
Inonotus hispidus Protects against Hyperlipidemia by Inhibiting Oxidative Stress and Inflammation through Nrf2/NF-κB Signaling in High Fat Diet Fed Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Detection of IH Components
2.2. Animal Experiments and Agent Administration Protocol
2.3. Cytokine Detection
2.4. Histopathological Analysis
2.5. Intestinal Microflora Analysis
2.6. Plasma Lipidome Analysis
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
3.1. Main Composition of IH
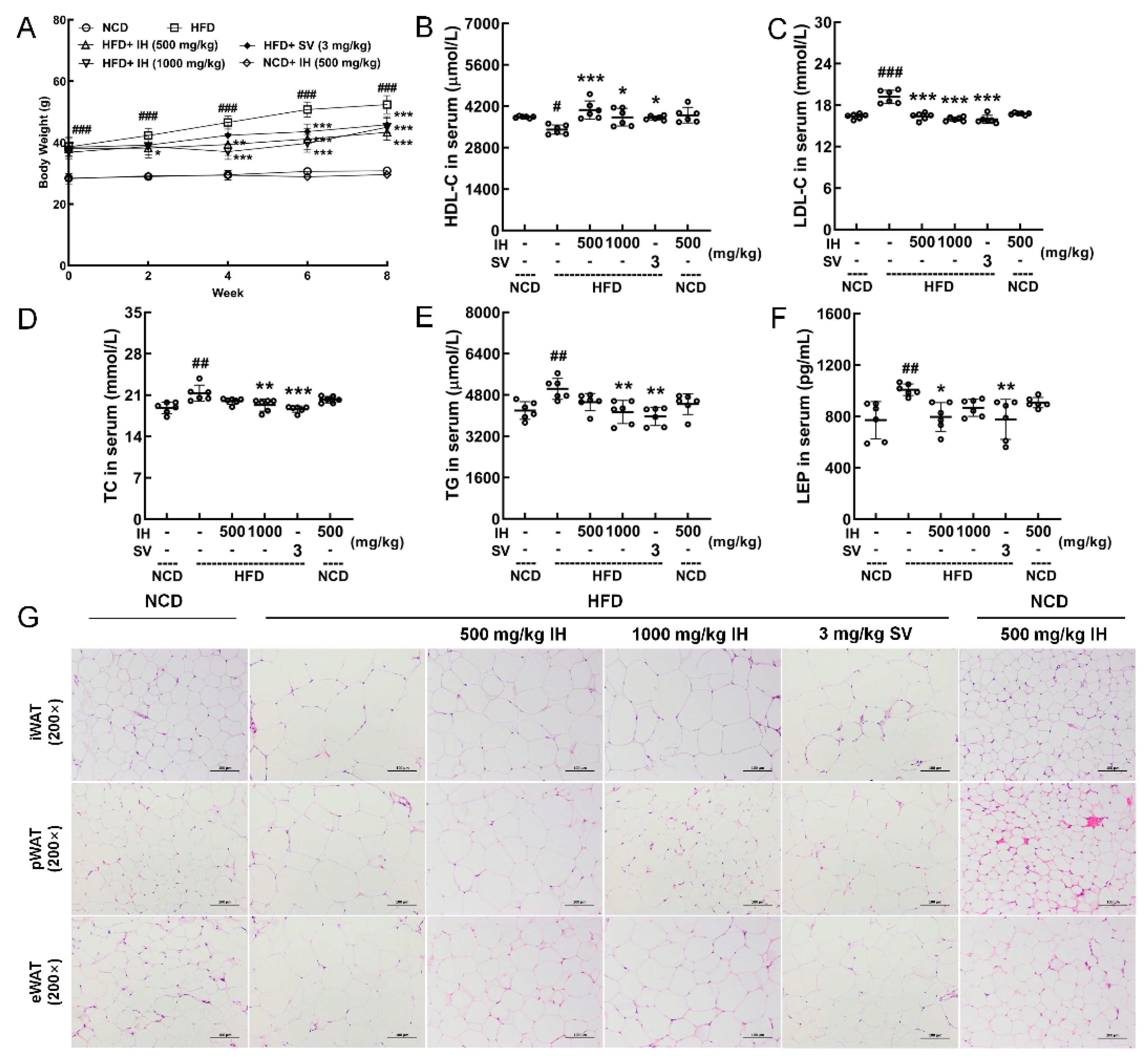
3.2. Hypolipidemic Effects of IH in HFD-Fed Mice
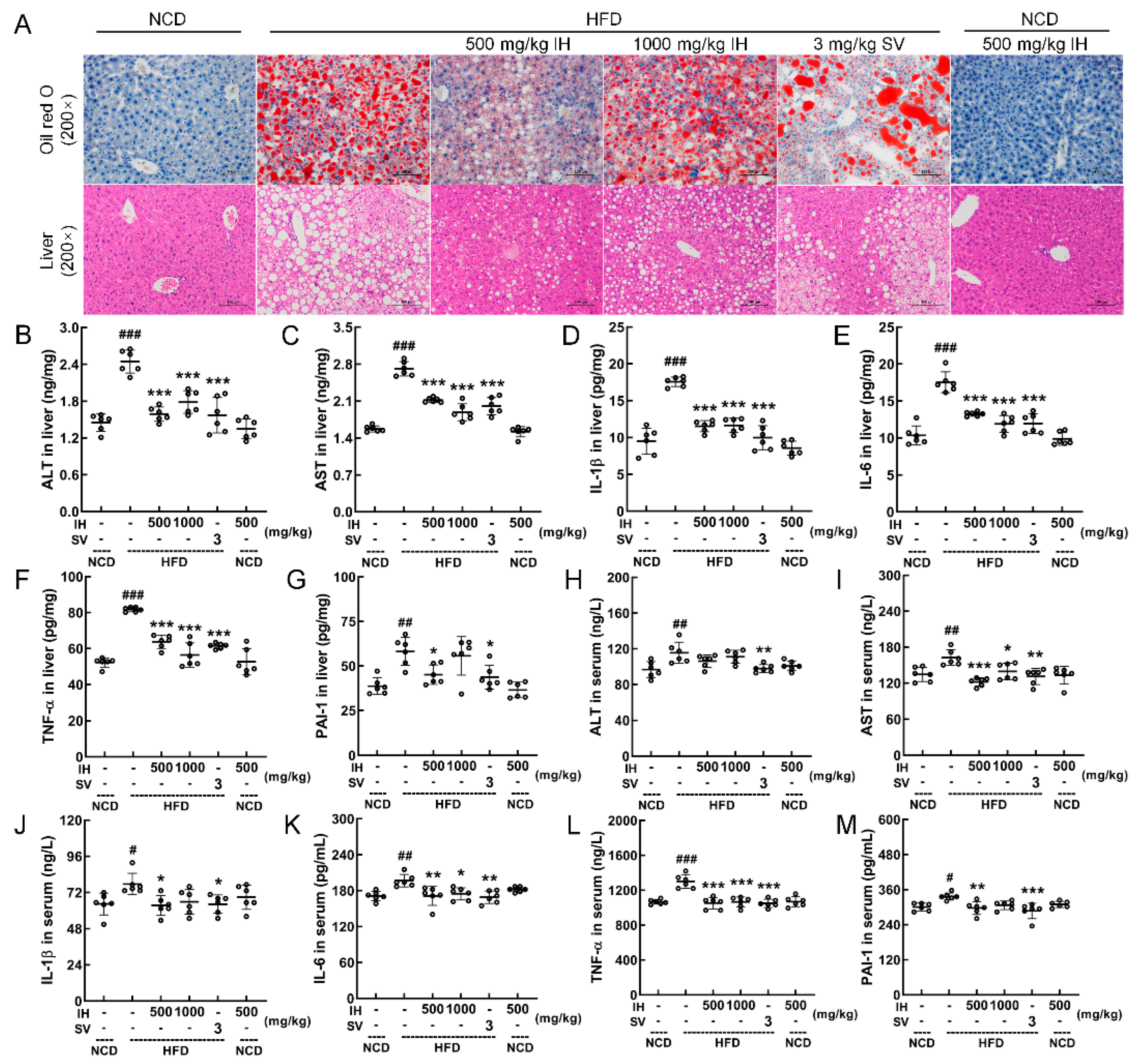
3.3. IH Ameliorated Hepatic Steatosis in HFD-Fed Mice
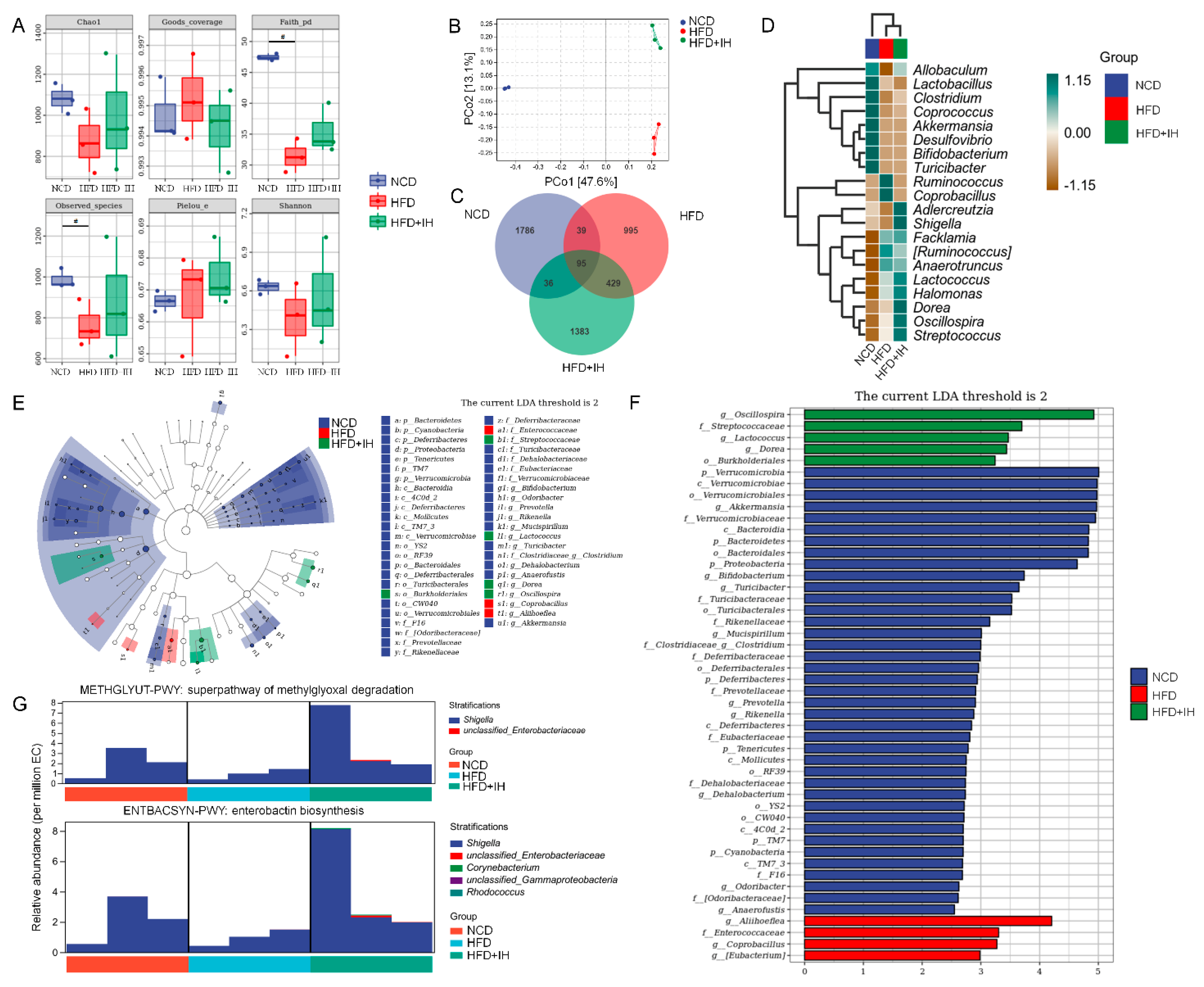
3.4. IH Regulated Intestinal Microflora in HFD-Fed Mice
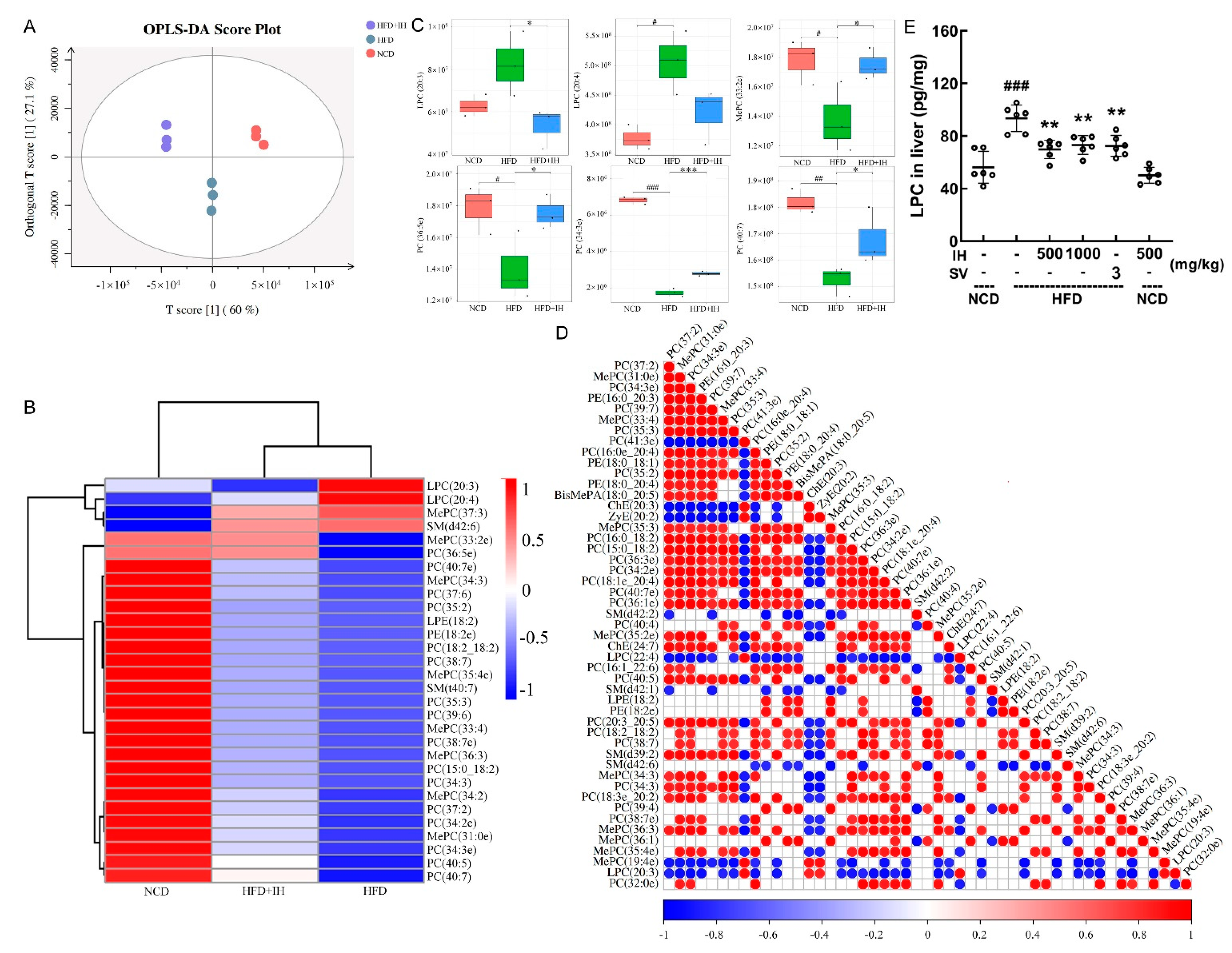
3.5. IH Regulated Lipid Metabolism
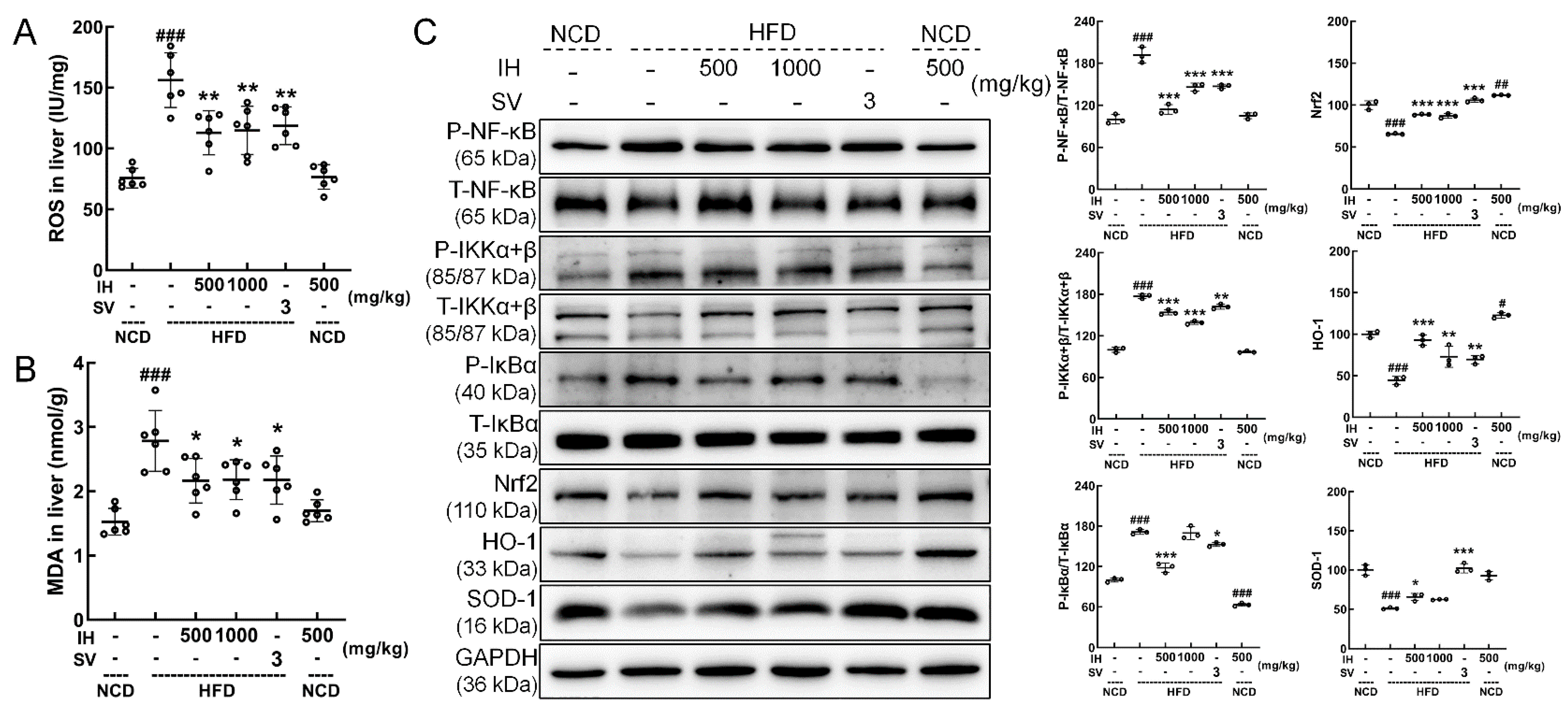
3.6. IH Regulated Nrf2/NF-κB Pathway and Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bluher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Yoo, A.; Kim, M.J.; Ahn, J.; Jung, C.H.; Seo, H.D.; Ly, S.Y.; Ha, T.Y. Fuzhuan brick tea extract prevents diet-induced obesity via stimulation of fat browning in mice. Food Chem. 2022, 377, 132006. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.B.; Oh, M.J.; Do, M.H.; Kim, Y.; Choi, I.; Kim, Y.S.; Park, H.Y. Dietary rhamnogalacturonan-I rich extracts of molokhia ameliorate high fat diet-induced obesity and gut dysbiosis. J. Nutr. Biochem. 2022, 103, 108954. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.F.; Wang, L.M.; Pan, A. Epidemiology and determinants of obesity in China. Lancet Diabetes Endocrinol. 2021, 9, 373–392. [Google Scholar] [CrossRef]
- Chakrabarty, S.; Bui, Q.Y.; Badeanlou, L.; Hester, K.; Chun, J.; Ruf, W.; Ciaraldi, T.P.; Samad, F. S1P/S1PR3 signalling axis protects against obesity-induced metabolic dysfunction. Adipocyte 2022, 11, 69–83. [Google Scholar] [CrossRef]
- Xue, C.Y.; Li, Y.; Lv, H.; Zhang, L.; Bi, C.P.; Dong, N.; Shan, A.S.; Wang, J.L. Oleanolic Acid Targets the Gut-Liver Axis to Alleviate Metabolic Disorders and Hepatic Steatosis. J. Agric. Food Chem. 2021, 69, 7884–7897. [Google Scholar] [CrossRef]
- Zhou, F.; Li, Y.L.; Zhang, X.; Wang, K.B.; Huang, J.A.; Liu, Z.H.; Zhu, M.Z. Polyphenols from Fu Brick Tea Reduce Obesity via Modulation of Gut Microbiota and Gut Microbiota-Related Intestinal Oxidative Stress and Barrier Function. J. Agric. Food Chem. 2021, 69, 14530–14543. [Google Scholar] [CrossRef]
- Cakir, I.; Lining Pan, P.; Hadley, C.K.; El-Gamal, A.; Fadel, A.; Elsayegh, D.; Mohamed, O.; Rizk, N.M.; Ghamari-Langroudi, M. Sulforaphane reduces obesity by reversing leptin resistance. eLife 2022, 11, e67368. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Wang, D.; Gu, Y.; Jiang, Z.X.; Zhou, Z.Q. Tangeretin prevents obesity by modulating systemic inflammation, fat browning, and gut microbiota in high-fat diet-induced obese C57BL/6 mice. J. Nutr. Biochem. 2022, 101, 108943. [Google Scholar] [CrossRef]
- Gutierrez-Cuevas, J.; Galicia-Moreno, M.; Monroy-Ramirez, H.C.; Sandoval-Rodriguez, A.; Garcia-Banuelos, J.; Santos, A.; Armendariz-Borunda, J. The Role of NRF2 in Obesity-Associated Cardiovascular Risk Factors. Antioxidants 2022, 11, 235. [Google Scholar] [CrossRef]
- Scheithauer, T.P.M.; Davids, M.; Winkelmeijer, M.; Verdoes, X.; Aydin, O.; de Brauw, M.; van de Laar, A.; Meijnikman, A.S.; Gerdes, V.E.A.; van Raalte, D.; et al. Compensatory intestinal antibody response against pro-inflammatory microbiota after bariatric surgery. Gut Microbes 2022, 14, 2034696. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, F.; Chopra, H.; Baig, A.A.; Avula, S.K.; Kumari, S.; Mohanta, T.K.; Saravanan, M.; Mishra, A.K.; Sharma, N.; Mohanta, Y.K. Edible Mushrooms as Novel Myco-Therapeutics: Effects on Lipid Level, Obesity, and BMI. J. Fungi 2022, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Xu, B.J. Anti-Obesity Effects of Medicinal and Edible Mushrooms. Molecules 2018, 23, 2880. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Hao, J.; Liu, Z.J.; Ma, X.T.; Feng, Y.X.; Teng, L.R.; Li, Y.; Wang, D. Anti-obesity effects of Grifola frondosa through the modulation of lipid metabolism via ceramide in mice fed a high-fat diet. Food Funct. 2021, 12, 6725–6739. [Google Scholar] [CrossRef]
- Yang, S.D.; Bao, H.Y.; Wang, H.; Li, Q.J. Anti-tumour Effect and Pharmacokinetics of an Active Ingredient Isolated from Inonotus hispidus. Biol. Pharm. Bull. 2019, 42, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Ali, N.A.A.; Jansen, R.; Pilgrim, H.; Liberra, K.; Lindequist, U. Hispolon, a yellow pigment from Inonotus hispidus. Phytochemistry 1996, 41, 927–929. [Google Scholar] [CrossRef]
- Liu, X.; Hou, R.L.; Xu, K.Q.; Chen, L.; Wu, X.P.; Lin, W.X.; Zheng, M.F.; Fu, J.S. Extraction, characterization and antioxidant activity analysis of the polysaccharide from the solid-state fermentation substrate of Inonotus hispidus. Int. J. Biol. Macromol. 2019, 123, 468–476. [Google Scholar] [CrossRef]
- Ren, Q.; Lu, X.Y.; Han, J.X.; Aisa, H.A.; Yuan, T. Triterpenoids and phenolics from the fruiting bodies of Inonotus hispidus and their activations of melanogenesis and tyrosinase. Chin. Chem. Lett. 2017, 28, 1052–1056. [Google Scholar] [CrossRef]
- Talhinhas, P.; Tavares, D.; Ramos, A.P.; Goncalves, S.; Loureiro, J. Validation of standards suitable for genome size estimation of fungi. J. Microbiol. Methods 2017, 142, 76–78. [Google Scholar] [CrossRef]
- Grundemann, C.; Arnhold, M.; Meier, S.; Backer, C.; Garcia-Kaufer, M.; Grunewald, F.; Steinborn, C.; Klemd, A.M.; Wille, R.; Huber, R.; et al. Effects of Inonotus hispidus Extracts and Compounds on Human Immunocompetent Cells. Planta Med. 2016, 82, 1359–1367. [Google Scholar] [CrossRef]
- Li, Z.; Bao, H. Anti-tumor effect of Inonotus hispidus petroleum ether extract in H22 tumor-bearing mice and analysis its mechanism by untargeted metabonomic. J. Ethnopharmacol. 2022, 285, 114898. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hou, R.L.; Yan, J.J.; Xu, K.Q.; Wu, X.P.; Lin, W.X.; Zheng, M.F.; Fu, J.S. Purification and characterization of Inonotus hispidus exopolysaccharide and its protective effect on acute alcoholic liver injury in mice. Int. J. Biol. Macromol. 2019, 129, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, X.; Zhang, Y.F.; Zhang, Y.Q.; Liu, S.Y.; Zhang, N.; Li, Y.; Wang, D. Protective effect of Gloeostereum incarnatum on ulcerative colitis via modulation of Nrf2/NF-kappa B signaling in C57BL/6 mice. Mol. Med. Rep. 2020, 22, 3418–3428. [Google Scholar] [CrossRef]
- Zhang, Z.; Lin, T.; Meng, Y.; Hu, M.; Shu, L.; Jiang, H.; Gao, R.; Ma, J.; Wang, C.; Zhou, X. FOS/GOS attenuates high-fat diet induced bone loss via reversing microbiota dysbiosis, high intestinal permeability and systemic inflammation in mice. Metabolism 2021, 119, 154767. [Google Scholar] [CrossRef]
- Wu, H.; Rao, Q.; Ma, G.-C.; Yu, X.-H.; Zhang, C.-E.; Ma, Z.-J. Effect of Triptolide on Dextran Sodium Sulfate-Induced Ulcerative Colitis and Gut Microbiota in Mice. Front. Pharm. 2020, 10, 1652. [Google Scholar] [CrossRef]
- Fu, X.; Guo, J.; Finkelbergs, D.; He, J.; Zha, L.; Guo, Y.; Cai, J. Fungal succession during mammalian cadaver decomposition and potential forensic implications. Sci. Rep. 2019, 9, 12907. [Google Scholar] [CrossRef]
- Baye, E.; Ukropec, J.; de Courten, M.P.; Vallova, S.; Krumpolec, P.; Kurdiova, T.; Aldini, G.; Ukropcova, B.; de Courten, B. Effect of carnosine supplementation on the plasma lipidome in overweight and obese adults: A pilot randomised controlled trial. Sci. Rep. 2017, 7, 17458. [Google Scholar] [CrossRef]
- Nie, Y.; Luo, F. Dietary Fiber: An Opportunity for a Global Control of Hyperlipidemia. Oxidative Med. Cell. Longev. 2021, 2021, 5542342. [Google Scholar] [CrossRef]
- Mayer, C.; Côme, M.; Ulmann, L.; Chini Zittelli, G.; Faraloni, C.; Nazih, H.; Ouguerram, K.; Chénais, B.; Mimouni, V. Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients 2019, 11, 1069. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Hållenius, F.F.; Lin, X.; Nyman, M.; Prykhodko, O. Monobutyrin and Monovalerin Affect Brain Short-Chain Fatty Acid Profiles and Tight-Junction Protein Expression in ApoE-Knockout Rats Fed High-Fat Diets. Nutrients 2020, 12, 1202. [Google Scholar] [CrossRef]
- Khwanchuea, R.; Punsawad, C. Associations Between Body Composition, Leptin, and Vitamin D Varied by the Body Fat Percentage in Adolescents. Front. Endocrinol 2022, 13, 876231. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Guo, Y.; Sun, L.; Lai, X.; Li, Q.; Zhang, W.; Xiang, L.; Sun, S.; Cao, F. Six types of tea reduce high-fat-diet-induced fat accumulation in mice by increasing lipid metabolism and suppressing inflammation. Food Funct. 2019, 10, 2061–2074. [Google Scholar] [CrossRef] [PubMed]
- Jiao, N.; Baker, S.S.; Nugent, C.A.; Tsompana, M.; Cai, L.T.; Wang, Y.; Buck, M.J.; Genco, R.J.; Baker, R.D.; Zhu, R.X.; et al. Gut microbiome may contribute to insulin resistance and systemic inflammation in obese rodents: A meta-analysis. Physiol. Genom. 2018, 50, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Wen, Z.; Liu, W.; Meng, L.; Huang, H. Oscillospira—A candidate for the next-generation probiotics. Gut Microbes 2021, 13, 1987783. [Google Scholar] [CrossRef] [PubMed]
- He, W.S.; Li, L.; Rui, J.; Li, J.; Sun, Y.; Cui, D.; Xu, B. Tomato seed oil attenuates hyperlipidemia and modulates gut microbiota in C57BL/6J mice. Food Funct. 2020, 11, 4275–4290. [Google Scholar] [CrossRef]
- Wang, X.; Shi, L.; Wang, X.; Feng, Y.; Wang, Y. MDG-1, an Ophiopogon polysaccharide, restrains process of non-alcoholic fatty liver disease via modulating the gut-liver axis. Int. J. Biol. Macromol. 2019, 141, 1013–1021. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis gut microbiota associated with inflammation and impaired mucosal immune function in intestine of humans with non-alcoholic fatty liver disease. Sci Rep. 2015, 5, 8096. [Google Scholar] [CrossRef]
- Stinson, L.F.; Gay, M.C.L.; Koleva, P.T.; Eggesbø, M.; Johnson, C.C.; Wegienka, G.; du Toit, E.; Shimojo, N.; Munblit, D.; Campbell, D.E.; et al. Human Milk From Atopic Mothers Has Lower Levels of Short Chain Fatty Acids. Front. Immunol. 2020, 11, 1427. [Google Scholar] [CrossRef]
- Fei, Y.; Wang, Y.; Pang, Y.; Wang, W.; Zhu, D.; Xie, M.; Lan, S.; Wang, Z. Xylooligosaccharide Modulates Gut Microbiota and Alleviates Colonic Inflammation Caused by High Fat Diet Induced Obesity. Front. Physiol. 2019, 10, 1601. [Google Scholar] [CrossRef]
- Raza, G.S.; Putaala, H.; Hibberd, A.A.; Alhoniemi, E.; Tiihonen, K.; Mäkelä, K.A.; Herzig, K.H. Polydextrose changes the gut microbiome and attenuates fasting triglyceride and cholesterol levels in Western diet fed mice. Sci. Rep. 2017, 7, 5294. [Google Scholar] [CrossRef]
- Terzo, S.; Mulè, F.; Caldara, G.F.; Baldassano, S.; Puleio, R.; Vitale, M.; Cassata, G.; Ferrantelli, V.; Amato, A. Pistachio Consumption Alleviates Inflammation and Improves Gut Microbiota Composition in Mice Fed a High-Fat Diet. Int. J. Mol. Sci 2020, 21, 365. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Wu, S.; Xia, Y.; Huang, J.; Ye, J.; Xuan, Z.; Li, P.; Du, B. Probiotic-fermented black tartary buckwheat alleviates hyperlipidemia and gut microbiota dysbiosis in rats fed with a high-fat diet. Food Funct. 2021, 12, 6045–6057. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Liu, J.; Gao, J.; Wu, X.; Cui, C.; Wei, H.; Zheng, R.; Peng, J. Combined Soluble Fiber-Mediated Intestinal Microbiota Improve Insulin Sensitivity of Obese Mice. Nutrients 2020, 12, 351. [Google Scholar] [CrossRef]
- Jin, H.; Leng, Q.; Zhang, C.; Zhu, Y.; Wang, J. P-cymene prevent high-fat diet-associated colorectal cancer by improving the structure of intestinal flora. J. Cancer 2021, 12, 4355–4361. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Zhang, F.; Zhang, J.; Zhang, X.; Guo, Y.; Yao, Q. A Holistic View of Berberine Inhibiting Intestinal Carcinogenesis in Conventional Mice Based on Microbiome-Metabolomics Analysis. Front. Immunol. 2020, 11, 588079. [Google Scholar] [CrossRef]
- Chen, J.; Chia, N.; Kalari, K.R.; Yao, J.Z.; Novotna, M.; Paz Soldan, M.M.; Luckey, D.H.; Marietta, E.V.; Jeraldo, P.R.; Chen, X.; et al. Multiple sclerosis patients have a distinct gut microbiota compared to healthy controls. Sci. Rep. 2016, 6, 28484. [Google Scholar] [CrossRef]
- Konikoff, T.; Gophna, U. Oscillospira: A Central, Enigmatic Component of the Human Gut Microbiota. Trends Microbiol. 2016, 24, 523–524. [Google Scholar] [CrossRef]
- Yu, M.; Yue, J.; Hui, N.; Zhi, Y.; Hayat, K.; Yang, X.; Zhang, D.; Chu, S.; Zhou, P. Anti-Hyperlipidemia and Gut Microbiota Community Regulation Effects of Selenium-Rich Cordyceps militaris Polysaccharides on the High-Fat Diet-Fed Mice Model. Foods 2021, 10, 2252. [Google Scholar] [CrossRef]
- Atarashi, M.; Izawa, T.; Miyagi, R.; Ohji, S.; Hashimoto, A.; Kuwamura, M.; Yamate, J. Dietary Iron Supplementation Alters Hepatic Inflammation in a Rat Model of Nonalcoholic Steatohepatitis. Nutrients 2018, 10, 175. [Google Scholar] [CrossRef]
- Ji, J.; Zhang, S.; Tang, L.; Zhang, M.; Yuan, M.; Wang, P.; Gao, X. Integrative analysis of fecal metabolome and gut microbiota in high-fat diet-induced hyperlipidemic rats treated with Rosa Roxburghii Tratt juice. J. Funct. Foods 2022, 90, 104978. [Google Scholar] [CrossRef]
- Xing, J.; Liu, G.; Zhang, X.; Bai, D.; Yu, J.; Li, L.; Wang, X.; Su, S.; Zhao, Y.; Bou, G.; et al. The Composition and Predictive Function of the Fecal Microbiota Differ Between Young and Adult Donkeys. Front. Microbiol. 2020, 11, 596394. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.; Corbalán, N.S.; Seyedsayamdost, M.R.; Pomares, M.F.; de Cristóbal, R.E.; Clardy, J.; Kolter, R.; Vincent, P.A. Catecholate siderophores protect bacteria from pyochelin toxicity. PLoS ONE 2012, 7, e46754. [Google Scholar] [CrossRef] [PubMed]
- Markova, I.; Huttl, M.; Oliyarnyk, O.; Kacerova, T.; Haluzik, M.; Kacer, P.; Seda, O.; Malinska, H. The effect of dicarbonyl stress on the development of kidney dysfunction in metabolic syndrome—A transcriptomic and proteomic approach. Nutr. Metab. 2019, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-H.; Lee, B.-H.; Li, C.-H.; Hsu, Y.-W.; Pan, T.-M. Monascin and AITC Attenuate Methylglyoxal-Induced PPARγ Phosphorylation and Degradation through Inhibition of the Oxidative Stress/PKC Pathway Depending on Nrf2 Activation. J. Agric. Food Chem. 2013, 61, 5996–6006. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhu, Y.; Wang, P.; Sang, S. Dietary Genistein Reduces Methylglyoxal and Advanced Glycation End Product Accumulation in Obese Mice Treated with High-Fat Diet. J. Agric. Food Chem. 2020, 68, 7416–7424. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Li, R.; Zhu, L.; Tribble, G.D.; Zheng, W.J.; Ferguson, B.; Maddipati, K.R.; Angelov, N.; Van Dyke, T.E. Subgingival Microbiome and Specialized Pro-Resolving Lipid Mediator Pathway Profiles Are Correlated in Periodontal Inflammation. Front. Immunol. 2021, 12, 691216. [Google Scholar] [CrossRef]
- Li, X.; Wang, L.; Fang, P.; Sun, Y.; Jiang, X.; Wang, H.; Yang, X.-F. Lysophospholipids induce innate immune transdifferentiation of endothelial cells, resulting in prolonged endothelial activation. J. Biol. Chem. 2018, 293, 11033–11045. [Google Scholar] [CrossRef]
- Wu, R.; Huang, Y.H.; Elinder, L.S.; Frostegård, J. Lysophosphatidylcholine is involved in the antigenicity of oxidized LDL. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 626–630. [Google Scholar] [CrossRef][Green Version]
- Kim, S.H.; Kim, B.K.; Park, S.; Park, S.K. Phosphatidylcholine Extends Lifespan via DAF-16 and Reduces Amyloid-Beta-Induced Toxicity in Caenorhabditis elegans. Oxid Med. Cell Longev. 2019, 2019, 2860642. [Google Scholar] [CrossRef]
- Lee, H.S.; Kim, B.K.; Nam, Y.; Sohn, U.D.; Park, E.S.; Hong, S.A.; Lee, J.H.; Chung, Y.H.; Jeong, J.H. Protective role of phosphatidylcholine against cisplatin-induced renal toxicity and oxidative stress in rats. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2013, 58, 388–393. [Google Scholar] [CrossRef]
- Zhang, W.; Randell, E.W.; Sun, G.; Likhodii, S.; Liu, M.; Furey, A.; Zhai, G. Hyperglycemia-related advanced glycation end-products is associated with the altered phosphatidylcholine metabolism in osteoarthritis patients with diabetes. PLoS ONE 2017, 12, e0184105. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Lv, W.; Liu, X.; Fan, Y.; Wang, K.; Feng, Z.; Liu, J.; Zang, W.; Xing, L.; Liu, J. Herba houttuyniae Extract Benefits Hyperlipidemic Mice via Activation of the AMPK/PGC-1α/Nrf2 Cascade. Nutrients 2020, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- Sarega, N.; Imam, M.U.; Ooi, D.J.; Chan, K.W.; Md Esa, N.; Zawawi, N.; Ismail, M. Phenolic Rich Extract from Clinacanthus nutans Attenuates Hyperlipidemia-Associated Oxidative Stress in Rats. Oxid Med. Cell Longev 2016, 2016, 4137908. [Google Scholar] [CrossRef]
- Slocum, S.L.; Skoko, J.J.; Wakabayashi, N.; Aja, S.; Yamamoto, M.; Kensler, T.W.; Chartoumpekis, D.V. Keap1/Nrf2 pathway activation leads to a repressed hepatic gluconeogenic and lipogenic program in mice on a high-fat diet. Arch. Biochem. Biophys. 2016, 591, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Ha Kim, K.; Sadikot, R.T.; Yeon Lee, J.; Jeong, H.S.; Oh, Y.K.; Blackwell, T.S.; Joo, M. Suppressed ubiquitination of Nrf2 by p47(phox) contributes to Nrf2 activation. Free Radic. Biol. Med. 2017, 113, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos Maia, M.; Rodrigues, G.C.S.; de Sousa, N.F.; Scotti, M.T.; Scotti, L.; Mendonça-Junior, F.J.B. Identification of New Targets and the Virtual Screening of Lignans against Alzheimer’s Disease. Oxidative Med. Cell. Longev. 2020, 2020, 3098673. [Google Scholar] [CrossRef]
- Laddha, N.C.; Dwivedi, M.; Gani, A.R.; Shajil, E.M.; Begum, R. Involvement of superoxide dismutase isoenzymes and their genetic variants in progression of and higher susceptibility to vitiligo. Free Radic. Biol. Med. 2013, 65, 1110–1125. [Google Scholar] [CrossRef]
- Liu, H.; Zhu, H.; Xia, H.; Yang, X.; Yang, L.; Wang, S.; Wen, J.; Sun, G. Different effects of high-fat diets rich in different oils on lipids metabolism, oxidative stress and gut microbiota. Food Res. Int. 2021, 141, 110078. [Google Scholar] [CrossRef]
- Qiu, M.; Xiao, F.; Wang, T.; Piao, S.; Zhao, W.; Shao, S.; Yan, M.; Zhao, D. Protective effect of Hedansanqi Tiaozhi Tang against non-alcoholic fatty liver disease in vitro and in vivo through activating Nrf2/HO-1 antioxidant signaling pathway. Phytomedicine 2020, 67, 153140. [Google Scholar] [CrossRef]
- Tan, S.M.; Sharma, A.; Stefanovic, N.; Yuen, D.Y.C.; Karagiannis, T.C.; Meyer, C.; Ward, K.W.; Cooper, M.E.; de Haan, J.B. Derivative of Bardoxolone Methyl, dh404, in an Inverse Dose-Dependent Manner Lessens Diabetes-Associated Atherosclerosis and Improves Diabetic Kidney Disease. Diabetes 2014, 63, 3091–3103. [Google Scholar] [CrossRef]
- Mabuchi, S.; Ohmichi, M.; Nishio, Y.; Hayasaka, T.; Kimura, A.; Ohta, T.; Kawagoe, J.; Takahashi, K.; Yada-Hashimoto, N.; Seino-Noda, H.; et al. Inhibition of inhibitor of nuclear factor-kappaB phosphorylation increases the efficacy of paclitaxel in in vitro and in vivo ovarian cancer models. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 7645–7654. [Google Scholar] [CrossRef] [PubMed]
- Kizilay Mancini, O.; Huynh, D.N.; Menard, L.; Shum-Tim, D.; Ong, H.; Marleau, S.; Colmegna, I.; Servant, M.J. Ex vivo Ikkβ ablation rescues the immunopotency of mesenchymal stromal cells from diabetics with advanced atherosclerosis. Cardiovasc Res. 2021, 117, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Celec, P. Nuclear factor kappa B—Molecular biomedicine: The next generation. Biomed. Pharmacother. 2004, 58, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Q.; Wu, Y.; Yang, M.; Wu, G.; Wang, Y.; Wang, H.; Feng, J.; Song, L.; Tong, B.; He, G.; et al. Nr2e1 ablation impairs liver glucolipid metabolism and induces inflammation, high-fat diets amplify the damage. Biomed. Pharmacother. 2019, 120, 109503. [Google Scholar] [CrossRef]
- Lei, Y.-F.; Chen, J.-L.; Wei, H.; Xiong, C.-M.; Zhang, Y.-H.; Ruan, J.-L. Hypolipidemic and anti-inflammatory properties of Abacopterin A from Abacopteris penangiana in high-fat diet-induced hyperlipidemia mice. Food Chem. Toxicol. 2011, 49, 3206–3210. [Google Scholar] [CrossRef]
- Lavrador, M.S.F.; Afonso, M.S.; Cintra, D.E.; Koike, M.; Nunes, V.S.; Demasi, M.; Lin, C.J.; Beda, L.M.M.; Gioielli, L.A.; Bombo, R.d.P.A.; et al. Interesterified Fats Induce Deleterious Effects on Adipose Tissue and Liver in LDLr-KO Mice. Nutrients 2019, 11, 466. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Contents (%) | Compounds | Contents (%) | |
| General nutritional composition | Total dietary fiber | 45.90 | Total triterpenes | 1.48 |
| Total sugar | 25.50 | Reducing sugar | 1.29 | |
| Protein | 15.90 | Soluble dietary fiber | 0.68 | |
| Ash | 9.90 | Soluble sugar | 0.57 | |
| Total flavonoids | 9.22 | Total saponins | 0.50 | |
| Moisture | 6.18 | Total alkaloids | 0.46 | |
| Total polyphenols | 5.04 | Total sterols | 0.31 | |
| Fructose | 4.81 | Glucose | UDa | |
| Mannitol | 4.81 | Lactose | UDa | |
| Fat | 4.70 | Maltose | UDa | |
| Soluble protein | 2.20 | Sucrose | UDa | |
| Amino acids | Glutamic acid | 1.83 | Isoleucine | 0.51 |
| Aspartic acid | 1.15 | Phenylalanine | 0.50 | |
| Leucine | 0.85 | Proline | 0.45 | |
| Lysine | 0.68 | Tryptophan | 0.29 | |
| Alanine | 0.65 | Tyrosine | 0.27 | |
| Valine | 0.61 | Histidine | 0.23 | |
| Arginine | 0.60 | Glutamine | 0.21 | |
| Threonine | 0.60 | Methionine | 0.09 | |
| Glycine | 0.59 | Asparagine | 0.07 | |
| Serine | 0.58 | Cysteine | 0.02 | |
| Fatty acids | C18:2n6c | 1.074 | C18:3n3 | UDb |
| C18:1n9c | 0.479 | C20:1 | UDb | |
| C16:0 | 0.291 | C20:2 | UDb | |
| C18:0 | 0.060 | C20:3n3 | UDb | |
| C22:1n9 | 0.025 | C20:3n6 | UDb | |
| C16:1 | 0.013 | C20:4n6 | UDb | |
| C17:0 | 0.010 | C20:5n3 | UDb | |
| C15:0 | 0.008 | C21:0 | UDb | |
| C24:1 | 0.004 | C22:2 | UDb | |
| C8:0 | UDb | C22:6n3 | UDb | |
| C11:0 | UDb | C23:0 | UDb | |
| C13:0 | UDb | C10:0 | UDc | |
| C14:0 | UDb | C12:0 | UDc | |
| C14:1 | UDb | C18:3n6 | UDc | |
| C15:1 | UDb | C20:0 | UDc | |
| C17:1 | UDb | C22:0 | UDc | |
| C18:1n9t | UDb | C24:0 | UDc | |
| C18:2n6t | UDb | |||
| Compounds | Contents (mg/kg) | Compounds | Contents (mg/kg) | |
| Minerals | K | 4.1 × 103 | Zn | 93.7 |
| Ca | 406.0 | Mn | 24.0 | |
| Fe | 310.0 | Se | 0.2 | |
| Na | 243.0 | |||
| Heavy metals | Cu | 31.000 | As | 0.249 |
| Cr | 30.900 | Pb | 0.188 | |
| Cd | 0.323 | Hg | 0.009 | |
| Vitamins | Vitamin B6 | 5.10 | Vitamin A | UDd |
| Vitamin D2 | 3.07 | Vitamin B1 | UDe | |
| Vitamin E | 2.02 | Vitamin C | UDf | |
| Vitamin B2 | 1.01 | Vitamin D3 | UDg | |
| Nucleotides | Urine purine nucleotides | 1092.13 | Cytosine nucleotides | UDh |
| Adenosine nucleotides | 146.92 | Inosinic acid | UDh | |
| Guanine nucleotides | 44.52 |
| Week | NCD | HFD | HFD + 500 mg/kg IH | HFD + 1000 mg/kg IH | HFD + 3 mg/kg SV | NCD + 500 mg/kg IH | |
|---|---|---|---|---|---|---|---|
| Plasma glucose (mmol/L) | 0 | 8.3 ± 1.6 | 9.8 ± 1.5 | 10.4 ± 1.9 | 10.9 ± 1.9 | 10.5 ± 2.1 | 8.7 ± 0.4 |
| 2 | 8.9 ± 1.3 | 11.9 ± 1.8 ## | 10.3 ± 1.6 | 10.4 ± 0.8 | 10.5 ± 0.7 | 9.2 ± 1.1 | |
| 4 | 8.8 ± 1.6 | 11.5 ± 1.8 # | 9.4 ± 1.4 | 9.8 ± 0.9 | 11.3 ± 1.5 | 9.0 ± 0.7 | |
| 6 | 8.3 ± 1.1 | 12.1 ± 2.1 ### | 9.4 ± 0.6 * | 8.7 ± 1.1 ** | 12.1 ± 1.5 | 8.6 ± 1.0 | |
| 8 | 8.8 ± 0.8 | 12.5 ± 1.1 ## | 11.6 ± 2.7 | 11.6 ± 1.4 | 12.1 ± 1.3 | 8.6 ± 0.4 | |
| Organ indices (%) | Heart | 0.560 ± 0.037 | 0.333 ± 0.055 ### | 0.367 ± 0.044 | 0.353 ± 0.028 | 0.313 ± 0.057 | 0.543 ± 0.053 |
| Liver | 3.465 ± 0.077 | 4.859 ± 0.576 ### | 3.310 ± 0.334 *** | 4.020 ± 0.551 | 4.225 ± 0.745 | 3.922 ± 0.281 | |
| Spleen | 0.259 ± 0.006 | 0.223 ± 0.029 | 0.225 ± 0.038 | 0.219 ± 0.031 | 0.186 ± 0.012 | 0.270 ± 0.011 | |
| Kidney | 1.198 ± 0.073 | 0.717 ± 0.044 ### | 0.800 ± 0.142 | 0.801 ± 0.052 | 0.718 ± 0.043 | 1.199 ± 0.064 | |
| Pancreas | 0.641 ± 0.064 | 0.375 ± 0.048 ### | 0.518 ± 0.127 | 0.437 ± 0.113 | 0.317 ± 0.048 | 0.662 ± 0.067 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Hao, J.; Liu, Z.; Li, Z.; Teng, L.; Wang, D. Inonotus hispidus Protects against Hyperlipidemia by Inhibiting Oxidative Stress and Inflammation through Nrf2/NF-κB Signaling in High Fat Diet Fed Mice. Nutrients 2022, 14, 3477. https://doi.org/10.3390/nu14173477
Zhang Y, Hao J, Liu Z, Li Z, Teng L, Wang D. Inonotus hispidus Protects against Hyperlipidemia by Inhibiting Oxidative Stress and Inflammation through Nrf2/NF-κB Signaling in High Fat Diet Fed Mice. Nutrients. 2022; 14(17):3477. https://doi.org/10.3390/nu14173477
Chicago/Turabian StyleZhang, Yongfeng, Jie Hao, Zijian Liu, Zhige Li, Lirong Teng, and Di Wang. 2022. "Inonotus hispidus Protects against Hyperlipidemia by Inhibiting Oxidative Stress and Inflammation through Nrf2/NF-κB Signaling in High Fat Diet Fed Mice" Nutrients 14, no. 17: 3477. https://doi.org/10.3390/nu14173477
APA StyleZhang, Y., Hao, J., Liu, Z., Li, Z., Teng, L., & Wang, D. (2022). Inonotus hispidus Protects against Hyperlipidemia by Inhibiting Oxidative Stress and Inflammation through Nrf2/NF-κB Signaling in High Fat Diet Fed Mice. Nutrients, 14(17), 3477. https://doi.org/10.3390/nu14173477