
Luteolin-7-O-Glucuronide Improves Depression-like and Stress Coping Behaviors in Sleep Deprivation Stress Model by Activation of the BDNF Signaling

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
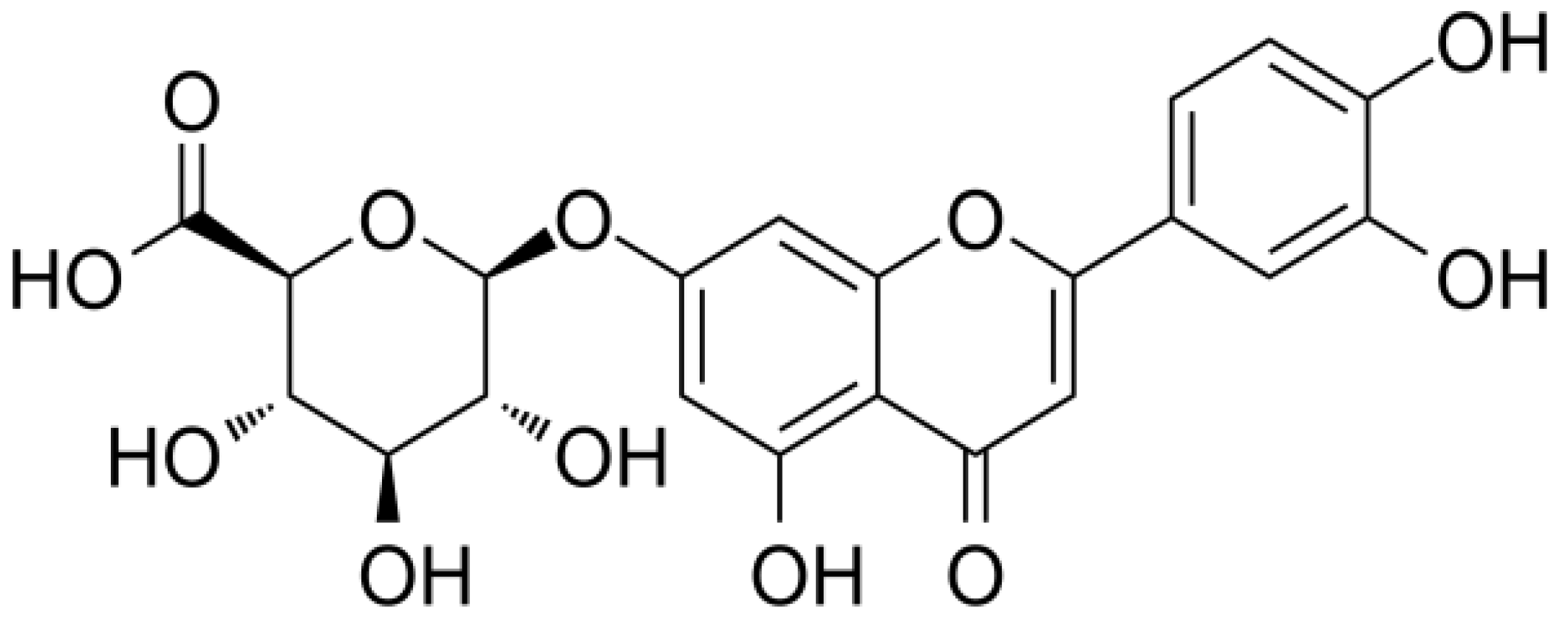
2.2. Isolation of L7Gn from Perilla frutescens Extarct
2.3. Animals
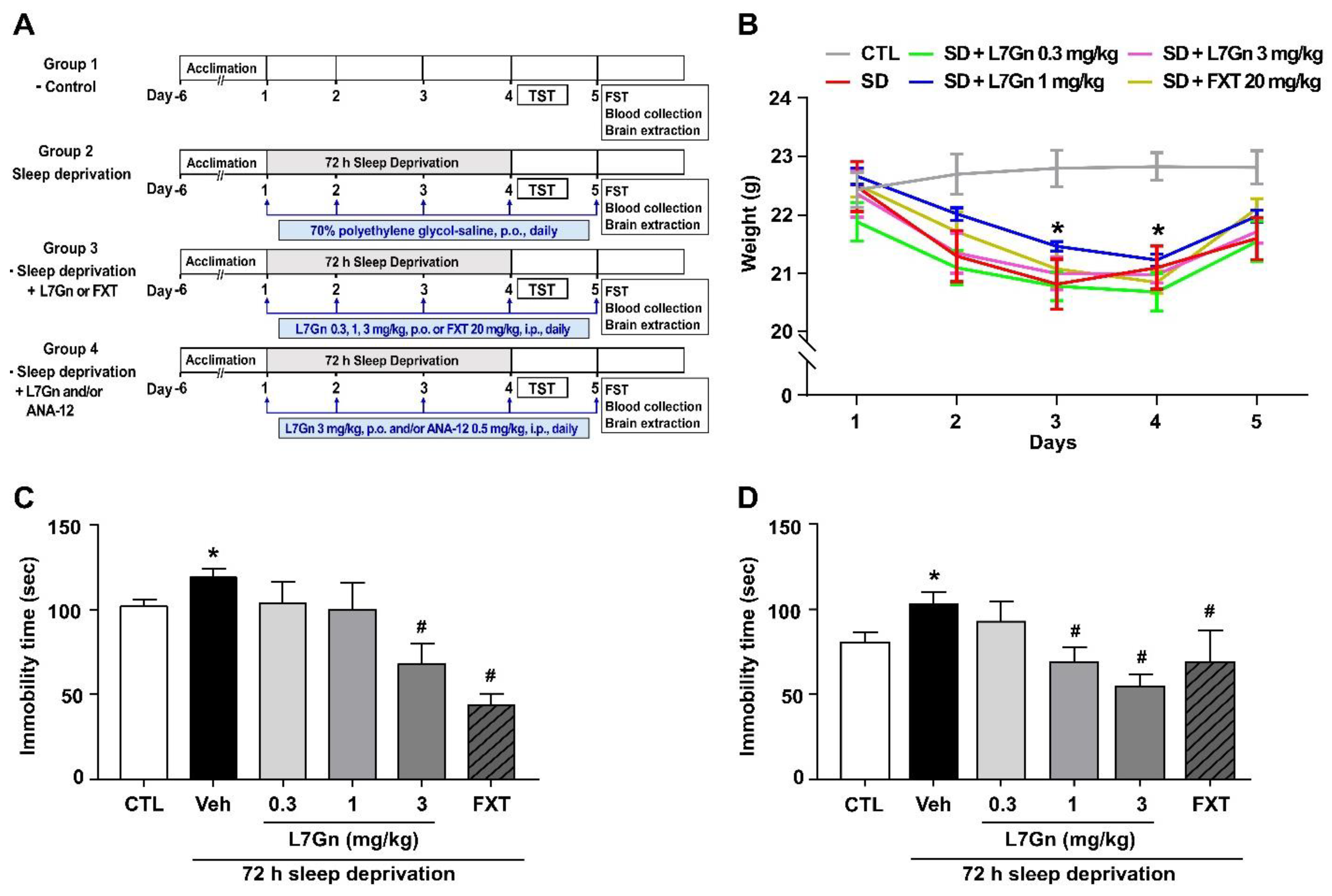
2.4. SD Model
2.5. TST
2.6. FST
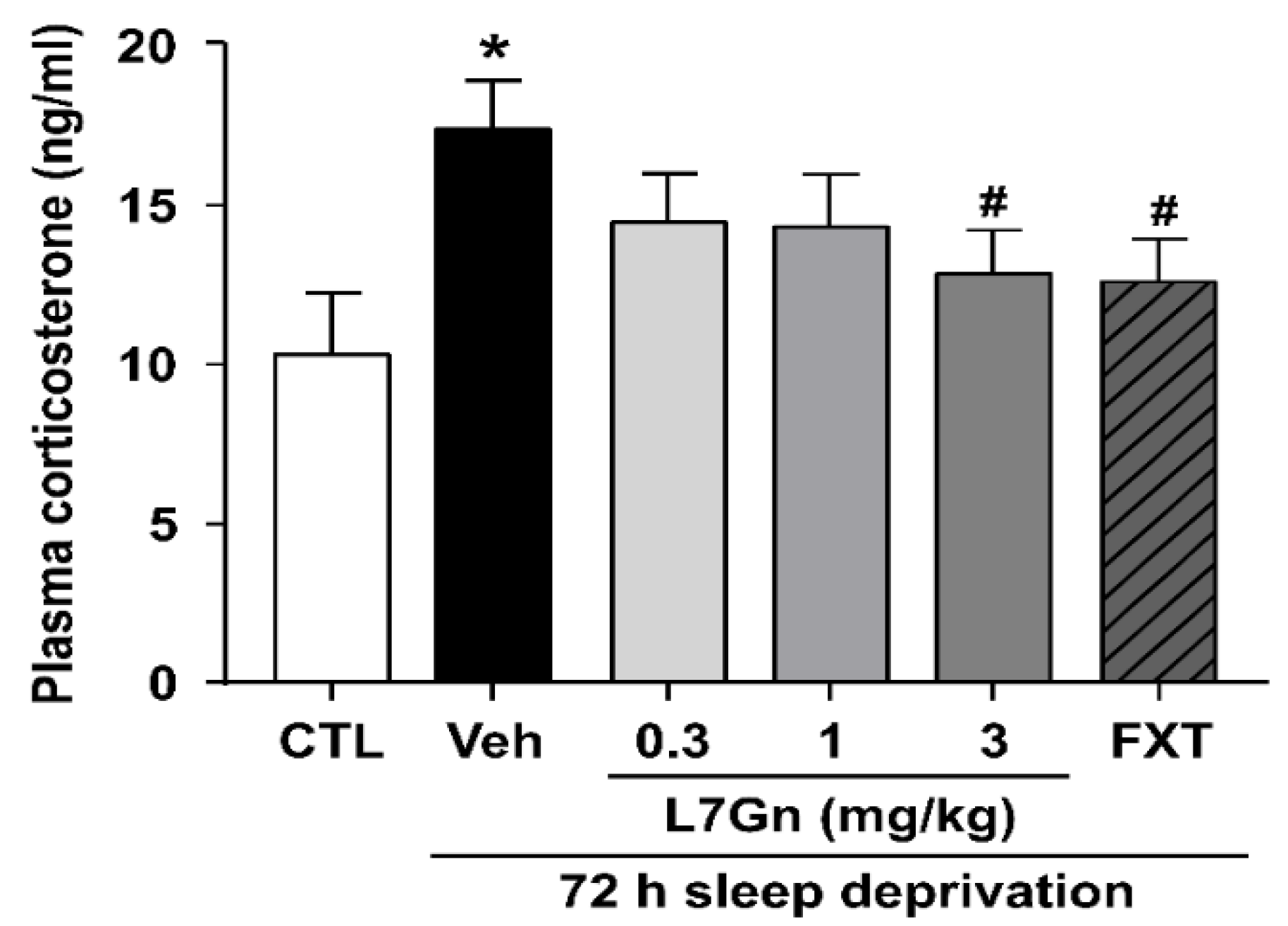
2.7. Plasma Corticosterone Level
2.8. Western Blot Analysis
2.9. Quantitative Reverse Transcription-PCR (qRT-PCR)
2.10. Statistical Analysis
3. Results
3.1. Effect of L7Gn on Depression-like and Stress Coping Behavior in the SD Stress Mouse Model
3.2. Effect of L7Gn Treatment on Plasma Corticosterone Level in Sleep-Deprived Mice
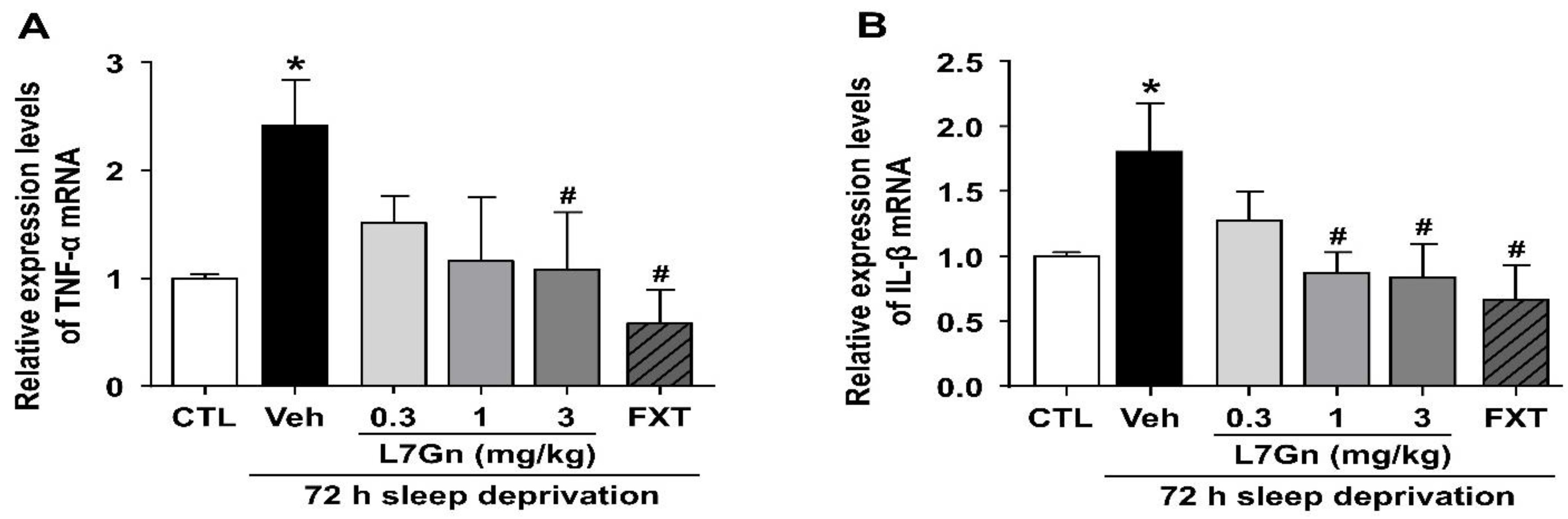
3.3. Effect of L7Gn on TNF-α and IL-1β Levels in Sleep-Deprived Mice
3.4. Effect of L7Gn on BDNF mRNA Expression in the Hippocampus of Sleep-Deprived Mice
3.5. Effect of L7Gn on the BDNF/TrkB/ERK/CREB Signaling Molecules in the Hippocampus of Sleep-Deprived Mice
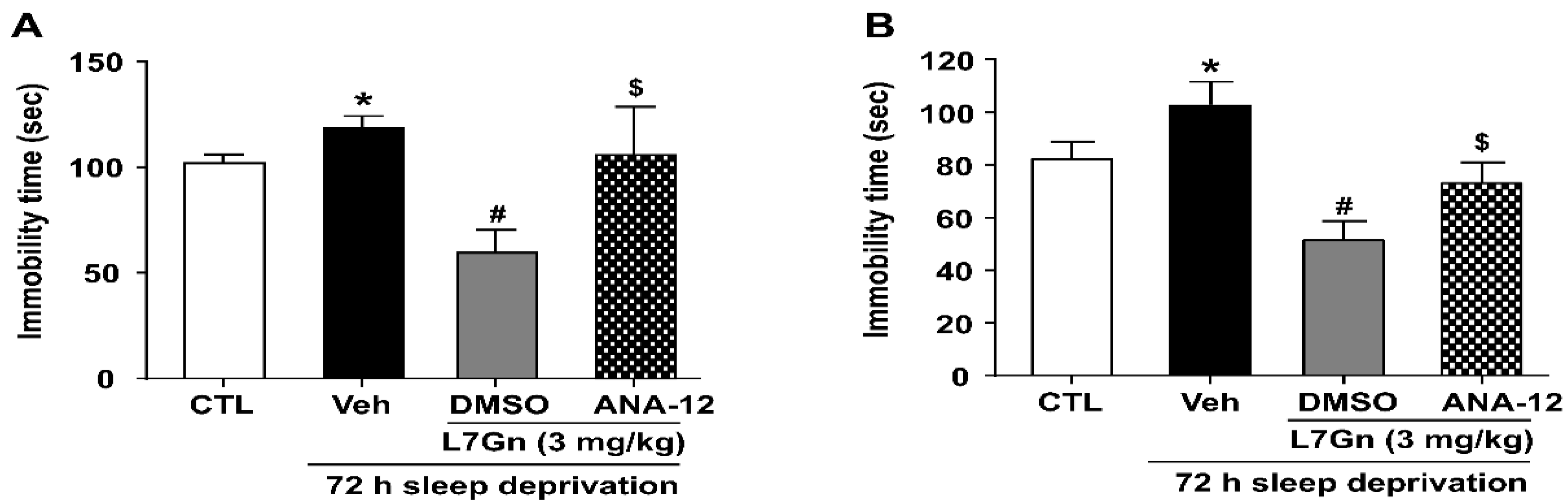
3.6. Effect of ANA-12 on the Antidepressant Effect and Active Stress Coping Response Induced by L7Gn
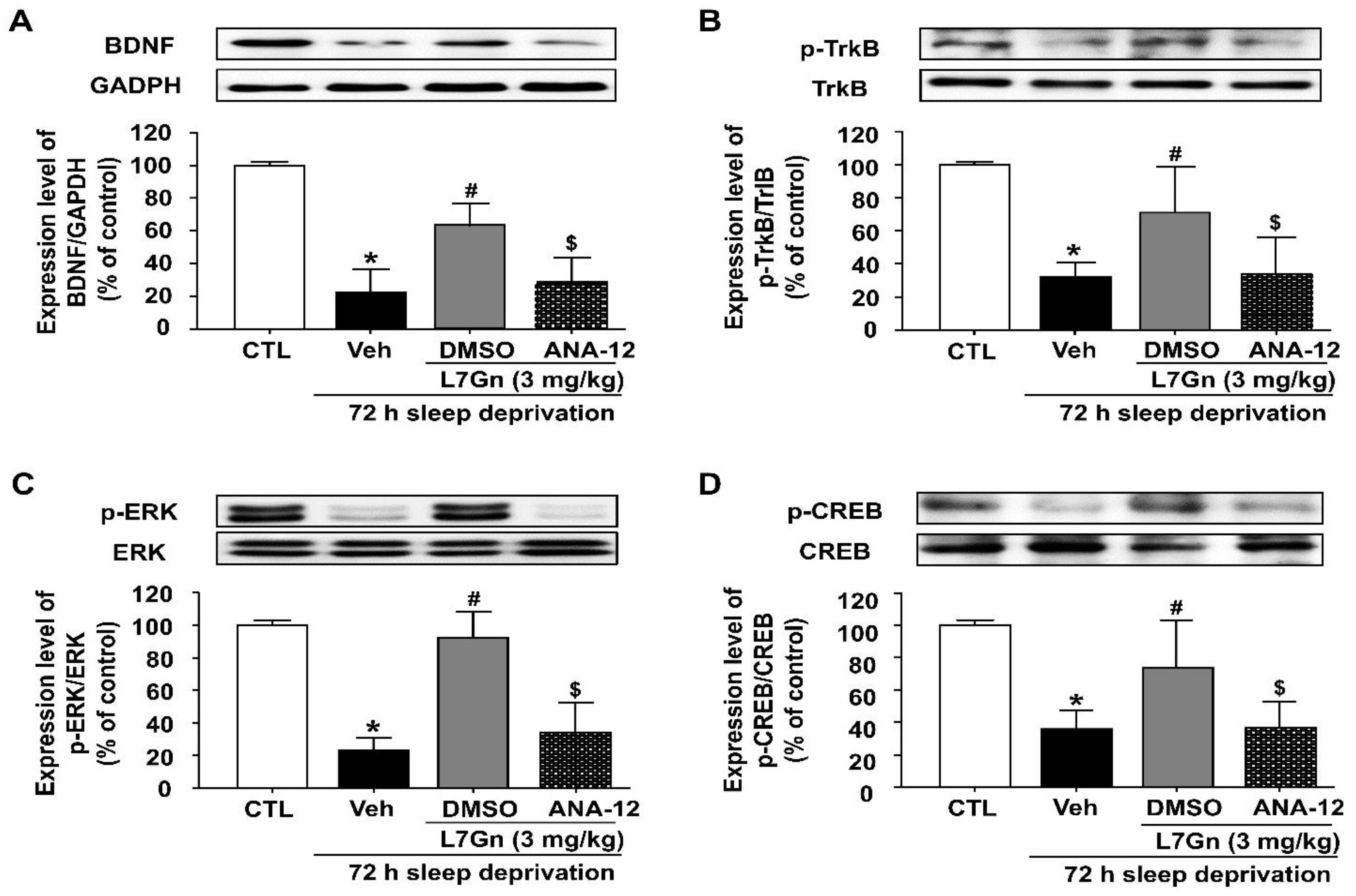
3.7. Effect of ANA-12 Treatment on the L7Gn-Induced Upregulation of BDNF, p-TrkB, p-CREB and p-ERK
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seo, J.-S.; Wei, J.; Qin, L.; Kim, Y.; Yan, Z.; Greengard, P. Cellular and molecular basis for stress-induced depression. Mol. Psychiatry 2017, 22, 1440–1447. [Google Scholar] [CrossRef] [PubMed]
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- Smith, K.; De Torres, I.J.N. A world of depression. Nature 2014, 515, 180–181. [Google Scholar] [CrossRef] [PubMed]
- Hammen, C. Stress and depression. Annu. Rev. Clin. Psychol. 2005, 1, 293–319. [Google Scholar] [CrossRef]
- Franzen, P.L.; Buysse, D.J. Sleep disturbances and depression: Risk relationships for subsequent depression and therapeutic implications. Dialogues Clin. Neurosci. 2022, 10, 473–481. [Google Scholar] [CrossRef]
- Geoffroy, P.A. Clock Genes and Light Signaling Alterations in Bipolar Disorder: When the Biological Clock Is Off. Biol. Psychiatry 2018, 84, 775–777. [Google Scholar] [CrossRef]
- Geoffroy, P.A.; Hoertel, N.; Etain, B.; Bellivier, F.; Delorme, R.; Limosin, F.; Peyre, H. Insomnia and hypersomnia in major depressive episode: Prevalence, sociodemographic characteristics and psychiatric comorbidity in a population-based study. J. Affect. Disord. 2018, 226, 132–141. [Google Scholar] [CrossRef]
- Gruber, R.; Cassoff, J. The interplay between sleep and emotion regulation: Conceptual framework empirical evidence and future directions. Curr. Psychiatry Rep. 2014, 16, 500. [Google Scholar] [CrossRef]
- Dimitrov, S.; Lange, T.; Tieken, S.; Fehm, H.L.; Born, J. Sleep associated regulation of T helper 1/T helper 2 cytokine balance in humans. Brain Behav. Immun. 2004, 18, 341–348. [Google Scholar] [CrossRef]
- Scheer, F.A.J.L.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef]
- Andersen, M.L.; Martins, P.J.F.; D’Almeida, V.; Bignotto, M.; Tufik, S. Endocrinological and catecholaminergic alterations during sleep deprivation and recovery in male rats. J. Sleep Res. 2005, 14, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Mirescu, C.; Peters, J.D.; Noiman, L.; Gould, E. Sleep deprivation inhibits adult neurogenesis in the hippocampus by elevating glucocorticoids. Proc. Natl. Acad. Sci. USA 2006, 103, 19170–19175. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.S.; Park, H.J.; Chung, J.-H.; Kim, J.W. REM sleep deprivation increases the expression of interleukin genes in mice hypothalamus. Neurosci. Lett. 2013, 556, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Havekes, R.; Park, A.J.; Tudor, J.C.; Luczak, V.G.; Hansen, R.T.; Ferri, S.L.; Bruinenberg, V.M.; Poplawski, S.G.; Day, J.P.; Aton, S.J.; et al. Sleep deprivation causes memory deficits by negatively impacting neuronal connectivity in hippocampal area CA1. eLife 2016, 5, e13424. [Google Scholar] [CrossRef]
- D'Sa, C.; Duman, R.S. Antidepressants and neuroplasticity. Bipolar Disord. 2002, 4, 183–194. [Google Scholar] [CrossRef]
- Park, H.; Poo, M.-M. Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 2013, 14, 7–23. [Google Scholar] [CrossRef]
- Hashimoto, K. Brain-derived neurotrophic factor as a biomarker for mood disorders: An historical overview and future directions. Psychiatry Clin. Neurosci. 2010, 64, 341–357. [Google Scholar] [CrossRef]
- Zhang, J.-C.; Yao, W.; Hashimoto, K. Brain-derived Neurotrophic Factor (BDNF)-TrkB Signaling in Inflammation-related Depression and Potential Therapeutic Targets. Curr. Neuropharmacol. 2016, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Jeon, I.H.; Kim, H.S.; Kang, H.J.; Lee, H.-S.; Jeong, S.I.; Kim, S.J.; Jang, S.I. Anti-inflammatory and antipruritic effects of luteolin from Perilla (P. frutescens L.) leaves. Molecules 2014, 19, 6941–6951. [Google Scholar] [CrossRef]
- Karki, S.; Park, H.J.; Nugroho, A.; Kim, E.J.; Jung, H.A.; Choi, J.S. Quantification of major compounds from Ixeris dentata, Ixeris dentata Var. albiflora, and Ixeris sonchifolia and their comparative anti-inflammatory activity in lipopolysaccharide-stimulated RAW 264.7 cells. J. Med. Food 2015, 18, 83–94. [Google Scholar] [CrossRef]
- Kim, E.; Yoon, K.D.; Lee, W.S.; Yang, W.S.; Kim, S.H.; Sung, N.Y.; Baek, K.-S.; Kim, Y.; Htwe, K.M.; Kim, Y.-D.; et al. Syk/Src-targeted anti-inflammatory activity of Codariocalyx motorius ethanolic extract. J. Ethnopharmacol. 2014, 155, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Rabelo, A.S.; Oliveira, I.D.; Guimarães, A.G.; Quintans, J.S.; Prata, A.P.; Gelain, D.P.; Venceslau, E.M.; Santos, J.P.; Quintans-Júnior, L.J.; Bonjardim, L.R.; et al. Antinociceptive, anti-inflammatory and antioxidant activities of aqueous extract from Remirea maritima (Cyperaceae). J. Ethnopharmacol. 2013, 145, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Petrović, S.; Ušjak, L.; Milenković, M.; Arsenijević, J.; Drobac, M.; Drndarević, A.; Niketić, M. Thymus dacicus as a new source of antioxidant and antimicrobial metabolites. J. Funct. Foods 2017, 28, 114–121. [Google Scholar] [CrossRef]
- Orhan, F.; Ceker, S.; Anar, M.; Agar, G.; Arasoglu, T.; Gulluce, M. Protective effects of three luteolin derivatives on aflatoxin B1-induced genotoxicity on human blood cells. Med. Chem. Res. 2016, 25, 2567–2577. [Google Scholar] [CrossRef]
- Jeyadevi, R.; Sivasudha, T.; Rameshkumar, A.; Kumar, L.D. Anti-arthritic activity of the Indian leafy vegetable Cardiospermum halicacabum in Wistar rats and UPLC–QTOF–MS/MS identification of the putative active phenolic components. Inflamm. Res. 2013, 62, 115–126. [Google Scholar] [CrossRef]
- Ma, Q.; Jiang, J.-G.; Zhang, X.-M.; Zhu, W. Identification of luteolin 7-O-β-D-glucuronide from Cirsium japonicum and its anti-inflammatory mechanism. J. Funct. Foods 2018, 46, 521–528. [Google Scholar] [CrossRef]
- Cho, Y.-C.; Park, J.; Cho, S. Anti-Inflammatory and Anti-Oxidative Effects of luteolin-7-O-glucuronide in LPS-Stimulated Murine Macrophages through TAK1 Inhibition and Nrf2 Activation. Int. J. Mol. Sci. 2020, 21, 2007. [Google Scholar] [CrossRef]
- Kim, J.; Choi, H.; Kim, M.-R.; Oh, D.-R.; Kim, Y.; Kang, H.; Jeong, K.-i.; Ryu, G.-c.; Pan, S.; Choi, C.-y. Amelioration of Visual Display Terminal-induced Ocular Fatigue by Aqueous Extracts of Perilla frutescens var. acuta. J. Food Nutr. Res. 2017, 5, 553–561. [Google Scholar] [CrossRef]
- Silva, R.H.; Abílio, V.C.; Takatsu, A.; Kameda, S.R.; Grassl, C.; Chehin, A.B.; Medrano, W.A.; Calzavara, M.B.; Registro, S.; Andersen, M.L. Role of hippocampal oxidative stress in memory deficits induced by sleep deprivation in mice. Neuropharmacology 2004, 46, 895–903. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Molendijk, M.L. Floating Rodents and Stress-Coping Neurobiology. Biol. Psychiatry 2021, 90, e19–e21. [Google Scholar] [CrossRef]
- Xu, X.; Zheng, P.; Zhao, H.; Song, B.; Wang, F. Effect of Electroacupuncture at GV20 on Sleep Deprivation-Induced Depression-Like Behavior in Mice. Evid. Based Complement. Altern. Med. 2020, 2020, 7481813. [Google Scholar] [CrossRef] [PubMed]
- Heim, C.; Newport, D.J.; Mletzko, T.; Miller, A.H.; Nemeroff, C.B. The link between childhood trauma and depression: Insights from HPA axis studies in humans. Psychoneuroendocrinology 2008, 33, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Anacker, C.; Zunszain, P.A.; Carvalho, L.A.; Pariante, C.M. The glucocorticoid receptor: Pivot of depression and of antidepressant treatment? Psychoneuroendocrinology 2011, 36, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Petit-Demouliere, B.; Chenu, F.; Bourin, M. Forced swimming test in mice: A review of antidepressant activity. Psychopharmacology 2005, 177, 245–255. [Google Scholar] [CrossRef]
- Gorman-Sandler, E.; Hollis, F. The forced swim test: Giving up on behavioral despair (Commentary on Molendijk & de Kloet, 2021). Eur. J. Neurosci. 2022, 55, 2832–2835. [Google Scholar] [CrossRef]
- Molendijk, M.L.; de Kloet, E.R. Coping with the forced swim stressor: Current state-of-the-art. Behav. Brain Res. 2019, 364, 1–10. [Google Scholar] [CrossRef]
- Grosso, C. Herbal Medicine in Depression: Traditional Medicine to Innovative Drug Delivery; Springer: Cham, Switzerland, 2016; pp. 551–552. [Google Scholar]
- Asarnow, L.D. Depression and sleep: What has the treatment research revealed and could the HPA axis be a potential mechanism? Curr. Opin. Psychol. 2020, 34, 112–116. [Google Scholar] [CrossRef]
- Origlia, N.; Bonadonna, C.; Rosellini, A.; Leznik, E.; Arancio, O.; Yan, S.S.; Domenici, L. Microglial receptor for advanced glycation end product-dependent signal pathway drives beta-amyloid-induced synaptic depression and long-term depression impairment in entorhinal cortex. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 11414–11425. [Google Scholar] [CrossRef]
- McAfoose, J.; Baune, B.T. Evidence for a cytokine model of cognitive function. Neurosci. Biobehav. Rev. 2009, 33, 355–366. [Google Scholar] [CrossRef]
- Aicardi, G.; Argilli, E.; Cappello, S.; Santi, S.; Riccio, M.; Thoenen, H.; Canossa, M. Induction of long-term potentiation and depression is reflected by corresponding changes in secretion of endogenous brain-derived neurotrophic factor. Proc. Natl. Acad. Sci. USA 2004, 101, 15788–15792. [Google Scholar] [CrossRef]
- Rossi, C.; Angelucci, A.; Costantin, L.; Braschi, C.; Mazzantini, M.; Babbini, F.; Fabbri, M.E.; Tessarollo, L.; Maffei, L.; Berardi, N.; et al. Brain-derived neurotrophic factor (BDNF) is required for the enhancement of hippocampal neurogenesis following environmental enrichment. Eur. J. Neurosci. 2006, 24, 1850–1856. [Google Scholar] [CrossRef] [PubMed]
- Autry, A.E.; Monteggia, L.M. Brain-derived neurotrophic factor and neuropsychiatric disorders. Pharmacol. Rev. 2012, 64, 238–258. [Google Scholar] [CrossRef] [PubMed]
- Shaywitz, A.J.; Greenberg, M.E. CREB: A stimulus-induced transcription factor activated by a diverse array of extracellular signals. Annu. Rev. Biochem. 1999, 68, 821–861. [Google Scholar] [CrossRef]
- Lim, J.Y.; Park, S.I.; Oh, J.H.; Kim, S.M.; Jeong, C.H.; Jun, J.A.; Lee, K.S.; Oh, W.; Lee, J.K.; Jeun, S.S. Brain-derived neurotrophic factor stimulates the neural differentiation of human umbilical cord blood-derived mesenchymal stem cells and survival of differentiated cells through MAPK/ERK and PI3K/Akt-dependent signaling pathways. J. Neurosci. Res. 2008, 86, 2168–2178. [Google Scholar] [CrossRef]
- Hoshaw, B.A.; Malberg, J.E.; Lucki, I. Central administration of IGF-I and BDNF leads to long-lasting antidepressant-like effects. Brain Res. 2005, 1037, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Advani, T.; Koek, W.; Hensler, J.G. Gender differences in the enhanced vulnerability of BDNF+/− mice to mild stress. Int. J. Neuropsychopharmacol. 2009, 12, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.-W.; Kim, J.; Myung, S.-K.; Hwang, S.-S.; Yoon, D.-H. Antidepressant use and risk of coronary heart disease: Meta-analysis of observational studies. Br. J. Clin. Pharmacol. 2014, 78, 727–737. [Google Scholar] [CrossRef]
- Eom, C.S.; Lee, H.K.; Ye, S.; Park, S.M.; Cho, K.H. Use of selective serotonin reuptake inhibitors and risk of fracture: A systematic review and meta-analysis. J. Bone Miner. Res. 2012, 27, 1186–1195. [Google Scholar] [CrossRef]
- Jia, Z.; Yang, J.; Cao, Z.; Zhao, J.; Zhang, J.; Lu, Y.; Chu, L.; Zhang, S.; Chen, Y.; Pei, L. Baicalin ameliorates chronic unpredictable mild stress-induced depression through the BDNF/ERK/CREB signaling pathway. Behav. Brain Res. 2021, 414, 113463. [Google Scholar] [CrossRef]
- Yu, Y.; Shi, Z.; Xu, D.; Li, Y.; Qin, J.; Zhang, Z.; Wang, H. Prenatal ethanol exposure increases susceptibility to depression- and anxiety-like behavior in adult female offspring and its underlying mechanism. Reprod. Toxicol. 2020, 96, 36–46. [Google Scholar] [CrossRef]
- Faria, A.; Mateus, N.; Calhau, C. Flavonoid transport across blood-brain barrier: Implication for their direct neuroprotective actions. Nutr. Aging 2012, 1, 89–97. [Google Scholar] [CrossRef]
- Sawmiller, D.; Li, S.; Shahaduzzaman, M.; Smith, A.J.; Obregon, D.; Giunta, B.; Borlongan, C.V.; Sanberg, P.R.; Tan, J. Luteolin reduces Alzheimer’s disease pathologies induced by traumatic brain injury. Int. J. Mol. Sci. 2014, 15, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Marshall, T.; Shult, P.; Busse, W.W. Release of lysosomal enzyme beta-glucuronidase from isolated human eosinophils. J. Allergy Clin. Immunol. 1988, 82, 550–555. [Google Scholar] [CrossRef]
- Shimoi, K.; Nakayama, T. Glucuronidase deconjugation in inflammation. Methods Enzymol. 2005, 400, 263–272. [Google Scholar] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ryu, D.; Jee, H.-J.; Kim, S.-Y.; Hwang, S.-H.; Pil, G.-B.; Jung, Y.-S. Luteolin-7-O-Glucuronide Improves Depression-like and Stress Coping Behaviors in Sleep Deprivation Stress Model by Activation of the BDNF Signaling. Nutrients 2022, 14, 3314. https://doi.org/10.3390/nu14163314
Ryu D, Jee H-J, Kim S-Y, Hwang S-H, Pil G-B, Jung Y-S. Luteolin-7-O-Glucuronide Improves Depression-like and Stress Coping Behaviors in Sleep Deprivation Stress Model by Activation of the BDNF Signaling. Nutrients. 2022; 14(16):3314. https://doi.org/10.3390/nu14163314
Chicago/Turabian StyleRyu, Dajung, Hye-Jin Jee, Sang-Yoon Kim, Seung-Hwan Hwang, Gam-Bang Pil, and Yi-Sook Jung. 2022. "Luteolin-7-O-Glucuronide Improves Depression-like and Stress Coping Behaviors in Sleep Deprivation Stress Model by Activation of the BDNF Signaling" Nutrients 14, no. 16: 3314. https://doi.org/10.3390/nu14163314
APA StyleRyu, D., Jee, H.-J., Kim, S.-Y., Hwang, S.-H., Pil, G.-B., & Jung, Y.-S. (2022). Luteolin-7-O-Glucuronide Improves Depression-like and Stress Coping Behaviors in Sleep Deprivation Stress Model by Activation of the BDNF Signaling. Nutrients, 14(16), 3314. https://doi.org/10.3390/nu14163314