
Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts
 ,
,  ,
,  ,
,  ,
,  ,
,  , , ,
, , ,  ,
,  ,
,  and
and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Extract Preparation
2.3. Thermogravimetric Analysis (TGA)
2.4. Attenuated Total Reflectance (ATR) Fourier Transform Infrared (FT-IR) Spectroscopy
2.5. Free Amino Acids and Ergosterol Identification
2.6. Analysis of Monosaccharide Composition
2.7. Analysis of Ergothioneine
2.8. Yeast Strains and Media
2.9. Chronological Lifespan Experiments (CLS)
2.10. Yeast Protein Extraction, Cell Fractionation and Immunoblotting
2.11. Immunofluorescence Analysis
2.12. Heat Shock Assay
2.13. Analysis of Reactive Oxygen Species (ROS) Levels
2.14. Analysis of Aggresomes
2.15. RNA Extraction and qRT-PCR
2.16. FM4-64 Internalization
2.17. In Vitro α-syn Fibrillation
2.18. Morphological Characterization by Electron Microscopy Imaging
2.19. Thioflavin T (ThT) Assay
2.20. Fly Stocks and Husbandry
2.21. Drosophila melanogaster Longevity Assay
2.22. Western Blot Analysis of D. melanogaster Heads
3. Results
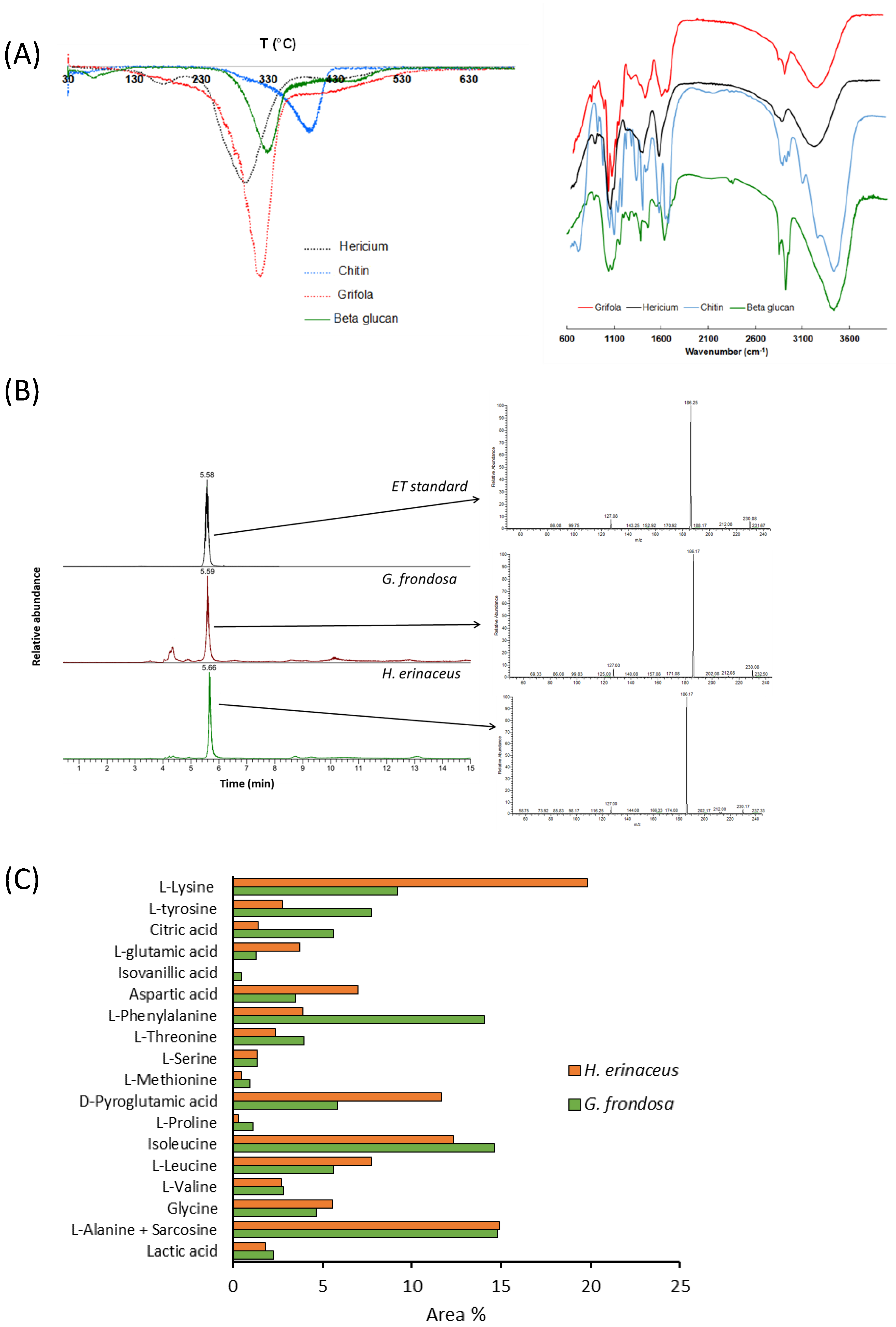
3.1. Analysis of Fungal Extract Composition
3.2. Fungal Extracts Extend Chronological Lifespan of Yeast Cells
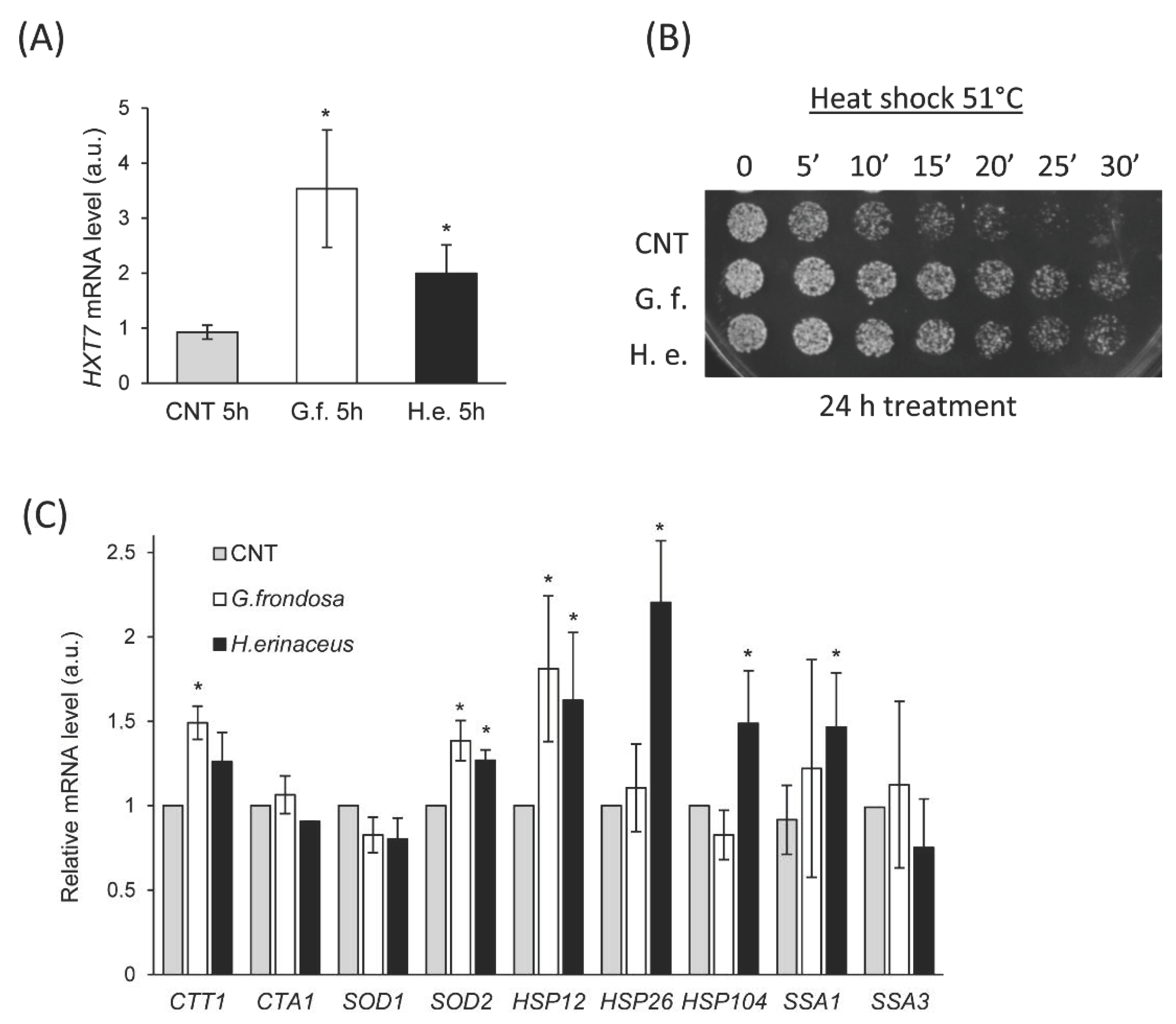
3.3. The Fungal Extracts Inhibit the Ras/PKA Pathway
3.4. Fungal Extracts Reduce α-Synuclein Toxicity in Yeast
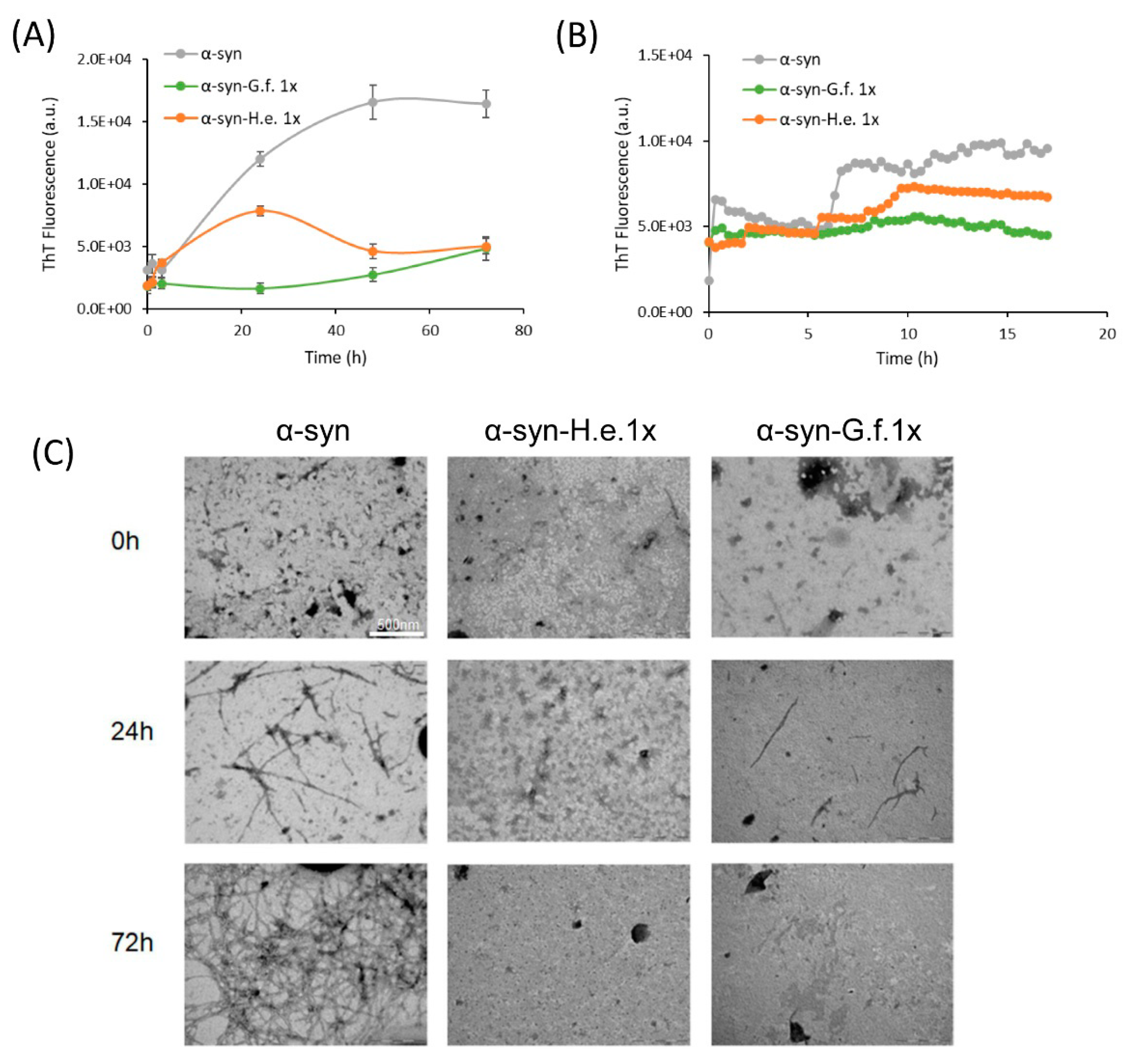
3.5. Fungal Extracts Inhibit α-Synuclein Aggregation In Vitro
3.6. G. frondosa Extract Extends Lifespan in a Drosophila melanogaster Model of Parkinson’s Disease
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ruetenik, A.; Barrientos, A. Exploiting Post-Mitotic Yeast Cultures to Model Neurodegeneration. Front. Mol. Neurosci. 2018, 11, 400. [Google Scholar] [CrossRef]
- Longo, V.D.; Shadel, G.S.; Kaeberlein, M.; Kennedy, B. Replicative and Chronological Aging in Saccharomyces Cerevisiae. Cell Metab. 2012, 16, 18–31. [Google Scholar] [CrossRef]
- Sampaio-Marques, B.; Pereira, H.; Santos, A.R.; Teixeira, A.; Ludovico, P. Caloric Restriction Rescues Yeast Cells from Alpha-Synuclein Toxicity through Autophagic Control of Proteostasis. Aging 2018, 10, 3821–3833. [Google Scholar] [CrossRef] [PubMed]
- Sampaio-Marques, B.; Guedes, A.; Vasilevskiy, I.; Gonçalves, S.; Outeiro, T.F.; Winderickx, J.; Burhans, W.C.; Ludovico, P. α-Synuclein Toxicity in Yeast and Human Cells Is Caused by Cell Cycle Re-Entry and Autophagy Degradation of Ribonucleotide Reductase 1. Aging Cell 2019, 18, e12922. [Google Scholar] [CrossRef]
- Sampaio-Marques, B.; Felgueiras, C.; Silva, A.; Rodrigues, M.; Tenreiro, S.; Franssens, V.; Reichert, A.S.; Outeiro, T.F.; Winderickx, J.; Ludovico, P. SNCA (α-Synuclein)-Induced Toxicity in Yeast Cells Is Dependent on Sir2-Mediated Mitophagy. Autophagy 2012, 8, 1494–1509. [Google Scholar] [CrossRef]
- Tenreiro, S.; Franssens, V.; Winderickx, J.; Outeiro, T.F. Yeast Models of Parkinson’s Disease-Associated Molecular Pathologies. Curr. Opin. Genet. Dev. 2017, 44, 74–83. [Google Scholar] [CrossRef]
- Dodel, R.; Csoti, I.; Ebersbach, G.; Fuchs, G.; Hahne, M.; Kuhn, W.; Oechsner, M.; Jost, W.; Reichmann, H.; Schulz, J.B. Lewy Body Dementia and Parkinson’s Disease with Dementia. J. Neurol. 2008, 255, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Van Den Eeden, S.K.; Tanner, C.M.; Bernstein, A.L.; Fross, R.D.; Leimpeter, A.; Bloch, D.A.; Nelson, L.M. Incidence of Parkinson’s Disease: Variation by Age, Gender, and Race/Ethnicity. Am. J. Epidemiol. 2003, 157, 1015–1022. [Google Scholar] [CrossRef]
- de Rijk, M.C.; Launer, L.J.; Berger, K.; Breteler, M.M.; Dartigues, J.F.; Baldereschi, M.; Fratiglioni, L.; Lobo, A.; Martinez-Lage, J.; Trenkwalder, C.; et al. Prevalence of Parkinson’s Disease in Europe: A Collaborative Study of Population-Based Cohorts. Neurologic Diseases in the Elderly Research Group. Neurology 2000, 54, S21–S23. [Google Scholar]
- Reiter, L.T.; Potocki, L.; Chien, S.; Gribskov, M.; Bier, E. A Systematic Analysis of Human Disease-Associated Gene Sequences in Drosophila Melanogaster. Genome Res. 2001, 11, 1114–1125. [Google Scholar] [CrossRef]
- Feany, M.B.; Bender, W.W. A Drosophila Model of Parkinson’s Disease. Nature 2000, 404, 394–398. [Google Scholar] [CrossRef]
- Sabaratnam, V.; Kah-Hui, W.; Naidu, M.; David, P. Neuronal Health—Can Culinary and Medicinal Mushrooms Help? J. Tradit. Complement. Med. 2013, 3, 62–68. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal Mushroom Science: Current Perspectives, Advances, Evidences, and Challenges. Biomed. J. 2014, 37, 345–356. [Google Scholar] [CrossRef]
- Venturella, G.; Ferraro, V.; Cirlincione, F.; Gargano, M.L. Medicinal Mushrooms: Bioactive Compounds, Use, and Clinical Trials. Int. J. Mol. Sci. 2021, 22, 634. [Google Scholar] [CrossRef]
- Wu, J.Y.; Siu, K.C.; Geng, P. Bioactive Ingredients and Medicinal Values of Grifola Frondosa (Maitake). Foods 2021, 10, 95. [Google Scholar] [CrossRef]
- Rossi, P.; Difrancia, R.; Quagliariello, V.; Savino, E.; Tralongo, P.; Randazzo, C.L.; Berretta, M. B-Glucans from Grifola Frondosa and Ganoderma Lucidum in Breast Cancer: An Example of Complementary and Integrative Medicine. Oncotarget 2018, 9, 24837–24856. [Google Scholar] [CrossRef]
- Li, I.C.; Lee, L.Y.; Tzeng, T.T.; Chen, W.P.; Chen, Y.P.; Shiao, Y.J.; Chen, C.C. Neurohealth Properties of Hericium Erinaceus Mycelia Enriched with Erinacines. Behav. Neurol. 2018, 2018, 5802634. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, Nutrition, and Health-Promoting Properties of Hericium Erinaceus (Lion’s Mane) Mushroom Fruiting Bodies and Mycelia and Their Bioactive Compounds. J. Agric. Food Chem. 2015, 63, 7108–7123. [Google Scholar] [CrossRef]
- Ratto, D.; Corana, F.; Mannucci, B.; Priori, E.C.; Cobelli, F.; Roda, E.; Ferrari, B.; Occhinegro, A.; Di Iorio, C.; De Luca, F.; et al. Hericium Erinaceus Improves Recognition Memory and Induces Hippocampal and Cerebellar Neurogenesis in Frail Mice during Aging. Nutrients 2019, 11, 715. [Google Scholar] [CrossRef]
- Vigna, L.; Morelli, F.; Agnelli, G.M.; Napolitano, F.; Ratto, D.; Occhinegro, A.; Di Iorio, C.; Savino, E.; Girometta, C.; Brandalise, F.; et al. Hericium Erinaceus Improves Mood and Sleep Disorders in Patients Affected by Overweight or Obesity: Could Circulating Pro-BDNF and BDNF Be Potential Biomarkers? Evid. Based. Complement. Alternat. Med. 2019, 2019, 7861297. [Google Scholar] [CrossRef]
- Corana, F.; Cesaroni, V.; Mannucci, B.; Baiguera, R.M.; Picco, A.M.; Savino, E.; Ratto, D.; Perini, C.; Kawagishi, H.; Girometta, C.E.; et al. Array of Metabolites in Italian Hericium Erinaceus Mycelium, Primordium, and Sporophore. Molecules 2019, 24, 3511. [Google Scholar] [CrossRef]
- Trovato Salinaro, A.; Pennisi, M.; Di Paola, R.; Scuto, M.; Crupi, R.; Cambria, M.T.; Ontario, M.L.; Tomasello, M.; Uva, M.; Maiolino, L.; et al. Neuroinflammation and Neurohormesis in the Pathogenesis of Alzheimer’s Disease and Alzheimer-Linked Pathologies: Modulation by Nutritional Mushrooms. Immun. Ageing 2018, 15, 8. [Google Scholar] [CrossRef]
- Lee, L.Y.; Chou, W.; Chen, W.P.; Wang, M.F.; Chen, Y.J.; Chen, C.C.; Tung, K.C. Erinacine A-Enriched Hericium Erinaceus Mycelium Delays Progression of Age-Related Cognitive Decline in Senescence Accelerated Mouse Prone 8 (SAMP8) Mice. Nutrients 2021, 13, 3659. [Google Scholar] [CrossRef]
- Song, L.; Zhou, Y.; Ni, S.; Wang, X.; Yuan, J.; Zhang, Y.; Zhang, S. Dietary Intake of β-Glucans Can Prolong Lifespan and Exert an Antioxidant Action on Aged Fish Nothobranchius Guentheri. Rejuvenation Res. 2020, 23, 293–301. [Google Scholar] [CrossRef]
- Aranaz, P.; Peña, A.; Vettorazzi, A.; Fabra, M.J.; Martínez-Abad, A.; López-Rubio, A.; Pera, J.; Parladé, J.; Castellari, M.; Milagro, F.I.; et al. Grifola Frondosa (Maitake) Extract Reduces Fat Accumulation and Improves Health Span in C. Elegans through the DAF-16/FOXO and SKN-1/NRF2 Signalling Pathways. Nutrients 2021, 13, 3968. [Google Scholar] [CrossRef]
- Tomas-hernandez, S.; Blanco, J.; Garcia-vallvé, S.; Pujadas, G.; Ojeda-montes, M.J.; Gimeno, A.; Arola, L.; Minghetti, L.; Beltrán-debón, R.; Mulero, M. Anti-Inflammatory and Immunomodulatory Effects of the Grifola Frondosa Natural Compound o-Orsellinaldehyde on LPS-Challenged Murine Primary Glial Cells. Roles of NF-Κβ and MAPK. Pharmaceutics 2021, 13, 806. [Google Scholar] [CrossRef]
- Cheng, J.H.; Tsai, C.L.; Lien, Y.Y.; Lee, M.S.; Sheu, S.C. High Molecular Weight of Polysaccharides from Hericium Erinaceus against Amyloid Beta-Induced Neurotoxicity. BMC Complement. Altern. Med. 2016, 16, 170. [Google Scholar] [CrossRef]
- Ciecierska, A.; Drywień, M.E.; Hamulka, J.; Sadkowski, T. Nutraceutical Functions of Beta-Glucans in Human Nutrition. Rocz. Panstw. Zakl. Hig. 2019, 70, 315–324. [Google Scholar] [CrossRef]
- La Rosa, F.; Clerici, M.; Ratto, D.; Occhinegro, A.; Licito, A.; Romeo, M.; Di Iorio, C.; Rossi, P. The Gut-Brain Axis in Alzheimer’s Disease and Omega-3. A Critical Overview of Clinical Trials. Nutrients 2018, 10, 1267. [Google Scholar] [CrossRef]
- Kalaras, M.D.; Richie, J.P.; Calcagnotto, A.; Beelman, R.B. Mushrooms: A Rich Source of the Antioxidants Ergothioneine and Glutathione. Food Chem. 2017, 233, 429–433. [Google Scholar] [CrossRef]
- Beelman, R.B.; Kalaras, M.D.; Phillips, A.T.; Richie, J.P. Is Ergothioneine a “longevity Vitamin” Limited in the American Diet? J. Nutr. Sci. 2020, 9, e52. [Google Scholar] [CrossRef] [PubMed]
- Roda, E.; Priori, E.C.; Ratto, D.; De Luca, F.; Di Iorio, C.; Angelone, P.; Locatelli, C.A.; Desiderio, A.; Goppa, L.; Savino, E.; et al. Neuroprotective Metabolites of Hericium Erinaceus Promote Neuro-Healthy Aging. Int. J. Mol. Sci. 2021, 22, 6379. [Google Scholar] [CrossRef] [PubMed]
- Roda, E.; Ratto, D.; De Luca, F.; Desiderio, A.; Ramieri, M.; Goppa, L.; Savino, E.; Bottone, M.G.; Locatelli, C.A.; Rossi, P. Searching for a Longevity Food, We Bump into Hericium Erinaceus Primordium Rich in Ergothioneine: The “Longevity Vitamin” Improves Locomotor Performances during Aging. Nutrients 2022, 14, 1177. [Google Scholar] [CrossRef] [PubMed]
- Gründemann, D.; Harlfinger, S.; Golz, S.; Geerts, A.; Lazar, A.; Berkels, R.; Jung, N.; Rubbert, A.; Schömig, E. Discovery of the Ergothioneine Transporter. Proc. Natl. Acad. Sci. USA 2005, 102, 5256–5261. [Google Scholar] [CrossRef]
- Mythri, R.B.; Harish, G.; Bharath, M.M. Therapeutic Potential of Natural Products in Parkinson’s Disease. Recent Pat. Endocr. Metab. Immune Drug Discov. 2012, 6, 181–200. [Google Scholar] [CrossRef]
- Cheah, I.K.; Halliwell, B. Ergothioneine, Recent Developments. Redox Biol. 2021, 42, 101868. [Google Scholar] [CrossRef] [PubMed]
- Kondoh, H.; Teruya, T.; Kameda, M.; Yanagida, M. Decline of Ergothioneine in Frailty and Cognition Impairment. FEBS Lett. 2022, 596, 1270–1278. [Google Scholar] [CrossRef]
- Li, Q.Z.; Wu, D.; Zhou, S.; Liu, Y.F.; Li, Z.P.; Feng, J.; Yang, Y. Structure Elucidation of a Bioactive Polysaccharide from Fruiting Bodies of Hericium Erinaceus in Different Maturation Stages. Carbohydr. Polym. 2016, 144, 196–204. [Google Scholar] [CrossRef]
- Pessina, S.; Tsiarentsyeva, V.; Busnelli, S.; Vanoni, M.; Alberghina, L.; Coccetti, P. Snf1/AMPK Promotes S-Phase Entrance by Controlling CLB5 Transcription in Budding Yeast. Cell Cycle 2010, 9, 2189–2200. [Google Scholar] [CrossRef]
- Tripodi, F.; Lombardi, L.; Guzzetti, L.; Panzeri, D.; Milanesi, R.; Leri, M.; Bucciantini, M.; Angeloni, C.; Beghelli, D.; Hrelia, S.; et al. Protective Effect of Vigna Unguiculata Extract against Aging and Neurodegeneration. Aging 2020, 12, 19785–19803. [Google Scholar] [CrossRef] [PubMed]
- Palazzi, L.; Fongaro, B.; Leri, M.; Acquasaliente, L.; Stefani, M.; Bucciantini, M.; Polverino de Laureto, P. Structural Features and Toxicity of α-Synuclein Oligomers Grown in the Presence of DOPAC. Int. J. Mol. Sci. 2021, 22, 6008. [Google Scholar] [CrossRef] [PubMed]
- LeVine, H. Quantification of Beta-Sheet Amyloid Fibril Structures with Thioflavin T. Methods Enzymol. 1999, 309, 274–284. [Google Scholar] [CrossRef] [PubMed]
- Piegholdt, S.; Rimbach, G.; Wagner, A.E. The Phytoestrogen Prunetin Affects Body Composition and Improves Fitness and Lifespan in Male Drosophila Melanogaster. FASEB J. 2016, 30, 948–958. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Nam, H.J.; Seo, M.; Han, S.K.; Choi, Y.; Nam, H.G.; Lee, S.J.; Kim, S. OASIS: Online Application for the Survival Analysis of Lifespan Assays Performed in Aging Research. PLoS ONE 2011, 6, e23525. [Google Scholar] [CrossRef] [PubMed]
- Zallocco, L.; Giusti, L.; Ronci, M.; Mussini, A.; Trerotola, M.; Mazzoni, M.R.; Lucacchini, A.; Sebastiani, L. Salivary Proteome Changes in Response to Acute Psychological Stress Due to an Oral Exam Simulation in University Students: Effect of an Olfactory Stimulus. Int. J. Mol. Sci. 2021, 22, 4295. [Google Scholar] [CrossRef]
- Girometta, C.; Dondi, D.; Baiguera, R.M.; Bracco, F.; Branciforti, D.S.; Buratti, S.; Lazzaroni, S.S.E. Characterization of Mycelia from Wood-Decay Species by TGA and IR Spectroscopy. Cellulose 2020, 27, 6133–6148. [Google Scholar] [CrossRef]
- Werner, K.; Pommer, L.; Broström, M. Thermal Decomposition of Hemicelluloses. J. Anal. Appl. Pyrolysis 2014, 110, 130–137. [Google Scholar] [CrossRef]
- Puanglek, S.; Kimura, S.; Enomoto-Rogers, Y.; Kabe, T.; Yoshida, M.; Wada, M.; Iwata, T. In Vitro Synthesis of Linear α-1,3-Glucan and Chemical Modification to Ester Derivatives Exhibiting Outstanding Thermal Properties. Sci. Reports 2016, 6, 30479. [Google Scholar] [CrossRef]
- Prime, R.B.; Bair, H.E.; Vyazovkin, S.; Gallagher, P.K.; Riga, A. Thermogravimetric Analysis (TGA). Therm. Anal. Polym. Fundam. Appl. 2008, 241–317. [Google Scholar] [CrossRef]
- Sun, B.; Hong, W.; Aziz, H.; Li, Y. Diketopyrrolopyrrole-Based Semiconducting Polymer Bearing Thermocleavable Side Chains. J. Mater. Chem. 2012, 22, 18950–18955. [Google Scholar] [CrossRef]
- Ospina Álvarez, S.P.; Ramírez Cadavid, D.A.; Escobar Sierra, D.M.; Ossa Orozco, C.P.; Rojas Vahos, D.F.; Zapata Ocampo, P.; Atehortúa, L. Comparison of Extraction Methods of Chitin from Ganoderma Lucidum Mushroom Obtained in Submerged Culture. Biomed Res. Int. 2014, 2014, 169071. [Google Scholar] [CrossRef] [PubMed]
- Dahmane, E.M.; Taourirte, M.; Eladlani, N.; Rhazi, M. Extraction and Characterization of Chitin and Chitosan from Parapenaeus Longirostris from Moroccan Local Sources. Int. J. Polym. Anal. Charact. 2014, 19, 342–351. [Google Scholar] [CrossRef]
- Gonzaga, M.L.C.; Menezes, T.M.F.; De Souza, J.R.R.; Ricardo, N.M.P.S.; Soares, S.D.A. Structural Characterization of β Glucans Isolated from Agaricus Blazei Murill Using NMR and FTIR Spectroscopy. Bioact. Carbohydrates Diet. Fibre 2013, 2, 152–156. [Google Scholar] [CrossRef]
- Przekora, A.; Benko, A.; Blazewicz, M.; Ginalska, G. Hybrid Chitosan/β-1,3-Glucan Matrix of Bone Scaffold Enhances Osteoblast Adhesion, Spreading and Proliferation via Promotion of Serum Protein Adsorption. Biomed. Mater. 2016, 11, 045001. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Ahmad, M.; Ashwar, B.A.; Gani, A.; Masoodi, F.A.; Wani, I.A.; Wani, S.M.; Gani, A. Effect of γ-Irradiation on Structure and Nutraceutical Potential of β-d-Glucan from Barley (Hordeum Vulgare). Int. J. Biol. Macromol. 2015, 72, 1168–1175. [Google Scholar] [CrossRef] [PubMed]
- Mau, J.L.; Lin, H.C.; Ma, J.T.; Song, S.F. Non-Volatile Taste Components of Several Speciality Mushrooms. Food Chem. 2001, 73, 461–466. [Google Scholar] [CrossRef]
- Sangtitanu, T.; Sangtanoo, P.; Srimongkol, P.; Saisavoey, T.; Reamtong, O.; Karnchanatat, A. Peptides Obtained from Edible Mushrooms: Hericium Erinaceus Offers the Ability to Scavenge Free Radicals and Induce Apoptosis in Lung Cancer Cells in Humans. Food Funct. 2020, 11, 4927–4939. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Razo, M.; González-Pérez, M. Analysis of Biomolecules of the Fungus Hericium Erinaceus Through the Theory of Electron Transfer of Quantum Chemistry and Its Relationship with the Primary Amino Acids. WORLD J. Pharm. Pharm. Sci. 2020, 9, 139–147. [Google Scholar] [CrossRef]
- Kim, S.P.; Kang, M.Y.; Kim, J.H.; Nam, S.H.; Friedman, M. Composition and Mechanism of Antitumor Effects of Hericium Erinaceus Mushroom Extracts in Tumor-Bearing Mice. J. Agric. Food Chem. 2011, 59, 9861–9869. [Google Scholar] [CrossRef]
- Tabata, T.; Yamasaki, Y.; Ogura, T. Comparison of Chemical Compositions of Maitake (Grifola Frondosa (Fr.) S. F. Gray) Cultivated on Logs and Sawdust Substrate. Food Sci. Technol. Res. 2004, 10, 21–24. [Google Scholar] [CrossRef][Green Version]
- Huang, S.J.; Tsai, S.Y.; Lin, S.Y.; Liang, C.H.; Mau, J.L. Nonvolatile Taste Components of Culinary-Medicinal Maitake Mushroom, Grifola Frondosa (Dicks.:Fr.) S.F. Gray. Int. J. Med. Mushrooms 2011, 13, 265–272. [Google Scholar] [CrossRef]
- Yang, F.; Wang, H.; Feng, G.; Zhang, S.; Wang, J.; Cui, L. Rapid Identification of Chemical Constituents in Hericium Erinaceus Based on LC-MS/MS Metabolomics. J. Food Qual. 2021, 2021, 233658894. [Google Scholar] [CrossRef]
- Zhang, Y.; Mills, G.L.; Nair, M.G. Cyclooxygenase Inhibitory and Antioxidant Compounds from the Mycelia of the Edible Mushroom Grifola Frondosa. J. Agric. Food Chem. 2002, 50, 7581–7585. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Zhang, J.; Han, Z.; Zhan, Y.; Wang, Y. Growth Parameters and Ergosterol Content of Mycelia and Fruit Bodies of Ten Hericium Erinaceus Strains Collected from the Wild in Heilongjiang Province,China. Acta Edulis Fungi 2014, 21, 45–49. [Google Scholar]
- Kim, S. Antioxidant Compounds for the Inhibition of Enzymatic Browning by Polyphenol Oxidases in the Fruiting Body Extract of the Edible Mushroom Hericium Erinaceus. Foods 2020, 9, 951. [Google Scholar] [CrossRef] [PubMed]
- Kawai, J.; Higuchi, Y.; Hirota, M.; Hirasawa, N.; Mori, K. Ergosterol and Its Derivatives from Grifola Frondosa Inhibit Antigen-Induced Degranulation of RBL-2H3 Cells by Suppressing the Aggregation of High Affinity IgE Receptors. Biosci. Biotechnol. Biochem. 2018, 82, 1803–1811. [Google Scholar] [CrossRef] [PubMed]
- Busti, S.; Coccetti, P.; Alberghina, L.; Vanoni, M. Glucose Signaling-Mediated Coordination of Cell Growth and Cell Cycle in Saccharomyces Cerevisiae. Sensors 2010, 10, 6195–6240. [Google Scholar] [CrossRef] [PubMed]
- Nicastro, R.; Tripodi, F.; Gaggini, M.; Castoldi, A.; Reghellin, V.; Nonnis, S.; Tedeschi, G.; Coccetti, P. Snf1 Phosphorylates Adenylate Cyclase and Negatively Regulates Protein Kinase A-Dependent Transcription in Saccharomyces Cerevisiae. J. Biol. Chem. 2015, 290, 24715–24726. [Google Scholar] [CrossRef]
- Belotti, F.; Tisi, R.; Paiardi, C.; Groppi, S.; Martegani, E. PKA-Dependent Regulation of Cdc25 RasGEF Localization in Budding Yeast. FEBS Lett. 2011, 585, 3914–3920. [Google Scholar] [CrossRef] [PubMed]
- Fruhmann, G.; Seynnaeve, D.; Zheng, J.; Ven, K.; Molenberghs, S.; Wilms, T.; Liu, B.; Winderickx, J.; Franssens, V. Yeast Buddies Helping to Unravel the Complexity of Neurodegenerative Disorders. Mech. Ageing Dev. 2017, 161, 288–305. [Google Scholar] [CrossRef] [PubMed]
- Lázaro, D.F.; Pavlou, M.A.S.; Outeiro, T.F. Cellular Models as Tools for the Study of the Role of Alpha-Synuclein in Parkinson’s Disease. Exp. Neurol. 2017, 298, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Outeiro, T.F.; Lindquist, S. Yeast Cells Provide Insight into Alpha-Synuclein Biology and Pathobiology. Science 2003, 302, 1772–1775. [Google Scholar] [CrossRef] [PubMed]
- Vida, T.A.; Emr, S.D. A New Vital Stain for Visualizing Vacuolar Membrane Dynamics and Endocytosis in Yeast. J. Cell Biol. 1995, 128, 779–792. [Google Scholar] [CrossRef]
- Yadav, D.; Negi, P.S. Bioactive Components of Mushrooms: Processing Effects and Health Benefits. Food Res. Int. 2021, 148, 110599. [Google Scholar] [CrossRef] [PubMed]
- Pan, H.Y.; Ye, Z.W.; Zheng, Q.W.; Yun, F.; Tu, M.Z.; Hong, W.G.; Chen, B.X.; Guo, L.Q.; Lin, J.F. Ergothioneine Exhibits Longevity-Extension Effect in Drosophila Melanogaster via Regulation of Cholinergic Neurotransmission, Tyrosine Metabolism, and Fatty Acid Oxidation. Food Funct. 2022, 13, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Cheah, I.K.; Ng, L.T.; Ng, L.F.; Lam, V.Y.; Gruber, J.; Huang, C.Y.W.; Goh, F.Q.; Lim, K.H.C.; Halliwell, B. Inhibition of Amyloid-Induced Toxicity by Ergothioneine in a Transgenic Caenorhabditis Elegans Model. FEBS Lett. 2019, 593, 2139–2150. [Google Scholar] [CrossRef] [PubMed]
- Mirisola, M.G.; Taormina, G.; Fabrizio, P.; Wei, M.; Hu, J.; Longo, V.D. Serine- and Threonine/Valine-Dependent Activation of PDK and Tor Orthologs Converge on Sch9 to Promote Aging. PLoS Genet. 2014, 10, e1004113. [Google Scholar] [CrossRef] [PubMed]
- Mirzaei, H.; Suarez, J.A.; Longo, V.D. Protein and Amino Acid Restriction, Aging and Disease: From Yeast to Humans. Trends Endocrinol. Metab. 2014, 25, 558–566. [Google Scholar] [CrossRef]
- Wu, Z.; Song, L.; Liu, S.Q.; Huang, D. Independent and Additive Effects of Glutamic Acid and Methionine on Yeast Longevity. PLoS ONE 2013, 8, e79319. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte | H. erinaceus | Reference (mg/g) Fruiting Body | G. frondosa | Reference (mg/g) Fruiting Body |
|---|---|---|---|---|
| Lactic acid | D | - | D | - |
| L-Alanine | D | 2.43 [56] 0.11 [57] D [58] D [59] | D | 2.15 [60] 3.13 [60] 5.22 [61] 2.77 [56] |
| Sarcosine | D | - | D | - |
| Glycine | D | 1.03 [56] 0.11 [57] D [58] | D | 1.53 [60] 1.53 [60] 2.46 [61] 0.57 [56] |
| L-Valine | D | 0.30 [56] 0.10 [57] D [58] | D | - |
| L-Leucine | D | 2.38 [56] 0.14 [57] D [58] | D | 0.05 [60] 0.,09 [60] 0.27 [61] 0.35 [56] |
| Isoleucine | D | 0.07 [57] D [58] | D | 0.12 [60] 0.12 [60] 0.56 [61] 0.33 [56] |
| L-Proline | D | 0.10 [57] D [58] | D | - |
| D-Pyroglutamic acid | D | - | D | - |
| L-Methionine | D | 1.08 [56] 0.03 [57] D [58] | D | 4.5 [61] 1.4 [56] |
| Adipic acid | - | - | D | - |
| L-Serine | D | 0.35 [56] 0.11 [57] D [58] | D | 2.91 [60] 2.82 [60] 2.01 [61] 0.97 [56] |
| L-Lysine | D | 0.47 [56] 0.28 [57] D [58] D [59] | D | 1.56 [60] 1.28 [60] 5.70 [61] 1.11 [56] |
| L-Threonine | D | 0.78 [56] 0.12 [57] D [58] | D | 1.43 [60] 1.44 [60] 4.4 [56] |
| L-Phenylalanine | D | 0.20 [56] 0.05 [57] D [58] | D | 0.26 [60] 0.28 [60] 2.71 [61] 0.8 [56] |
| Aspartic acid | D | 0.50 [56] 0.23 [57] D [58] | D | 1.61 [60] 1.25 [60] 1.66 [61] 0.64 [56] |
| Isovanillic acid | - | - | D | 1.61 [60] 1.25 [60] 1.88 [61] 0.42 [56] |
| L-Glutamic acid | D | 0.50 [56] 0.42 [57] D [58] D [59] | D | 8.01 [60] 9.10 [60] 12.62 [61] 0.67 [56] |
| Citric acid | D | D [62] D [59] | D | - |
| L-Tyrosine | D | 0.14 [57] D [58] D [62] | D | - |
| Crotonic acid | D | - | D | - |
| Glyceric acid | D | - | D | - |
| Palmitic acid | D | - | D | D [63] |
| Azelaic acid | - | - | D | - |
| Linoelaidic acid | D | - | D | D [63] |
| Oleic acid | D | D [59] | D | D [63] |
| Stearic acid | - | - | D | - |
| Ergosterol | D | 2.58 [64] 2.72 [64] 2.45 [64] D [65] | D | D [66] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tripodi, F.; Falletta, E.; Leri, M.; Angeloni, C.; Beghelli, D.; Giusti, L.; Milanesi, R.; Sampaio-Marques, B.; Ludovico, P.; Goppa, L.; et al. Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts. Nutrients 2022, 14, 4368. https://doi.org/10.3390/nu14204368
Tripodi F, Falletta E, Leri M, Angeloni C, Beghelli D, Giusti L, Milanesi R, Sampaio-Marques B, Ludovico P, Goppa L, et al. Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts. Nutrients. 2022; 14(20):4368. https://doi.org/10.3390/nu14204368
Chicago/Turabian StyleTripodi, Farida, Ermelinda Falletta, Manuela Leri, Cristina Angeloni, Daniela Beghelli, Laura Giusti, Riccardo Milanesi, Belém Sampaio-Marques, Paula Ludovico, Lorenzo Goppa, and et al. 2022. "Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts" Nutrients 14, no. 20: 4368. https://doi.org/10.3390/nu14204368
APA StyleTripodi, F., Falletta, E., Leri, M., Angeloni, C., Beghelli, D., Giusti, L., Milanesi, R., Sampaio-Marques, B., Ludovico, P., Goppa, L., Rossi, P., Savino, E., Bucciantini, M., & Coccetti, P. (2022). Anti-Aging and Neuroprotective Properties of Grifola frondosa and Hericium erinaceus Extracts. Nutrients, 14(20), 4368. https://doi.org/10.3390/nu14204368