

Crossbred Sows Fed a Western Diet during Pre-Gestation, Gestation, Lactation, and Post-Lactation Periods Develop Signs of Lean Metabolic Syndrome That Are Partially Attenuated by Spirulina Supplementation

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
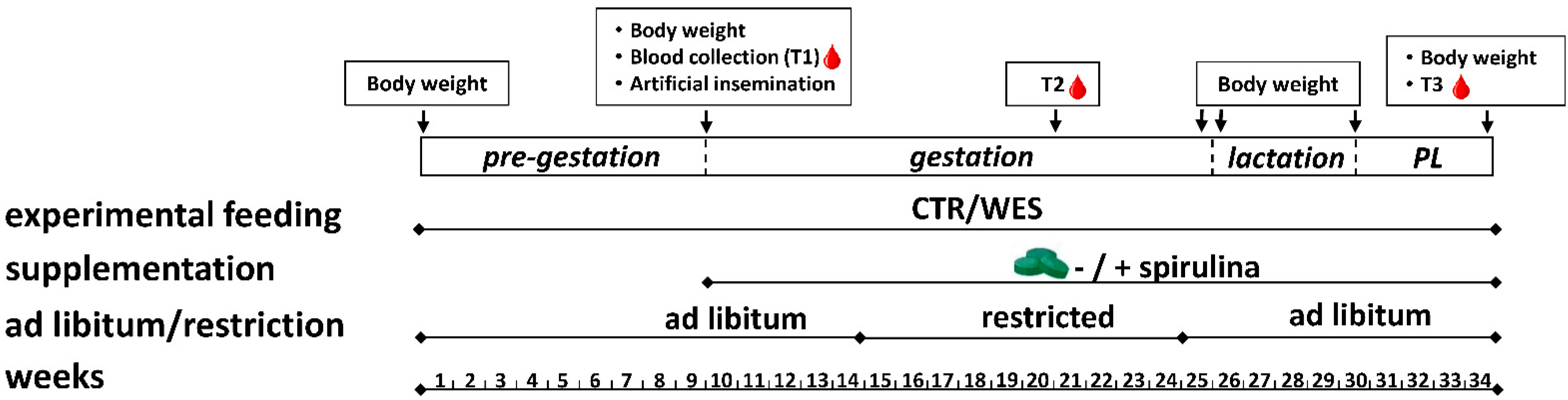
2.1. Animals, Housing, and Experimental Design
2.2. Experimental Diets
2.3. Data and Sample Collection during the Feeding Experiment
2.4. Organ Collection at Slaughter
2.5. Isolation and Quality Assessment of Liver and Muscle RNA
2.6. Transcriptome Analyses
2.7. Validation of Transcriptome Data via qPCR
2.8. Compositional Analyses of Diets, Liver, Skeletal Muscle, and Feces
2.9. Histological Analyses of Liver, Skeletal Muscle, and Visceral Adipose Tissue
2.10. Blood Collection
2.11. Serum and Plasma Assays
2.12. Fatty Acid Analyses in Diets and Tissues
2.13. Statistical Analyses
3. Results
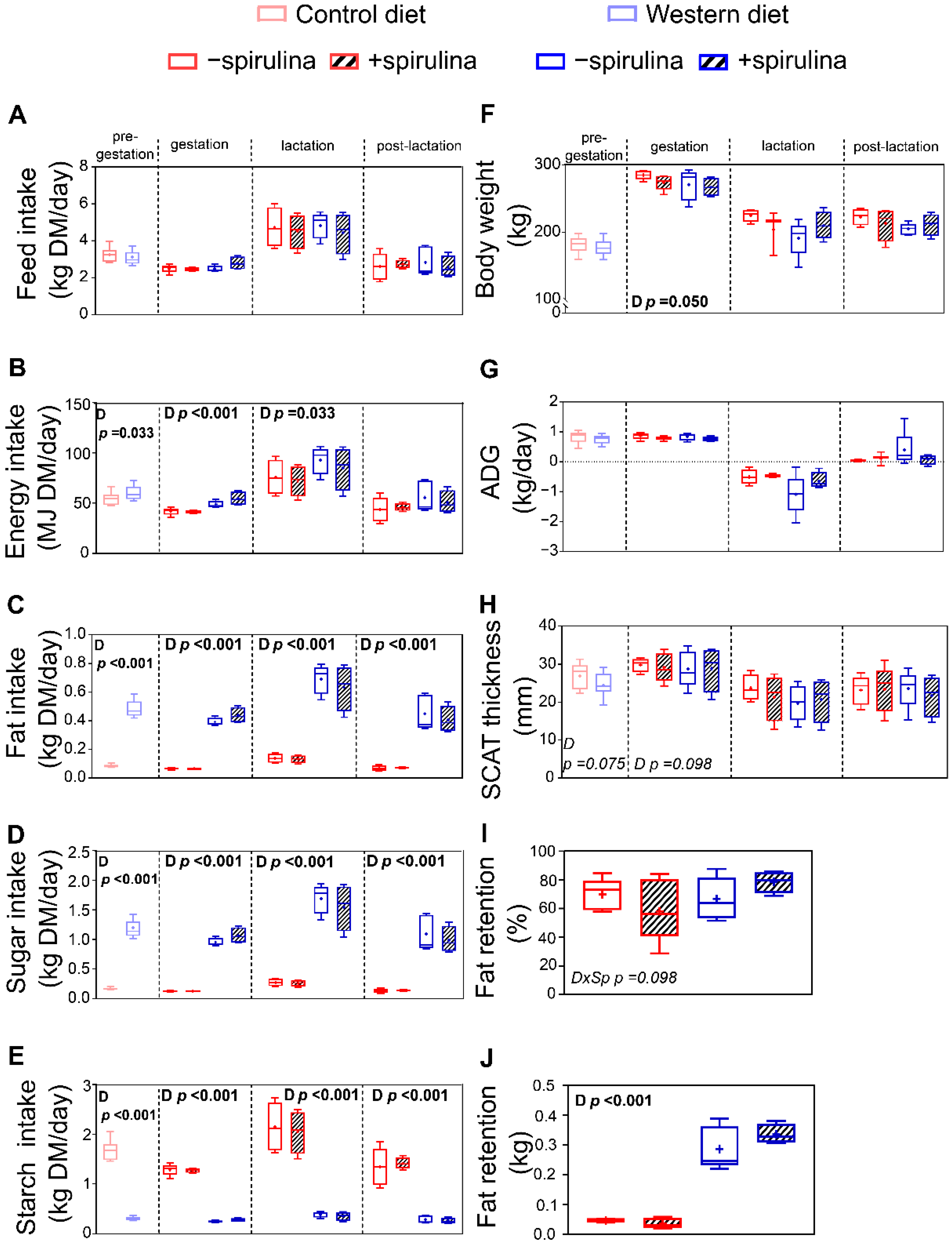
3.1. Dietary Intake and Growth Performance
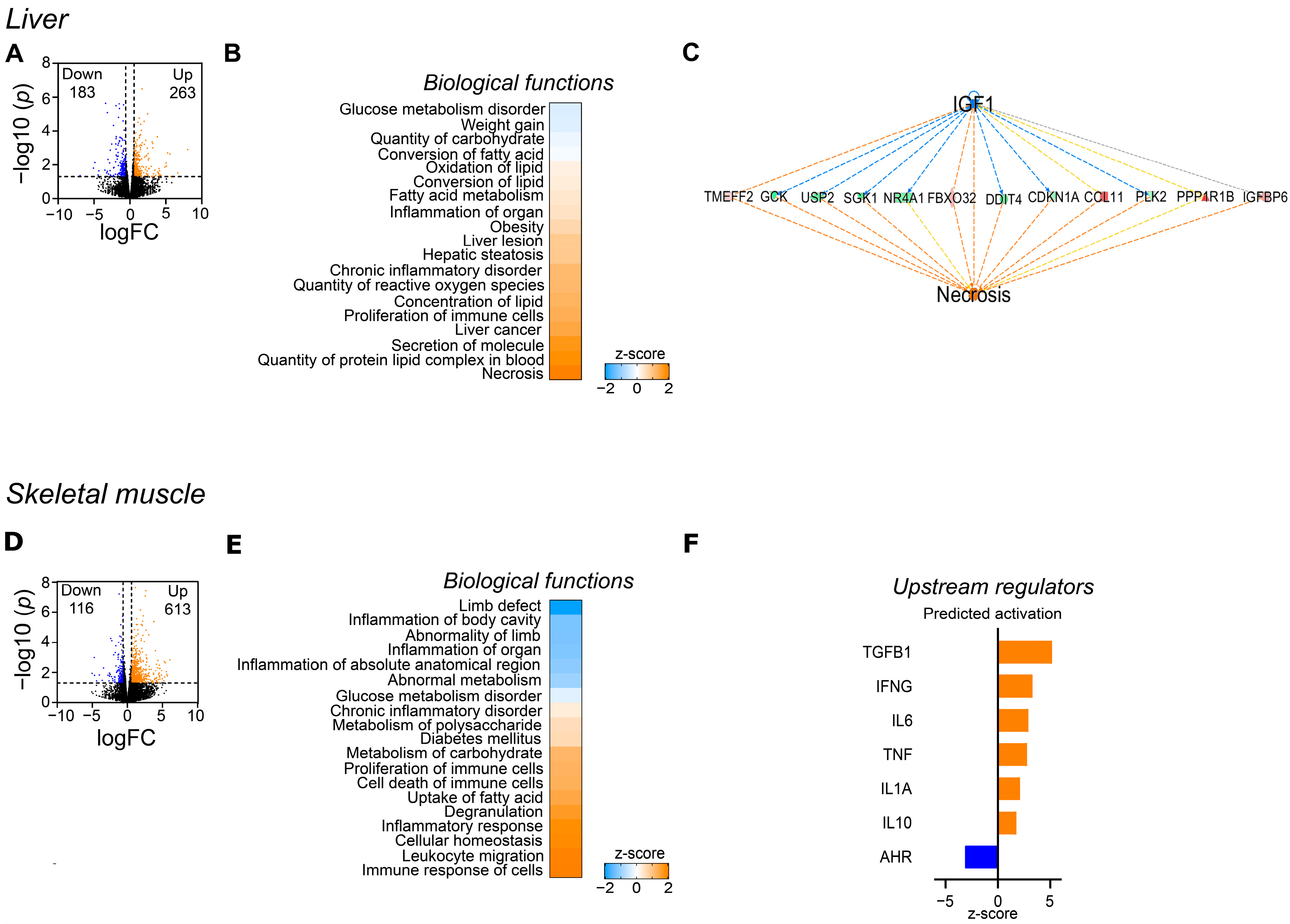
3.2. Differential Impact of WES Diet on Hepatic and Skeletal Muscle Transcriptome
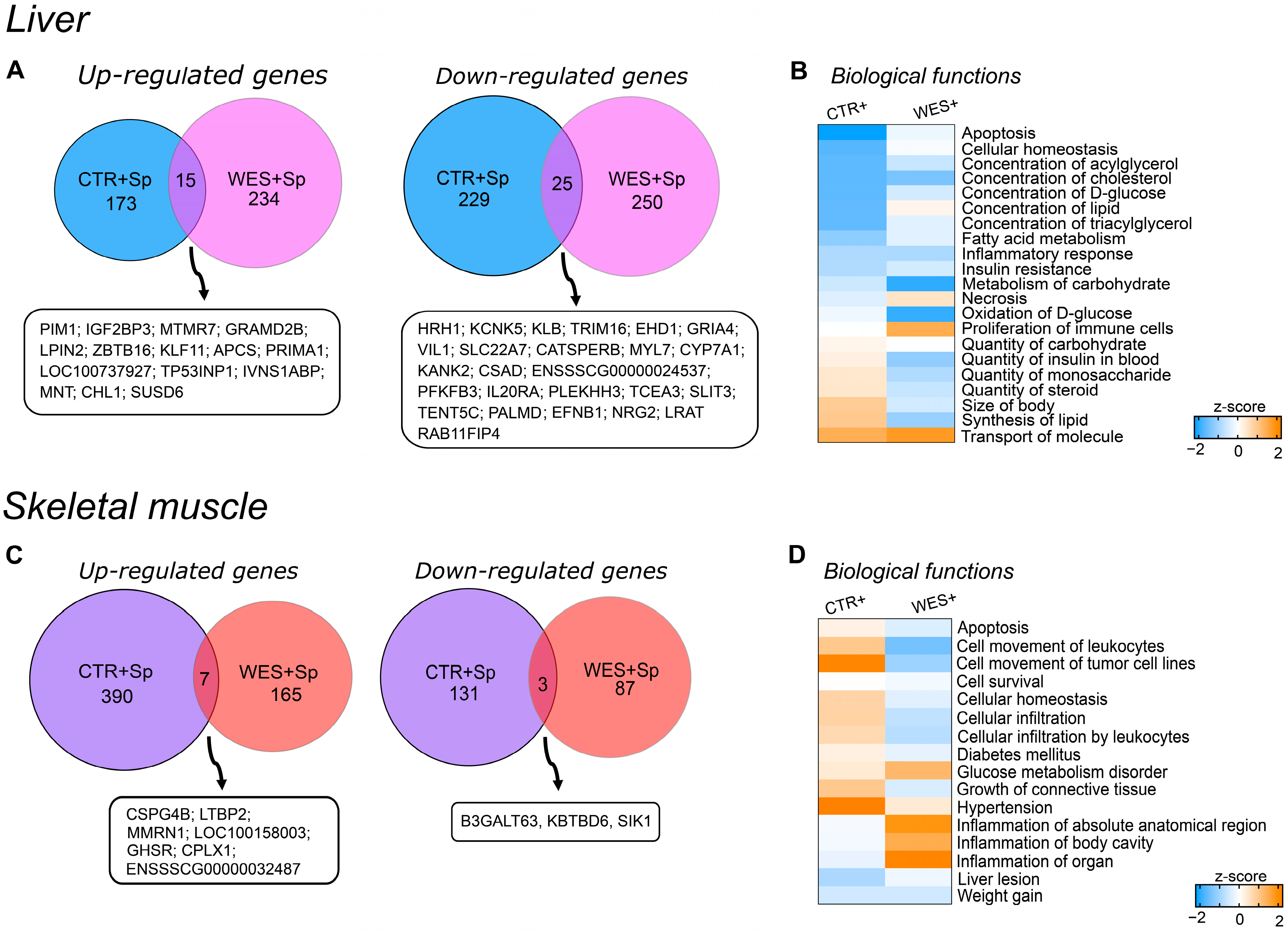
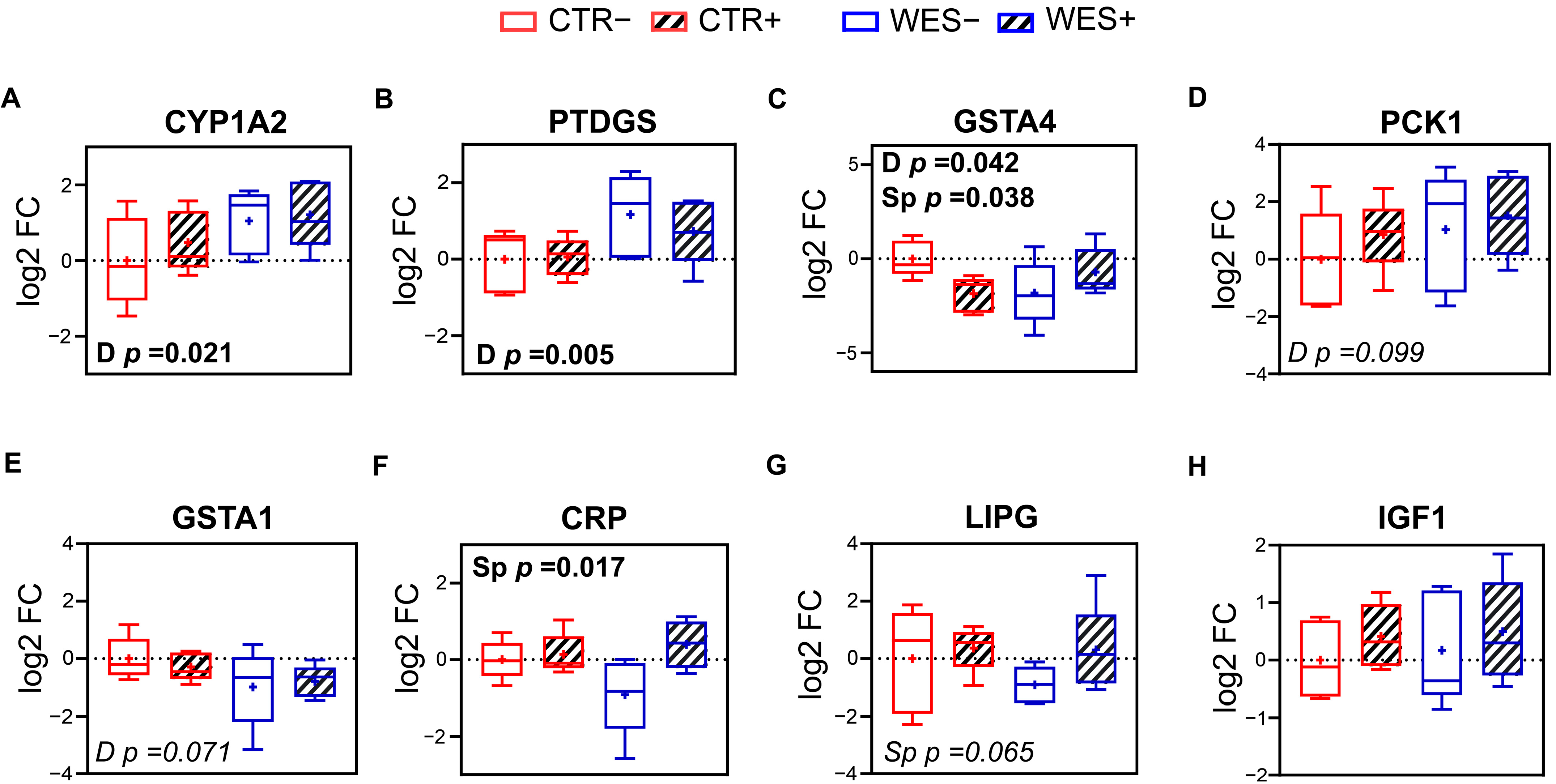
3.3. Changes in Gene Expression Influenced by Spirulina
3.4. Validation of RNAseq by qPCR
3.5. Organ Proportions, Composition, and Histology
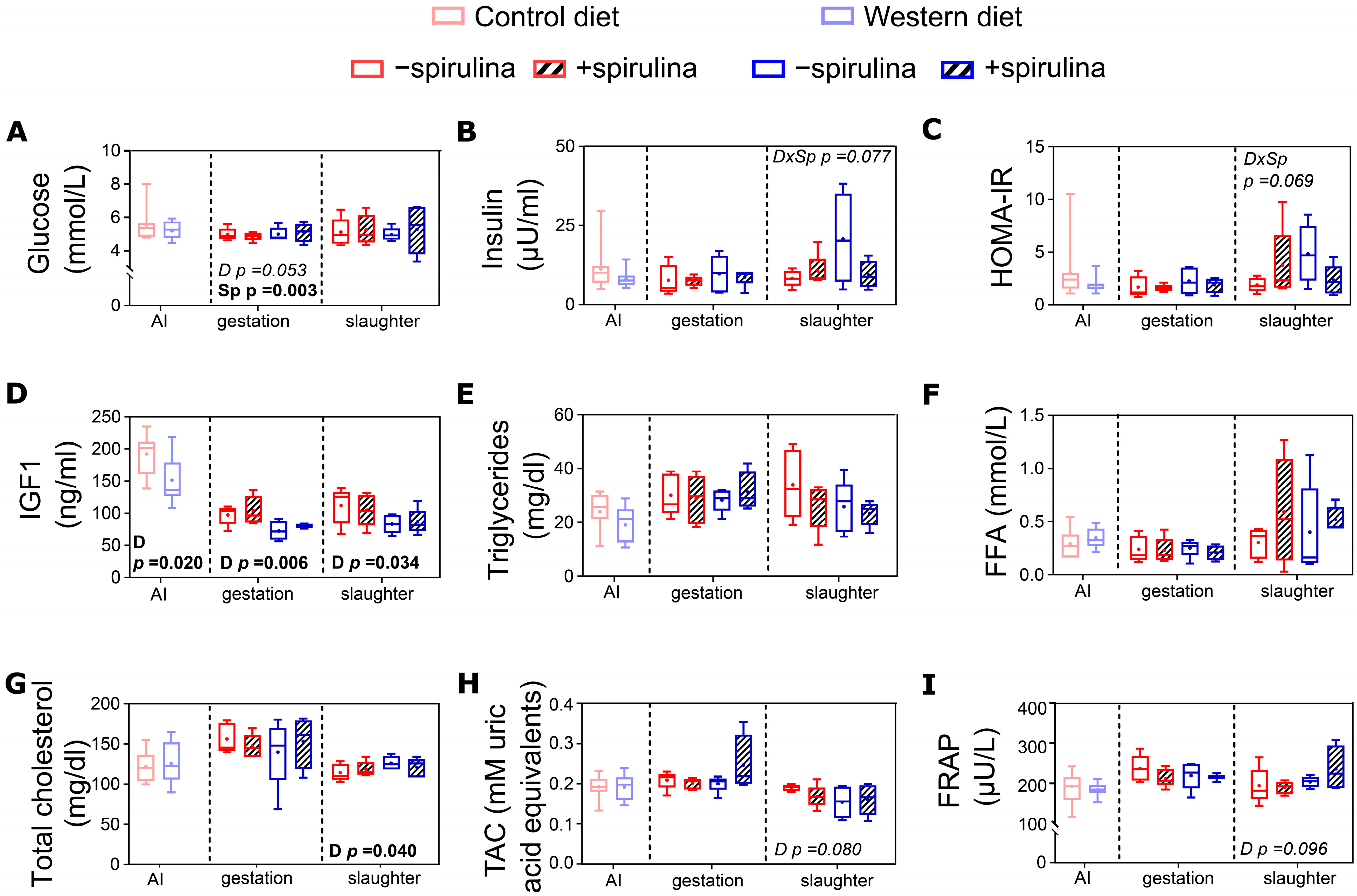
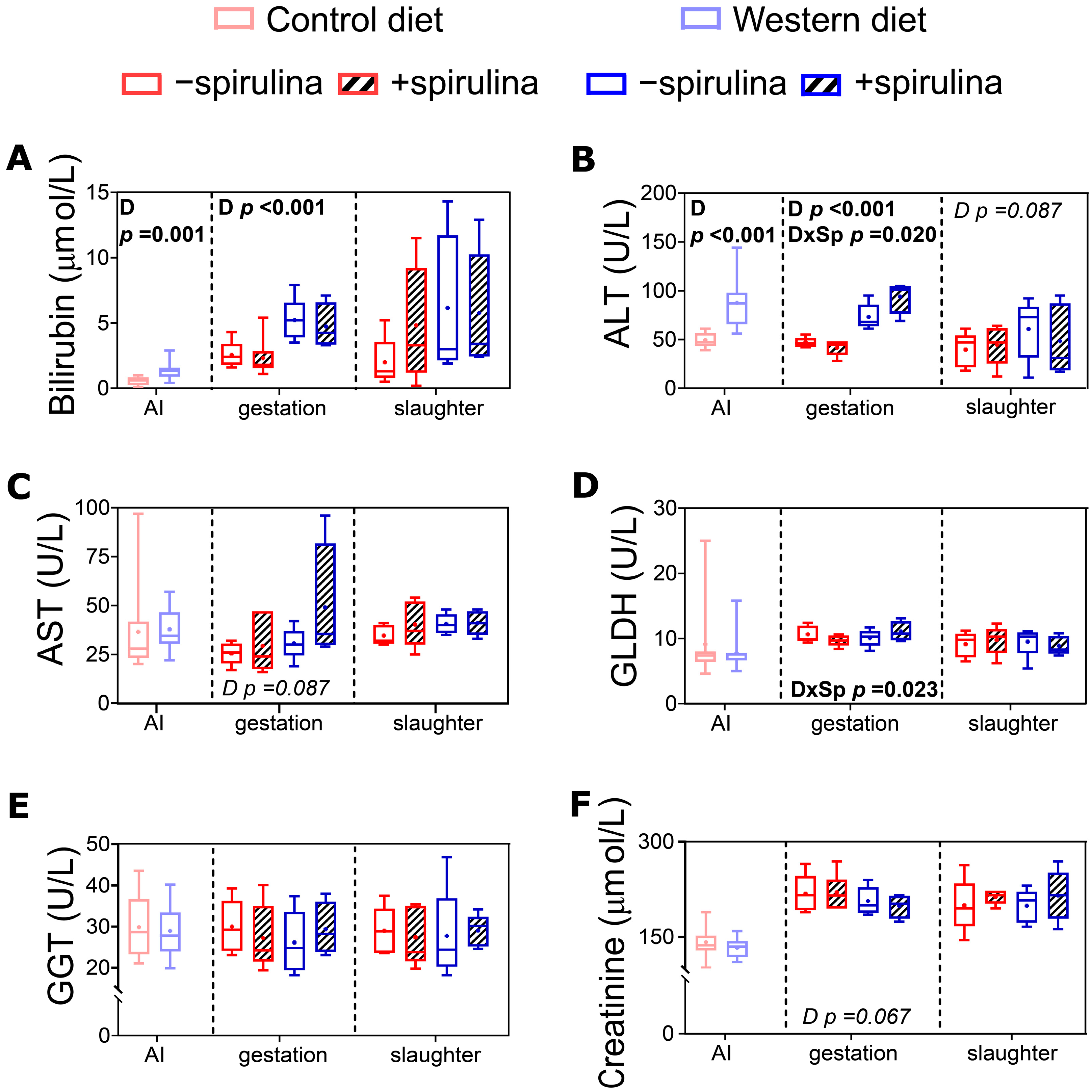
3.6. Blood Biochemical Biomarkers
3.7. Fatty Acid Composition of Liver and Skeletal Muscle
4. Discussion
4.1. Effects of a Western Diet on the Metabolic Phenotype of Female Domestic Pigs
4.1.1. Western Diet Intake Induced Regulatory Processes to Maintain Body Weight and Visceral Adipose Tissue Proportion
4.1.2. Liver Functionality Was Adversely Affected by the Western Diet
4.1.3. The Impaired Glucose Metabolism Induced by the Western Diet Was Likely Mediated by Metabolic Changes Predominantly in the Skeletal Muscle
4.2. Effects of Spirulina Supplementation on the Metabolic Phenotype of Female Domestic Pigs
4.2.1. Spirulina Supplementation Did Not Affect Body Weight but Potentially Aggravated WD-Induced Liver Damage
4.2.2. Spirulina Supplementation Had Only Small Diet-Dependent Effects on Systemic Metabolic Biomarkers
4.2.3. Spirulina Supplementation Had Only Minor Effects on the Tissue Fatty Acid Profiles
4.3. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Population Review. Most Obese Countries. Available online: https://worldpopulationreview.com/country-rankings/most-obese-countries (accessed on 1 October 2021).
- Tovar, A.R.; Díaz-Villaseñor, A.; Cruz-Salazar, N.; Ordáz, G.; Granados, O.; Palacios-González, B.; Tovar-Palacio, C.; López, P.; Torres, N. Dietary Type and Amount of Fat Modulate Lipid Metabolism Gene Expression in Liver and in Adipose Tissue in High-fat Diet-fed Rats. Arch. Med Res. 2011, 42, 540–553. [Google Scholar] [CrossRef] [PubMed]
- Parks, E.J.; Skokan, L.E.; Timlin, M.T.; Dingfelder, C.S. Dietary Sugars Stimulate Fatty Acid Synthesis in Adults. J. Nutr. 2008, 138, 1039–1046. [Google Scholar] [CrossRef]
- Gabbia, D.; Roverso, M.; Guido, M.; Sacchi, D.; Scaffidi, M.; Carrara, M.; Orso, G.; Russo, F.P.; Floreani, A.; Bogialli, S.; et al. Western Diet-Induced Metabolic Alterations Affect Circulating Markers of Liver Function before the Development of Steatosis. Nutrients 2019, 11, 1602. [Google Scholar] [CrossRef]
- Barnard, R.J.; Roberts, C.K.; Varon, S.M.; Berger, J.J. Diet-induced insulin resistance precedes other aspects of the metabolic syndrome. J. Appl. Physiol. 1998, 84, 1311–1315. [Google Scholar] [CrossRef] [PubMed]
- Misra, B.B.; Upadhayay, R.P.; Mattern, V.; Parks, J.S.; Cox, L.A.; Comuzzie, A.G.; Olivier, M. Metabolic programming: Short term western diet exposure induces sustained changes in plasma metabolites. bioRXiv 2019. [Google Scholar] [CrossRef]
- Shi, Z.; Wu, X.; Yu, S.; Huynh, M.; Jena, P.K.; Nguyen, M.; Wan, Y.Y.; Hwang, S.T. Short-term exposure to a Western diet induces psoriasiform dermatitis by promoting accumulation of IL-17A-producing γδ T cells. J. Investig. Dermatol. 2020, 140, 1815–1823. [Google Scholar] [CrossRef] [PubMed]
- Simon, A.; Pratt, M.; Hutton, B.; Skidmore, B.; Fakhraei, R.; Rybak, N.; Corsi, D.J.; Walker, M.; Velez, M.P.; Smith, G.N.; et al. Guidelines for the management of pregnant women with obesity: A systematic review. Obes. Rev. 2019, 21, e12972. [Google Scholar] [CrossRef]
- Butte, N.F. Carbohydrate and lipid metabolism in pregnancy: Normal compared with gestational diabetes mellitus. Am. J. Clin. Nutr. 2000, 71, 1256S–1261S. [Google Scholar] [CrossRef]
- Hyatt, H.; Zhang, Y.; Hood, W.R.; Kavazis, A.N. Lactation has persistent effects on a mother’s metabolism and mitochondrial function. Sci. Rep. 2017, 7, 17118. [Google Scholar] [CrossRef]
- Musial, B.; Vaughan, O.; Twinn, D.; Voshol, P.; Ozanne, S.; Fowden, A.; Sferruzzi-Perri, A. A Western-style obesogenic diet alters maternal metabolic physiology with consequences for fetal nutrient acquisition in mice. J. Physiol. 2017, 14, 4875–4892. [Google Scholar] [CrossRef] [Green Version]
- Holman, B.W.B.; Malau-Aduli, A.E.O. Spirulina as a livestock supplement and animal feed. J. Anim. Physiol. Anim. Nutr. 2013, 97, 615–623. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Liu, L.; Miron, A.; Klímová, B.; Wan, D.; Kuča, K. The antioxidant, immunomodulatory, and anti-inflammatory activities of Spirulina: An overview. Arch. Toxicol. 2016, 90, 1817–1840. [Google Scholar] [CrossRef] [PubMed]
- Coué, M.; Tesse, A.; Falewée, J.; Aguesse, A.; Croyal, M.; Fizanne, L.; Chaigneau, J.; Boursier, J.; Ouguerram, K. Spirulina Liquid Extract Protects against Fibrosis Related to Non-Alcoholic Steatohepatitis and Increases Ursodeoxycholic Acid. Nutrients 2019, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Heo, M.-G.; Choung, S.-Y. Anti-obesity effects of Spirulina maxima in high fat diet induced obese rats via the activation of AMPK pathway and SIRT1. Food Funct. 2018, 9, 4906–4915. [Google Scholar] [CrossRef]
- Yang, Y.; Du, L.; Hosokawa, M.; Miyashita, K. Spirulina Lipids Alleviate Oxidative Stress and Inflammation in Mice Fed a High-Fat and High-Sucrose Diet. Mar. Drugs 2020, 18, 148. [Google Scholar] [CrossRef]
- Deng, R.; Chow, T.J. Hypolipidemic, Antioxidant, and Anti-inflammatory Activities of Microalgae Spirulina. Cardiovasc. Ther. 2010, 28, e33–e45. [Google Scholar] [CrossRef]
- Koopmans, S.J.; Schuurman, T. Considerations on pig models for appetite, metabolic syndrome and obese type 2 diabetes: From food intake to metabolic disease. Eur. J. Pharmacol. 2015, 759, 231–239. [Google Scholar] [CrossRef]
- Mathias, P.C.F.; Elmhiri, G.; de Oliveira, J.; Delayre-Orthez, C.; Barella, L.F.; Tófolo, L.P.; Fabricio, G.S.; Chango, A.; Abdennebi-Najar, L. Maternal diet, bioactive molecules, and exercising as reprogramming tools of metabolic programming. Eur. J. Nutr. 2014, 53, 711–722. [Google Scholar] [CrossRef]
- Pruis, M.G.M.; Lendvai, A.; Bloks, V.W.; Zwier, M.V.; Baller, J.F.W.; de Bruin, A.; Groen, A.K.; Plösch, T. Maternal western diet primes non-alcoholic fatty liver disease in adult mouse offspring. Acta Physiol. 2014, 210, 215–227. [Google Scholar] [CrossRef]
- Kleinert, M.; Clemmensen, C.; Hofmann, S.; Moore, M.C.; Renner, S.; Woods, S.C.; Huypens, P.; Beckers, J.; de Angelis, M.H.; Schürmann, A.; et al. Animal models of obesity and diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 140–162. [Google Scholar] [CrossRef] [Green Version]
- Merrifiel, C.A.; Lewis, M.; Claus, S.P.; Beckonert, O.P.; Dumas, M.E.; Duncker, S.; Kochhar, S.; Rezzi, S.; Lindon, J.C.; Bailey, M.; et al. A metabolic system-wide characterization of the pig: A model for human physiology. Mol. Bio. Syst. 2011, 7, 2577–2588. [Google Scholar]
- Bergen, W.G.; Mersmann, H.J. Comparative Aspects of Lipid Metabolism: Impact on Contemporary Research and Use of Animal Models. J. Nutr. 2005, 135, 2499–2502. [Google Scholar] [CrossRef] [PubMed]
- Litten-Brown, J.C.; Corson, A.M.; Clarke, L. Porcine models for the metabolic syndrome, digestive and bone disorders: A general overview. Animals 2010, 4, 899–920. [Google Scholar] [CrossRef] [PubMed]
- Lugarà, R.; Grześkowiak, Ł.; Zentek, J.; Meese, S.; Kreuzer, M.; Giller, K. A High-Energy Diet and Spirulina Supplementation during Pre-Gestation, Gestation, and Lactation do Not Affect the Reproductive and Lactational Performance of Primiparous Sows. Animals 2022, 12, 1171. [Google Scholar] [CrossRef] [PubMed]
- Affentranger, P.; Gerwig, C.; Seewer, G.; Schwörer, D.; Künzi, N. Growth and carcass characteristics as well as meat and fat quality of three types of pigs under different feeding regimens. Livest. Prod. Sci. 1996, 45, 187–196. [Google Scholar] [CrossRef]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. The Subread aligner: Fast, accurate and scalable read mapping by seed-and-vote. Nucleic Acids Res. 2013, 41, e108. [Google Scholar] [CrossRef]
- Xie, F.; Xiao, P.; Chen, D.; Xu, L.; Zhang, B. miRDeepFinder: A miRNA analysis tool for deep sequencing of plant small RNAs. Plant Mol. Biol. 2012, 80, 75–84. [Google Scholar] [CrossRef]
- Madzlan, K. Determination of cholesterol in several types of eggs by gas chromatography. J. Trop. Agric. Food Sci. 2008, 36, 205–210. [Google Scholar]
- Lan, F.; Misu, H.; Chikamoto, K.; Takayama, H.; Kikuchi, A.; Mohri, K.; Takata, N.; Hayashi, H.; Matsuzawa-Nagata, N.; Takeshita, Y.; et al. LECT2 Functions as a Hepatokine That Links Obesity to Skeletal Muscle Insulin Resistance. Diabetes 2014, 63, 1649–1664. [Google Scholar] [CrossRef]
- Vicari, T.; Borne, J.V.D.; Gerrits, W.; Zbinden, Y.; Blum, J. Postprandial blood hormone and metabolite concentrations influenced by feeding frequency and feeding level in veal calves. Domest. Anim. Endocrinol. 2008, 34, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P. Homeostasis model assessment: Insulin resistance and β-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, C.; Ulbrich, S.; Kreuzer, M.; Berard, J.; Giller, K. Differential partitioning of rumen-protected n–3 and n–6 fatty acids into muscles with different metabolism. Meat Sci. 2018, 137, 106–113. [Google Scholar] [CrossRef]
- Collomb, M.; Bühler, T. Analyse de la composition en acides gras de la graisse de lait. Mitt. Lebensm. Hyg. 2000, 91, 306–332. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 19 April 2022).
- Xi, S.; Yin, W.; Wang, Z.; Kusunoki, M.; Lian, X.; Koike, T.; Fan, J.; Zhang, Q. A minipig model of high-fat/high-sucrose diet-induced diabetes and atherosclerosis. Int. J. Exp. Pathol. 2004, 85, 223–231. [Google Scholar] [CrossRef]
- Sébert, S.P.; Lecannu, G.; Sené, S.; Hucteau, S.; Chetiveaux, M.; Ouguerram, K.; Champ, M.M.-J. Obesity induced during sexual maturation is linked to LDL-triacylglycerols in Yucatan miniature swine. Br. J. Nutr. 2005, 94, 282–289. [Google Scholar] [CrossRef]
- Dyson, M.C.; Alloosh, M.; Vuchetich, J.P.; A Mokelke, E.; Sturek, M. Components of metabolic syndrome and coronary artery disease in female Ossabaw swine fed excess atherogenic diet. Comp. Med. 2006, 56, 35–45. [Google Scholar]
- Christoffersen, B.O.; Golozoubova, V.; Pacini, G.; Svendsen, O.; Raun, K. The young göttingen minipig as a model of childhood and adolescent obesity: Influence of diet and gender. Obesity 2007, 21, 149–158. [Google Scholar] [CrossRef] [PubMed]
- Newell-Fugate, A.E.; Taibl, J.N.; Alloosh, M.; Sturek, M.; Bahr, J.M.; Nowak, R.A.; Krisher, R.L. Effects of Obesity and Metabolic Syndrome on Steroidogenesis and Folliculogenesis in the Female Ossabaw Mini-Pig. PLoS ONE 2015, 10, e0128749. [Google Scholar] [CrossRef]
- Yang, X.; Ma, X.; Wang, L.; Gao, K.; Jiang, Z. 454 A high-fat diet expands body fat mass and up-regulates expression of genes involved in adipogenesis and inflammation in a genetically lean pig. J. Anim. Sci. 2018, 95, 223. [Google Scholar] [CrossRef]
- Renner, S.; Blutke, A.; Dobenecker, B.; Dhom, G.; Müller, T.D.; Finan, B.; Clemmensen, C.; Bernau, M.; Novak, I.; Rathkolb, B.; et al. Metabolic syndrome and extensive adipose tissue inflammation in morbidly obese Göttingen minipigs. Mol. Metab. 2018, 16, 180–190. [Google Scholar] [CrossRef]
- Goodpaster, B.H.; Sparks, L.M. Metabolic Flexibility in Health and Disease. Cell Metab. 2017, 25, 1027–1036. [Google Scholar] [CrossRef] [PubMed]
- Buscemi, S.; Chiarello, P.; Buscemi, C.; Corleo, D.; Massenti, M.F.; Barile, A.M.; Rosafio, G.; Maniaci, V.; Settipani, V.; Cosentino, L.; et al. Characterization of Metabolically Healthy Obese People and Metabolically Unhealthy Normal-Weight People in a General Population Cohort of the ABCD Study. J. Diabetes Res. 2017, 2017, 9294038. [Google Scholar] [CrossRef] [PubMed]
- Hamer, M.; Stamatakis, E. Metabolically Healthy Obesity and Risk of All-Cause and Cardiovascular Disease Mortality. J. Clin. Endocrinol. Metab. 2012, 97, 2482–2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefan, N.; Schick, F.; Häring, H.-U. Causes, Characteristics, and Consequences of Metabolically Unhealthy Normal Weight in Humans. Cell Metab. 2017, 26, 292–300. [Google Scholar] [CrossRef] [PubMed]
- Salvaggio, A.; Periti, M.; Miano, L.; Tavanelli, M.; Marzorati, D. Body mass index and liver enzyme activity in serum. Clin. Chem. 1991, 37, 720–723. [Google Scholar] [CrossRef]
- Duval, C.; Thissen, U.; Keshtkar, S.; Accart, B.; Stienstra, R.; Boekschoten, M.V.; Roskams, T.; Kersten, S.; Müller, M. Adipose tissue dysfunction signals progression of hepatic steatosis towards non-alcoholic steatohepatitis in C57Bl/6 mice. Diabetes 2010, 59, 3181–31919. [Google Scholar] [CrossRef]
- Hjelkrem, M.; Morales, A.; Williams, C.D.; Harrison, S.A. Unconjugated hyperbilirubinemia is inversely associated with non-alcoholic steatohepatitis (NASH). Aliment. Pharmacol. Ther. 2012, 35, 1416–1423. [Google Scholar] [CrossRef]
- Bril, F.; McPhaul, M.J.; Caufield, M.P.; Clark, V.C.; Soldevilla-Pico, C.; Firpi-Morell, R.J.; Lai, J.; Shiffman, D.; Rowland, C.M.; Cusi, K. Performance of plasma biomarkers and diagnostic panels for non-alcoholic steatohepatitis and advanced fibrosis in patients with type 2 diabetes. Diabetes Care 2020, 43, 290–297. [Google Scholar] [CrossRef]
- Stuebe, A.S.; Rich-Edwards, J.W. The reset hypothesis: Lactation and maternal metabolism. Am. J. Perinatol. 2010, 26, 81–88. [Google Scholar] [CrossRef]
- Paczkowska-Abdulsalam, M.; Niemira, M.; Bielska, A.; Szałkowska, A.; Raczkowska, B.A.; Junttila, S.; Gyenesei, A.; Adamska-Patruno, E.; Maliszewska, K.; Citko, A.; et al. Evaluation of Transcriptomic Regulations behind Metabolic Syndrome in Obese and Lean Subjects. Int. J. Mol. Sci. 2020, 21, 1455. [Google Scholar] [CrossRef]
- Biobaku, F.; Ghanim, H.; Batra, M.; Dandona, P. Macronutrient-Mediated Inflammation and Oxidative Stress: Relevance to Insulin Resistance, Obesity, and Atherogenesis. J. Clin. Endocrinol. Metab. 2019, 104, 6118–6128. [Google Scholar] [CrossRef]
- Frova, C. Glutathione transferases in the genomics era: New insights and perspectives. Biomol. Eng. 2006, 23, 149–169. [Google Scholar] [CrossRef]
- Giannini, E.; Risso, D.; Ceppa, P.; Botta, F.; Chiarbonello, B.; Fasoli, A.; Malfatti, F.; Romagnoli, P.; Lantieri, P.B.; Testa, R. Utility of α-Glutathione S-transferase assessment in chronic hepatitis C patients with near normal alanine aminotransferase levels. Clin. Biochem. 2000, 33, 297–301. [Google Scholar] [CrossRef]
- Mavis, C.K.; Kinney, S.R.M.; Foster, B.A.; Karpf, A.R. Expression level and DNA methylation status of glutathione-S-transferase genes in normal murine prostate and TRAMP tumors. Prostate 2009, 69, 1312–1324. [Google Scholar] [CrossRef]
- Bessone, F.; Razori, M.V.; Roma, M.G. Molecular pathways of nonalcoholic fatty liver disease development and progression. Cell. Mol. Life Sci. 2019, 76, 99–128. [Google Scholar] [CrossRef]
- Gentile, C.L.; Pagliassotti, M.J. The role of fatty acids in the development and progression of nonalcoholic fatty liver disease. J. Nutr. Biochem. 2008, 19, 567–576. [Google Scholar] [CrossRef]
- Younossi, Z.M.; Stepanova, M.; Negro, F.; Hallaji, S.; Younossi, Y.; Lam, B.; Srishord, M. Nonalcoholic Fatty Liver Disease in Lean Individuals in the United States. Medicine 2012, 91, 319–327. [Google Scholar] [CrossRef]
- Tobari, M.; Hashimoto, E.; Taniai, M.; Ikarashi, Y.; Kodama, K.; Kogiso, T.; Tokushige, K.; Takayoshi, N.; Hashimoto, N. Characteristics of non-alcoholic steatohepatitis among lean patients in Japan: Not uncommon and not always benign. J. Gastroenterol. Hepatol. 2018, 34, 1404–1410. [Google Scholar] [CrossRef]
- Adamek, A.; Kasprzak, A. Insulin-Like Growth Factor (IGF) System in Liver Diseases. Int. J. Mol. Sci. 2018, 19, 1308. [Google Scholar] [CrossRef]
- Renner, S.; Dobenecker, B.; Blutke, A.; Zöls, S.; Wanke, R.; Ritzmann, M.; Wolf, E. Comparative aspects of rodent and nonrodent animal models for mechanistic and translational diabetes research. Theriogenology 2016, 86, 406–421. [Google Scholar] [CrossRef]
- Teppala, S.; Shankar, A. Association Between Serum IGF-1 and Diabetes Among U.S. Adults. Diabetes Care 2010, 33, 2257–2259. [Google Scholar] [CrossRef]
- He, J.; Kelley, D.E. Muscle glycogen content in type 2 diabetes mellitus. Am. J. Physiol. Metab. 2004, 287, E1002–E1007. [Google Scholar] [CrossRef] [PubMed]
- Moradi, S.; Ziaei, R.; Foshati, S.; Mohammadi, H.; Nachvak, S.M.; Rouhani, M.H. Effects of Spirulina supplementation on obesity: A systematic review and meta-analysis of randomized clinical trials. Complement. Ther. Med. 2019, 47, 102211. [Google Scholar] [CrossRef]
- Cattley, R.C.; Cullen, J.M. Liver and Gall bladder. In Haschek and Rousseaux’s Handbook of Toxicologic Pathology, 3rd ed.; Academic Press: London, UK, 2013; pp. 1509–1566. [Google Scholar]
- Yang, Y.; Du, L.; Hosokawa, M.; Miyashita, K. Effect of Spirulina lipids on high-fat and high-sucrose diet induced obesity and hepatic lipid accumulation in C57BL/6J mice. J. Funct. Foods 2020, 65, 103741. [Google Scholar] [CrossRef]
- Ide, T.; Iwase, H.; Amano, S.; Sunahara, S.; Tachihara, A.; Yagi, M.; Watanabe, T. Physiological effects of γ-linolenic acid and sesamin on hepatic fatty acid synthesis and oxidation. J. Nutr. Biochem. 2017, 41, 42–55. [Google Scholar] [CrossRef]
- Schinckel, A.; Ferrel, J.; Einstein, M.; Pearce, S.; Boyd, R. Analysis of Pig Growth from Birth to Sixty Days of Age1. Prof. Anim. Sci. 2004, 20, 79–86. [Google Scholar] [CrossRef]
- Fisher, K.D.; Scheffler, T.L.; Kasten, S.C.; Reinholt, B.M.; van Eyk, G.R.; Escobar, J.; Scheffler, J.M.; Gerrard, D.E. Energy Dense, Protein Restricted Diet Increases Adiposity and Perturbs Metabolism in Young, Genetically Lean Pigs. PLoS ONE 2013, 8, e72320. [Google Scholar] [CrossRef] [PubMed]
- Feng, R.; Du, S.; Chen, Y.; Zheng, S.; Zhang, W.; Na, G.; Li, Y.; Sun, C. High carbohydrate intake from starchy foods is positively associated with metabolic disorders: A Cohort Study from a Chinese population. Sci. Rep. 2015, 5, 16919. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Han, Y.S.; Kim, I.H. Growth performance, nutrient digestibility, antioxidant enzyme activity, and fecal microbiota flora in growing pigs fed diets containing Spirulina. Can. J. Anim. Sci. 2019, 99, 840–847. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gestation Diet | Lactation Diet | ||||
|---|---|---|---|---|---|
| CTRG | WESG | CTRL | WESL | Spirulina | |
| Ingredients (g/kg as fed) | |||||
| Hydrogenated palm oil | - | 150 | - | 150 | - |
| Saccharose | - | 200 | - | 200 | - |
| D-Fructose | - | 150 | - | 150 | - |
| Cholesterol | - | 2.0 | - | 2.0 | - |
| Corn germ | 590 | 298 | 488 | 200 | - |
| Wheat | 300 | - | 300 | - | - |
| Soybean meal | 45.0 | 130 | 145 | 217 | - |
| Lignocellulose † | 26.6 | 29.0 | 26.6 | 29.0 | - |
| Monocalcium phosphate | 13.0 | 20.0 | 11.2 | 20.0 | - |
| Calcium carbonate | 11.0 | 6.00 | 11.6 | 6.00 | - |
| NaCl | 6.2 | 6.0 | 6.2 | 6.0 | - |
| Vitamin and mineral premix ‡ | 5.0 | 5.0 | 5.0 | 5.0 | - |
| L-Lysine | 2.3 | 2.5 | 3.6 | 5.4 | - |
| DL-Methionine | - | 1.0 | 0.7 | 1.9 | - |
| DL-Tryptophan | 0.2 | 0.4 | 0.5 | 1.0 | - |
| L-Threonine | 0.7 | 1.3 | 1.8 | 3.4 | - |
| Valine | - | - | - | 2.2 | - |
| Chemical composition (g/kg dry matter (DM) if not stated otherwise) | |||||
| DM (g/kg fresh matter) | 893 | 936 | 898 | 934 | 935 |
| Total ash | 47.8 | 47.1 | 54.9 | 54.3 | 90.2 |
| Crude protein | 110 | 93 | 150 | 162 | 638 |
| Ether extract | 26 | 158 | 29 | 143 | 47 |
| Starch | 462 | 93 | 408 | 74 | 51 |
| Total sugars | 46 | 359 | 52 | 326 | <0.5 |
| Crude fiber | 6.7 | 8.7 | 6.2 | 8.1 | n.a. |
| Cholesterol | 0.03 | 0.48 | 0.02 | 0.45 | n.a. |
| Gross energy (MJ/kg DM) | 13.4 | 17.2 | 13.0 | 16.8 | n.a. |
| Fatty acids (FA) composition (g/100 g total FA) § | |||||
| C12:0 | 0.14 | 0.64 | 0.30 | 0.08 | 0.03 |
| C14:0 | 0.15 | 0.91 | 0.12 | 0.11 | 0.17 |
| C16:0 | 15.9 | 30.7 | 12.3 | 5.9 | 42.9 |
| iso C16:0 | 0.06 | 0.00 | 0.06 | 0.00 | 1.93 |
| C16:1 n–7 | 0.15 | 0.01 | 0.12 | 0.03 | 5.44 |
| C16:1x | 0.34 | 0.00 | 0.04 | 0.00 | 0.05 |
| C17:1 | 0.03 | 0.00 | 0.04 | 0.03 | 0.33 |
| C18:0 | 4.4 | 56.4 | 14.8 | 81.4 | 3.4 |
| C18:1 cis-9 | 26.0 | 3.3 | 21.9 | 2.8 | 1.6 |
| C18:1 cis-11 | 0.88 | 0.11 | 0.67 | 0.12 | 0.73 |
| C18:2 n–6 (LA) | 47.6 | 5.0 | 45.1 | 5.0 | 16.5 |
| C18:3 n–3 (ALA) | 1.93 | 0.34 | 2.28 | 0.68 | 0.43 |
| C18:3 n–6 (GLA) | 0.0 | 0.0 | 0.0 | 0.0 | 23.2 |
| C20:0 | 0.49 | 0.89 | 0.64 | 2.00 | 0.09 |
| C20:1 n–9 | 0.42 | 0.04 | 0.34 | 0.06 | 0.12 |
| C20:2 n–6 | 0.11 | 0.01 | 0.05 | 0.00 | 0.30 |
| C20:3 n–6 | 0.00 | 0.00 | 0.00 | 0.00 | 0.38 |
| C22:0 | 0.22 | 0.35 | 0.35 | 1.15 | 0.00 |
| ∑ Saturated FA | 22.0 | 90.3 | 29.1 | 91.1 | 49.2 |
| ∑ Monounsaturated FA | 28.0 | 4.3 | 23.3 | 3.1 | 8.9 |
| ∑ Polyunsaturated FA | 50.0 | 5.40 | 47.6 | 5.75 | 41.9 |
| ∑n–6 | 47.7 | 5.0 | 45.2 | 5.0 | 40.4 |
| ∑n–3 | 1.9 | 0.3 | 2.3 | 0.7 | 0.4 |
| n–6/n–3 | 25.1 | 14.7 | 19.6 | 7.4 | 101 |
| Gene | Forward Primer (5′→3′) | Reverse Primer (5′→3′) | Amplicon Length | Accession Number |
|---|---|---|---|---|
| ACTB | GATCTGGCACCACACCTTCT | AGAGACAGCACAGCCTGGAT | 174 | NM_173979.3 |
| CYP1A2 | GCCCAGCCCTACTCTGCAA | CCAGGAGATGGCTGTGGTAA | 250 | XM_005666124.3 |
| CRP | TGAACACAGGCTCTCACATCC | CAAGCCAGACACTTGAATGCC | 70 | XM_003355107.4 |
| GSTA1 | CAGGACACCCAGGACCAATC | GTCTCAGGTACATTCCGGGAG | 202 | NM_214389.2 |
| GSTA4 | GCTCGGAGTGGACCCAGAAAA | TTCGGGTCTGCACCAACTTC | 243 | NM_001243379.1 |
| H3F3A | AGGAGGTCTCTATACCATGGCTC | GAGCAATTTCCCGCACCAGA | 245 | NM_213930.1 |
| HPRT1 | TGCTGAGGATTTGGAGAAGG | CAACAGGTCGGCAAAGAACT | 154 | NM_001034035.2 |
| IGF1 | TGGTGGACGCTCTTCAGTTCG | ACAGTACATCTCCAGCCTCCTC | 155 | NM_214256.1 |
| LIPG | CGAAACTCAGTTCCTCTGCTCT | TGGCTGTTGCATTGAAGCCA | 247 | XM_013992843.2 |
| PCK1 | GGGCATCATCTTCGGAGGG | AGTTGTAGCCGAAGAACGGC | 182 | XM_005673043.3 |
| PTDGS | AAGAACTACGCCCTGCTCCA | ATGGCCAGGTCCTGAGAGT | 231 | NM_214228.1 |
| SDHA | GCAGAACCTGATGCTTTGTG | CGTAGGAGAGCGTGTGCTT | 185 | NM_174178.2 |
| UBB | CATTGTTGGCGGTTTCGCT | TTGACCTGTGAGTGAAGGCA | 85 | NM_001105309.1 |
| YWHAZ | ATTGGGTCTGGCCCTTAACT | GCGTGCTGTCTTTGTATGACTC | 146 | XM_021088756.1 |
| Diet (D) | Control Diet | Western Diet | SEM | Significance | ||||
|---|---|---|---|---|---|---|---|---|
| Spirulina (Sp) | − | + | − | + | D | Sp | D × Sp | |
| Liver | ||||||||
| Proportion (g/kg BW) | 11.1 | 10.1 | 11.8 | 10.8 | 1.10 | # | * | n.s. |
| Glycogen (µM/g) | 2.17 | 1.14 | 2.10 | 1.41 | 0.826 | n.s. | n.s. | n.s. |
| Triglycerides (mg/g) | 4.20 | 4.17 | 4.13 | 4.23 | 1.057 | n.s. | n.s. | n.s. |
| Cholesterol (mg/g) | 1.33 | 1.47 | 1.85 | 1.95 | 0.321 | # | n.s. | n.s. |
| Skeletal muscle | ||||||||
| Intramuscular fat (g/kg DM) | 55.7 | 82.7 | 56.0 | 52.3 | 17.30 | n.s. | n.s. | n.s. |
| Protein (g/kg DM) | 18.5 | 19.0 | 19.3 | 17.9 | 1.19 | n.s. | n.s. | n.s. |
| Cholesterol (mg/100g) | 51.1 | 54.9 | 55.1 | 56.7 | 2.53 | * | * | n.s. |
| Glycogen (µM/g) | 13.7 | 12.8 | 9.81 | 4.17 | 2.650 | ** | n.s. | n.s. |
| Visceral adipose tissue | ||||||||
| Proportion (g/kg BW) | 12.6 | 11.6 | 10.1 | 12.2 | 1.86 | n.s. | n.s. | n.s. |
| Adipocyte area (µm2 × 103) | 16.3 | 12.2 | 14.2 | 16.5 | 3.23 | n.s | n.s. | # |
| Adipocyte proportion (%) | ||||||||
| <5 (µm2 × 103) | 5.10 | 5.86 | 4.05 | 6.00 | 6.300 | n.s. | n.s. | n.s. |
| 5–15 | 41.0 | 66.8 | 55.5 | 39.6 | 24.68 | n.s. | n.s. | n.s. |
| 15–25 | 52.0 | 23.4 | 36.1 | 49.0 | 15.76 | n.s. | n.s. | * |
| >25 | 5.68 | 2.37 | 4.37 | 8.85 | 8.58 | n.s. | n.s. | n.s. |
| Diet (D) | Control Diet | Western Diet | Significance | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Spirulina (Sp) | − | + | − | + | SEM | D | Sp | D × Sp | |
| Tissue | |||||||||
| ∑SFA | Liver | 41.6 | 41.6 | 43.3 | 42.6 | 0.78 | * | n.s. | n.s. |
| Muscle | 41.0 | 39.9 | 42.6 | 40.9 | 1.10 | n.s. | # | n.s. | |
| ∑MUFA | Liver | 18.8 | 18.4 | 18.3 | 21.8 | 3.47 | n.s. | n.s. | n.s. |
| Muscle | 46.8 | 47.3 | 44.9 | 47.5 | 2.14 | n.s. | n.s. | n.s. | |
| ∑PUFA | Liver | 39.7 | 40.0 | 38.3 | 35.6 | 2.87 | # | n.s. | n.s. |
| Muscle | 12.5 | 12.7 | 13.1 | 10.6 | 2.42 | n.s. | n.s. | n.s. | |
| ∑n–3 | Liver | 3.60 | 3.72 | 3.90 | 3.58 | 4.500 | n.s. | n.s. | n.s. |
| Muscle | 0.58 | 0.62 | 0.78 | 0.63 | 0.119 | n.s. | n.s. | n.s. | |
| ∑n–6 | Liver | 36.0 | 36.1 | 34.3 | 31.9 | 2.49 | * | n.s. | n.s. |
| Muscle | 11.7 | 12.0 | 12.3 | 9.8 | 2.31 | n.s. | n.s. | n.s. | |
| n–6/n–3 | Liver | 10.0 | 9.7 | 8.8 | 8.9 | 0.01 | n.s. | n.s. | n.s. |
| Muscle | 20.2 | 19.4 | 15.8 | 15.6 | 0.003 | *** | n.s. | n.s. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lugarà, R.; Renner, S.; Wolf, E.; Liesegang, A.; Bruckmaier, R.; Giller, K. Crossbred Sows Fed a Western Diet during Pre-Gestation, Gestation, Lactation, and Post-Lactation Periods Develop Signs of Lean Metabolic Syndrome That Are Partially Attenuated by Spirulina Supplementation. Nutrients 2022, 14, 3574. https://doi.org/10.3390/nu14173574
Lugarà R, Renner S, Wolf E, Liesegang A, Bruckmaier R, Giller K. Crossbred Sows Fed a Western Diet during Pre-Gestation, Gestation, Lactation, and Post-Lactation Periods Develop Signs of Lean Metabolic Syndrome That Are Partially Attenuated by Spirulina Supplementation. Nutrients. 2022; 14(17):3574. https://doi.org/10.3390/nu14173574
Chicago/Turabian StyleLugarà, Rosamaria, Simone Renner, Eckhard Wolf, Annette Liesegang, Rupert Bruckmaier, and Katrin Giller. 2022. "Crossbred Sows Fed a Western Diet during Pre-Gestation, Gestation, Lactation, and Post-Lactation Periods Develop Signs of Lean Metabolic Syndrome That Are Partially Attenuated by Spirulina Supplementation" Nutrients 14, no. 17: 3574. https://doi.org/10.3390/nu14173574
APA StyleLugarà, R., Renner, S., Wolf, E., Liesegang, A., Bruckmaier, R., & Giller, K. (2022). Crossbred Sows Fed a Western Diet during Pre-Gestation, Gestation, Lactation, and Post-Lactation Periods Develop Signs of Lean Metabolic Syndrome That Are Partially Attenuated by Spirulina Supplementation. Nutrients, 14(17), 3574. https://doi.org/10.3390/nu14173574