

Phytochemical Profiles, Antioxidant Activity and Antiproliferative Mechanism of Rhodiola rosea L. Phenolic Extract


Abstract
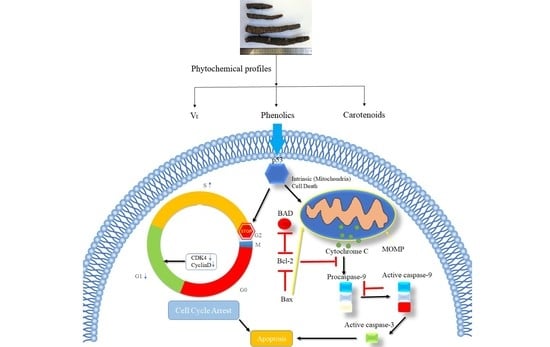
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Extraction of Rhodiola Phenolics
2.2.1. Extraction of RFE
2.2.2. Extraction of RBE
2.3. Determination of Total Polyphenols
2.4. Analysis of Phenolic Profiles in Rhodiola rosea by HPLC
2.5. Profiles of Volatile Compounds in Rhodiola rosea Detected by Static Headspace-Gas Chromatography–Tandem Mass Spectrometry (GC-MS)
2.6. Extraction and Profiles of Vitamin E and Carotenoids
2.6.1. Extraction of Vitamin E and Carotenoids
2.6.2. Vitamin E Analysis by NP-HPLC
2.6.3. Carotenoid Analysis by HPLC
2.7. Cell Culture
2.8. Antioxidant Activity Assay
2.8.1. In Vitro Antioxidant Assay
2.8.2. Cellular Antioxidant Activity (CAA) Assay
2.9. Cytotoxicity and Antiproliferative Activity Assays
2.10. The Determination of Synergy Effects
2.11. Cell Cycle and Cell Apoptosis Detection Assay
2.12. RT-qPCR Analysis
2.13. Western Blot Analysis
2.14. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Phenolic Contents in RFE
3.2. Profiles of Volatile Compounds in Rhodiola rosea
3.3. Analysis of VE and Carotenoid Contents in Rhodiola rosea
3.4. In Vitro Antioxidant Activity of Rhodiola rosea Phenolics
3.5. Cellular Antioxidant Activity of RE
3.6. Cytotoxicity and Antiproliferative Effects of RFE
3.7. Combined Effect of GA, EGC and EGCG on Inhibiting the Proliferation of HepG2 Cells
3.8. Analysis of CI and DRI Values
3.9. PCA Analysis
3.10. Effect of RFE on Cell Cycle Arrest
3.11. Effect of RFE on Apoptosis Induction
3.12. Effect of RFE on Mitochondrial Pathway-Related Gene and Protein Expression
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reddy, L.; Odhav, B.; Bhoola, K. Natural products for cancer prevention: A global perspective. Pharmacol. Ther. 2003, 99, 1–13. [Google Scholar] [CrossRef]
- Liu, R.H. Health-Promoting Components of Fruits and Vegetables in the Diet. Adv. Nutr. Int. Rev. J. 2013, 4, 384S–392S. [Google Scholar] [CrossRef]
- Surh, Y.-J. Cancer chemoprevention with dietary phytochemicals. Nat. Rev. Cancer 2003, 3, 768–780. [Google Scholar] [CrossRef]
- Kwon, K.H.; Barve, A.; Yu, S.; Huang, M.-T.; Kong, A.-N.T. Cancer chemoprevention by phytochemicals: Potential molecular targets, biomarkers and animal models. Acta Pharmacol. Sin. 2007, 28, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, M.; Roy, K.; Janarthan, M.; Das, S.; Chatterjee, M. Biological Activity of Carotenoids: Its Implications in Cancer Risk and Prevention. Curr. Pharm. Biotechnol. 2011, 13, 180–190. [Google Scholar] [CrossRef]
- Cianciosi, D.; Forbes-Hernández, T.Y.; Afrin, S.; Gasparrini, M.; Reboredo-Rodriguez, P.; Manna, P.P.; Zhang, J.; Bravo Lamas, L.; Martínez Flórez, S.; Agudo Toyos, P.; et al. Phenolic Compounds in Honey and Their Associated Health Benefits: A Review. Molecules 2018, 23, 2322. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Li, H.-B.; Sui, Z.-Q.; Corke, H. Absorption, metabolism, anti-cancer effect and molecular targets of epigallocatechin gallate (EGCG): An updated review. Crit. Rev. Food Sci. Nutr. 2018, 58, 924–941. [Google Scholar] [CrossRef]
- Ko, E.-B.; Jang, Y.-G.; Kim, C.-W.; Go, R.-E.; Lee, H.K.; Choi, K.-C. Gallic Acid Hindered Lung Cancer Progression by Inducing Cell Cycle Arrest and Apoptosis in A549 Lung Cancer Cells via PI3K/Akt Pathway. Biomol. Ther. 2021, 30, 151–161. [Google Scholar] [CrossRef]
- Chakrabarti, I.; Ai, W.; Banik, N.L.; Ray, S.K. Overexpression of miR-7-1 Increases Efficacy of Green Tea Polyphenols for Induction of Apoptosis in Human Malignant Neuroblastoma SH-SY5Y and SK-N-DZ Cells. Neurochem. Res. 2012, 38, 420–432. [Google Scholar] [CrossRef]
- Zhang, Y.-Z.; Zhu, R.-W.; Zhong, D.-L.; Zhang, J.-Q. Nunataks or massif de refuge? A phylogeographic study of Rhodiola crenulata (Crassulaceae) on the world’s highest sky islands. BMC Evol. Biol. 2018, 18, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Michels, B.; Franke, K.; Weiglein, A.; Sultani, H.; Gerber, B.; Wessjohann, L.A. Rewarding compounds identified from the medicinal plant Rhodiola rosea. J. Exp. Biol. 2020, 223, b223982. [Google Scholar] [CrossRef] [PubMed]
- Rajizadeh, M.A.; Najafpour, H.; Fekri, M.S.; Rostamzadeh, F.; Jafari, E.; Bejeshk, M.A.; Masoumi-Ardakani, Y. Anti-inflammatory and anti-oxidative effects of Myrtenol in the rats with allergic asthma. Iran. J. Pharm. Res. 2019, 18, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Viljoen, A.M. Geraniol—A review of a commercially important fragrance material. S. Afr. J. Bot. 2010, 76, 643–651. [Google Scholar] [CrossRef]
- Panossian, A.; Wikman, G.; Sarris, J. Rosenroot (Rhodiola rosea): Traditional use, chemical composition, pharmacology and clinical efficacy. Phytomedicine 2010, 17, 481–493. [Google Scholar] [CrossRef] [PubMed]
- Rajan, D.K.; Mohan, K.; Zhang, S.; Ganesan, A.R. Dieckol: A brown algal phlorotannin with biological potential. Biomed. Pharmacother. 2021, 142, 111988. [Google Scholar] [CrossRef]
- Hu, X.; Lin, S.; Yu, D.; Qiu, S.; Zhang, X.; Mei, R. A preliminary study: The anti-proliferation effect of salidroside on different human cancer cell lines. Cell Biol. Toxicol. 2010, 26, 499–507. [Google Scholar] [CrossRef]
- Jiang, S.; Deng, N.; Zheng, B.; Li, T.; Liu, R.H. Rhodiola extract promotes longevity and stress resistance of Caenorhabditis elegans via DAF-16 and SKN-1. Food Funct. 2021, 12, 4471–4483. [Google Scholar] [CrossRef]
- Yang, Q.-Q.; Cheng, L.-Z.; Zhang, T.; Yaron, S.; Jiang, H.-X.; Sui, Z.-Q.; Corke, H. Phenolic profiles, antioxidant, and antiproliferative activities of turmeric (Curcuma longa). Ind. Crops Prod. 2020, 152, 112561. [Google Scholar] [CrossRef]
- Xiang, N.; Li, C.; Li, G.; Yu, Y.; Hu, J.; Guo, X. Comparative Evaluation on Vitamin E and Carotenoid Accumulation in Sweet Corn (Zea mays L.) Seedlings under Temperature Stress. J. Agric. Food Chem. 2019, 67, 9772–9781. [Google Scholar] [CrossRef]
- Xu, Q.; Wang, H.; Li, T.; Chen, L.; Zheng, B.; Liu, R.H. Comparison of phenolics, antioxidant, and antiproliferative activities of two Hypsizygus marmoreus varieties. J. Food Sci. 2020, 85, 2227–2235. [Google Scholar] [CrossRef]
- Yang, J.; Liu, R.H. Synergistic Effect of Apple Extracts and Quercetin 3-β-d-Glucoside Combination on Antiproliferative Activity in MCF-7 Human Breast Cancer Cells in Vitro. J. Agric. Food Chem. 2009, 57, 8581–8586. [Google Scholar] [CrossRef]
- Xie, L.-W.; Cai, S.; Zhao, T.-S.; Li, M.; Tian, Y. Green tea derivative (−)-epigallocatechin-3-gallate (EGCG) confers protection against ionizing radiation-induced intestinal epithelial cell death both in vitro and in vivo. Free Radic. Biol. Med. 2020, 161, 175–186. [Google Scholar] [CrossRef]
- Oshima, T.; Ito, M. Sedative effects of l-menthol, d-camphor, phenylethyl alcohol, and geraniol. J. Nat. Med. 2021, 75, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Sawada, S.; Okano, J.; Imamoto, R.; Yasunaka, Y.; Abe, R.; Koda, M.; Murawaki, Y.; Isomoto, H. Preventive Effect of Ge-raniol on Diethylnitrosamine-Induced Hepatocarcinogenesis in Rats. Yonago Acta Med. 2016, 59, 37–43. [Google Scholar]
- Trytek, M.; Paduch, R.; Pięt, M.; Kozieł, A.; Kandefer-Szerszeń, M.; Szajnecki, Ł.; Gromada, A. Biological activity of oxygenated pinene derivatives on human colon normal and carcinoma cells. Flavour Fragr. J. 2018, 33, 428–437. [Google Scholar] [CrossRef]
- Mączka, W.; Wińska, K.; Grabarczyk, M. One Hundred Faces of Geraniol. Molecules 2020, 25, 3303. [Google Scholar] [CrossRef]
- Yang, M.; Li, R.; Wang, Y.; Tan, J.; Tang, S.-H.; Jiang, Z.-T. Rapid screening of antioxidant bioactive components in blue ginger (Dichorisandra thyrsiflora) essential oil by GC electronic-nose and radical scavenging mechanisms. J. Food Meas. Charact. 2020, 14, 2343–2351. [Google Scholar] [CrossRef]
- Yang, S.-M.; Wang, T.; Wen, D.-G.; Hou, J.-Q.; Li, H.-B. Protective effect of Rhodiola rosea polysaccharides on cryopreserved boar sperm. Carbohydr. Polym. 2016, 135, 44–47. [Google Scholar] [CrossRef]
- Galli, F.; Azzi, A.; Birringer, M.; Cook-Mills, J.M.; Eggersdorfer, M.; Frank, J.; Cruciani, G.; Lorkowski, S.; Özer, N.K. Vitamin E: Emerging aspects and new directions. Free Radic. Biol. Med. 2016, 102, 16–36. [Google Scholar] [CrossRef]
- Yuan, H.; Zhang, J.; Nageswaran, D.; Li, L. Carotenoid metabolism and regulation in horticultural crops. Hortic. Res. 2015, 2, 15036. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids of biotechnological importance. In Biotechnology of Isoprenoids, Advances in Biochemical Engineering/Biotechnology; Schrader, J., Bohlmann, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 148, pp. 449–467. [Google Scholar]
- Radomska-Leśniewska, D.M.; Skopiński, P.; Bałan, B.J.; Bialoszewska, A.; Jóźwiak, J.; Rokicki, D.; Skopińska-Różewska, E.; Borecka, A.; Hevelke, A. Review paper Angiomodulatory properties of Rhodiola spp. and other natural antioxidants. Cent. Eur. J. Immunol. 2015, 2, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Tusevski, O.; Kostovska, A.; Iloska, A.; Trajkovska, L.; Simic, S. Phenolic production and antioxidant properties of some Macedonian medicinal plants. Open Life Sci. 2014, 9, 888–900. [Google Scholar] [CrossRef]
- Saeed, K.M.; You, L.; Chen, C.; Zhao, Z.; Fu, X.; Liu, R.H. Comparative assessment of phytochemical profiles and antioxidant and antiproliferative activities of kiwifruit (Actinidia deliciosa) cultivars. J. Food Biochem. 2019, 43, e13025. [Google Scholar] [CrossRef]
- Xia, W.; Lin, Y.; Gong, E.; Li, T.; Lian, F.; Zheng, B.; Liu, R. Wild pink bayberry fruit: The effect of in vitro gastrointestinal digestion on phytochemical profiles, and antioxidant and antiproliferative activities. Food Funct. 2021, 12, 2126–2136. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- Veeriah, S.; Kautenburger, T.; Habermann, N.; Sauer, J.; Dietrich, H.; Will, F.; Pool-Zobel, B.L. Apple flavonoids inhibit growth of HT29 human colon cancer cells and modulate expression of genes involved in the biotransformation of xenobiotics. Mol. Carcinog. 2006, 45, 164–174. [Google Scholar] [CrossRef]
- Du, G.-J.; Zhang, Z.; Wen, X.-D.; Yu, C.; Calway, T.; Yuan, C.-S.; Wang, C.-Z. Epigallocatechin Gallate (EGCG) Is the Most Effective Cancer Chemopreventive Polyphenol in Green Tea. Nutrients 2012, 4, 1679–1691. [Google Scholar] [CrossRef]
- Chou, T.-C. Drug combination studies and their synergy quantification using the Chou-Talalay method. Cancer Res. 2010, 70, 440–446. [Google Scholar] [CrossRef]
- Li, T.; Kon, N.; Jiang, L.; Tan, M.; Ludwig, T.; Zhao, Y.; Baer, R.; Gu, W. Tumor Suppression in the Absence of p53-Mediated Cell-Cycle Arrest, Apoptosis, and Senescence. Cell 2012, 149, 1269–1283. [Google Scholar] [CrossRef]
- Oakes, V.; Wang, W.; Harrington, B.; Lee, W.J.; Beamish, H.; Chia, K.M.; Pinder, A.; Goto, H.; Inagaki, M.; Pavey, S.; et al. Cyclin A/Cdk2 regulates Cdh1 and claspin during late S/G2 phase of the cell cycle. Cell Cycle 2014, 13, 3302–3311. [Google Scholar] [CrossRef]
- Liu, H.; Li, Z.; Huo, S.; Wei, Q.; Ge, L. Induction of G0/G1 phase arrest and apoptosis by CRISPR/Cas9-mediated knockout of CDK2 in A375 melanocytes. Mol. Clin. Oncol. 2020, 12, 9–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Xu, K.; Gao, F.; Huang, J.; Guan, X. Clinical considerations of CDK4/6 inhibitors in triple-negative breast cancer. Biochim. Biophys. Acta 2021, 1876, 188590. [Google Scholar] [CrossRef] [PubMed]
- Speidel, D. The role of DNA damage responses in p53 biology. Arch. Toxicol. 2015, 89, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Y.; Rahmat, A.; Pihie, A.H.L.; Abdullah, N.R.; Houghton, P.J. Eurycomanone induce apoptosis in HepG2 cells via up-regulation of p53. Cancer Cell Int. 2009, 9, 16. [Google Scholar] [CrossRef] [PubMed]
- D’Orsi, B.; Mateyka, J.; Prehn, J.H. Control of mitochondrial physiology and cell death by the Bcl-2 family proteins Bax and Bok. Neurochem. Int. 2017, 109, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Qi, L.; Li, L.; Li, Y. The caspase-3/GSDME signal pathway as a switch between apoptosis and pyroptosis in cancer. Cell Death Discov. 2020, 6, 1–11. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenol | |
|---|---|
| Free polyphenol (mg GAE/g FW) | 120.13 ± 2.88 |
| Bound polyphenol (mg GAE/g FW) | 2.58 ± 0.22 |
| Flavonoids | |
| Free flavonoids (mg CE/g FW) | 125.71 ± 6.06 |
| Bound flavonoids (mg CE/g FW) | 3.28 ± 0.24 |
| GA (mg/100 g FW) | 319.17 ± 41.18 |
| EGC (mg/100 g FW) | 267.04 ± 16.09 |
| EGCG (mg/100 g FW) | 722.10 ± 54.26 |
| Catechin (mg/100 g FW) | 35.93 ± 7.39 |
| Compound Name | Contents (ng/100 g FW) |
|---|---|
| 1-Hexanol | 105.51 ± 4.13 |
| Acetic acid | 514.48 ± 6.77 |
| Benzaldehyde | 68.36 ± 19.21 |
| 1-Octanol | 848.14 ± 19.60 |
| Myrtenal | 90.59 ± 4.51 |
| L-α-Terpineol | 67.14 ± 4.40 |
| 2-Methyl-2-butenolide | 326.18 ± 2.95 |
| Pentanoic acid | 52.72 ± 13.02 |
| (-)-Myrtenol | 1477.80 ± 54.76 |
| Phenethyl acetate | 125.98 ± 11.56 |
| Hexanoic acid | 293.22 ± 10.18 |
| Geraniol | 550.87 ± 21.27 |
| Benzyl alcohol | 416.38 ± 11.03 |
| Phenylethyl alcohol | 7869.33 ± 174.51 |
| 2-(Hydroxymethyl) but-2-enenitrile | 67.66 ± 9.45 |
| Octanoic acid | 99.44 ± 7.55 |
| Compounds (Vitamin E) | Contents (μg/100 g FW) |
|---|---|
| α-Tocopherol | 338.51 ± 17.75 |
| α-Tocotrienol | 16.82 ± 0.47 |
| β-Tocotrienol | 70.47 ± 0.78 |
| γ-Tocopherols | 19.88 ± 1.32 |
| γ-Tocotrienol | 29.38 ± 3.44 |
| δ-Tocotrienol | 27.89 ± 0.53 |
| Total | 502.94 ± 17.41 |
| Compounds (Carotenoids) | Contents (μg/100 g FW) |
| Lutein | 3.48 ± 0.11 |
| β-Carotene | 3.95 ± 0.40 |
| Total | 7.43 ± 0.72 |
| Fractions | EC50 (µg/mL) | CAA Quality (µmol of QE/100 µmol of Polyphenols) | CAA Quality (µmol of QE/100 µmol of Flavonoids) | |||
|---|---|---|---|---|---|---|
| PBS Wash | No PBS Wash | PBS Wash | No PBS Wash | PBS Wash | No PBS Wash | |
| RFE | 118.49 ± 7.58 | 56.09 ± 0.34 | 10.51 ± 0.25 | 14.71 ± 0.35 | 15.51 ± 0.67 | 21.74 ± 0.94 |
| RBE | 10,048.57 ± 131.91 | 3018.09 ± 133.12 | 0.10 ± 0.00 | 0.15 ± 0.01 | 0.30 ± 0.01 | 0.44 ± 0.02 |
| Total | - | - | 10.64 ± 0.25 | 15.01 ± 0.36 | 15.67 ± 0.68 | 22.15 ± 0.96 |
| EGCG | 176.09 ± 9.83 | 52.31 ± 2.55 | 0.96 ± 0.04 | 1.64 ± 0.07 | 2.15 ± 0.09 | 3.66 ± 0.15 |
| Samples | EC50 (μg/mL) | CC50 (μg/mL) | SI (CC50/EC50) |
|---|---|---|---|
| RFE | 276.32 ± 8.47 | >1400 | >2 |
| Samples | EC50 (μM) | CC50 (μM) | SI (CC50/EC50) |
| GA | 73.91 ± 3.23 | >600 | >2 |
| EGC | 107.21 ± 4.64 | >600 | >2 |
| EGCG | 61.66 ± 7.97 | >500 | >2 |
| Concentration | Inhibitory Effects of Combinations (%) | |||
|---|---|---|---|---|
| GA + EGC | GA + EGCG | EGC + EGCG | GA + EGC + EGCG (2/3) | |
| 0.125 × EC50 | 18.97 ± 3.28 | 48.74 ± 3.18 | 33.81 ± 2.76 | 30.58 ± 5.34 |
| 0.25 × EC50 | 32.29 ± 7.09 | 47.97 ± 9.53 | 54.22 ± 5.66 | 37.18 ± 4.55 |
| 0.50 × EC50 | 53.57 ± 3.86 | 55.79 ± 2.05 | 57.56 ± 10.95 | 47.22 ± 3.89 |
| 0.75 × EC50 | 63.37 ± 9.51 | 72.09 ± 5.75 | 70.04 ± 0.92 | 57.69 ± 2.67 |
| 1.0 × EC50 | 68.80 ± 1.31 | 67.38 ± 3.14 | 75.63 ± 3.13 | 61.80 ± 0.43 |
| 1.25 × EC50 | 78.67 ± 1.60 | 77.40 ± 1.78 | 75.81 ± 4.24 | 67.89 ± 0.59 |
| Compound | Range of DRI A | |||
|---|---|---|---|---|
| GA + EGC | GA + EGCG | EGC + EGCG | GA + EGC + EGCG | |
| GA | 1.18 ± 0.18–4.23 ± 0.91 | 0.72 ± 0.12–34.48 ± 5.51 | - B | 0.72 ± 0.09–14.26 ± 4.97 |
| EGC | 1.77 ± 0.27–3.15 ± 0.68 | - | 1.65 ± 0.67–7.43 ± 1.62 | 1.08 ± 0.13–10.63 ± 3.70 |
| EGCG | - | 1.21 ± 0.07–14.24 ± 0.95 | 2.00 ± 0.81–4.41 ± 0.96 | 1.20 ± 0.04–7.16 ± 1.03 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Jiang, S.; Deng, N.; Zheng, B.; Li, T.; Liu, R.H. Phytochemical Profiles, Antioxidant Activity and Antiproliferative Mechanism of Rhodiola rosea L. Phenolic Extract. Nutrients 2022, 14, 3602. https://doi.org/10.3390/nu14173602
Zhang S, Jiang S, Deng N, Zheng B, Li T, Liu RH. Phytochemical Profiles, Antioxidant Activity and Antiproliferative Mechanism of Rhodiola rosea L. Phenolic Extract. Nutrients. 2022; 14(17):3602. https://doi.org/10.3390/nu14173602
Chicago/Turabian StyleZhang, Sheng, Siqi Jiang, Na Deng, Bisheng Zheng, Tong Li, and Rui Hai Liu. 2022. "Phytochemical Profiles, Antioxidant Activity and Antiproliferative Mechanism of Rhodiola rosea L. Phenolic Extract" Nutrients 14, no. 17: 3602. https://doi.org/10.3390/nu14173602
APA StyleZhang, S., Jiang, S., Deng, N., Zheng, B., Li, T., & Liu, R. H. (2022). Phytochemical Profiles, Antioxidant Activity and Antiproliferative Mechanism of Rhodiola rosea L. Phenolic Extract. Nutrients, 14(17), 3602. https://doi.org/10.3390/nu14173602