrease Aβ peptide formation and tau phosphorylation by elevating the expression of PP2A and by reducing the expression of APP, BACE1 and GSK3β | 2016 [57] |
| Anti-neuroinflammation | Ganoderma lucidum | GLP | 15 | / | BV2 microglia and primary mouse microglia; zebrafish | AD | Decrease pro-inflammatory cytokines and promotes anti-inflammatory cytokine expressions in BV-2 and primary microglia; attenuate microglial migration, morphological alterations and phagocytosis probabilities | 2017 [71] |
| Antrodia camphorata | APC | / | / | 6-hydroxydopamine treated C57BL/6J mice | PD | Reduce the activation of NLRP3 and the expression of related inflammatory factors | 2019 [72] | |
| APC | / | / | Dopaminergic neuron cell line MES23.5; 6-hydroxydopamine treated mice | PD | Inhibit ROS-NLRP3 signaling | 2020 [73] | ||
| Amanita caesarea | ACPS2 | 16.6 | Gal, Glc and Man (35.40: 31.77: 29.47) | APP/PS1 mice | AD | Reduce inflammatory cell infiltration in brains, decrease serum concentrations of TNF-α and IL-1β, regulate neuroinflammation by regulating Nrf2 signaling and inhibiting NF-κB activation | 2021 [91] | |
| Anticholinesterase activities | Hericium erinaceus | HE | / | / | L-Glu-exposed PC12 cells; AlCl3 and D-Gal-treated balb/c mice | AD | Enhance the Ach and ChAT concentrations in mouse serum and hypothalamus | 2016 [87] |
| Flammulina velutipes | FVP | / | / | Scopolamine-treated Wistar rats | Cognitive impairment | Elevate the expression of CaMK II and connexin 36, and then regulated the activities of ChAT and AChE to normalize the level of ACh | 2015 [92] | |
| Armillaria mellea | AMPS | / | / | L-Glu induced HT22 cell; AlCl3 and D-Gal-treated balb/c mice | AD | Increase Ach and ChAT concentrations, decrease AchE concentrations in serum and hypothalamus of mice | 2017 [83] | |
| Amanita caesarea | ACPS | 18.62 | Xyl, Man, Gal and Glc | L-Glu exposed HT22 cells; D-Gal and AlCl3-treated balb/c mice | AD | Decrease AchE levels, increase Ach and chAT levels, improve cholinergic neurotransmission | 2019 [60] | |
| Anti-apoptotic activities | Pleurotus eryngii | PEP | / | / | PC12 Cells induced by β-Amyloid; aging rats | Aging | Decrease intracellular calcium levels, and attenuated the β-amyloid-mediated cell apoptosis in PC12 cells; decrease iNOS, and COX-2 levels in aging rats | 2020 [93] |
| Armillaria mellea | AMPS | / | / | L-Glu induced HT22 cell; AlCl3 and D-Gal-treated balb/c mice | AD | enhanced cell viability, suppressed nuclear apoptosis, inhibited intracellular ROS accumulation, prevented caspase-3 activation, and restored MMP | 2017 [83] | |
| Tremella fuciformis | TL04 | 2033 | Rha, Man and Glc (1:5.04:1.87) | Glu-induced differentiated PC12 cells | NDs | Enhance Bcl-2 levels, suppress Bax expression and cytochrome c release, decrease activities of caspase-3 caspase-8, caspase-9 | 2016 [84] | |
| Amauroderma rugosum | AR | / | / | PC12 rat pheochromocytoma cells induced by 6-OHDA | Neurotoxicity | Upregulate the expressions of proapoptotic proteins and downregulate the Akt/mTOR and MEK/ERK dependent pathways | 2021 [85] | |
| Morchella importuna | MIP | H2O2-induced PC12 cells | NDs | Inhibit cell apoptosis via down-regulation of the NF-κB pathway and the p38-JNK pathway and activating of the ERK | 2016 [94] | |||
| Anti-neurotoxicity activities | Hericium erinaceus | HE | / | / | L-Glu induced PC12 Cells; AlCl3 and D-Gal-treated balb/c mice | AD | Increase cell survival, induce cell differentiation, block intracellular Ca2+ overload, inhibit ROS production and prevent MMP depolarization in L-Glu induced HT22 cell apoptosis model; reduce the apoptosis rate, Aβ deposition, oxidative damage, and p-Tau aggregations in the hippocampus of AD mouse | 2016 [87] |
5. Structure–Activity Relationship of Mushroom Polysaccharides
6. Limitations and Future Perspective
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Phan, C.-W.; David, P.; Naidu, M.; Wong, K.-H.; Sabaratnam, V. Therapeutic potential of culinary-medicinal mushrooms for the management of neurodegenerative diseases: Diversity, metabolite, and mechanism. Crit. Rev. Biotechnol. 2015, 35, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Gitler, A.D.; Dhillon, P.; Shorter, J. Neurodegenerative disease: Models, mechanisms, and a new hope. Dis. Model Mech. 2017, 10, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Fu, H.; Hardy, J.; Duff, K.E. Selective vulnerability in neurodegenerative diseases. Nat. Neurosci. 2018, 21, 1350–1358. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.-L.; Li, S.; Zhang, R.; Le, W.-D. Neuroprotective effects of naturally sourced bioactive polysaccharides: An update. Neural Regen. Res. 2022, 17, 1907–1912. [Google Scholar] [CrossRef]
- Spires-Jones, T.L.; Hyman, B.T. The Intersection of Amyloid Beta and Tau at Synapses in Alzheimer’s Disease. Neuron 2014, 82, 756–771. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, R.; Yang, Z.; Wen, Q.; Cao, X.; Zhao, N.; Yan, J. Protective Effects of Polysaccharides in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 917629. [Google Scholar] [CrossRef]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics. Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef]
- Ostadkarampour, M.; Putnins, E.E. Monoamine Oxidase Inhibitors: A Review of Their Anti-Inflammatory Therapeutic Potential and Mechanisms of Action. Front. Pharmacol. 2021, 12, 676239. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Vigo, D.E.; Olivar, N.; Vidal, M.F.; Brusco, L.I. Melatonin Therapy in Patients with Alzheimer’s Disease. Antioxidants 2014, 3, 245–277. [Google Scholar] [CrossRef]
- Nimmrich, V.; Eckert, A. Calcium channel blockers and dementia. Br. J. Pharmacol. 2013, 169, 1203–1210. [Google Scholar] [CrossRef]
- Blume, T.; Filser, S.; Jaworska, A.; Blain, J.F.; Koenig, G.; Moschke, K.; Lichtenthaler, S.F.; Herms, J. BACE1 Inhibitor MK-8931 Alters Formation but Not Stability of Dendritic Spines. Front. Aging Neurosci. 2018, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Balestrino, R.; Schapira, A.H.V. Parkinson disease. Eur. J. Neurol. 2020, 27, 27–42. [Google Scholar] [CrossRef]
- Dickson, D.W. Neuropathology of Parkinson disease. Park. Relat. Disord. 2018, 46 (Suppl. S1), S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Salamon, A.; Zádori, D.; Szpisjak, L.; Klivényi, P.; Vécsei, L. What is the impact of catechol-O-methyltransferase (COMT) on Parkinson’s disease treatment? Expert Opin. Pharmacother. 2022, 23, 1123–1128. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, R.; Mishra, N.; Rana, R.; Kaur, G.; Ghoneim, M.M.; Alshehri, S.; Mustafa, G.; Ahmad, J.; Alhakamy, N.A.; Mishra, A. Molecular Mechanisms and Therapeutic Strategies for Levodopa-Induced Dyskinesia in Parkinson’s Disease: A Perspective Through Preclinical and Clinical Evidence. Front. Pharm. 2022, 13, 805388. [Google Scholar] [CrossRef] [PubMed]
- Nagatsu, T.; Sawada, M. Molecular mechanism of the relation of monoamine oxidase B and its inhibitors to Parkinson’s disease: Possible implications of glial cells. J. Neural Transm. Suppl. 2006, 71, 53–65. [Google Scholar] [CrossRef]
- Jenner, P. Pharmacology of dopamine agonists in the treatment of Parkinson’s disease. Neurology 2002, 58, S1–S8. [Google Scholar] [CrossRef]
- Jimenez-Sanchez, M.; Licitra, F.; Underwood, B.R.; Rubinsztein, D.C. Huntington’s Disease: Mechanisms of Pathogenesis and Therapeutic Strategies. Cold Spring Harb. Perspect. Med. 2017, 7, a024240. [Google Scholar] [CrossRef]
- Potkin, K.T.; Potkin, S.G. New directions in therapeutics for Huntington disease. Future Neurol. 2018, 13, 101–121. [Google Scholar] [CrossRef]
- Kukharsky, M.S.; Skvortsova, V.I.; Bachurin, S.O.; Buchman, V.L. In a search for efficient treatment for amyotrophic lateral sclerosis: Old drugs for new approaches. Med. Res. Rev. 2021, 41, 2804–2822. [Google Scholar] [CrossRef]
- Chio, A.; Mazzini, L.; Mora, G. Disease-modifying therapies in amyotrophic lateral sclerosis. Neuropharmacology 2020, 167, 107986. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Ir, R.; Jeewon, R.; Doble, M.; Hyde, K.D.; Kaliappan, I.; Jeyaraman, R.; Reddi, R.N.; Krishnan, J.; Li, M.; et al. A Mechanistic Review on Medicinal Mushrooms-Derived Bioactive Compounds: Potential Mycotherapy Candidates for Alleviating Neurological Disorders. Planta Med. 2020, 86, 1161–1175. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-Z.; Liao, Y.; Li, W.; Guo, L.-M. Neuroprotective effects of ganoderma lucidum polysaccharides against oxidative stress-induced neuronal apoptosis. Neural Regen. Res. 2017, 12, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Shen, H.; Yang, Y.; Sheng, Y.; Wang, J.; Li, W.; Zhou, X.; Guo, L.; Zhai, L.; Guan, Q. Antrodia camphorata polysaccharide resists 6-OHDA-induced dopaminergic neuronal damage by inhibiting ROS-NLRP3 activation. Brain Behav. 2020, 10, e01824. [Google Scholar] [CrossRef]
- Bai, Y.; Chen, L.; Chen, Y.; Chen, X.; Dong, Y.; Zheng, S.; Zhang, L.; Li, W.; Du, J.; Li, H. A Maitake (Grifola frondosa) polysaccharide ameliorates Alzheimer’s disease-like pathology and cognitive impairments by enhancing microglial amyloid-β clearance. RSC Adv. 2019, 9, 37127–37135. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.W.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef]
- Bertram, L.; Tanzi, R.E. The genetic epidemidogy of neurodegenerative disease. J. Clin. Investig. 2005, 115, 1449–1457. [Google Scholar] [CrossRef]
- Kametani, F.; Hasegawa, M. Reconsideration of Amyloid Hypothesis and Tau Hypothesis in Alzheimer’s Disease. Front. Neurosci. 2018, 12, 25. [Google Scholar] [CrossRef]
- Hamilton, R.L. Lewy bodies in Alzheimer’s disease: A neuropathological review of 145 cases using alpha-synuclein immunohistochemistry. Brain Pathol. 2000, 10, 378–384. [Google Scholar] [CrossRef]
- Brun, A. Frontal lobe degeneration of non-Alzheimer type. I. Neuropathology. Arch. Gerontol. Geriatr. 1987, 6, 193–208. [Google Scholar] [CrossRef]
- Shukla, V.; Mishra, S.K.; Pant, H.C. Oxidative stress in neurodegeneration. Adv. Pharmacol. Sci. 2011, 2011, 572634. [Google Scholar] [CrossRef] [PubMed]
- Rego, A.C.; Oliveira, C.R. Mitochondrial dysfunction and reactive oxygen species in excitotoxicity and apoptosis: Implications for the pathogenesis of neurodegenerative diseases. Neurochem. Res. 2003, 28, 1563–1574. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.-H.; Fu, X.; Zhang, R.; Wang, Z.; Guo, M. Neuroprotective effects of plant polysaccharides: A review of the mechanisms. Int. J. Biol. Macromol. 2018, 106, 749–754. [Google Scholar] [CrossRef]
- Moosavi, F.; Hosseini, R.; Saso, L.; Firuzi, O. Modulation of neurotrophic signaling pathways by polyphenols. Drug Des. Dev. Ther. 2016, 10, 23–42. [Google Scholar] [CrossRef]
- Dhahri, M.; Alghrably, M.; Mohammed, H.A.; Badshah, S.L.; Noreen, N.; Mouffouk, F.; Rayyan, S.; Qureshi, K.A.; Mahmood, D.; Lachowicz, J.I.; et al. Natural Polysaccharides as Preventive and Therapeutic Horizon for Neurodegenerative Diseases. Pharmaceutics 2021, 14, 1. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.S.; Al Mamun, A.; Rahman, M.A.; Behl, T.; Perveen, A.; Hafeez, A.; Bin-Jumah, M.N.; Abdel-Daim, M.M.; Ashraf, G.M. Emerging Proof of Protein Misfolding and Interactions in Multifactorial Alzheimer’s Disease. Curr. Top. Med. Chem. 2020, 20, 2380–2390. [Google Scholar] [CrossRef]
- Liu, R.; Su, R.; Liang, M.; Huang, R.; Wang, M.; Qi, W.; He, Z. Physicochemical Strategies for Inhibition of Amyloid Fibril Formation: An Overview of Recent Advances. Curr. Med. Chem. 2012, 19, 4157–4174. [Google Scholar] [CrossRef] [PubMed]
- Forman, M.S.; Trojanowski, J.Q.; Lee, V.M.Y. Neurodegenerative diseases: A decade of discoveries paves the way for therapeutic breakthroughs. Nat. Med. 2004, 10, 1055–1063. [Google Scholar] [CrossRef]
- Xanthos, D.N.; Sandkuehler, J. Neurogenic neuroinflammation: Inflammatory CNS reactions in response to neuronal activity. Nat. Rev. Neurosci. 2014, 15, 43–53. [Google Scholar] [CrossRef]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflammation 2014, 11, 98. [Google Scholar] [CrossRef]
- Lyman, M.; Lloyd, D.G.; Ji, X.; Vizcaychipi, M.P.; Ma, D. Neuroinflammation: The role and consequences. Neurosci. Res. 2014, 79, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Wu, Q.; Mak, S.; Liu, E.Y.L.; Zheng, B.Z.Y.; Dong, T.T.X.; Pi, R.; Tsim, K.W.K. Regulation of acetylcholinesterase during the lipopolysaccharide-induced inflammatory responses in microglial cells. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2022, 36, e22189. [Google Scholar] [CrossRef] [PubMed]
- Tata, A.M.; Velluto, L.; D’Angelo, C.; Reale, M. Cholinergic System Dysfunction and Neurodegenerative Diseases: Cause or Effect? CNS Neurol. Disord. -Drug Targets 2014, 13, 1294–1303. [Google Scholar] [CrossRef]
- Stanga, S.; Caretto, A.; Boido, M.; Vercelli, A. Mitochondrial Dysfunctions: A Red Thread across Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 3719. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, W.; Li, L.; Perry, G.; Lee, H.-G.; Zhu, X. Oxidative stress and mitochondrial dysfunction in Alzheimer’s disease. Biochim. Et Biophys. Acta-Mol. Basis Dis. 2014, 1842, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef]
- Bove, J.; Perier, C. Neurotoxin-based models of parkinson’s disease. Neuroscience 2012, 211, 51–76. [Google Scholar] [CrossRef]
- Li, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases. Nutrients 2017, 9, 778. [Google Scholar] [CrossRef]
- Zhou, J.; Jin, Y.; Lei, Y.; Liu, T.; Wan, Z.; Meng, H.; Wang, H. Ferroptosis Is Regulated by Mitochondria in Neurodegenerative Diseases. Neurodegener. Dis. 2020, 20, 20–34. [Google Scholar] [CrossRef]
- Galluzzi, L.; Vitale, I.; Aaronson, S.A.; Abrams, J.M.; Adam, D.; Agostinis, P.; Alnemri, E.S.; Altucci, L.; Amelio, I.; Andrews, D.W.; et al. Molecular mechanisms of cell death: Recommendations of the Nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018, 25, 486–541. [Google Scholar] [CrossRef]
- Reichert, C.O.; de Freitas, F.A.; Sampaio-Silva, J.; Rokita-Rosa, L.; Barros, P.L.; Levy, D.; Bydlowski, S.P. Ferroptosis Mechanisms Involved in Neurodegenerative Diseases. Int. J. Mol. Sci. 2020, 21, 8765. [Google Scholar] [CrossRef]
- Hirschhorn, T.; Stockwell, B.R. The development of the concept of ferroptosis. Free Radic. Biol. Med. 2019, 133, 130–143. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Maier, O. Interrelation of Oxidative Stress and Inflammation in Neurodegenerative Disease: Role of TNF. Oxidative Med. Cell. Longev. 2015, 2015, 610813. [Google Scholar] [CrossRef]
- Fox, J.H.; Connor, T.; Stiles, M.; Kama, J.; Lu, Z.; Dorsey, K.; Liebermann, G.; Sapp, E.; Cherny, R.A.; Banks, M.; et al. Cysteine Oxidation within N-terminal Mutant Huntingtin Promotes Oligomerization and Delays Clearance of Soluble Protein. J. Biol. Chem. 2011, 286, 18320–18330. [Google Scholar] [CrossRef]
- Ding, Q.; Yang, D.; Zhang, W.; Lu, Y.; Zhang, M.; Wang, L.; Li, X.; Zhou, L.; Wu, Q.; Pan, W.; et al. Antioxidant and anti-aging activities of the polysaccharide TLH-3 from Tricholoma lobayense. Int. J. Biol. Macromol. 2016, 85, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Li, W.-J.; Nie, S.-P.; Peng, X.-P.; Liu, X.-Z.; Li, C.; Chen, Y.; Li, J.-E.; Song, W.-R.; Xie, M.-Y. Ganoderma atrum Polysaccharide Improves Age-Related Oxidative Stress and Immune Impairment in Mice. J. Agric. Food Chem. 2012, 60, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, X.; Jin, G.; Yang, X.; Zhang, Y. Polysaccharides from Pleurotus ostreatus alleviate cognitive impairment in a rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 92, 935–941. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Z.-Y.; Zhang, Z.; Wang, X. Purified Auricularia auricular-judae polysaccharide (AAP I-a) prevents oxidative stress in an ageing mouse model. Carbohydr. Polym. 2011, 84, 638–648. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Yang, X.; Jin, G.; Zhang, Y. Cognitive-enhancing effect of polysaccharides from Flammulina velutipes on Alzheimer’s disease by compatibilizing with ginsenosides. Int. J. Biol. Macromol. 2018, 112, 788–795. [Google Scholar] [CrossRef]
- Li, Z.; Chen, X.; Zhang, Y.; Liu, X.; Wang, C.; Teng, L.; Wang, D. Protective roles of Amanita caesarea polysaccharides against Alzheimer’s disease via Nrf2 pathway. Int. J. Biol. Macromol. 2019, 121, 29–37. [Google Scholar] [CrossRef]
- Han, Y.; Nan, S.; Fan, J.; Chen, Q.; Zhang, Y. Inonotus obliquus polysaccharides protect against Alzheimer’s disease by regulating Nrf2 signaling and exerting antioxidative and antiapoptotic effects. Int. J. Biol. Macromol. 2019, 131, 769–778. [Google Scholar] [CrossRef] [PubMed]
- Furukawa-Hibi, Y.; Kobayashi, Y.; Chen, C.; Motoyama, N. FOXO transcription factors in cell-cycle regulation and the response to oxidative stress. Antioxid. Redox Signal. 2005, 7, 752–760. [Google Scholar] [CrossRef]
- Huang, S.; Mao, J.; Ding, K.; Zhou, Y.; Zeng, X.; Yang, W.; Wang, P.; Zhao, C.; Yao, J.; Xia, P.; et al. Polysaccharides from Ganoderma lucidum Promote Cognitive Function and Neural Progenitor Proliferation in Mouse Model of Alzheimer’s Disease. Stem Cell Rep. 2017, 8, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.S.-W.; Yu, M.-S.; Yuen, W.-H.; So, K.-F.; Zee, S.-Y.; Chang, R.C.-C. Antagonizing beta-amyloid peptide neurotoxicity of the anti-aging fungus Ganoderma lucidum. Brain Res. 2008, 1190, 215–224. [Google Scholar] [CrossRef]
- Zhou, S.; Wang, S.Q.; Sun, C.Y.; Mao, H.Y.; Di, W.H.; Ma, X.R.; Liu, L.; Liu, J.X.; Wang, F.F.; Kelly, P.; et al. Investigation into anti-epileptic effect and mechanisms of Ganoderma lucidum polysaccharides in in vivo and in vitro models. Proc. Nutr. Soc. 2015, 74, E65. [Google Scholar] [CrossRef]
- Hoshi, M. Multi-angle development of therapeutic methods for Alzheimer’s disease. Br. J. Pharm. 2021, 178, 770–783. [Google Scholar] [CrossRef] [PubMed]
- Lopez Sanchez, M.I.G.; van Wijngaarden, P.; Trounce, I.A. Amyloid precursor protein-mediated mitochondrial regulation and Alzheimer’s disease. Br. J. Pharm. 2019, 176, 3464–3474. [Google Scholar] [CrossRef]
- Dobson, C.M. The Amyloid Phenomenon and Its Links with Human Disease. Cold Spring Harb. Perspect. Biol. 2017, 9, a023648. [Google Scholar] [CrossRef]
- Vassar, R. Beta-Secretase, APP and Abeta in Alzheimer’s disease. Sub-Cell. Biochem. 2005, 38, 79–103. [Google Scholar]
- Barbalace, M.C.; Malaguti, M.; Giusti, L.; Lucacchini, A.; Hrelia, S.; Angeloni, C. Anti-Inflammatory Activities of Marine Algae in Neurodegenerative Diseases. Int. J. Mol. Sci. 2019, 20, 3061. [Google Scholar] [CrossRef]
- Cai, Q.; Li, Y.; Pei, G. Polysaccharides from Ganoderma lucidum attenuate microglia-mediated neuroinflammation and modulate microglial phagocytosis and behavioural response. J. Neuroinflammation 2017, 14, 63. [Google Scholar] [CrossRef] [PubMed]
- Han, C.; Guo, L.; Yang, Y.; Li, W.; Sheng, Y.; Wang, J.; Guan, Q.; Zhang, X. Study on antrodia camphorata polysaccharide in alleviating the neuroethology of PD mice by decreasing the expression of NLRP3 inflammasome. Phytother. Res. PTR 2019, 33, 2288–2297. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Singh, B. A review on cholinesterase inhibitors for Alzheimer’s disease. Arch. Pharmacal Res. 2013, 36, 375–399. [Google Scholar] [CrossRef] [PubMed]
- Pejin, B.; Tesanovic, K.; Jakovljevic, D.; Kaisarevic, S.; Sibul, F.; Raseta, M.; Karaman, M. The polysaccharide extracts from the fungi Coprinus comatus and Coprinellus truncorum do exhibit AChE inhibitory activity. Nat. Prod. Res. 2019, 33, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Deveci, E.; Tel-Çayan, G.; Duru, M.E.; Öztürk, M. Isolation, characterization, and bioactivities of compounds from Fuscoporia torulosa mushroom. J. Food Biochem. 2019, 43, e13074. [Google Scholar] [CrossRef]
- Badshah, S.L.; Riaz, A.; Muhammad, A.; Tel Cayan, G.; Cayan, F.; Emin Duru, M.; Ahmad, N.; Emwas, A.-H.; Jaremko, M. Isolation, Characterization, and Medicinal Potential of Polysaccharides of Morchella esculenta. Molecules 2021, 26, 1459. [Google Scholar] [CrossRef]
- Rahmani-Nezhad, S.; Dianat, S.; Mandizadeh, V.; Fooladi, Z.; Hariri, R.; Najafi, Z.; Firuzi, O.; Vahedi-Mazdabadi, Y.; Farjadmand, F.; Akbarzadeh, T.; et al. Investigation of polysaccharide extracts from Iranian and French strains of Agaricus subrufescens against enzymes involved in Alzheimer’s disease. Bol. Latinoam. Y Del Caribe De Plantas Med. Y Aromat. 2019, 18, 544–554. [Google Scholar] [CrossRef]
- Yuan, J.; Amin, P.; Ofengeim, D. Necroptosis and RIPK1-mediated neuroinflammation in CNS diseases. Nat. Rev. Neurosci. 2019, 20, 19–33. [Google Scholar] [CrossRef]
- Chi, H.; Chang, H.-Y.; Sang, T.-K. Neuronal Cell Death Mechanisms in Major Neurodegenerative Diseases. Int. J. Mol. Sci. 2018, 19, 3082. [Google Scholar] [CrossRef]
- Peña-Blanco, A.; García-Sáez, A.J. Bax, Bak and beyond-mitochondrial performance in apoptosis. FEBS J. 2018, 285, 416–431. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Tan, H.; Yang, P.; Li, Y.; Xu, L.; Duan, B.; Liu, Y. Advances in polysaccharides of natural source of the anti-Alzheimer’s disease effect and mechanism. Carbohydr. Polym. 2022, 296, 119961. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Mei, Q.; Wang, L.; Feng, X.; Tao, X.; Qiu, C.; Zhu, J. TIGAR suppresses seizures induced by kainic acid through inhibiting oxidative stress and neuronal apoptosis. Biochem. Biophys. Res. Commun. 2019, 515, 436–441. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Lu, W.; Zhang, Y.; Yuan, Q.; Wang, D. Pharmacological Basis for Use of Armillaria mellea Polysaccharides in Alzheimer’s Disease: Antiapoptosis and Antioxidation. Oxid. Med. Cell. Longev. 2017, 2017, 4184562. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Hu, X.; Zhang, Y.; Liu, T. Studies on the purification of polysaccharides separated from Tremella fuciformis and their neuroprotective effect. Mol. Med. Rep. 2016, 13, 3985–3992. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, R.; Wu, X.; Hoo, R.L.; Lee, S.M.; Cheung, T.M.; Ho, B.S.; Leung, G.P. Amauroderma rugosum Protects PC12 Cells against 6-OHDA-Induced Neurotoxicity through Antioxidant and Antiapoptotic Effects. Oxid. Med. Cell. Longev. 2021, 2021, 6683270. [Google Scholar] [CrossRef] [PubMed]
- Armada-Moreira, A.; Gomes, J.I.; Pina, C.C.; Savchak, O.K.; Goncalves-Ribeiro, J.; Rei, N.; Pinto, S.; Morais, T.P.; Martins, R.S.; Ribeiro, F.F.; et al. Going the Extra (Synaptic) Mile: Excitotoxicity as the Road Toward Neurodegenerative Diseases. Front. Cell. Neurosci. 2020, 14, 90. [Google Scholar] [CrossRef]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The Neuroprotective Properties of Hericium erinaceus in Glutamate-Damaged Differentiated PC12 Cells and an Alzheimer’s Disease Mouse Model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef]
- Lin, Y.; Yang, J.; Luo, L.; Zhang, X.; Deng, S.; Chen, X.; Li, Y.; Bekhit, A.E.A.; Xu, B.; Huang, R. Ferroptosis Related Immunomodulatory Effect of a Novel Extracellular Polysaccharides from Marine Fungus Aureobasidium melanogenum. Mar. Drugs 2022, 20, 332. [Google Scholar] [CrossRef]
- Li, W.; Zhou, X.; Xu, S.; Cao, N.; Li, B.; Chen, W.; Yang, B.; Yuan, M.; Xu, D. Lipopolysaccharide-induced splenic ferroptosis in goslings was alleviated by polysaccharide of atractylodes macrocephala koidz associated with proinflammatory factors. Poult. Sci. 2022, 101, 101725. [Google Scholar] [CrossRef]
- Li, Z.; Mei, J.; Jiang, L.; Geng, C.; Li, Q.; Yao, X.; Cao, J. Chaga Medicinal Mushroom Inonotus obliquus (Agaricomycetes) Polysaccharides Suppress Tacrine-Induced Apoptosis by Reactive Oxygen Species-Scavenging and Mitochondrial Pathway in HepG2 Cells. Int. J. Med. Mushrooms 2019, 21, 583–593. [Google Scholar] [CrossRef]
- Hu, W.; Li, Z.; Wang, W.; Song, M.; Dong, R.; Zhou, Y.; Li, Y.; Wang, D. Structural characterization of polysaccharide purified from Amanita caesarea and its pharmacological basis for application in Alzheimer’s disease: Endoplasmic reticulum stress. Food Funct. 2021, 12, 11009–11023. [Google Scholar] [CrossRef]
- Yang, W.; Yu, J.; Zhao, L.; Ma, N.; Fang, Y.; Pei, F.; Mariga, A.M.; Hu, Q. Polysaccharides from Flammulina velutipes improve scopolamine-induced impairment of learning and memory of rats. J. Funct. Foods 2015, 18, 411–422. [Google Scholar] [CrossRef]
- Zhang, C.J.; Guo, J.Y.; Cheng, H.; Lin, L.; Liu, Y.; Shi, Y.; Xu, J.; Yu, H.T. Protective Effects of the King Oyster Culinary-Medicinal Mushroom, Pleurotus eryngii (Agaricomycetes), Polysaccharides on β-Amyloid-Induced Neurotoxicity in PC12 Cells and Aging Rats, In Vitro and In Vivo Studies. Int. J. Med. Mushrooms 2020, 22, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Li, Q.; Chen, C.; Chen, Z.; Huang, W. Neuroprotective effect of crude polysaccharide isolated from the fruiting bodies of Morchella importuna against H(2)O(2)-induced PC12 cell cytotoxicity by reducing oxidative stress. Biomed. Pharmacother. 2016, 83, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Meng, W.; Li, L.; Meng, Z.; Wang, D. Adjuvant Therapy with Mushroom Polysaccharides for Diabetic Complications. Front. Pharmacol. 2020, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Y.-C.; Siu, K.-C.; Liu, Y.-S.; Wu, J.-Y. Molecular properties and antioxidant activities of polysaccharide–protein complexes from selected mushrooms by ultrasound-assisted extraction. Process Biochem. 2012, 47, 892–895. [Google Scholar] [CrossRef]
- You, R.; Wang, K.; Liu, J.; Liu, M.; Luo, L.; Zhang, Y. A comparison study between different molecular weight polysaccharides derived from Lentinus edodes and their antioxidant activities in vivo. Pharm. Biol. 2011, 49, 1298–1305. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, M.; Yang, Y.; Lin, L.; Xu, N.; Zhao, H.; Jia, L. Purification, characterization and hepatoprotective activities of mycelia zinc polysaccharides by Pleurotus djamor. Carbohydr. Polym. 2016, 136, 588–597. [Google Scholar] [CrossRef]
- Ren, Z.; Liu, W.; Song, X.; Qi, Y.; Zhang, C.; Gao, Z.; Zhang, J.; Jia, L. Antioxidant and anti-inflammation of enzymatic-hydrolysis residue polysaccharides by Lentinula edodes. Int. J. Biol. Macromol. 2018, 120, 811–822. [Google Scholar] [CrossRef]
- Mingyi, Y.; Belwal, T.; Devkota, H.P.; Li, L.; Luo, Z. Trends of utilizing mushroom polysaccharides (MPs) as potent nutraceutical components in food and medicine: A comprehensive review. Trends. Food Sci. Technol. 2019, 92, 94–110. [Google Scholar] [CrossRef]
- Tu, W.; Zhu, J.; Bi, S.; Chen, D.; Song, L.; Wang, L.; Zi, J.; Yu, R. Isolation, characterization and bioactivities of a new polysaccharide from Annona squamosa and its sulfated derivative. Carbohydr. Polym. 2016, 152, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Lu, Y.; Zhang, W.; Yuan, S.; Zhou, L.; Wang, L.; Ding, Q.; Wang, D.; Yang, W.; Cai, Z.; et al. Antioxidant capacity and cytotoxicity of sulfated polysaccharide TLH-3 from Tricholoma lobayense. Int. J. Biol. Macromol. 2016, 82, 913–919. [Google Scholar] [CrossRef] [PubMed]
- Sit, S.Y. Dopamine agonists in the treatment of Parkinson s disease past, present and future. Curr. Pharmaceutical Des. 2000, 6, 1211–1248. [Google Scholar] [CrossRef]
- Miller, R.G.; Mitchell, J.D.; Lyon, M.; Moore, D.H. Riluzole for amyotrophic lateral sclerosis (ALS)/motor neuron disease (MND). Amyotroph. Lateral Scler. 2003, 4, 191–206. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, X.; Li, S.; Feng, X.; Li, L.; Hao, J.; Wang, D.; Wang, Q. Mushroom Polysaccharides as Potential Candidates for Alleviating Neurodegenerative Diseases. Nutrients 2022, 14, 4833. https://doi.org/10.3390/nu14224833
Jiang X, Li S, Feng X, Li L, Hao J, Wang D, Wang Q. Mushroom Polysaccharides as Potential Candidates for Alleviating Neurodegenerative Diseases. Nutrients. 2022; 14(22):4833. https://doi.org/10.3390/nu14224833
Chicago/Turabian StyleJiang, Xue, Siqi Li, Xiangru Feng, Lanzhou Li, Jie Hao, Di Wang, and Qingshuang Wang. 2022. "Mushroom Polysaccharides as Potential Candidates for Alleviating Neurodegenerative Diseases" Nutrients 14, no. 22: 4833. https://doi.org/10.3390/nu14224833
APA StyleJiang, X., Li, S., Feng, X., Li, L., Hao, J., Wang, D., & Wang, Q. (2022). Mushroom Polysaccharides as Potential Candidates for Alleviating Neurodegenerative Diseases. Nutrients, 14(22), 4833. https://doi.org/10.3390/nu14224833