
Vitamin K Contribution to DNA Damage—Advantage or Disadvantage? A Human Health Response



Abstract
1. Introduction
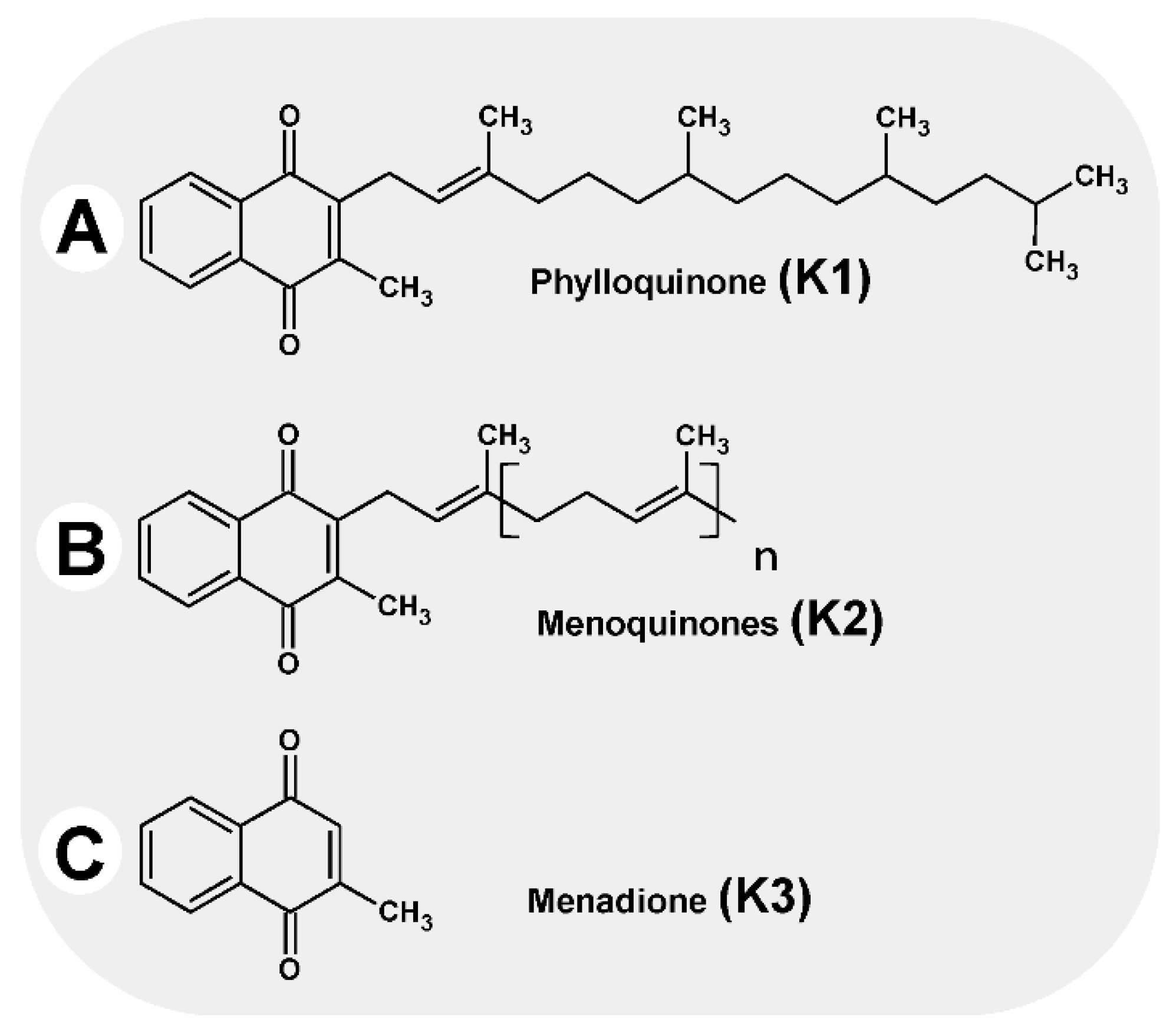
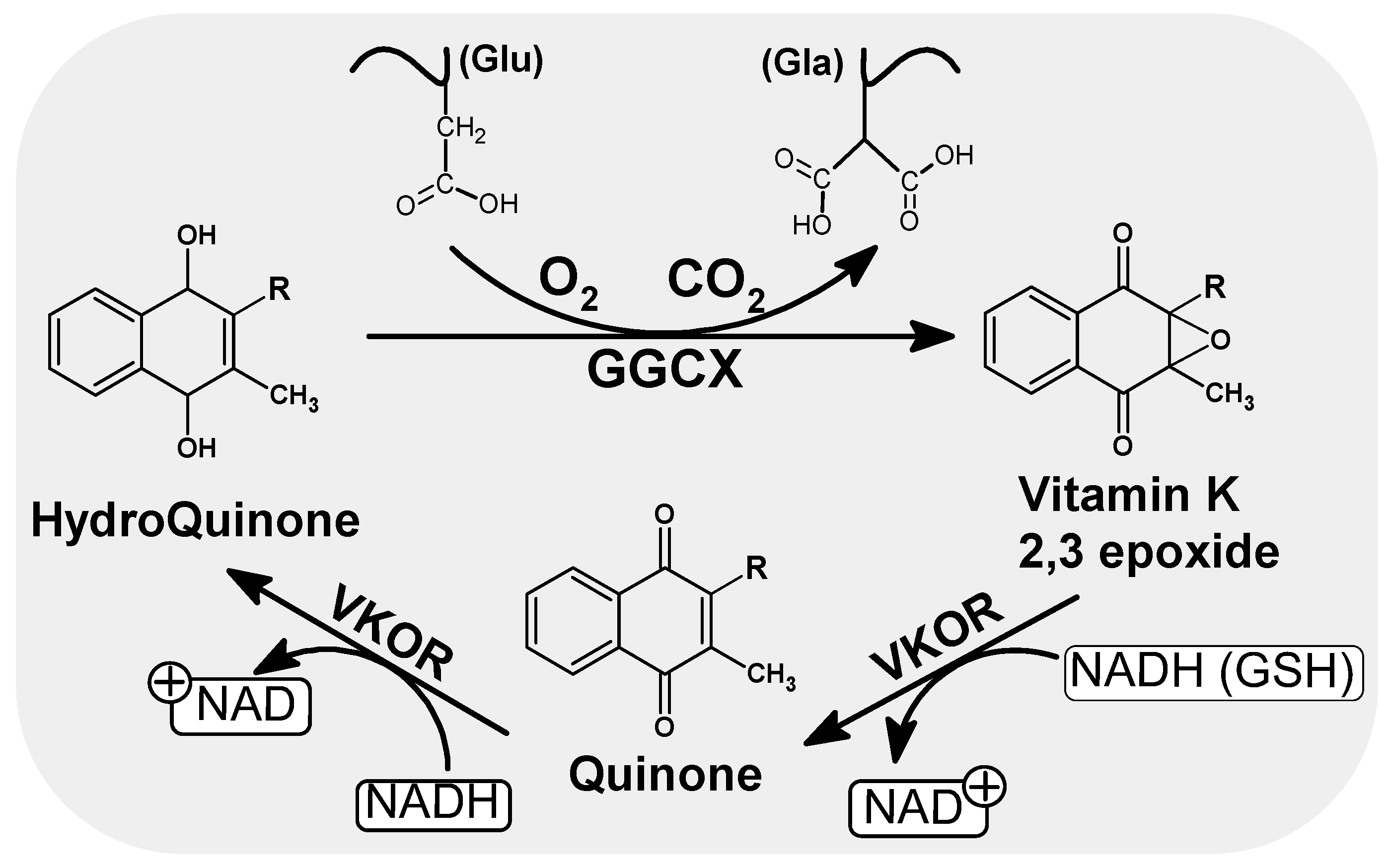
2. Vitamin K in Cell Biology
3. Recommended Daily Intake (RDI) and Deficiency of Vitamin K

{kind=link}
{kind=link}
{kind=link}
| RDI (μg/day) | References | |
|---|---|---|
| IOM 1 (2001) | 90 μg/day women 120 μg/day men | [53] |
| WHO/FAO 2 (2004) | 55 μg/day women 65 μg/day men | [54] |
| D-A-CH 3 (2015) | 60 μg/day women 70 μg/day men | [54] |
| EFSA 4 Panel (2017) | 70 μg/day for all adults | [54] |
| NIZP-PZH 5 (2020) | 55 μg/day women 65 μg/day men | [55] |
4. Anti-Cancer Properties
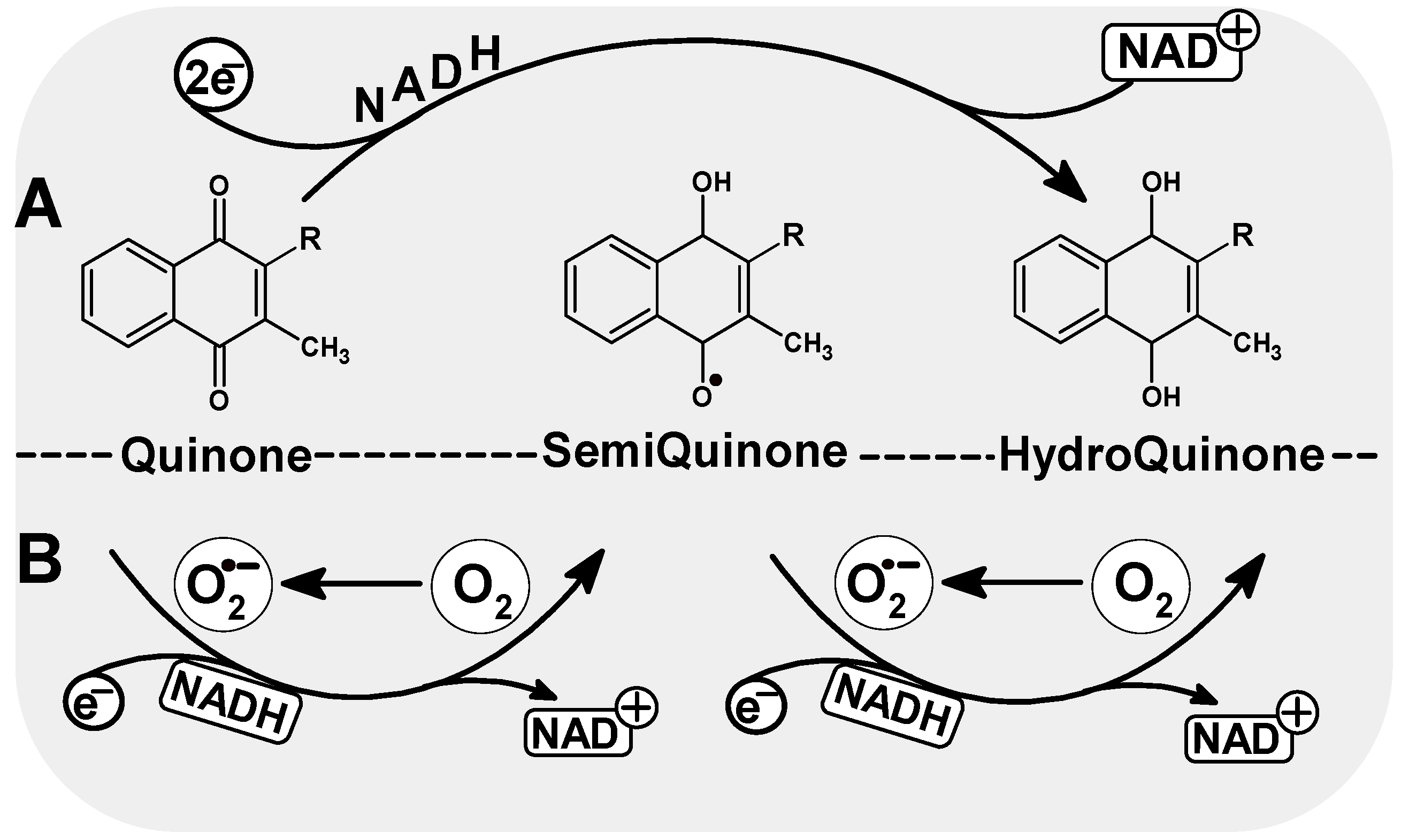
4.1. DNA Damage
4.2. Anti-cancer Effect
5. Summary
Author Contributions
Funding
Conflicts of Interest
References
- National Center for Biotechnology Information. PubChem Compound Summary for CID 5280483, Vitamin, K. 2022. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Vitamin-K. (accessed on 5 January 2022).
- Schurgers, L.; Geleijnse, J.M.; Grobbee, D.E.; Pols, H.A.P.; Hofman, A.; Witteman, J.C.M.; Vermeer, C. Nutritional Intake of Vitamins K1 (Phylloquinone) and K2 (Menaquinone) in The Netherlands. J. Nutr. Environ. Med. 1999, 9, 115–122. [Google Scholar] [CrossRef]
- Holmes, M.V.; Hunt, B.J.; Shearer, M.J. The role of dietary vitamin K in the management of oral vitamin K antagonists. Blood Rev. 2012, 26, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kegel, L.P.; Crane, F.L. Vitamin K1 in Chloroplasts. Nature 1962, 194, 1282. [Google Scholar] [CrossRef]
- Basset, G.J.; Latimer, S.; Fatihi, A.; Soubeyrand, E.; Block, A. Phylloquinone (Vitamin K1): Occurrence, Biosynthesis and Functions. Mini-Rev. Med. Chem. 2017, 17, 1028–1038. [Google Scholar] [CrossRef]
- Booth, S.L. Roles for Vitamin K Beyond Coagulation. Annu. Rev. Nutr. 2009, 29, 89–110. [Google Scholar] [CrossRef]
- Booth, S.L. Vitamin K: Food composition and dietary intakes. Food Nutr. Res. 2012, 56, 1–5. [Google Scholar] [CrossRef]
- Newman, P.; Shearer, M.J. Metabolism and cell biology of vitamin K. Thromb. Haemost. 2008, 100, 530–547. [Google Scholar] [CrossRef]
- Shearer, M.J. Vitamin K. Lancet 1995, 345, 229–234. [Google Scholar] [CrossRef]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K status in human tissues: Tissue-specific accumulation ofphylloquinone and menaquinone-4. Br. J. Nutr. 1996, 75, 121–127. [Google Scholar] [CrossRef]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Vermeer, C.; Braam, L. Role of K vitamins in the regulation of tissue calcification. J. Bone Miner. Metab. 2001, 19, 201–206. [Google Scholar] [CrossRef]
- Wallin, R.; Sane, D.C.; Hutson, S.M. Vitamin K 2,3-epoxide reductase and the vitamin K-dependent g-carboxylation system. Thromb Res. 2003, 108, 221–226. [Google Scholar] [CrossRef]
- Morrissey, J.H. Deeper understanding of carboxylase. Blood 2016, 127, 1841–1842. [Google Scholar] [CrossRef]
- Vermeer, C.; Shearer, M.J.; Zittermann, A. Beyond Deficiency: Potential benefits of increased intakes of vitamin K for bone and vascular health. Eur. J. Nutr. 2004, 43, 325–335. [Google Scholar] [CrossRef]
- Tie, J.-K.; Stafford, D. Functional Study of the Vitamin K Cycle Enzymes in Live Cells. Methods Enzymol. 2017, 584, 349–394. [Google Scholar] [CrossRef]
- Coutu, D.; Wu, J.H.; Monette, A.; Rivard, G.; Blostein, M.D.; Galipeau, J. Periostin, a Member of a Novel Family of Vitamin K-dependent Proteins, Is Expressed by Mesenchymal Stromal Cells. J. Biol. Chem. 2008, 283, 17991–18001. [Google Scholar] [CrossRef]
- McCann, J.C.; Ames, B.N. Vitamin K, an example of triage theory: Is micronutrient inadequacy linked to diseases of aging? Am. J. Clin. Nutr. 2009, 90, 889–907. [Google Scholar] [CrossRef]
- Lai, Y.; Masatoshi, H.; Ma, Y.; Guo, Y.; Zhang, B. Role of Vitamin K in Intestinal Health. Front. Immunol. 2022, 12, 791565. [Google Scholar] [CrossRef]
- Weijs, B.; Blaauw, Y.; Rennenberg, R.J.; Schurgers, L.; Timmermans, C.C.M.M.; Pison, L.; Nieuwlaat, R.; Hofstra, L.; Kroon, A.A.; Wildberger, J.; et al. Patients using vitamin K antagonists show increased levels of coronary calcification: An observational study in low-risk atrial fibrillation patients. Eur. Heart, J. 2011, 32, 2555–2562. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Joosen, I.A.; Laufer, E.M.; Chatrou, M.L.L.; Herfs, M.; Winkens, M.H.M.; Westenfeld, R.; Veulemans, V.; Krueger, T.; Shanahan, C.; et al. Vitamin K-Antagonists Accelerate Atherosclerotic Calcification and Induce a Vulnerable Plaque Phenotype. PLoS ONE 2012, 7, e43229. [Google Scholar] [CrossRef]
- Brandenburg, V.M.; Reinartz, S.; Kaesler, N.; Krüger, T.; Dirrichs, T.; Kramann, R. Slower progress of aortic valve calcification with Vitamin K supplementation: Results from a prospective interventional proof-of-concept study. Circulation 2017, 135, 2081–2083. [Google Scholar] [CrossRef]
- Aksoy, A.; Al Zaidi, M.; Repges, E.; Becher, M.U.; Müller, C.; Oldenburg, J.; Zimmer, S.; Nickenig, G.; Tiyerili, V. Vitamin K Epoxide Reductase Complex Subunit 1-Like 1 (VKORC1L1) Inhibition Induces a Proliferative and Pro-inflammatory Vascular Smooth Muscle Cell Phenotype. Front. Cardiovasc. Med. 2021, 8, 708946. [Google Scholar] [CrossRef] [PubMed]
- Kieronska-Rudek, A.; Kij, A.; Kaczara, P.; Tworzydlo, A.; Napiorkowski, M.; Sidoryk, K.; Chlopicki, S. Exogenous Vitamins K Exert Anti-Inflammatory Effects Dissociated from Their Role as Substrates for Synthesis of Endogenous MK-4 in Murine Macrophages Cell Line. Cells 2021, 10, 1571. [Google Scholar] [CrossRef]
- Li, J.; Wang, H.; Rosenberg, P.A. Vitamin K prevents oxidative cell death by inhibiting activation of 12-lipoxygenase in developing oligodendrocytes. J. Neurosci. Res. 2009, 87, 1997–2005. [Google Scholar] [CrossRef]
- Li, J.; Lin, J.C.; Wang, H.; Peterson, J.W.; Furie, B.C.; Furie, B.; Booth, S.L.; Volpe, J.J.; Rosenberg, P. Novel Role of Vitamin K in Preventing Oxidative Injury to Developing Oligodendrocytes and Neurons. J. Neurosci. 2003, 23, 5816–5826. [Google Scholar] [CrossRef]
- Vervoort, L.M.; Ronden, J.E.; Thijssen, H.H. The potent antioxidant activity of the vitamin K cycle in microsomal lipid peroxidation. Biochem. Pharmacol. 1997, 54, 871–876. [Google Scholar] [CrossRef]
- Carrié, I.; Portoukalian, J.; Vicaretti, R.; Rochford, J.; Potvin, S.; Ferland, G. Menaquinone-4 Concentration Is Correlated with Sphingolipid Concentrations in Rat Brain. J. Nutr. 2004, 134, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Alisi, L.; Cao, R.; De Angelis, C.; Cafolla, A.; Caramia, F.; Cartocci, G.; Librando, A.; Fiorelli, M. The Relationships Between Vitamin K and Cognition: A Review of Current Evidence. Front. Neurol. 2019, 10, 239. [Google Scholar] [CrossRef]
- Ferland, G. Vitamin K, an emerging nutrient in brain function. BioFactors 2012, 38, 151–157. [Google Scholar] [CrossRef]
- Chatrou, M.L.; Winckers, K.; Hackeng, T.M.; Reutelingsperger, C.P.; Schurgers, L.J. Vascular calcification: The price to pay for anticoagulation therapy with vitamin K-antagonists. Blood Rev. 2012, 26, 155–166. [Google Scholar] [CrossRef] [PubMed]
- Namba, S.; Yamaoka-Tojo, M.; Hashikata, T.; Ikeda, Y.; Kitasato, L.; Hashimoto, T.; Shimohama, T.; Tojo, T.; Takahira, N.; Masuda, T.; et al. Long-term warfarin therapy and biomarkers for osteoporosis and atherosclerosis. BBA Clin. 2015, 4, 76–80. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Shirakawa, H.; Hiwatashi, K.; Furukawa, Y.; Mizutani, T.; Komai, M. Vitamin K Suppresses Lipopolysaccharide-Induced Inflammation in the Rat. Biosci. Biotechnol. Biochem. 2006, 70, 926–932. [Google Scholar] [CrossRef]
- Wang, J.; Li, J. Activated protein C: A potential cardioprotective factor against ischemic injury during ischemia/reperfusion. Am. J. Transl. Res. 2009, 1, 381–392. [Google Scholar]
- Joyce, D.E.; Grinnell, B.W. Recombinant human activated protein C attenuates the inflammatory response in endothelium and monocytes by modulating nuclear factor-kappaB. Crit. Care Med. 2002, 30, 288–293. [Google Scholar] [CrossRef]
- Griffin, J.; Fernandez, J.; Liu, D.; Cheng, T.; Guo, H.; Zlokovic, B.V. Activated protein C and ischemic stroke. Crit. Care Med. 2004, 32, S247–S253. [Google Scholar] [CrossRef]
- Tousoulis, D.; Oikonomou, E.; Economou, E.K.; Crea, F.; Kaski, J.C. Inflammatory cytokines in atherosclerosis: Current therapeutic approaches. Eur. Heart J. 2016, 37, 1723–1732. [Google Scholar] [CrossRef]
- Senn, J.J.; Klover, P.J.; Nowak, I.A.; Mooney, R.A. Interleukin-6 Induces Cellular Insulin Resistance in Hepatocytes. Diabetes 2002, 51, 3391–3399. [Google Scholar] [CrossRef]
- Samson, L.D.; Buisman, A.M.; Ferreira, J. Inflammatory Marker Trajectories Associated with Frailty and Ageing in A 20-yearTutinal Study. Clin. Transl. Immunol. 2022, 11, E1374. [Google Scholar] [CrossRef]
- Hubbard, R.E.; O’Mahony, M.S.; Calver, B.L.; Woodhouse, K.W. Nutrition, Inflammation, and Leptin Levels in Aging and Frailty. J. Am. Geriatr. Soc. 2008, 56, 279–284. [Google Scholar] [CrossRef]
- Brábek, J.; Jakubek, M.; Vellieux, F.; Novotný, J.; Kolář, M.; Lacina, L.; Szabo, P.; Strnadová, K.; Rösel, D.; Dvořánková, B.; et al. Interleukin-6: Molecule in the Intersection of Cancer, Ageing and COVID-19. Int. J. Mol. Sci. 2020, 21, 7937. [Google Scholar] [CrossRef]
- Omokehinde, T.; Johnson, R.W. GP130 Cytokines in Breast Cancer and Bone. Cancers 2020, 12, 326. [Google Scholar] [CrossRef] [PubMed]
- Kodama, H.; Fukuda, K.; Pan, J.; Sano, M.; Takahashi, T.; Kato, T.; Makino, S.; Manabe, T.; Murata, M.; Ogawa, S. Significance of ERK cascade compared with JAK/STAT and PI3-K pathway in gp130-mediated cardiac hypertrophy. Am. J. Physiol. Heart Circ. Physiol. 2000, 279, H1635–H1644. [Google Scholar] [CrossRef] [PubMed]
- Felcher, C.M.; Bogni, E.S.; Kordon, E.C. IL-6 Cytokine Family: A Putative Target for Breast Cancer Prevention and Treatment. Int. J. Mol. Sci. 2022, 23, 1809. [Google Scholar] [CrossRef] [PubMed]
- Reddi, K.; Henderson, B.; Meghji, S.; Wilson, M.; Poole, S.; Hopper, C.; Harris, M.; Hodges, S. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by Naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef]
- Thijssen, H.H.W.; Vervoort, L.M.T.; Schurgers, L.; Shearer, M.J. Menadione is a metabolite of oral vitamin K. Br. J. Nutr. 2006, 95, 260–266. [Google Scholar] [CrossRef]
- Hirota, Y.; Tsugawa, N.; Nakagawa, K.; Suhara, Y.; Tanaka, K.; Uchino, Y.; Takeuchi, A.; Sawada, N.; Kamao, M.; Wada, A.; et al. Menadione (Vitamin K3) Is a Catabolic Product of Oral Phylloquinone (Vitamin K1) in the Intestine and a Circulating Precursor of Tissue Menaquinone-4 (Vitamin K2) in Rats. J. Biol. Chem. 2013, 288, 33071–33080. [Google Scholar] [CrossRef]
- Okano, T.; Shimomura, Y.; Yamane, M.; Suhara, Y.; Kamao, M.; Sugiura, M.; Nakagawa, K. Conversion of Phylloquinone (Vitamin K1) into Menaquinone-4 (Vitamin K2) in Mice. J. Biol. Chem. 2008, 283, 11270–11279. [Google Scholar] [CrossRef]
- Ronden, J.E.; Thijssen, H.W.H.; Vermeer, C. Tissue distribution of K-vitamers under different nutritional regimens in the rat. Biochim. Biophys. Acta 1998, 1379, 16–22. [Google Scholar] [CrossRef]
- McKeown, N.M.; Jacques, P.F.; Gundberg, C.M.; Peterson, J.W.; Tucker, K.; Kiel, D.; Wilson, P.W.F.; Booth, S.L. Dietary and Nondietary Determinants of Vitamin K Biochemical Measures in Men and Women. J. Nutr. 2002, 132, 1329–1334. [Google Scholar] [CrossRef]
- Booth, S.L.; Martini, L.; Peterson, J.W.; Saltzman, E.; Dallal, G.E.; Wood, R.J. Dietary Phylloquinone Depletion and Repletion in Older Women. J. Nutr. 2003, 133, 2565–2569. [Google Scholar] [CrossRef]
- Booth, S.L.; Lichtenstein, A.; O’Brien-Morse, M.; McKeown, N.M.; Wood, R.J.; Saltzman, E.; Gundberg, C.M. Effects of a Hydrogenated Form of Vitamin K On Bone Formation and Resorption. Am. J. Clin. Nutr. 2001, 74, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Food and Nutrition Board, Insititue of Medicne. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academy Press: Washington, DC, USA, 2002.
- EFSA NDA Panel (EFSA Panel on Dietetic Products, Nutrition and Allergies); Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; et al. Scientific Opinion on the dietary reference values for vitamin K. EFSA J. 2017, 15, 4780. [Google Scholar] [CrossRef]
- Jarosz, M.; Rychlik, E.; Stoś, K.; Charzewska, J. Normy żywienia dla Populacji Polski i Ich Zastosowanie; NIZP-PZH: Warszawa, Poland, 2020; ISBN 978-83-65870-28-5. [Google Scholar]
- Simes, D.C.; Viegas, C.S.B.; Araújo, N.; Marreiros, C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients 2020, 12, 138. [Google Scholar] [CrossRef]
- Chen, L.-J.; Hsiao, F.-Y.; Shen, L.-J.; Wu, F.-L.L.; Tsay, W.; Hung, C.-C.; Lin, S.-W. Use of Hypoprothrombinemia-Inducing Cephalosporins and the Risk of Hemorrhagic Events: A Nationwide Nested Case-Control Study. PLoS ONE 2016, 11, e0158407. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.L. Vitamin K status In the elderly. Curr. Opin. Clin. Nutr. Metab. Care 2007, 10, 20–29. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Vermeer, C. Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2002, 1570, 27–32. [Google Scholar] [CrossRef]
- Kamao, M.; Suhara, Y.; Tsugawa, N.; Uwano, M.; Yamaguchi, N.; Uenishi, K.; Ishida, H.; Sasaki, S.; Okano, T. Vitamin K Content of Foods and Dietary Vitamin K Intake in Japanese Young Women. J. Nutr. Sci. Vitaminol. 2007, 53, 464–470. [Google Scholar] [CrossRef]
- Badr, M.; Yoshihara, H.; Kauffman, F.; Thurman, R. Menadione causes selective toxicity to periportal regions of the liver lobule. Toxicol. Lett. 1987, 35, 241–246. [Google Scholar] [CrossRef]
- Qadri, S.M.; Eberhard, M.; Mahmud, H.; Föller, M.; Lang, F. Stimulation of ceramide formation and suicidal erythrocyte death by vitamin K3 (menadione). Eur. J. Pharmacol. 2009, 623, 10–13. [Google Scholar] [CrossRef]
- Popa, D.-S.; Bigman, G.; Rusu, M. The Role of Vitamin K in Humans: Implication in Aging and Age-Associated Diseases. Antioxidants 2021, 10, 566. [Google Scholar] [CrossRef]
- Morrison, H.; Jernström, B.; Nordenskjöld, M.; Thor, H.; Orrenius, S. Induction of dna damage by menadione (2-methyl-1,4-naphthoquinone) in primary cultures of rat hepatocytes. Biochem. Pharmacol. 1984, 33, 1763–1769. [Google Scholar] [CrossRef]
- Asmus, K.D.; Bonifačić, M. Part I, Chapter 1—Free radical chemistry. In Handbook of Oxidants and Antioxidants in Exercise; Chandan, K.S., Packer, L., Hänninen, O.O.P., Eds.; Elsevier Science B.V.: Amsterdam, The Netherlands, 2000; pp. 3–54. [Google Scholar] [CrossRef]
- Ngo, E.O.; Sun, T.-P.; Chang, J.-Y.; Wang, C.-C.; Chi, K.-H.; Cheng, A.-L.; Nutter, L.M. Menadione-induced DNA damage in a human tumor cell line. Biochem. Pharmacol. 1991, 42, 1961–1968. [Google Scholar] [CrossRef]
- Morgan, W.; Hartley, J.A.; Cohen, G.M. Quinone-induced DNA single strand breaks in rat hepatocytes and human chronic myelogenous leukaemic K562 cells. Biochem. Pharmacol. 1992, 44, 215–221. [Google Scholar] [CrossRef]
- Morgan, W.A. DNA single-strand breakage in mammalian cells induced by redox cycling quinones in the absence of oxidative stress. J. Biochem. Toxicol. 1995, 10, 227–232. [Google Scholar] [CrossRef]
- Loor, G.; Kondapalli, J.; Schriewer, J.M.; Chandel, N.S.; Vanden Hoek, T.L.; Schumacker, P.T. Menadione triggers cell death through ROS-dependent mechanisms involving PARP activation without requiring apoptosis. Free Radic. Biol. Med. 2010, 49, 1925–1936. [Google Scholar] [CrossRef]
- Boetefuer, E.L.; Lake, R.J.; Dreval, K.; Fan, H.-Y. Poly(ADP-ribose) polymerase 1 (PARP1) promotes oxidative stress–induced association of Cockayne syndrome group B protein with chromatin. J. Biol. Chem. 2018, 293, 17863–17874. [Google Scholar] [CrossRef] [PubMed]
- Sinha, B.; Chignell, C. Binding mode of chemically activated semiquinone free radicals from quinone anticancer agents to DNA. Chem. Interact. 1979, 28, 301–308. [Google Scholar] [CrossRef]
- Giulivi, C.; Cadenas, E. One- and two-electron reduction of 2-methyl-1,4-naphthoquinone bioreductive alkylating agents: Kinetic studies, free-radical production, thiol oxidation and DNA-strand-break formation. Biochem. J. 1994, 301, 21–30. [Google Scholar] [CrossRef]
- Verrax, J.; Cadrobbi, J.; Marques, C.; Taper, H.; Habraken, Y.; Piette, J.; Calderon, P.B. Ascorbate potentiates the cytotoxicity of menadione leading to an oxidative stress that kills cancer cells by a non-apoptotic caspase-3 independent form of cell death. Apoptosis 2004, 9, 223–233. [Google Scholar] [CrossRef]
- Aherne, S.A.; O’Brien, N.M. Mechanism of protection by the flavonoids, quercetin and rutin, against tert-butylhydroperoxide- and menadione-induced DNA single strand breaks in Caco-2 cells. Free Radic. Biol. Med. 2000, 29, 507–514. [Google Scholar] [CrossRef]
- Woods, J.A.; Young, A.J.; Gilmore, I.T.; Morris, A.; Bilton, R.F. Measurement of Menadione-Mediated DNA Damage in Human Lymphocytes Using the Comet Assay. Free Radic. Res. 1997, 26, 113–124. [Google Scholar] [CrossRef]
- D’Odorico, A.; Sturniolo, G.C.; Bilton, R.F.; Morris, A.I.; Gilmore, I.T.; Naccarato, R. Quinone-induced DNA single strand breaks in a human colon carcinoma cell line. Carcinogenesis 1997, 18, 43–46. [Google Scholar] [CrossRef] [PubMed]
- Klaus, V.; Hartmann, T.; Gambini, J.; Graf, P.; Stahl, W.; Hartwig, A.; Klotz, L.-O. 1,4-Naphthoquinones as inducers of oxidative damage and stress signaling in HaCaT human keratinocytes. Arch. Biochem. Biophys. 2010, 496, 93–100. [Google Scholar] [CrossRef]
- Halilovic, A.; Schmedt, T.; Benischke, A.-S.; Hamill, C.; Chen, Y.; Santos, J.H.; Jurkunas, U.V. Menadione-Induced DNA Damage Leads to Mitochondrial Dysfunction and Fragmentation During Rosette Formation in Fuchs Endothelial Corneal Dystrophy. Antioxid. Redox Signal. 2016, 24, 1072–1083. [Google Scholar] [CrossRef]
- Gerasimenko, J.; Gerasimenko, O.; Palejwala, A.; Tepikin, A.; Petersen, O.; Watson, A.J.M. Menadione-induced apoptosis: Roles of cytosolic Ca2+ elevations and the mitochondrial permeability transition pore. J. Cell Sci. 2002, 115, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, B.; Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Calcium and mitochondria in the regulation of cell death. Biochem. Biophys. Res. Commun. 2015, 460, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Cojocel, C.; Novotny, L.; Vachalkova, A. Mutagenic and carcinogenic potential of menadione. Neoplasma 2006, 53, 316. [Google Scholar]
- Criddle, N.; Gillies, S.; Baumgartner-Wilson, K.; Jaffar, M.; Chinje, E.C.; Passmore, S.; Petersen, H. Menadione-induced Reactive Oxygen Species Generation via Redox Cycling Promotes Apoptosis of Murine Pancreatic Acinar Cells. J. Biol. Chem. 2006, 281, 40485–40492. [Google Scholar] [CrossRef]
- Shibayama-Imazu, T.; Sonoda, I.; Sakairi, S.; Aiuchi, T.; Ann, W.-W.; Nakajo, S.; Itabe, H.; Nakaya, K. Production of superoxide and dissipation of mitochondrial transmembrane potential by vitamin K2 trigger apoptosis in human ovarian cancer TYK-nu cells. Apoptosis 2006, 11, 1535–1543. [Google Scholar] [CrossRef]
- Li, Z.; Yao, Y.; Zhang, Y.; Zhang, Y.; Shao, Y.; Tang, C.; Qu, W.; Zhou, Y. Classification and Temporal Variability in Urinary 8-oxodG and 8-oxoGuo: Analysis by UHPLC-MS/MS. Sci. Rep. 2019, 9, 8187. [Google Scholar] [CrossRef]
- Topp, H.; Fusch, G.; Schöch, G.; Fusch, C. Noninvasive Markers of Oxidative DNA Stress, RNA Degradation and Protein Degradation Are Differentially Correlated With Resting Metabolic Rate and Energy Intake in Children and Adolescents. Pediatr. Res. 2008, 64, 246–250. [Google Scholar] [CrossRef]
- Espinosa, O.; Jiménez-Almazán, J.; Chaves, F.J.; Tormos, M.C.; Clapes, S.; Iradi, A.; Salvador, A.; Fandos, M.; Redón, J.; Sáez, G.T. Urinary 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxo-dG), a reliable oxidative stress marker in hypertension. Free Radic. Res. 2007, 41, 546–554. [Google Scholar] [CrossRef]
- Evans, M.D. Toward consensus in the analysis of urinary 8-oxo-7,8-dihydro-2′-deoxyguanosine as a noninvasive biomarker of oxidative stress. FASEB J. 2010, 24, 1249–1260. [Google Scholar]
- Il’Yasova, D.; Scarbrough, P.; Spasojevic, I. Urinary biomarkers of oxidative status. Clin. Chim. Acta 2012, 413, 1446–1453. [Google Scholar] [CrossRef]
- Sova, H.; Jukkolavuorinen, A.; Puistola, U.; Kauppila, S.; Karihtala, P. 8-Hydroxydeoxyguanosine: A new potential independent prognostic factor in breast cancer. Br. J. Cancer 2010, 102, 1018–1023. [Google Scholar] [CrossRef]
- Fischer-Nielsen, A.; Corcoran, G.B.; Poulsen, H.E.; Kamendulist, L.M.; Loft, S. Menadione-Induced DNA fragmentation without 8-oxo-2′-deoxyguanosine formation in isolated rat hepatocytes biochemical. Pharmacology 1995, 49, 1469–1474. [Google Scholar]
- Briedé, J.J.; Van Delft, J.M.H.; De Kok, T.M.C.M.; Van Herwijnen, M.H.M.; Maas, L.M.; Gottschalk, R.W.H.; Kleinjans, J.C.S. Global Gene Expression Analysis Reveals Differences in Cellular Responses to Hydroxyl- and Superoxide Anion Radical–Induced Oxidative Stress in Caco-2 Cells. Toxicol. Sci. 2009, 114, 193–203. [Google Scholar] [CrossRef]
- Legault, J.; Carrier, C.; Petrov, P.; Renard, P.; Remacle, J.; Mirault, M.-E. Mitochondrial GPx1 Decreases Induced but Not Basal Oxidative Damage to mtDNA in T47D Cells. Biochem. Biophys. Res. Commun. 2000, 272, 416–422. [Google Scholar] [CrossRef]
- Sibayama-Imazu, T.; Fujisawa, Y.; Masuda, Y.; Aiuchi, T.; Nakajo, S.; Itabe, H.; Nakaya, K. Induction of apoptosis in PA-1 ovarian cancer cells by vitamin K2 is associated with an increase in the level of TR3/Nur77 and its accumulation in mitochondria and nuclei. J. Cancer Res. Clin. Oncol. 2008, 134, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, M.; Tsuchida, A.; Miyazawa, K.; Yokoyama, T.; Kawakita, H.; Tokita, H.; Naito, M.; Itoh, M.; Ohyashiki, K.; Aoki, T. Vitamin K2-induced cell growth inhibition via autophagy formation in cholangiocellular carcinoma cell lines. Int. J. Mol. Med. 2007, 20, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Funato, K.; Miyazawa, K.; Yaguchi, M.; Gotoh, A.; Ohyashiki, K. Combination of 22-oxa-1,25-dihydroxyvitamin D (3), a vitamin D (3) Derivative, with vitamin k (2) (VK2) synergistically enhances Cell Differentiation But Suppresses VK2-In-Ducing Apoptosis in HL-60CELS. Leukemia 2002, 16, 1519–1527. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lu, H.; Mei, C.; Yang, L.; Zheng, J.; Tong, J.; Duan, F.; Liang, H.; Hong, L. PPM-18, an Analog of Vitamin K, Induces Autophagy and Apoptosis in Bladder Cancer Cells Through ROS and AMPK Signaling Pathways. Front. Pharmacol. 2021, 12, 684915. [Google Scholar] [CrossRef]
- Saibu, M.; Sagar, S.; Green, I.; Ameer, F.; Meyer, M. Evaluating the Cytotoxic Effects of Novel Quinone Compounds. Anticancer Res. 2014, 34, 4077–4086. [Google Scholar]
- Hitomi, M.; Yokoyama, F.; Kita, Y.; Nonomura, T.; Masaki, T.; Yoshiji, H.; Inoue, H.; Kinekawa, F.; Kurokohchi, K.; Uchida, N.; et al. Antitumor effects of vitamins K1, K2 and K3 on hepatocellular carcinoma in vitro and in vivo. Int. J. Oncol. 2005, 26, 713–720. [Google Scholar] [CrossRef]
- Ozaki, I.; Zhang, H.; Mizuta, T.; Ide, Y.; Eguchi, Y.; Yasutake, T.; Sakamaki, T.; Pestell, R.G.; Yamamoto, K. Menatetrenone, a vitamin K2 analogue, inhibits hepatocellular carcinoma cell growth by suppressing cyclin D1 expression through inhibition of nuclear factor kappaB activation. Clin. Cancer Res. 2007, 13, 2236–2245. [Google Scholar] [CrossRef]
- Yamamoto, T.; Nakamura, H.; Liu, W.; Cao, K.; Yoshikawa, S.; Enomoto, H.; Iwata, Y.; Koh, N.; Saito, M.; Imanishi, H. Involvement of hepatoma-derived growth factor in the growth inhibition of hepatocellular carcinoma cells by vitamin K (2). J. Gastroenterol. 2009, 44, 228–235. [Google Scholar] [CrossRef]
- Kaneda, M.; Zhang, D.; Bhattacharjee, R.; Nakahama, K.-I.; Arii, S.; Morita, I. Vitamin K2 suppresses malignancy of HuH7 hepatoma cells via inhibition of connexin 43. Cancer Lett. 2008, 263, 53–60. [Google Scholar] [CrossRef]
- Kandouz, M.; Batist, G. Gap junctions and connexins as therapeutic targets in cancer. Expert Opin. Ther. Targets 2010, 14, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, S.; Azuma, M.; Kasama, T.; Sakamoto, S.; Kabe, Y.; Imai, T.; Yamaguchi, Y.; Miyazawa, K.; Handa, H. Vitamin K2 Covalently Binds to Bak and Induces Bak-Mediated Apoptosis. Mol. Pharmacol. 2012, 83, 613–620. [Google Scholar] [CrossRef]
- Showalter, S.L.; Wang, Z.; Costantino, C.L.; Witkiewicz, A.K.; Yeo, C.J.; Brody, J.R.; Carr, B.I. Naturally occurring K vitamins inhibit pancreatic cancer cell survival through a caspase-dependent pathway. J. Gastroenterol. Hepatol. 2010, 25, 738–744. [Google Scholar] [CrossRef]
- Yokoyama, T.; Miyazawa, K.; Naito, M.; Toyotake, J.; Tauchi, T.; Itoh, M.; Yuo, A.; Hayashi, Y.; Georgescu, M.-M.; Kondo, Y.; et al. Vitamin K2 induces autophagy and apoptosis simultaneously in leukemia cells. Autophagy 2008, 4, 629–640. [Google Scholar] [CrossRef]
- Duan, F.; Mei, C.; Yang, L.; Zheng, J.; Lu, H.; Xia, Y.; Hsu, S.; Liang, H.; Hong, L. Vitamin K2 promotes PI3K/AKT/HIF-1α-mediated glycolysis that leads to AMPK-dependent autophagic cell death in bladder cancer cells. Sci. Rep. 2020, 10, 7714. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.; Hegazy, M.-E.F.; Elbadawi, M.; Fleischer, E.; Klinger, A.; Bringmann, G.; Kuntner, C.; Shan, L.; Efferth, T. Vitamin K3 chloro derivative (VKT-2) inhibits HDAC6, activates autophagy and apoptosis, and inhibits aggresome formation in hepatocellular carcinoma cells. Biochem. Pharmacol. 2020, 180, 114176. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, K.; Kayashima, T.; Mori, M.; Yoshida, H.; Mizushina, Y. Inhibitory effects of vitamin K3 on DNA polymerase and angiogenesis. Int. J. Mol. Med. 2008, 22, 381–387. [Google Scholar] [CrossRef]
- Mokhtari, R.B.; Homayouni, T.S.; Baluch, N.; Morgatskaya, E.; Kumar, S.; Das, B.; Yeger, H. Combination therapy in combating cancer. Oncotarget 2017, 8, 38022–38043. [Google Scholar] [CrossRef]
- Nutter, L.M.; Ann-Lii, C.; Hsiao-Ling, H.; Ruey-Kun, H.; Ngo, E.O.; Tsang-Wu, L. Menadione: Spectrum of anticancer activity and effects on nucleotide metabolism in human neoplastic cell lines. Biochem. Pharmacol. 1991, 41, 1283–1292. [Google Scholar] [CrossRef]
- D’Alessandro, R.; Refolo, M.G.; Lippolis, C.; Carella, N.; Messa, C.; Cavallini, A.; Carr, B.I. Strong enhancement by IGF1-R antagonists of hepatocellular carcinoma cell migration inhibition by Sorafenib and/or vitamin K1. Cell. Oncol. 2018, 41, 283–296. [Google Scholar] [CrossRef] [PubMed]
- Carr, B.I.; Wang, Z.; Wang, M.; Cavallini, A.; D’Alessandro, R.; Refolo, M.G. c-Met-Akt pathway-mediated enhancement of inhibitory c-Raf phosphorylation is involved in vitamin K1 and sorafenib synergy on HCC growth inhibition. Cancer Biol. Ther. 2011, 12, 531–538. [Google Scholar] [CrossRef]
- Refolo, M.G.; D’Alessandro, R.; Lippolis, C.; Carella, N.; Cavallini, A.; Messa, C.; Carr, B.I. IGF-1R tyrosine kinase inhibitors and Vitamin K1 enhance the antitumor effects of Regorafenib in HCC cell lines. Oncotarget 2017, 8, 103465–103476. [Google Scholar] [CrossRef]
- Ha, T.-Y.; Hwang, S.; Hong, H.-N.; Choi, Y.-I.; Yoon, S.-Y.; Won, Y.-J.; Song, G.-W.; Kim, N.; Tak, E.; Ryoo, B.-Y. Synergistic effect of sorafenib and vitamin K on suppression of hepatocellular carcinoma cell migration and metastasis. Anticancer Res. 2015, 35, 1985–1995. [Google Scholar] [PubMed]
- Wei, G.; Wang, M.; Hyslop, T.; Wang, Z.; Carr, B. Vitamin K enhancement of sorafenib-mediated HCC cell growth inhibition in vitro and in vivo. Int. J. Cancer 2010, 127, 2949–2958. [Google Scholar] [CrossRef]
- Wei, G.; Wang, M.; Carr, B.I. Sorafenib combined vitamin K induces apoptosis in human pancreatic cancer cell lines through RAF/MEK/ERK and c-Jun NH2-terminal kinase pathways. J. Cell. Physiol. 2010, 224, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, B.; Zhang, A.; Zhao, Y.; Zhao, J.; Liu, J.; Gao, J.; Fang, D.; Rao, Z. Synergistic growth inhibition by sorafenib and vitamin K2 in human hepatocellular carcinoma cells. Clinics 2012, 67, 1093–1099. [Google Scholar] [CrossRef]
- Zhang, H.; Ozaki, I.; Hamajima, H. Vitamin K2 augments 5-fluorouracil-induced growth inhibition of human hepatocellular carcinoma cells by inhibiting NF-κB activation. Oncol. Rep. 2011, 25, 159–166. [Google Scholar]
- Kitagawa, J.; Hara, T.; Tsurumi, H.; Ninomiya, S.; Ogawa, K.; Adachi, S.; Kanemura, N.; Kasahara, S.; Shimizu, M.; Moriwaki, H. Synergistic growth inhibition in HL-60 cells by the combination of acyclic retinoid and vitamin K2. J. Cancer Res. Clin. Oncol. 2010, 137, 779–787. [Google Scholar] [CrossRef]
- Kanamori, T.; Shimizu, M.; Okuno, M.; Matsushima-Nishiwaki, R.; Tsurumi, H.; Kojima, S.; Moriwaki, H. Synergistic growth inhibition by acyclic retinoid and vitamin K2 in human hepatocellular carcinoma cells. Cancer Sci. 2007, 98, 431–437. [Google Scholar] [CrossRef]
- Tomasetti, M.; Nocchi, L.; Neuzil, J.; Goodwin, J.; Nguyen, M.; Dong, L.; Manzella, N.; Staffolani, S.; Milanese, C.; Garrone, B.; et al. Alpha-Tocopheryl Succinate Inhibits Autophagic Survival of Prostate Cancer Cells Induced by Vitamin K3 and Ascorbate to Trigger Cell Death. PLoS ONE 2012, 7, e52263. [Google Scholar] [CrossRef]
- Ren, X.; Santhosh, S.M.; Coppo, L.; Ogata, F.T.; Lu, J.; Holmgren, A. The combination of ascorbate and menadione causes cancer cell death by oxidative stress and replicative stress. Free Radic. Biol. Med. 2019, 134, 350–358. [Google Scholar] [CrossRef]
- Bonilla-Porras, A.R.; Jimenez-Del-Rio, M.; Velez-Pardo, C. Vitamin K3 and vitamin C alone or in combination induced apoptosis in leukemia cells by a similar oxidative stress signalling mechanism. Cancer Cell Int. 2011, 11, 19. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, D.; Zhelev, Z.; Getsov, P.; Nikolova, B.; Aoki, I.; Higashi, T.; Bakalova, R. Vitamin K: Redox-modulation, prevention of mitochondrial dysfunction and anticancer effect. Redox Biol. 2018, 16, 352–358. [Google Scholar] [CrossRef]
- Bakalova, R.; Semkova, S.; Ivanova, D.; Zhelev, Z.; Miller, T.; Takeshima, T.; Shibata, S.; Lazarova, D.; Aoki, I.; Higashi, T. Selective Targeting of Cancerous Mitochondria and Suppression of Tumor Growth Using Redox-Active Treatment Adjuvant. Oxidative Med. Cell. Longev. 2020, 2020, 6212935. [Google Scholar] [CrossRef]
- Nimptsch, K.; Rohrmann, S.; Kaaks, R.; Linseisen, J. Dietary vitamin K intake in relation to cancer incidence and mortality: Results from the Heidelberg cohort of the European Prospective Investigation into Cancer and Nutrition (EPIC-Heidelberg). Am. J. Clin. Nutr. 2010, 91, 1348–1358. [Google Scholar] [CrossRef]
- Yu, D.-W.; Li, Q.-J.; Cheng, L.; Yang, P.-F.; Sun, W.-P.; Peng, Y.; Hu, J.-J.; Wu, J.-J.; Gong, J.-P.; Zhong, G.-C. Dietary Vitamin K Intake and the Risk of Pancreatic Cancer: A Prospective Study of 101,695 American Adults. Am. J. Epidemiol. 2021, 190, 2029–2041. [Google Scholar] [CrossRef] [PubMed]
- Mizuta, T.; Ozaki, I.; Eguchi, Y.; Yasutake, T.; Kawazoe, S.; Fujimoto, K.; Yamamoto, K. The effect of menatetrenone, a vitamin K2 analog, on disease recurrence and survival in patients with hepatocellular carcinoma after curative treatment. Cancer 2006, 106, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.-H.; Mo, X.-S.; Xiang, B.-D.; Yuan, W.-P.; Jiang, J.-F.; Xie, G.-S.; Li, L.-Q. Postoperative Use of the Chemopreventive Vitamin K2 Analog in Patients with Hepatocellular Carcinoma. PLoS ONE 2013, 8, e58082. [Google Scholar] [CrossRef]
- Booth, S.L.; Rajabi, A.A. Determinants of Vitamin K Status in Humans. Vitamin, K. Vitam. Horm. 2008, 78, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ferland, G.; Sadowski, J.A.; O’Brien, M.E. Dietary induced subclinical vitamin K deficiency in normal human subjects. J. Clin. Investig. 1993, 91, 1761–1768. [Google Scholar] [CrossRef]
- Desai, A.P.; Dirajlal-Fargo, S.; Durieux, J.C.; Tribout, H.; Labbato, D.; McComsey, G.A. Vitamin K & D Deficiencies Are Independently Associated With COVID-19 Disease Severity. Open Forum Infect. Dis. 2021, 8, ofab408. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaźmierczak-Barańska, J.; Karwowski, B.T. Vitamin K Contribution to DNA Damage—Advantage or Disadvantage? A Human Health Response. Nutrients 2022, 14, 4219. https://doi.org/10.3390/nu14204219
Kaźmierczak-Barańska J, Karwowski BT. Vitamin K Contribution to DNA Damage—Advantage or Disadvantage? A Human Health Response. Nutrients. 2022; 14(20):4219. https://doi.org/10.3390/nu14204219
Chicago/Turabian StyleKaźmierczak-Barańska, Julia, and Bolesław T. Karwowski. 2022. "Vitamin K Contribution to DNA Damage—Advantage or Disadvantage? A Human Health Response" Nutrients 14, no. 20: 4219. https://doi.org/10.3390/nu14204219
APA StyleKaźmierczak-Barańska, J., & Karwowski, B. T. (2022). Vitamin K Contribution to DNA Damage—Advantage or Disadvantage? A Human Health Response. Nutrients, 14(20), 4219. https://doi.org/10.3390/nu14204219