
Plasma Phospholipid Fatty Acids and Risk of Venous Thromboembolism: Mendelian Randomization Investigation

 ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
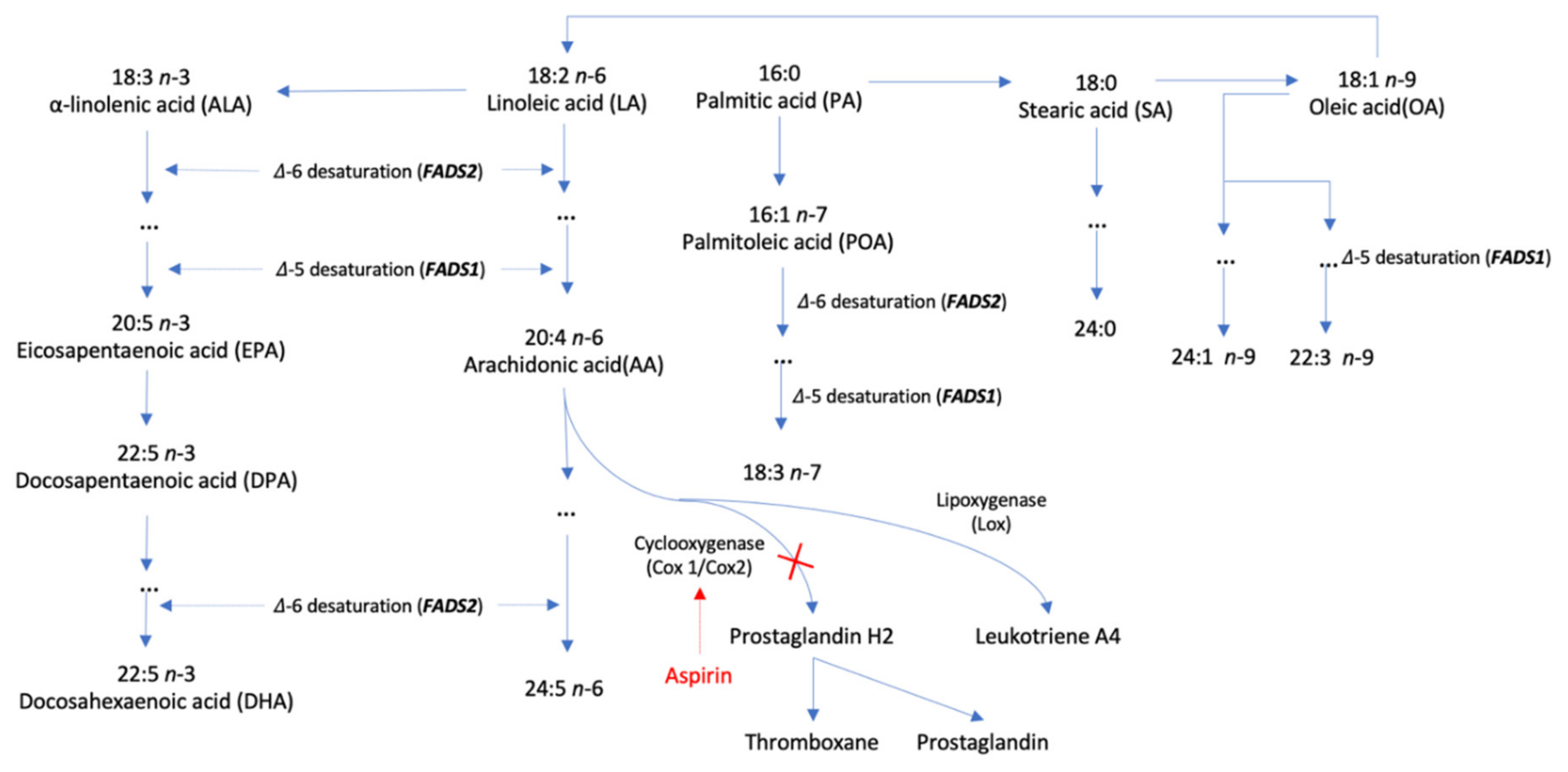
2.1. Genetic Instrument Selection
2.2. Data Sources for Outcomes
2.3. Statistical Analysis
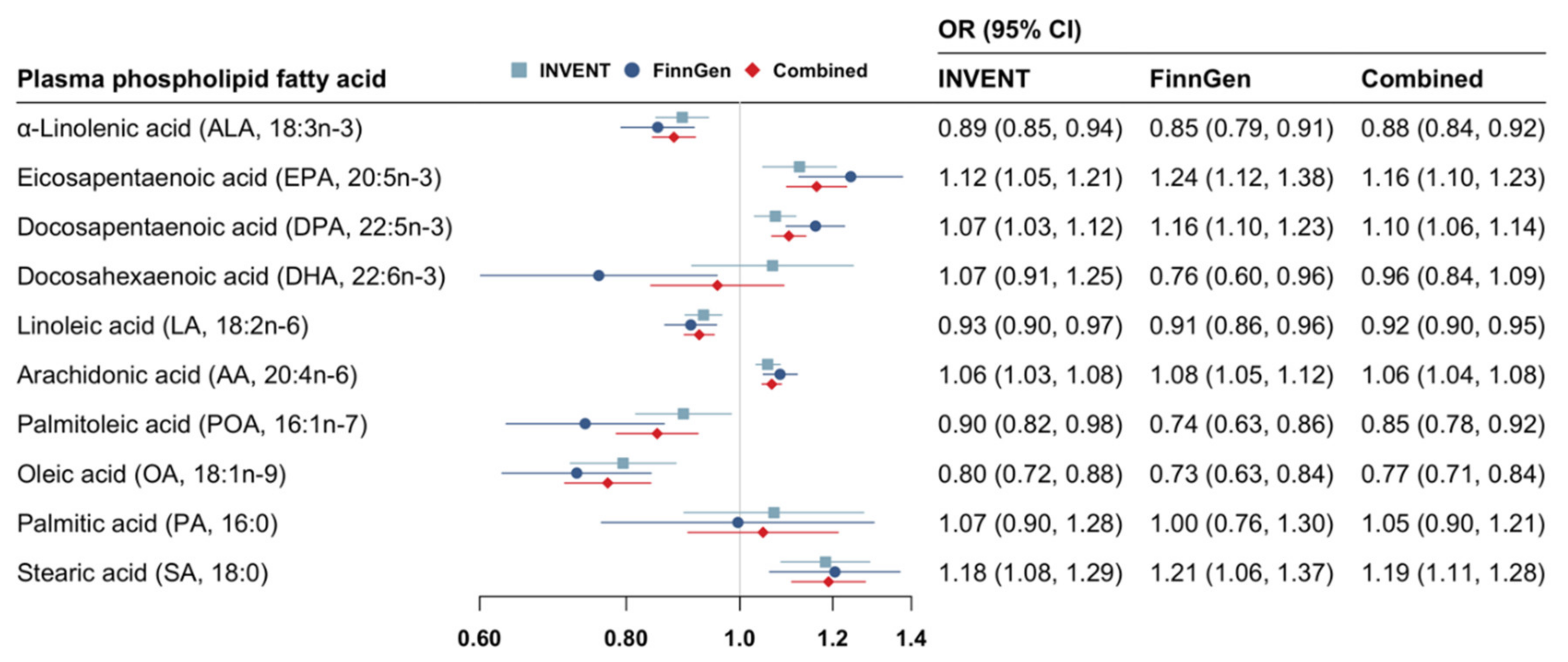
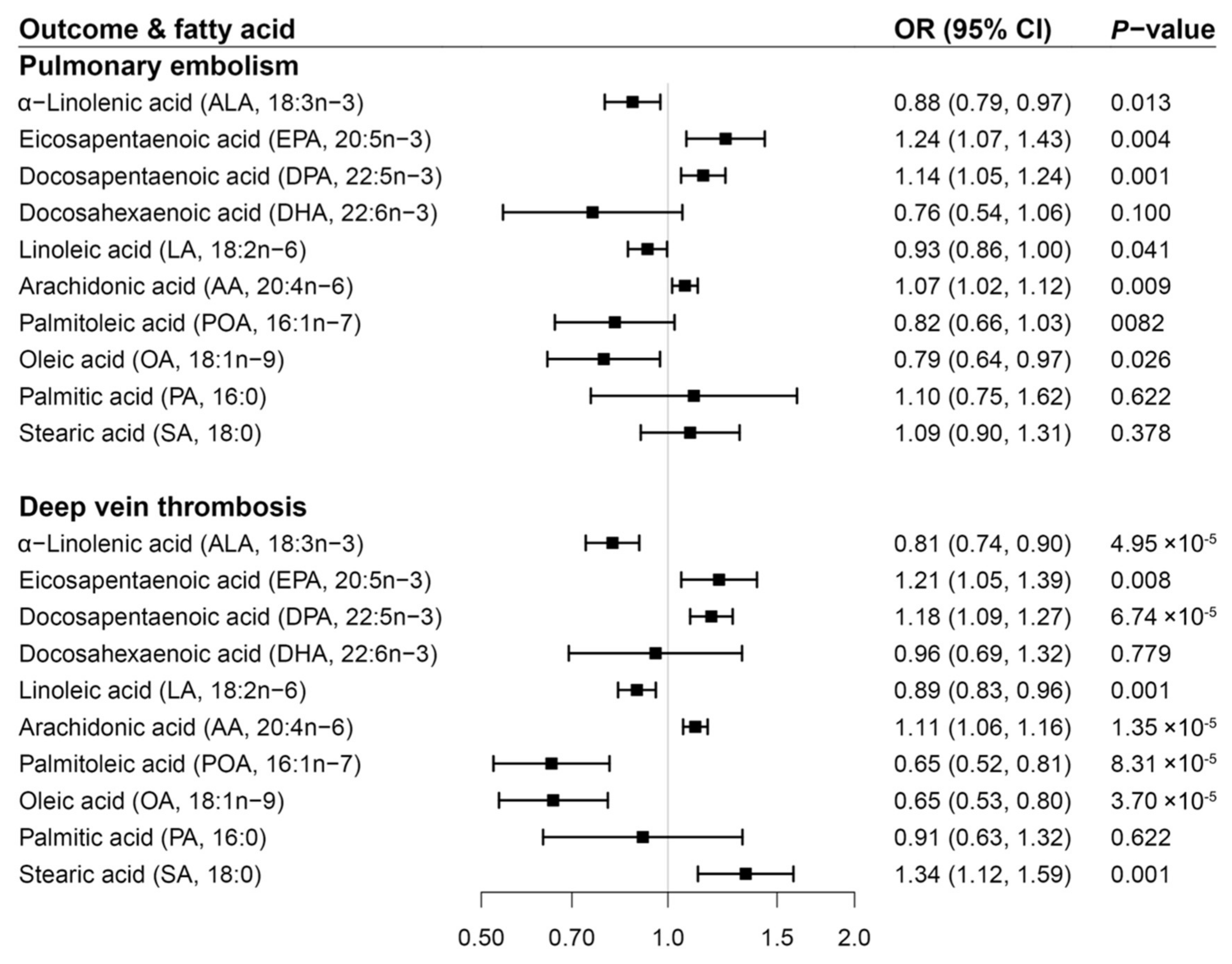
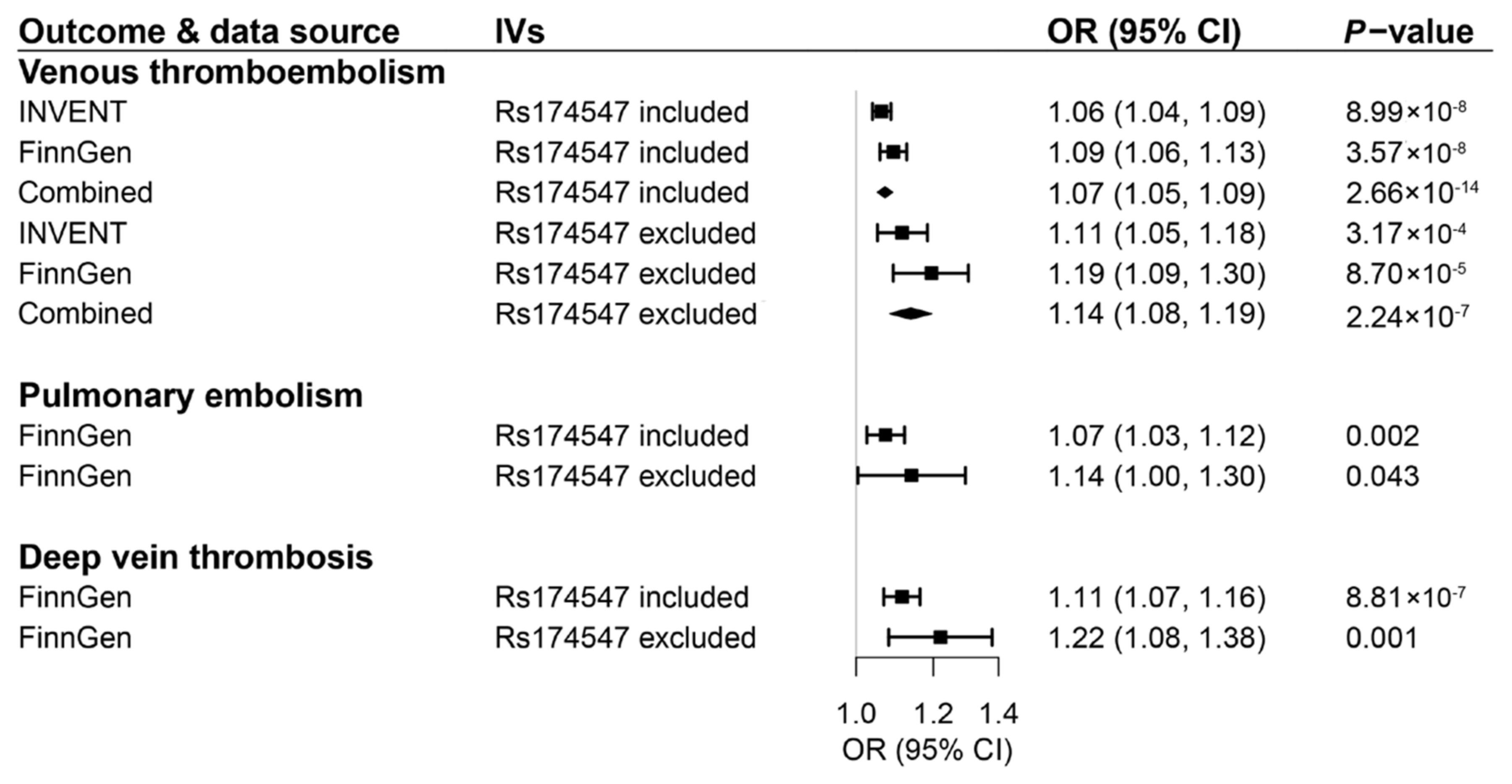
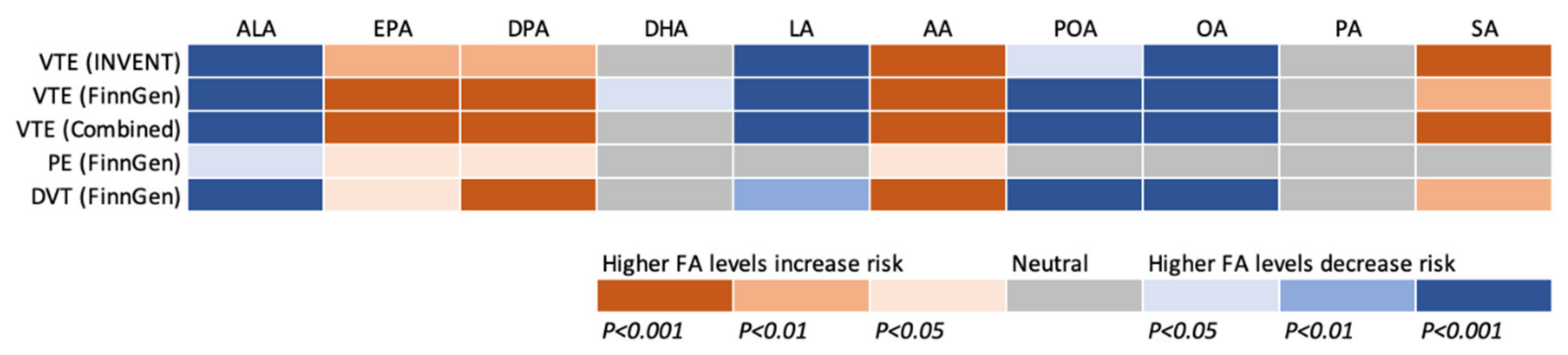
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heit, J.A. Epidemiology of venous thromboembolism. Nat. Rev. Cardiol. 2015, 12, 464–474. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.A., Jr.; Spencer, F.A. Risk factors for venous thromboembolism. Circulation 2003, 107, I9–I16. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Bruzelius, M.; Xiong, Y.; Håkansson, N.; Åkesson, A.; Larsson, S.C. Overall and abdominal obesity in relation to venous thromboembolism. J. Thromb. Haemost. 2021, 19, 460–469. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Bruzelius, M.; Håkansson, N.; Åkesson, A.; Larsson, S.C. Lifestyle factors and venous thromboembolism in two cohort studies. Thromb. Res. 2021, 202, 119–124. [Google Scholar] [CrossRef]
- Yuan, S.; Bruzelius, M.; Damrauer, S.M.; Håkansson, N.; Wolk, A.; Åkesson, A.; Larsson, S.C. Anti-inflammatory diet and venous thromboembolism: Two prospective cohort studies. Nutr. Metab. Cardiovasc. Dis. 2021, 31, 2831–2838. [Google Scholar] [CrossRef]
- Zhang, Y.; Ding, J.; Guo, H.; Liang, J.; Li, Y. Associations of Fish and Omega-3 Fatty Acids Consumption with the Risk of Venous Thromboembolism. A Meta-Analysis of Prospective Cohort Studies. Front. Nutr. 2020, 7, 614784. [Google Scholar] [CrossRef]
- Isaksen, T.; Evensen, L.H.; Brækkan, S.K.; Hansen, J.B. Dietary Intake of Marine Polyunsaturated n-3 Fatty Acids and Risk of Recurrent Venous Thromboembolism. Thromb. Haemost. 2019, 119, 2053–2063. [Google Scholar] [CrossRef]
- Zheng, X.; Jia, R.; Li, Y.; Liu, T.; Wang, Z. Omega-3 fatty acids reduce post-operative risk of deep vein thrombosis and pulmonary embolism after surgery for elderly patients with proximal femoral fractures: A randomized placebo-controlled, double-blind clinical trial. Int. Orthop. 2020, 44, 2089–2093. [Google Scholar] [CrossRef]
- Anderson, B.M.; Ma, D.W. Are all n-3 polyunsaturated fatty acids created equal? Lipids Health Dis. 2009, 8, 33. [Google Scholar] [CrossRef]
- Reiner, M.F.; Martinod, K.; Stivala, S.; Savarese, G.; Camici, G.G.; Lüscher, T.F.; Wagner, D.D.; Beer, J.H. Dietary omega-3 alpha-linolenic acid does not prevent venous thrombosis in mice. Thromb. Haemost. 2015, 113, 177–184. [Google Scholar] [CrossRef]
- Holy, E.W.; Forestier, M.; Richter, E.K.; Akhmedov, A.; Leiber, F.; Camici, G.G.; Mocharla, P.; Lüscher, T.F.; Beer, J.H.; Tanner, F.C. Dietary α-linolenic acid inhibits arterial thrombus formation, tissue factor expression, and platelet activation. Arter. Thromb. Vasc. Biol. 2011, 31, 1772–1780. [Google Scholar] [CrossRef]
- Li, D.; Sinclair, A.; Wilson, A.; Nakkote, S.; Kelly, F.; Abedin, L.; Mann, N.; Turner, A. Effect of dietary alpha-linolenic acid on thrombotic risk factors in vegetarian men. Am. J. Clin. Nutr. 1999, 69, 872–882. [Google Scholar] [CrossRef]
- Nelson, G.J.; Schmidt, P.S.; Bartolini, G.L.; Kelley, D.S.; Kyle, D. The effect of dietary docosahexaenoic acid on platelet function, platelet fatty acid composition, and blood coagulation in humans. Lipids 1997, 32, 1129–1136. [Google Scholar] [CrossRef]
- Frelinger, A.L., III; Furman, M.I.; Linden, M.D.; Li, Y.; Fox, M.L.; Barnard, M.R.; Michelson, A.D. Residual arachidonic acid-induced platelet activation via an adenosine diphosphate-dependent but cyclooxygenase-1- and cyclooxygenase-2-independent pathway: A 700-patient study of aspirin resistance. Circulation 2006, 113, 2888–2896. [Google Scholar] [CrossRef]
- Pacheco, Y.M.; Bermúdez, B.; López, S.; Abia, R.; Villar, J.; Muriana, F.J. Ratio of oleic to palmitic acid is a dietary determinant of thrombogenic and fibrinolytic factors during the postprandial state in men. Am. J. Clin. Nutr. 2006, 84, 342–349. [Google Scholar] [CrossRef]
- Kelly, F.D.; Sinclair, A.J.; Mann, N.J.; Turner, A.H.; Abedin, L.; Li, D. A stearic acid-rich diet improves thrombogenic and atherogenic risk factor profiles in healthy males. Eur. J. Clin. Nutr. 2001, 55, 88–96. [Google Scholar] [CrossRef]
- Burgess, S.; Thompson, S.G. Mendelian Randomization: Methods for Using Genetic Variants in Causal Estimation; Chapman and Hall/CRC: London, UK, 2015. [Google Scholar]
- Lemaitre, R.N.; Tanaka, T.; Tang, W.; Manichaikul, A.; Foy, M.; Kabagambe, E.K.; Nettleton, J.A.; King, I.B.; Weng, L.C.; Bhattacharya, S.; et al. Genetic loci associated with plasma phospholipid n-3 fatty acids: A meta-analysis of genome-wide association studies from the CHARGE Consortium. PLoS Genet. 2011, 7, e1002193. [Google Scholar] [CrossRef]
- Guan, W.; Steffen, B.T.; Lemaitre, R.N.; Wu, J.H.Y.; Tanaka, T.; Manichaikul, A.; Foy, M.; Rich, S.S.; Wang, L.; Nettleton, J.A.; et al. Genome-wide association study of plasma N6 polyunsaturated fatty acids within the cohorts for heart and aging research in genomic epidemiology consortium. Circ. Cardiovasc. Genet. 2014, 7, 321–331. [Google Scholar] [CrossRef]
- Wu, J.H.; Lemaitre, R.N.; Manichaikul, A.; Guan, W.; Tanaka, T.; Foy, M.; Kabagambe, E.K.; Djousse, L.; Siscovick, D.; Fretts, A.M.; et al. Genome-wide association study identifies novel loci associated with concentrations of four plasma phospholipid fatty acids in the de novo lipogenesis pathway: Results from the Cohorts for Heart and Aging Research in Genomic Epidemiology (CHARGE) consortium. Circ. Cardiovasc. Genet. 2013, 6, 171–183. [Google Scholar] [CrossRef]
- Yuan, S.; Larsson, S.C. Association of genetic variants related to plasma fatty acids with type 2 diabetes mellitus and glycaemic traits: A Mendelian randomisation study. Diabetologia 2020, 63, 116–123. [Google Scholar] [CrossRef]
- Yamagishi, K.; Nettleton, J.A.; Folsom, A.R. Plasma fatty acid composition and incident heart failure in middle-aged adults: The Atherosclerosis Risk in Communities (ARIC) Study. Am. Heart J. 2008, 156, 965–974. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, J.V.; Schooling, C.M. The associations of plasma phospholipid arachidonic acid with cardiovascular diseases: A Mendelian randomization study. EBioMedicine 2021, 63, 103189. [Google Scholar] [CrossRef]
- Lindström, S.; Wang, L.; Smith, E.N.; Gordon, W.; van Hylckama Vlieg, A.; de Andrade, M.; Brody, J.A.; Pattee, J.W.; Haessler, J.; Brumpton, B.M.; et al. Genomic and transcriptomic association studies identify 16 novel susceptibility loci for venous thromboembolism. Blood 2019, 134, 1645–1657. [Google Scholar] [CrossRef]
- FinnGen. R6 Release of the FinnGen Study. Available online: https://finngen.gitbook.io/documentation/ (accessed on 1 March 2022).
- Burgess, S.; Davies, N.M.; Thompson, S.G. Bias due to participant overlap in two-sample Mendelian randomization. Genet. Epidemiol. 2016, 40, 597–608. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent Estimation in Mendelian Randomization with Some Invalid Instruments Using a Weighted Median Estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Brion, M.J.; Shakhbazov, K.; Visscher, P.M. Calculating statistical power in Mendelian randomization studies. Int. J. Epidemiol. 2013, 42, 1497–1501. [Google Scholar] [CrossRef]
- Spiller, W.; Davies, N.M.; Palmer, T.M. Software application profile: Mrrobust—A tool for performing two-sample summary Mendelian randomization analyses. Int. J. Epidemiol. 2019, 48, 684–690. [Google Scholar] [CrossRef]
- Nordoy, A.; Bonaa, K.H.; Sandset, P.M.; Hansen, J.B.; Nilsen, H. Effect of omega-3 fatty acids and simvastatin on hemostatic risk factors and postprandial hyperlipemia in patients with combined hyperlipemia. Arter. Thromb. Vasc. Biol. 2000, 20, 259–265. [Google Scholar] [CrossRef]
- Ramberg, C.; Hindberg, K.; Biedermann, J.S.; Cannegieter, S.C.; van der Meer, F.J.; Snir, O.; Leebeek, F.W.G.; Kruip, M.; Hansen, J.B.; Lijfering, W.M. Rosuvastatin treatment decreases plasma procoagulant phospholipid activity after a VTE: A randomized controlled trial. J. Thromb. Haemost. 2022, 20, 877–887. [Google Scholar] [CrossRef]
- Gajos, G.; Zalewski, J.; Rostoff, P.; Nessler, J.; Piwowarska, W.; Undas, A. Reduced thrombin formation and altered fibrin clot properties induced by polyunsaturated omega-3 fatty acids on top of dual antiplatelet therapy in patients undergoing percutaneous coronary intervention (OMEGA-PCI clot). Arter. Thromb. Vasc. Biol. 2011, 31, 1696–1702. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, S.M.; Ali, M.A.; Churchill, D.N. Effect of n-3 fatty acids from fish oil on hemostasis, blood pressure, and lipid profile of dialysis patients. J. Am. Soc. Nephrol. 1992, 2, 1634–1639. [Google Scholar] [CrossRef] [PubMed]
- Chiva-Blanch, G.; Bratseth, V.; Laake, K.; Arnesen, H.; Solheim, S.; Schmidt, E.B.; Badimon, L.; Seljeflot, I. One year of omega 3 polyunsaturated fatty acid supplementation does not reduce circulating prothrombotic microvesicles in elderly subjects after suffering a myocardial infarction. Clin. Nutr. 2021, 40, 5674–5677. [Google Scholar] [CrossRef] [PubMed]
- Nieuwenhuys, C.M.; Hornstra, G. The effects of purified eicosapentaenoic and docosahexaenoic acids on arterial thrombosis tendency and platelet function in rats. Biochim. Biophys. Acta 1998, 1390, 313–322. [Google Scholar] [CrossRef]
- Dyerberg, J.; Bang, H.O.; Stoffersen, E.; Moncada, S.; Vane, J.R. Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 1978, 2, 117–119. [Google Scholar] [CrossRef]
- Yuan, S.; Bäck, M.; Bruzelius, M.; Mason, A.M.; Burgess, S.; Larsson, S. Plasma Phospholipid Fatty Acids, FADS1 and Risk of 15 Cardiovascular Diseases: A Mendelian Randomisation Study. Nutrients 2019, 11, 3001. [Google Scholar] [CrossRef]
- Fuke, G.; Nornberg, J.L. Systematic evaluation on the effectiveness of conjugated linoleic acid in human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1–7. [Google Scholar] [CrossRef]
- Jones, C.M.; Hall, E.R.; Hester, J.P.; Wu, K.K. Arachidonic acid metabolites produced by platelet-depleted human blood monocytes: A possible role in thrombogenesis. Am. J. Hematol. 1989, 31, 145–152. [Google Scholar] [CrossRef]
- Etscheidt, J.; Shahien, A.; Gainey, M.; Kronenfeld, D.; Niu, R.; Freccero, D.; Smith, E. Review of Therapeutic Options for the Prevention of VTE in Total Joint Arthroplasty. Geriatrics 2020, 5, 18. [Google Scholar] [CrossRef]
- Simes, J.; Becattini, C.; Agnelli, G.; Eikelboom, J.W.; Kirby, A.C.; Mister, R.; Prandoni, P.; Brighton, T.A. Aspirin for the prevention of recurrent venous thromboembolism: The INSPIRE collaboration. Circulation 2014, 130, 1062–1071. [Google Scholar] [CrossRef]
- Badimon, L.; Vilahur, G.; Rocca, B.; Patrono, C. The key contribution of platelet and vascular arachidonic acid metabolism to the pathophysiology of atherothrombosis. Cardiovasc. Res. 2021, 117, 2001–2015. [Google Scholar] [CrossRef]
- Yuan, S.; Burgess, S.; Laffan, M.; Mason, A.M.; Dichgans, M.; Gill, D.; Larsson, S.C. Genetically Proxied Inhibition of Coagulation Factors and Risk of Cardiovascular Disease: A Mendelian Randomization Study. J. Am. Heart Assoc. 2021, 10, e019644. [Google Scholar] [CrossRef]
- Kelly, F.D.; Mann, N.J.; Turner, A.H.; Sinclair, A.J. Stearic acid-rich diets do not increase thrombotic risk factors in healthy males. Lipids 1999, 34, S199. [Google Scholar] [CrossRef]
- Sanders, T.A.; Oakley, F.R.; Cooper, J.A.; Miller, G.J. Influence of a stearic acid-rich structured triacylglycerol on postprandial lipemia, factor VII concentrations, and fibrinolytic activity in healthy subjects. Am. J. Clin. Nutr. 2001, 73, 715–721. [Google Scholar] [CrossRef]
- Thijssen, M.A.; Hornstra, G.; Mensink, R.P. Stearic, oleic, and linoleic acids have comparable effects on markers of thrombotic tendency in healthy human subjects. J. Nutr. 2005, 135, 2805–2811. [Google Scholar] [CrossRef]
- Sahebkar, A.; Simental-Mendía, L.E.; Pedone, C.; Ferretti, G.; Nachtigal, P.; Bo, S.; Derosa, G.; Maffioli, P.; Watts, G.F. Statin therapy and plasma free fatty acids: A systematic review and meta-analysis of controlled clinical trials. Br. J. Clin. Pharmacol. 2016, 81, 807–818. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, S.; Li, X.; Morange, P.-E.; Bruzelius, M.; Larsson, S.C.; on behalf of the INVENT Consortium. Plasma Phospholipid Fatty Acids and Risk of Venous Thromboembolism: Mendelian Randomization Investigation. Nutrients 2022, 14, 3354. https://doi.org/10.3390/nu14163354
Yuan S, Li X, Morange P-E, Bruzelius M, Larsson SC, on behalf of the INVENT Consortium. Plasma Phospholipid Fatty Acids and Risk of Venous Thromboembolism: Mendelian Randomization Investigation. Nutrients. 2022; 14(16):3354. https://doi.org/10.3390/nu14163354
Chicago/Turabian StyleYuan, Shuai, Xue Li, Pierre-Emmanuel Morange, Maria Bruzelius, Susanna C. Larsson, and on behalf of the INVENT Consortium. 2022. "Plasma Phospholipid Fatty Acids and Risk of Venous Thromboembolism: Mendelian Randomization Investigation" Nutrients 14, no. 16: 3354. https://doi.org/10.3390/nu14163354
APA StyleYuan, S., Li, X., Morange, P.-E., Bruzelius, M., Larsson, S. C., & on behalf of the INVENT Consortium. (2022). Plasma Phospholipid Fatty Acids and Risk of Venous Thromboembolism: Mendelian Randomization Investigation. Nutrients, 14(16), 3354. https://doi.org/10.3390/nu14163354