
Functional Alternatives to Alcohol

 ,
, 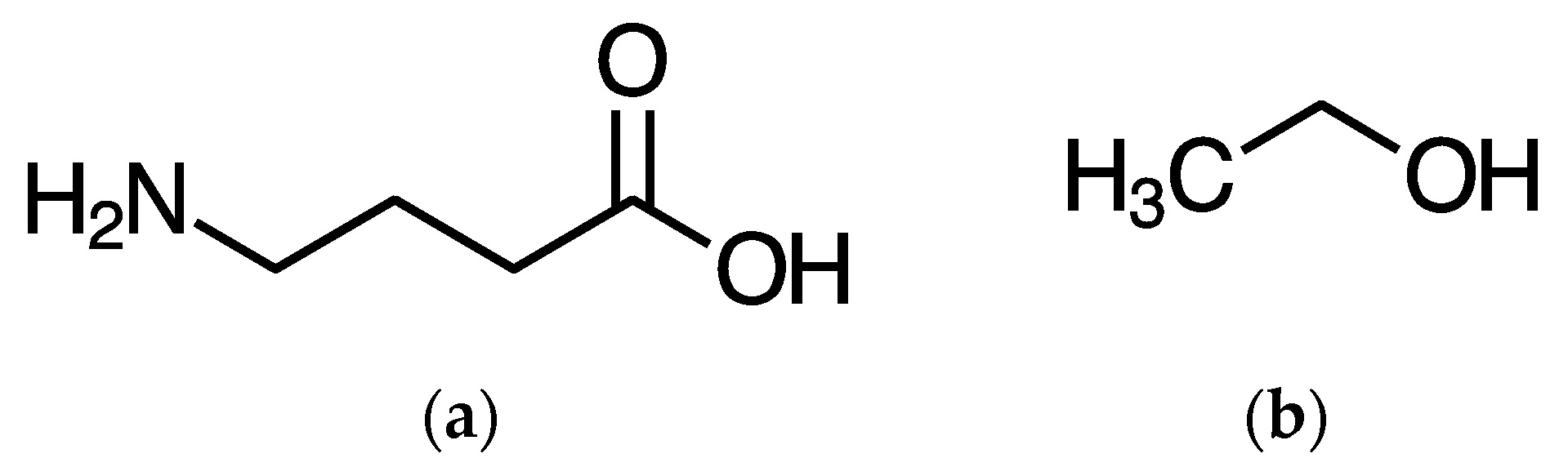{kind=link}
Abstract
:1. Introduction
2. How Does Alcohol Work?
3. Can Targeting the GABA Receptor Provide Us with an Alcohol Alternative?
4. Other Molecules That Work to Enhance GABA
5. Neurosteroids and GABAA Receptors
6. The Search for Synthetic Alcohol Alternatives
7. Botanical Alternatives to Alcohol
NS and NS-like Molecules in Plants: An Alcohol Alternative?
8. Other Possible Molecular GABA Targets
GABAB and GHB Receptors
9. From Bench to Bar: What Are the Regulatory and Other Challenges?
Author Contributions
Funding
Conflicts of Interest
References
- Nutt, D.J.; King, L.A.; Phillips, L.D.; Independent Scientific Committee on Drugs. Drug harms in the UK: A multicriteria decision analysis. Lancet 2010, 376, 1558–1565. [Google Scholar] [CrossRef]
- van Amsterdam, J.; Nutt, D.; Phillips, L.; van den Brink, W. European rating of drug harms. J. Psychopharmacol. 2015, 29, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Bonomo, Y.; Norman, A.; Biondo, S.; Bruno, R.; Daglish, M.; Dawe, S.; Egerton-Warburton, D.; Karro, J.; Kim, C.; Lenton, S.; et al. The Australian drug harms ranking study. J. Psychopharmacol. 2019, 33, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Nutt, D.J.; Gual, A.; Anderson, P.; Rehm, J. Why Less Is Always More in the Treatment of Alcohol Use Disorders. JAMA Psychiatry 2019, 76, 359–360. [Google Scholar] [CrossRef]
- Nutt, D.J. Alcohol alternatives—A goal for psychopharmacology? J. Psychopharmacol. 2006, 20, 318–320. [Google Scholar] [CrossRef]
- Foresight, R. How Future Scientific Advances May Affect Our Understanding of Addiction and Drug Use. Available online: https://www.gov.uk/government/publications/drugs-futures-2025 (accessed on 16 August 2022).
- Nutt, D.; Hayes, A.; Fonville, L.; Zafar, R.; Palmer, E.O.C.; Paterson, L.; Lingford-Hughes, A. Alcohol and the Brain. Nutrients 2021, 13, 3938. [Google Scholar] [CrossRef]
- Abrahao, K.P.; Salinas, A.G.; Lovinger, D.M. Alcohol and the Brain: Neuronal Molecular Targets, Synapses, and Circuits. Neuron 2017, 96, 1223–1238. [Google Scholar] [CrossRef]
- Roerecke, M. Alcohol’s Impact on the Cardiovascular System. Nutrients 2021, 13, 3419. [Google Scholar] [CrossRef]
- Pohl, K.; Moodley, P.; Dhanda, A.D. Alcohol’s Impact on the Gut and Liver. Nutrients 2021, 13, 3170. [Google Scholar] [CrossRef]
- Rumgay, H.; Murphy, N.; Ferrari, P.; Soerjomataram, I. Alcohol and Cancer: Epidemiology and Biological Mechanisms. Nutrients 2021, 13, 3173. [Google Scholar] [CrossRef]
- Slingerland, E. Drunk: How We Sipped, Danced, and Stumbled Our Way to Civilization; Little, Brown Spark: New York, NY, USA, 2021. [Google Scholar]
- Lembke, A. Dopamine Nation: Finding Balance in the Age of Indulgence.; Penguin: London, UK, 2021. [Google Scholar]
- Hoffman, P.L.; Rabe, C.S.; Grant, K.A.; Valverius, P.; Hudspith, M.; Tabakoff, B. Ethanol and the NMDA receptor. Alcohol 1990, 7, 229–231. [Google Scholar] [CrossRef]
- Kobayashi, N.H.C.; Farias, S.V.; Luz, D.A.; Machado-Ferraro, K.M.; Conceicao, B.C.D.; Silveira, C.; Fernandes, L.M.P.; Cartagenes, S.C.; Ferreira, V.M.M.; Fontes-Junior, E.A.; et al. Ketamine plus Alcohol: What We Know and What We Can Expect about This. Int. J. Mol. Sci. 2022, 23, 7800. [Google Scholar] [CrossRef] [PubMed]
- Lovinger, D.M. 5-HT3 receptors and the neural actions of alcohols: An increasingly exciting topic. Neurochem. Int. 1999, 35, 125–130. [Google Scholar] [CrossRef]
- Koob, G.F.; Buck, C.L.; Cohen, A.; Edwards, S.; Park, P.E.; Schlosburg, J.E.; Schmeichel, B.; Vendruscolo, L.F.; Wade, C.L.; Whitfield, T.W., Jr.; et al. Addiction as a stress surfeit disorder. Neuropharmacology 2014, 76, 370–382. [Google Scholar] [CrossRef] [PubMed]
- Crews, F.T.; Lawrimore, C.J.; Walter, T.J.; Coleman, L.G., Jr. The role of neuroimmune signaling in alcoholism. Neuropharmacology 2017, 122, 56–73. [Google Scholar] [CrossRef] [PubMed]
- Quillin, S.J.; Tran, P.; Prindle, A. Potential Roles for Gamma-Aminobutyric Acid Signaling in Bacterial Communities. Bioelectricity 2021, 3, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; O’Riordan, K.J.; Cowan, C.S.M.; Sandhu, K.V.; Bastiaanssen, T.F.S.; Boehme, M.; Codagnone, M.G.; Cussotto, S.; Fulling, C.; Golubeva, A.V.; et al. The Microbiota-Gut-Brain Axis. Physiol. Rev. 2019, 99, 1877–2013. [Google Scholar] [CrossRef]
- Strandwitz, P.; Kim, K.H.; Terekhova, D.; Liu, J.K.; Sharma, A.; Levering, J.; McDonald, D.; Dietrich, D.; Ramadhar, T.R.; Lekbua, A.; et al. GABA-modulating bacteria of the human gut microbiota. Nat. Microbiol. 2019, 4, 396–403. [Google Scholar] [CrossRef]
- Barrett, E.; Ross, R.P.; O’Toole, P.W.; Fitzgerald, G.F.; Stanton, C. gamma-Aminobutyric acid production by culturable bacteria from the human intestine. J. Appl. Microbiol. 2012, 113, 411–417. [Google Scholar] [CrossRef]
- Otaru, N.; Ye, K.; Mujezinovic, D.; Berchtold, L.; Constancias, F.; Cornejo, F.A.; Krzystek, A.; de Wouters, T.; Braegger, C.; Lacroix, C.; et al. GABA Production by Human Intestinal Bacteroides spp.: Prevalence, Regulation, and Role in Acid Stress Tolerance. Front. Microbiol. 2021, 12, 656895. [Google Scholar] [CrossRef]
- Tillisch, K.; Mayer, E.A.; Gupta, A.; Gill, Z.; Brazeilles, R.; Le Neve, B.; van Hylckama Vlieg, J.E.T.; Guyonnet, D.; Derrien, M.; Labus, J.S. Brain Structure and Response to Emotional Stimuli as Related to Gut Microbial Profiles in Healthy Women. Psychosom. Med. 2017, 79, 905–913. [Google Scholar] [CrossRef] [PubMed]
- Belelli, D.; Hales, T.G.; Lambert, J.J.; Luscher, B.; Olsen, R.; Peters, J.A.; Rudolph, U.; Sieghart, W. GABAA receptors in GtoPdb v.2021.3. IUPHAR/BPS Guide Pharmacol. CITE 2021, 2021. [Google Scholar] [CrossRef]
- Alexander, S.P.; Christopoulos, A.; Davenport, A.P.; Kelly, E.; Mathie, A.; Peters, J.A.; Veale, E.L.; Armstrong, J.F.; Faccenda, E.; Harding, S.D.; et al. THE CONCISE GUIDE TO PHARMACOLOGY 2021/22: G protein-coupled receptors. Br. J. Pharmacol. 2021, 178 (Suppl. S1), S27–S156. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, D. Keeping the Balance: GABAB Receptors in the Developing Brain and Beyond. Brain Sci. 2022, 12, 419. [Google Scholar] [CrossRef] [PubMed]
- Wisden, W.; Laurie, D.J.; Monyer, H.; Seeburg, P.H. The distribution of 13 GABAA receptor subunit mRNAs in the rat brain. I. Telencephalon, diencephalon, mesencephalon. J. Neurosci. 1992, 12, 1040–1062. [Google Scholar] [CrossRef]
- Maramai, S.; Benchekroun, M.; Ward, S.E.; Atack, J.R. Subtype Selective gamma-Aminobutyric Acid Type A Receptor (GABAAR) Modulators Acting at the Benzodiazepine Binding Site: An Update. J. Med. Chem. 2020, 63, 3425–3446. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.; Stephens, D.N.; Duka, T. Lorazepam substitutes for the alcohol stimulus in social drinkers. Psychopharmacology 2003, 166, 181–187. [Google Scholar] [CrossRef]
- Belelli, D.; Lambert, J.J. Neurosteroids: Endogenous regulators of the GABA(A) receptor. Nat. Rev. Neurosci. 2005, 6, 565–575. [Google Scholar] [CrossRef]
- Hosie, A.M.; Wilkins, M.E.; da Silva, H.M.; Smart, T.G. Endogenous neurosteroids regulate GABAA receptors through two discrete transmembrane sites. Nature 2006, 444, 486–489. [Google Scholar] [CrossRef]
- Laverty, D.; Thomas, P.; Field, M.; Andersen, O.J.; Gold, M.G.; Biggin, P.C.; Gielen, M.; Smart, T.G. Crystal structures of a GABAA-receptor chimera reveal new endogenous neurosteroid-binding sites. Nat. Struct. Mol. Biol. 2017, 24, 977–985. [Google Scholar] [CrossRef]
- Miller, P.S.; Scott, S.; Masiulis, S.; De Colibus, L.; Pardon, E.; Steyaert, J.; Aricescu, A.R. Structural basis for GABAA receptor potentiation by neurosteroids. Nat. Struct. Mol. Biol. 2017, 24, 986–992. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wells, M.M.; Arjunan, P.; Tillman, T.S.; Cohen, A.E.; Xu, Y.; Tang, P. Structural basis of neurosteroid anesthetic action on GABAA receptors. Nat. Commun. 2018, 9, 3972. [Google Scholar] [CrossRef] [PubMed]
- Carter, R.B.; Wood, P.L.; Wieland, S.; Hawkinson, J.E.; Belelli, D.; Lambert, J.J.; White, H.S.; Wolf, H.H.; Mirsadeghi, S.; Tahir, S.H.; et al. Characterization of the anticonvulsant properties of ganaxolone (CCD 1042; 3alpha-hydroxy-3beta-methyl-5alpha-pregnan-20-one), a selective, high-affinity, steroid modulator of the gamma-aminobutyric acid(A) receptor. J. Pharmacol. Exp. Ther. 1997, 280, 1284–1295. [Google Scholar] [PubMed]
- Knight, E.M.P.; Amin, S.; Bahi-Buisson, N.; Benke, T.A.; Cross, J.H.; Demarest, S.T.; Olson, H.E.; Specchio, N.; Fleming, T.R.; Aimetti, A.A.; et al. Safety and efficacy of ganaxolone in patients with CDKL5 deficiency disorder: Results from the double-blind phase of a randomised, placebo-controlled, phase 3 trial. Lancet Neurol. 2022, 21, 417–427. [Google Scholar] [CrossRef]
- Belelli, D.; Hogenkamp, D.; Gee, K.W.; Lambert, J.J. Realising the therapeutic potential of neuroactive steroid modulators of the GABAA receptor. Neurobiol. Stress 2020, 12, 100207. [Google Scholar] [CrossRef]
- Althaus, A.L.; Ackley, M.A.; Belfort, G.M.; Gee, S.M.; Dai, J.; Nguyen, D.P.; Kazdoba, T.M.; Modgil, A.; Davies, P.A.; Moss, S.J.; et al. Preclinical characterization of zuranolone (SAGE-217), a selective neuroactive steroid GABAA receptor positive allosteric modulator. Neuropharmacology 2020, 181, 108333. [Google Scholar] [CrossRef]
- Deligiannidis, K.M.; Meltzer-Brody, S.; Gunduz-Bruce, H.; Doherty, J.; Jonas, J.; Li, S.; Sankoh, A.J.; Silber, C.; Campbell, A.D.; Werneburg, B.; et al. Effect of Zuranolone vs Placebo in Postpartum Depression: A Randomized Clinical Trial. JAMA Psychiatry 2021, 78, 951–959. [Google Scholar] [CrossRef]
- Gunduz-Bruce, H.; Takahashi, K.; Huang, M.Y. Development of neuroactive steroids for the treatment of postpartum depression. J. Neuroendocrinol. 2022, 34, e13019. [Google Scholar] [CrossRef]
- Bowen, C.A.; Purdy, R.H.; Grant, K.A. Ethanol-like discriminative stimulus effects of endogenous neuroactive steroids: Effect of ethanol training dose and dosing procedure. J. Pharmacol. Exp. Ther. 1999, 289, 405–411. [Google Scholar]
- Allen, D.C.; Ford, M.M.; Grant, K.A. Cross-Species Translational Findings in the Discriminative Stimulus Effects of Ethanol. Curr. Top. Behav. Neurosci. 2018, 39, 95–111. [Google Scholar] [CrossRef]
- Helms, C.M.; McCracken, A.D.; Heichman, S.L.; Moschak, T.M. Ovarian hormones and the heterogeneous receptor mechanisms mediating the discriminative stimulus effects of ethanol in female rats. Behav. Pharmacol. 2013, 24, 95–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finn, D.A.; Beadles-Bohling, A.S.; Beckley, E.H.; Ford, M.M.; Gililland, K.R.; Gorin-Meyer, R.E.; Wiren, K.M. A new look at the 5alpha-reductase inhibitor finasteride. CNS Drug Rev. 2006, 12, 53–76. [Google Scholar] [CrossRef] [PubMed]
- Khisti, R.T.; VanDoren, M.J.; O’Buckley, T.; Morrow, A.L. Neuroactive steroid 3 alpha-hydroxy-5 alpha-pregnan-20-one modulates ethanol-induced loss of righting reflex in rats. Brain Res. 2003, 980, 255–265. [Google Scholar] [CrossRef]
- Hirani, K.; Sharma, A.N.; Jain, N.S.; Ugale, R.R.; Chopde, C.T. Evaluation of GABAergic neuroactive steroid 3alpha-hydroxy-5alpha-pregnane-20-one as a neurobiological substrate for the anti-anxiety effect of ethanol in rats. Psychopharmacology 2005, 180, 267–278. [Google Scholar] [CrossRef]
- VanDoren, M.J.; Matthews, D.B.; Janis, G.C.; Grobin, A.C.; Devaud, L.L.; Morrow, A.L. Neuroactive steroid 3alpha-hydroxy-5alpha-pregnan-20-one modulates electrophysiological and behavioral actions of ethanol. J. Neurosci. 2000, 20, 1982–1989. [Google Scholar] [CrossRef]
- Hirani, K.; Khisti, R.T.; Chopde, C.T. Behavioral action of ethanol in Porsolt’s forced swim test: Modulation by 3 alpha-hydroxy-5 alpha-pregnan-20-one. Neuropharmacology 2002, 43, 1339–1350. [Google Scholar] [CrossRef]
- Torres, J.M.; Ortega, E. Alcohol intoxication increases allopregnanolone levels in female adolescent humans. Neuropsychopharmacology 2003, 28, 1207–1209. [Google Scholar] [CrossRef]
- Torres, J.M.; Ortega, E. Alcohol intoxication increases allopregnanolone levels in male adolescent humans. Psychopharmacology 2004, 172, 352–355. [Google Scholar] [CrossRef]
- Morrow, A.L.; Boero, G.; Porcu, P. A Rationale for Allopregnanolone Treatment of Alcohol Use Disorders: Basic and Clinical Studies. Alcohol. Clin. Exp. Res. 2020, 44, 320–339. [Google Scholar] [CrossRef] [PubMed]
- Durkin, E.J.; Muessig, L.; Herlt, T.; Lumb, M.J.; Patel, R.; Thomas, P.; Bright, D.P.; Jurd, R.; Moss, S.J.; Dickenson, A.H.; et al. Brain neurosteroids are natural anxiolytics targeting α2 subunit γ-aminobutyric acid type-A receptors. BioRXiv 2018. [Google Scholar] [CrossRef]
- Belelli, D.; Phillips, G.D.; Atack, J.R.; Lambert, J.J. Relating neurosteroid modulation of inhibitory neurotransmission to behaviour. J. Neuroendocrinol. 2022, 34, e13045. [Google Scholar] [CrossRef] [PubMed]
- Zolpidem. Available online: https://www.drugs.com/zolpidem.html# (accessed on 24 August 2022).
- Zoplicone. Available online: https://www.drugs.com/zopiclone.html (accessed on 24 August 2022).
- Sikdar, S. Physical dependence on zopiclone. Prescribing this drug to addicts may give rise to iatrogenic drug misuse. BMJ 1998, 317, 146. [Google Scholar] [CrossRef] [PubMed]
- June, H.L.; Foster, K.L.; McKay, P.F.; Seyoum, R.; Woods, J.E.; Harvey, S.C.; Eiler, W.J.; Grey, C.; Carroll, M.R.; McCane, S.; et al. The reinforcing properties of alcohol are mediated by GABA(A1) receptors in the ventral pallidum. Neuropsychopharmacology 2003, 28, 2124–2137. [Google Scholar] [CrossRef] [PubMed]
- Atack, J.R. GABAA receptor subtype-selective modulators. I. alpha2/alpha3-selective agonists as non-sedating anxiolytics. Curr. Top. Med. Chem. 2011, 11, 1176–1202. [Google Scholar] [CrossRef] [PubMed]
- AZD7325 Clinical Trial. Available online: https://www.clinicaltrials.gov/ct2/results?term=AZD7325&Search=Sear (accessed on 16 August 2022).
- June, H.L.; Harvey, S.C.; Foster, K.L.; McKay, P.F.; Cummings, R.; Garcia, M.; Mason, D.; Grey, C.; McCane, S.; Williams, L.S.; et al. GABA(A) receptors containing (alpha)5 subunits in the CA1 and CA3 hippocampal fields regulate ethanol-motivated behaviors: An extended ethanol reward circuitry. J. Neurosci. 2001, 21, 2166–2177. [Google Scholar] [CrossRef]
- Zhu, S.; Noviello, C.M.; Teng, J.; Walsh, R.M., Jr.; Kim, J.J.; Hibbs, R.E. Structure of a human synaptic GABAA receptor. Nature 2018, 559, 67–72. [Google Scholar] [CrossRef]
- Masiulis, S.; Desai, R.; Uchanski, T.; Serna Martin, I.; Laverty, D.; Karia, D.; Malinauskas, T.; Zivanov, J.; Pardon, E.; Kotecha, A.; et al. GABAA receptor signalling mechanisms revealed by structural pharmacology. Nature 2019, 565, 454–459. [Google Scholar] [CrossRef]
- Kim, J.J.; Gharpure, A.; Teng, J.; Zhuang, Y.; Howard, R.J.; Zhu, S.; Noviello, C.M.; Walsh, R.M., Jr.; Lindahl, E.; Hibbs, R.E. Shared structural mechanisms of general anaesthetics and benzodiazepines. Nature 2020, 585, 303–308. [Google Scholar] [CrossRef]
- Sauguet, L.; Shahsavar, A.; Poitevin, F.; Huon, C.; Menny, A.; Nemecz, A.; Haouz, A.; Changeux, J.P.; Corringer, P.J.; Delarue, M. Crystal structures of a pentameric ligand-gated ion channel provide a mechanism for activation. Proc. Natl. Acad. Sci. USA 2014, 111, 966–971. [Google Scholar] [CrossRef]
- Forstera, B.; Castro, P.A.; Moraga-Cid, G.; Aguayo, L.G. Potentiation of Gamma Aminobutyric Acid Receptors (GABAAR) by Ethanol: How Are Inhibitory Receptors Affected? Front. Cell. Neurosci. 2016, 10, 114. [Google Scholar] [CrossRef]
- Greenblatt, D.J.; Shader, R.I.; Divoll, M.; Harmatz, J.S. Benzodiazepines: A summary of pharmacokinetic properties. Br. J. Clin. Pharmacol. 1981, 11 (Suppl. S1), 11S–16S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnersley, A.M.; Turano, F.J. Gamma aminobutyric acid (GABA) and plant responses to stress. Crit. Rev. Plant Sci. 2000, 19, 479–509. [Google Scholar] [CrossRef]
- Li, L.; Dou, N.; Zhang, H.; Wu, C.X. The versatile GABA in plants. Plant Signal. Behav. 2021, 16, 1862565–1862576. [Google Scholar] [CrossRef] [PubMed]
- Cullinan, W.E.; Ziegler, D.R.; Herman, J.P. Functional role of local GABAergic influences on the HPA axis. Brain Struct. Funct. 2008, 213, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Cicek, S.S. Structure-Dependent Activity of Natural GABA(A) Receptor Modulators. Molecules 2018, 23, 1512. [Google Scholar] [CrossRef]
- Atal, C.K.; Dubey, R.K.; Singh, J. Biochemical Basis of Enhanced Drug Bioavailability by Piperine—Evidence That Piperine Is a Potent Inhibitor of Drug-Metabolism. J. Pharmacol. Exp. Ther. 1985, 232, 258–262. [Google Scholar]
- Zhang, Q.L.; Fu, B.M.M.; Zhang, Z.J. Borneol, a novel agent that improves central nervous system drug delivery by enhancing blood-brain barrier permeability. Drug Deliv. 2017, 24, 1037–1044. [Google Scholar] [CrossRef]
- Sinyard, A. Sentia: Is this Alcohol-Free Spirit that Makes You “Tipsy” the Future of Drinking? Available online: https://www.stylist.co.uk/health/alcohol-free-spirit-tipsy-sentia/574441 (accessed on 16 August 2022).
- Schuster-Bruce, C. I Tried an Alcohol-Free, No-Hangover Drink Made by a Top Professor that Claims to Make You as Relaxed as Alcohol Does. It Hits the Spot—but Make Sure You Read the Label. Available online: https://www.businessinsider.com/i-tried-alcohol-free-drink-professor-relaxed-alcohol-no-hangover-2021-3?r=US&IR=T (accessed on 16 August 2022).
- Derrick, F. This Alcohol-Free Spirit Promises to Give You All the Good Bits of Booze—And None of the Bad. Available online: https://metro.co.uk/2021/01/22/alcohol-free-spirit-sentia-promises-good-effects-of-booze-without-a-hangover-13947226/ (accessed on 16 August 2022).
- Kin Euphorics. Available online: https://www.kineuphorics.com/ (accessed on 16 August 2022).
- Lapin, I. Phenibut (beta-phenyl-GABA): A tranquilizer and nootropic drug. CNS Drug Rev. 2001, 7, 471–481. [Google Scholar] [CrossRef]
- U.S. Food and Drug Administration (FDA). FDA Acts on Dietary Supplements Containing DMHA and Phenibut. Available online: https://www.fda.gov/food/cfsan-constituent-updates/fda-acts-dietary-supplements-containing-dmha-and-phenibut (accessed on 16 August 2022).
- Boonstra, E.; de Kleijn, R.; Colzato, L.S.; Alkemade, A.; Forstmann, B.U.; Nieuwenhuis, S. Neurotransmitters as food supplements: The effects of GABA on brain and behavior. Front. Psychol. 2015, 6, 1520–1525. [Google Scholar] [CrossRef]
- Three Spirit. Available online: https://threespiritdrinks.com/ (accessed on 16 August 2022).
- Grundmann, O.; Wang, J.; McGregor, G.P.; Butterweck, V. Anxiolytic Activity of a Phytochemically Characterized Passiflora incarnata Extract is Mediated via the GABAergic System. Planta Med. 2008, 74, 1769–1773. [Google Scholar] [CrossRef]
- Awad, R.; Muhammad, A.; Durst, T.; Trudeau, V.L.; Arnason, J.T. Bioassay-guided Fractionation of Lemon Balm (Melissa officinalis L.) using an In Vitro Measure of GABA Transaminase Activity. Phytother. Res. 2009, 23, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Rosati, F.; Danza, G.; Guarna, A.; Cini, N.; Racchi, M.L.; Serio, M. New evidence of similarity between human and plant steroid metabolism: 5alpha-reductase activity in Solanum malacoxylon. Endocrinology 2003, 144, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Rosati, F.; Bardazzi, I.; De Blasi, P.; Simi, L.; Scarpi, D.; Guarna, A.; Serio, M.; Racchi, M.L.; Danza, G. 5alpha-Reductase activity in Lycopersicon esculentum: Cloning and functional characterization of LeDET2 and evidence of the presence of two isoenzymes. J. Steroid Biochem. Mol. Biol. 2005, 96, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Janzen, D.; Slavik, B.; Zehe, M.; Sotriffer, C.; Loos, H.M.; Buettner, A.; Villmann, C. Sesquiterpenes and sesquiterpenoids harbor modulatory allosteric potential and affect inhibitory GABAA receptor function in vitro. J. Neurochem. 2021, 159, 101–115. [Google Scholar] [CrossRef]
- Tyacke, R.J.; Lingford-Hughes, A.; Reed, L.J.; Nutt, D.J. GABAB receptors in addiction and its treatment. Adv. Pharmacol. 2010, 58, 373–396. [Google Scholar] [CrossRef] [PubMed]
- Durant, C.F.; Paterson, L.M.; Turton, S.; Wilson, S.J.; Myers, J.F.M.; Muthukumaraswamy, S.; Venkataraman, A.; Mick, I.; Paterson, S.; Jones, T.; et al. Using Baclofen to Explore GABA-B Receptor Function in Alcohol Dependence: Insights From Pharmacokinetic and Pharmacodynamic Measures. Front. Psychiatry 2018, 9, 664. [Google Scholar] [CrossRef] [PubMed]
- Rolland, B.; Simon, N.; Franchitto, N.; Aubin, H.J. France Grants an Approval to Baclofen for Alcohol Dependence. Alcohol Alcohol. 2020, 55, 44–45. [Google Scholar] [CrossRef] [PubMed]
- Chick, J.; Nutt, D.J. Substitution therapy for alcoholism: Time for a reappraisal? J. Psychopharmacol. 2012, 26, 205–212. [Google Scholar] [CrossRef]
- Augier, E. Recent Advances in the Potential of Positive Allosteric Modulators of the GABAB Receptor to Treat Alcohol Use Disorder. Alcohol Alcohol. 2021, 56, 139–148. [Google Scholar] [CrossRef]
- EU Novel Food Catalogue 2022. Available online: https://webgate.ec.europa.eu/fip/novel_food_catalogue/# (accessed on 16 August 2022).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nutt, D.J.; Tyacke, R.J.; Spriggs, M.; Jacoby, V.; Borthwick, A.D.; Belelli, D. Functional Alternatives to Alcohol. Nutrients 2022, 14, 3761. https://doi.org/10.3390/nu14183761
Nutt DJ, Tyacke RJ, Spriggs M, Jacoby V, Borthwick AD, Belelli D. Functional Alternatives to Alcohol. Nutrients. 2022; 14(18):3761. https://doi.org/10.3390/nu14183761
Chicago/Turabian StyleNutt, David J., Robin J. Tyacke, Meg Spriggs, Vanessa Jacoby, Alan D. Borthwick, and Delia Belelli. 2022. "Functional Alternatives to Alcohol" Nutrients 14, no. 18: 3761. https://doi.org/10.3390/nu14183761
APA StyleNutt, D. J., Tyacke, R. J., Spriggs, M., Jacoby, V., Borthwick, A. D., & Belelli, D. (2022). Functional Alternatives to Alcohol. Nutrients, 14(18), 3761. https://doi.org/10.3390/nu14183761