
Fetal Brain Damage during Maternal COVID-19: Emerging Hypothesis, Mechanism, and Possible Mitigation through Maternal-Targeted Nutritional Supplementation


 ,
,  , and
, and
Abstract
:1. Introduction
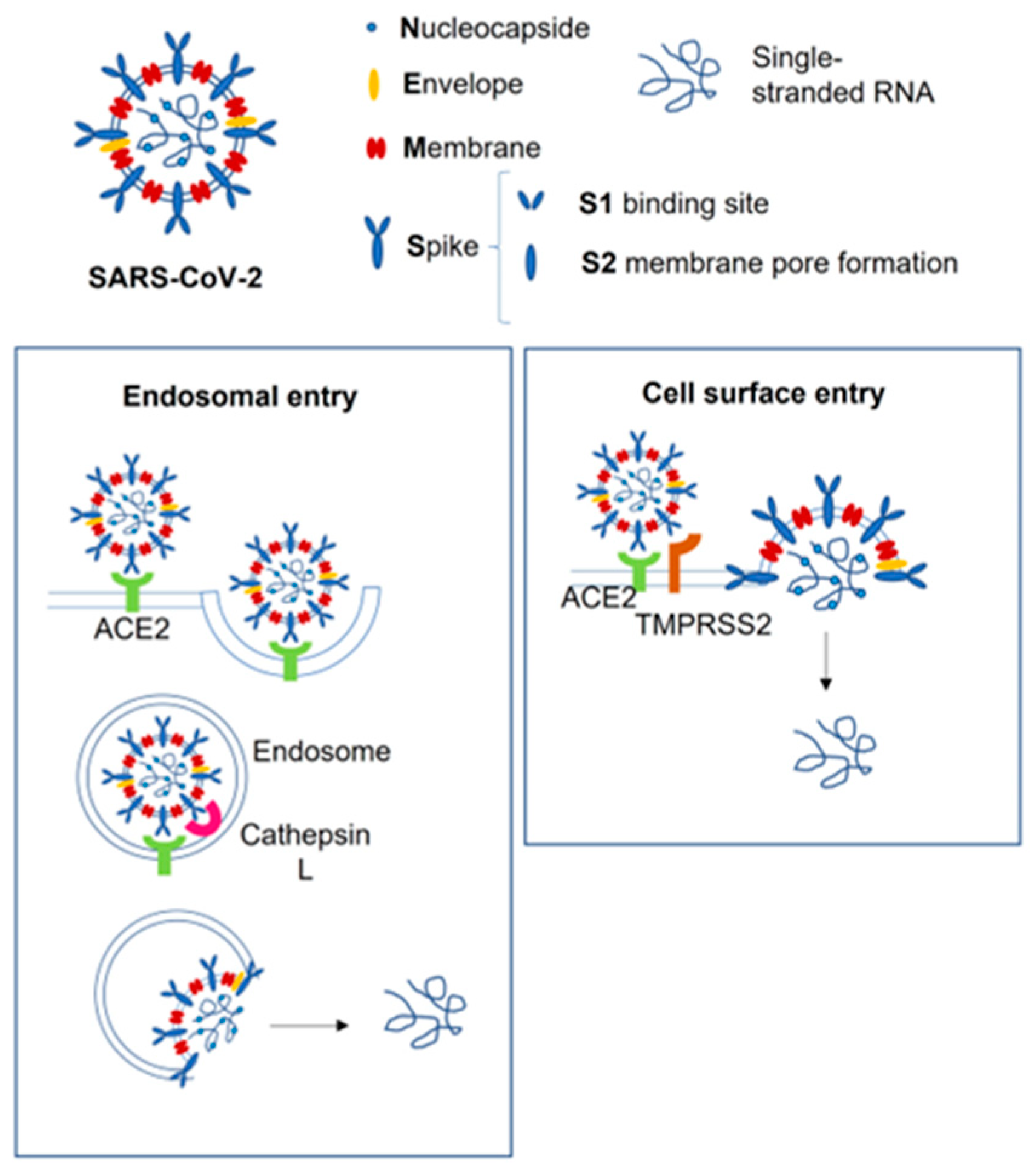
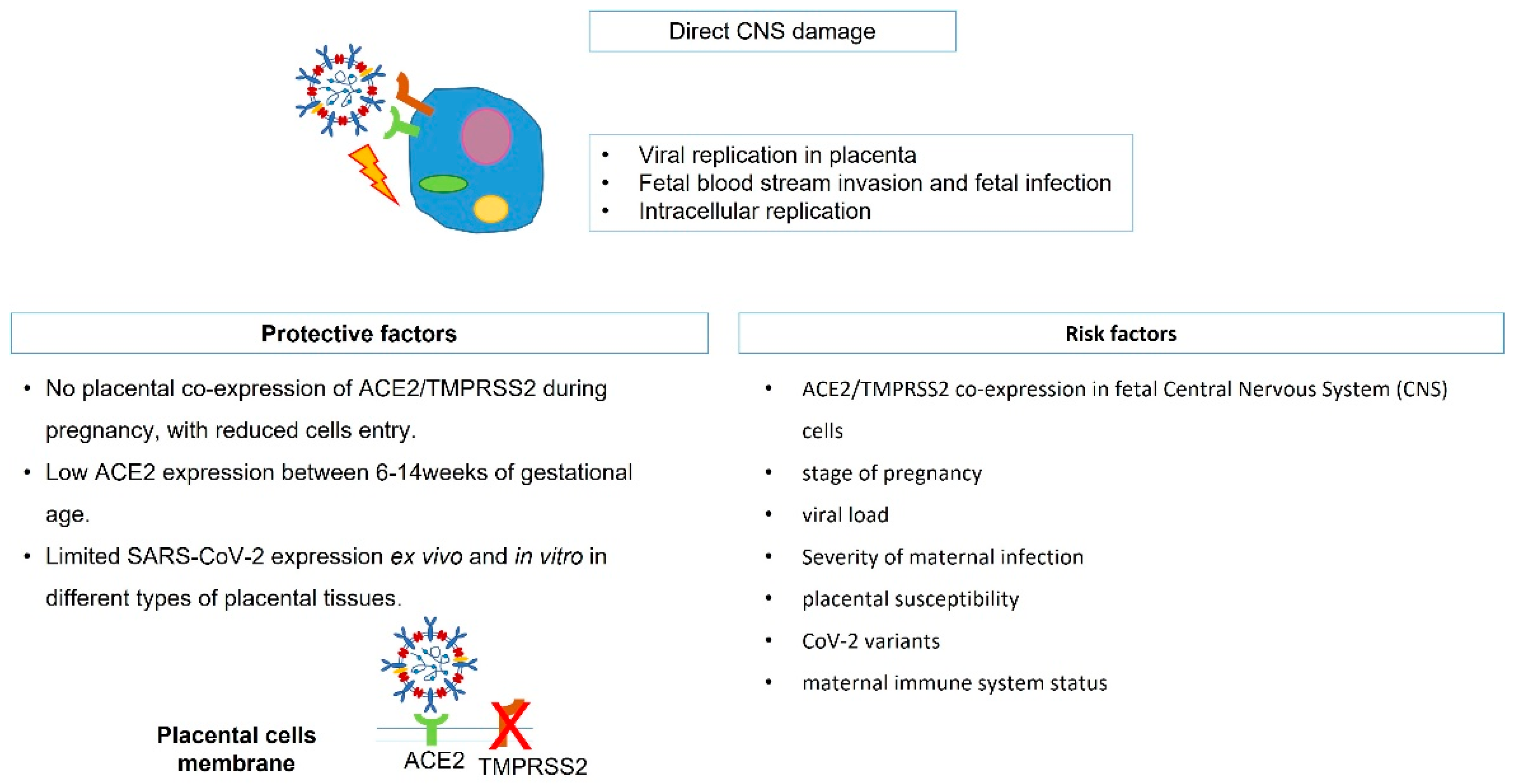
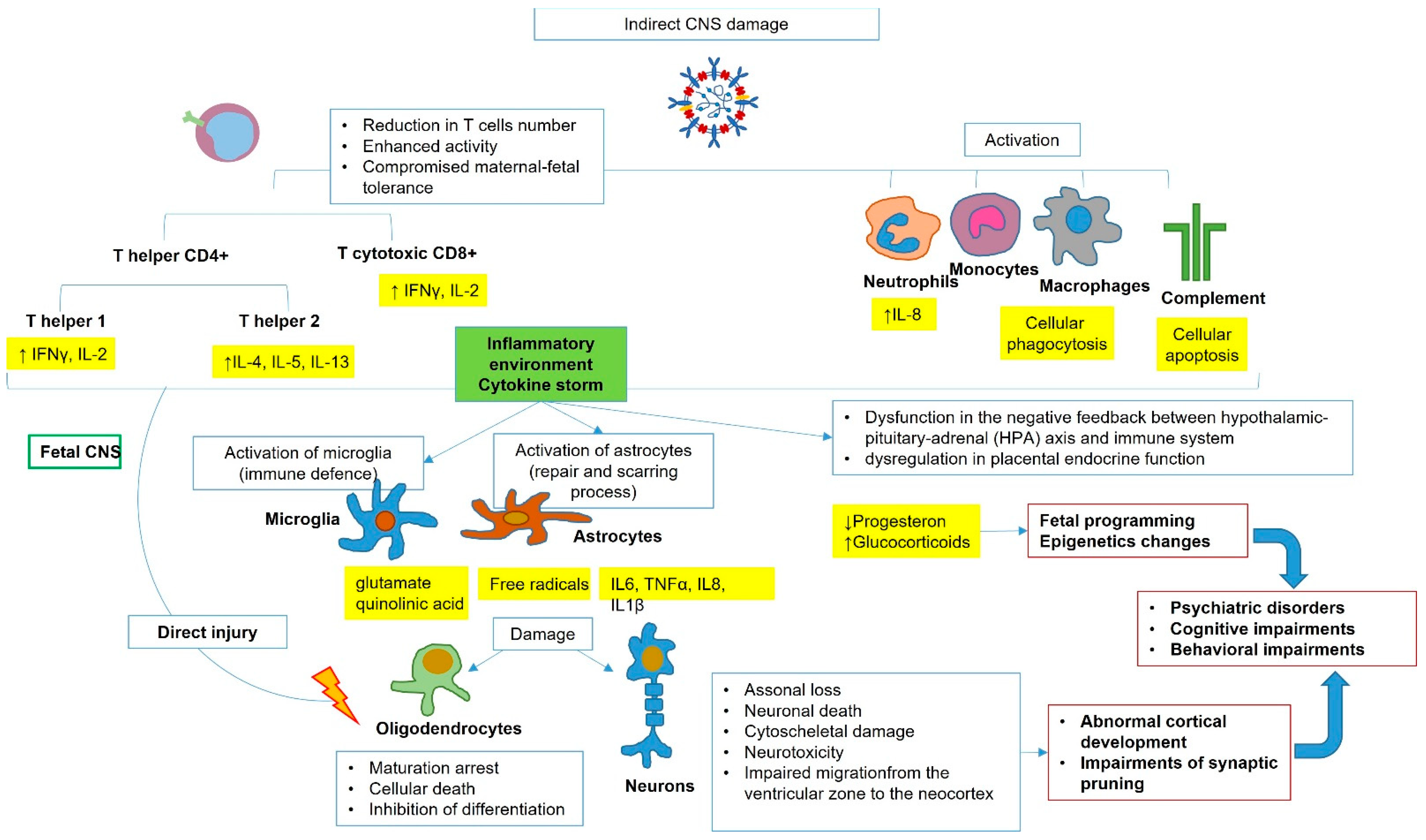
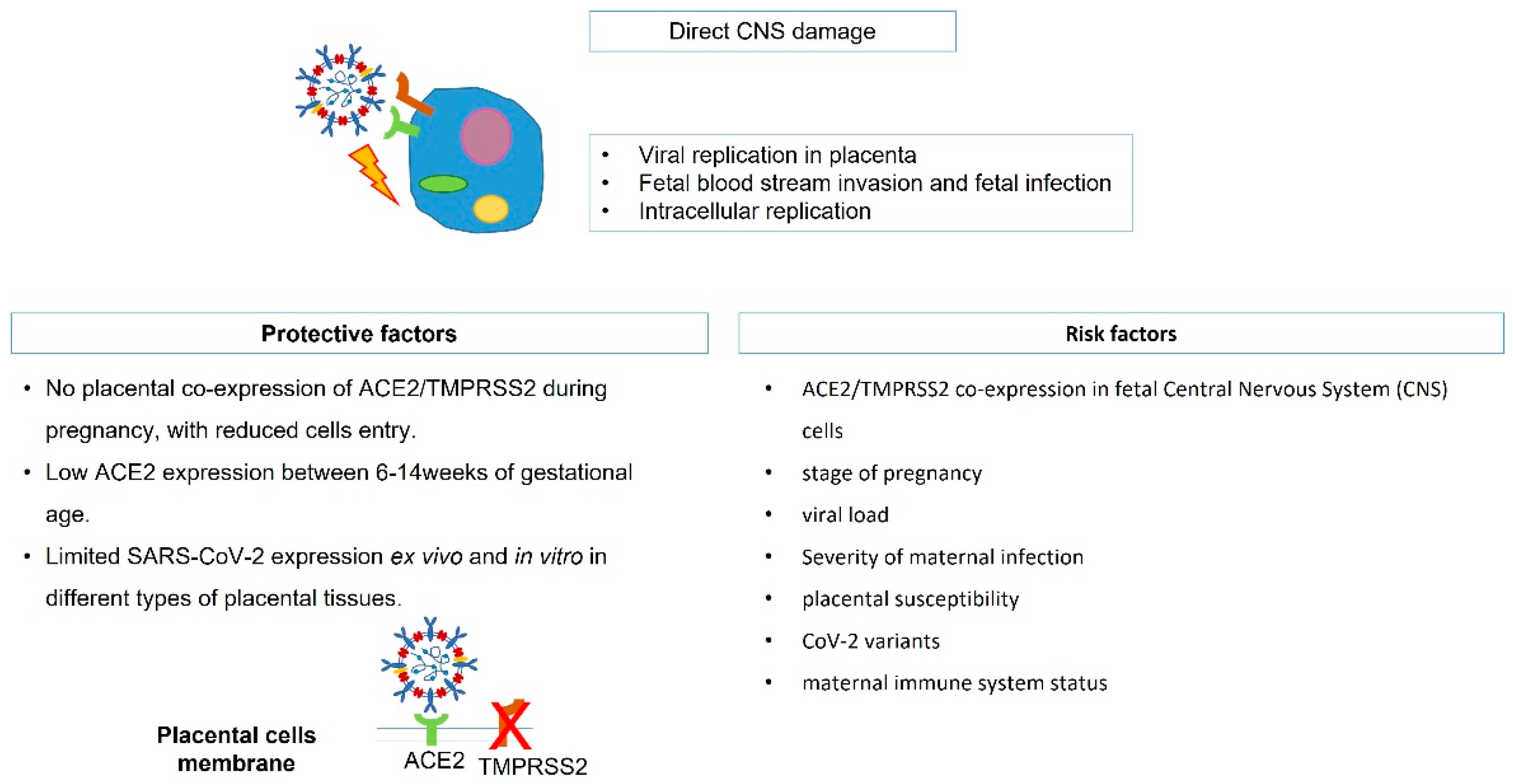
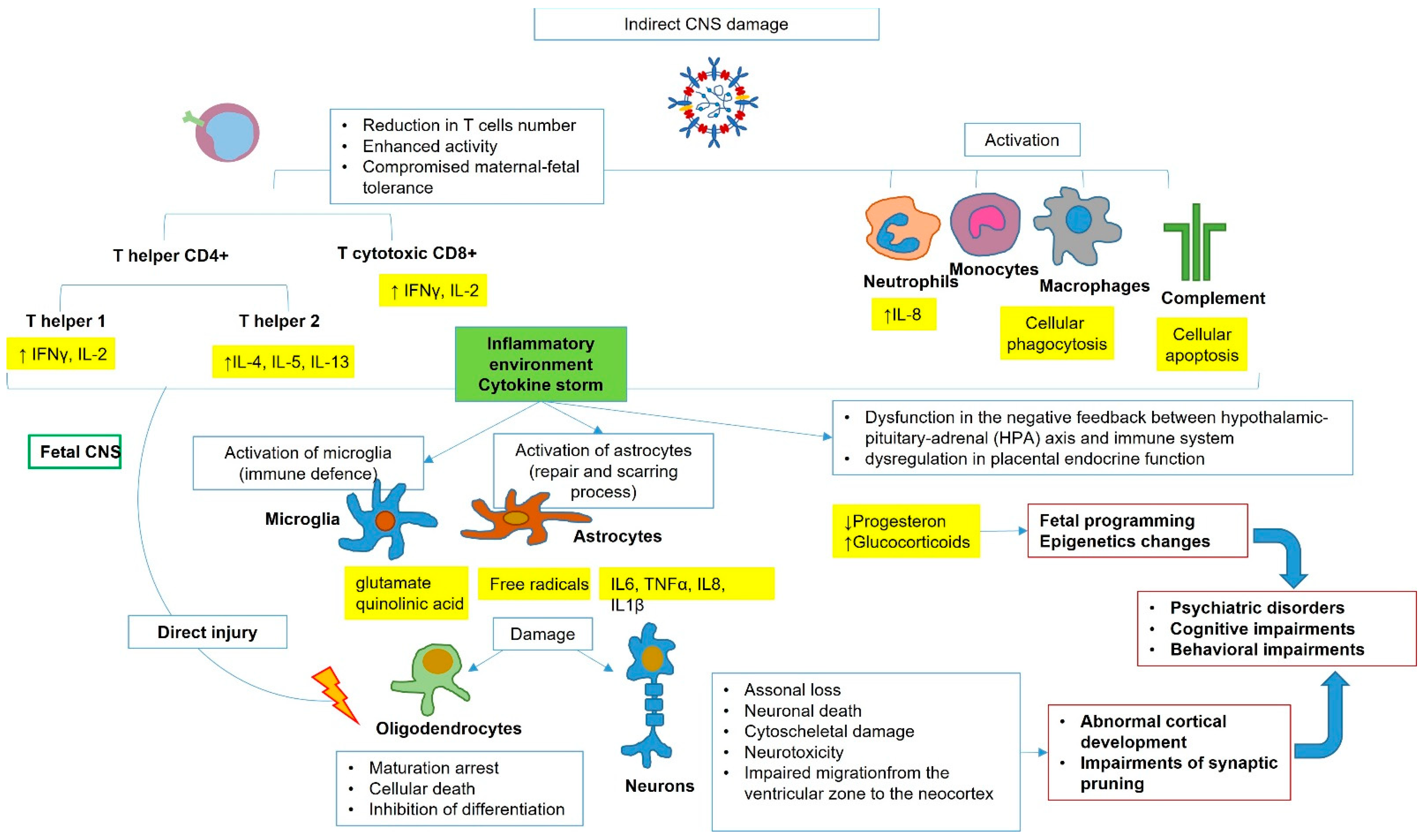
2. SARS-CoV-2 Vertical Transmission, Fetal Infection, and Potential Brain Damage
3. Strategies of Prevention
4. Nutrition and Supplementation in Preventing a Negative Fetal Outcome of SARS-CoV-2 Infection
4.1. Amino Acids
4.2. Omega-3-Poly-Unsaturated Fatty Acids
4.3. Fatty Acids (S/MCFA)
4.4. Vitamin A (All-Trans-Retinol)
4.5. Vitamin D (25(OH)D)
4.6. Vitamin E (Tocopherol)
4.7. Vitamin B1 (Thiamine)
4.8. Vitamin B6 (Pyridoxine)
4.9. Vitamin B9 (Folic Acid)
4.10. Vitamin B12 (Cobalamin)
4.11. Vitamin C (Ascorbic Acid)
4.12. Iron
4.13. Zinc
4.14. Selenium
4.15. Copper
4.16. Magnesium
4.17. Choline
4.18. Phytonutrients
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- de Vries, L.S. Viral Infections and the Neonatal Brain. Semin. Pediatr. Neurol. 2019, 32, 100769. [Google Scholar] [CrossRef]
- Kraus, T.A.; Engel, S.M.; Sperling, R.S.; Kellerman, L.; Lo, Y.; Wallenstein, S.; Escribese, M.M.; Garrido, J.L.; Singh, T.; Loubeau, M.; et al. Characterizing the pregnancy immune phenotype: Results of the viral immunity and pregnancy (VIP) study. J. Clin. Immunol. 2012, 32, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Pazos, M.; Sperling, R.S.; Moran, T.M.; Kraus, T.A. The influence of pregnancy on systemic immunity. Immunol. Res. 2012, 54, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Priddy, K.D. Immunologic adaptations during pregnancy. J. Obstet. Gynecol. Neonatal. Nurs. 1997, 26, 388–394. [Google Scholar] [CrossRef]
- Sappenfield, E.; Jamieson, D.J.; Kourtis, A.P. Pregnancy and susceptibility to infectious diseases. Infect. Dis. Obstet. Gynecol. 2013, 2013, 752852. [Google Scholar] [CrossRef] [PubMed]
- Harapan, B.N.; Yoo, H.J. Neurological symptoms, manifestations, and complications associated with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) and coronavirus disease 19 (COVID-19). J. Neurol. 2021, 268, 3059–3071. [Google Scholar] [CrossRef] [PubMed]
- Maury, A.; Lyoubi, A.; Peiffer-Smadja, N.; de Broucker, T.; Meppiel, E. Neurological manifestations associated with SARS-CoV-2 and other coronaviruses: A narrative review for clinicians. Rev. Neurol. 2021, 177, 51–64. [Google Scholar] [CrossRef]
- Román, G.C.; Spencer, P.S.; Reis, J.; Buguet, A.; Faris, M.E.A.; Katrak, S.M.; Láinez, M.; Medina, M.T.; Meshram, C.; Mizusawa, H.; et al. The neurology of COVID-19 revisited: A proposal from the Environmental Neurology Specialty Group of the World Federation of Neurology to implement international neurological registries. J. Neurol. Sci. 2020, 414, 116884. [Google Scholar] [CrossRef]
- Sharifian-Dorche, M.; Huot, P.; Osherov, M.; Wen, D.; Saveriano, A.; Giacomini, P.S.; Antel, J.P.; Mowla, A. Neurological complications of coronavirus infection; a comparative review and lessons learned during the COVID-19 pandemic. J. Neurol. Sci. 2020, 417, 117085. [Google Scholar] [CrossRef] [PubMed]
- Desforges, M.; Le Coupanec, A.; Brison, É.; Meessen-Pinard, M.; Talbot, P.J. Human respiratory coronaviruses: Neuroinvasive, neurotropic and potentially neurovirulent pathogens. Virologie 2014, 18, 5–16. (In English) [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, C.P.; Fontes-Dantas, F.L.; da Poian, A.T.; Clarke, J.R. SARS-CoV-2-associated cytokine storm during pregnancy as a possible risk factor for neuropsychiatric disorder development in post-pandemic infants. Neuropharmacology 2021, 201, 108841. [Google Scholar] [CrossRef] [PubMed]
- Leyser, M.; Marques, F.J.P.; Nascimento, O.J.M.D. Potential risk of brain damage and poor developmental outcomes in children prenatally exposed to SARS-CoV-2: A systematic review. Rev. Paul. Pediatr. 2021, 40, e2020415. [Google Scholar] [CrossRef] [PubMed]
- Juan, J.; Gil, M.M.; Rong, Z.; Zhang, Y.; Yang, H.; Poon, L.C. Effect of coronavirus disease 2019 (COVID-19) on maternal, perinatal and neonatal outcome: Systematic review. Ultrasound Obstet. Gynecol. 2020, 56, 15–27. [Google Scholar] [CrossRef]
- Curcio, A.M.; Shekhawat, P.; Reynolds, A.S.; Thakur, K.T. Neurologic infections during pregnancy. Handb. Clin. Neurol. 2020, 172, 79–104. [Google Scholar] [CrossRef]
- Mate, A.; Reyes-Goya, C.; Santana-Garrido, Á.; Vázquez, C.M. Lifestyle, Maternal Nutrition and Healthy Pregnancy. Curr. Vasc. Pharmacol. 2021, 19, 132–140. [Google Scholar] [CrossRef]
- Katona, P.; Katona-Apte, J. The interaction between nutrition and infection. Clin. Infect. Dis. 2008, 46, 1582–1588. [Google Scholar] [CrossRef]
- Jackson, C.B.; Farzan, M.; Chen, B.; Choe, H. Mechanisms of SARS-CoV-2 entry into cells. Nat. Rev. Mol. Cell Biol. 2022, 23, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Beeraka, N.M.; Sadhu, S.P.; Madhunapantula, S.V.; Rao Pragada, R.; Svistunov, A.A.; Nikolenko, V.N.; Mikhaleva, L.M.; Aliev, G. Strategies for Targeting SARS CoV-2: Small Molecule Inhibitors—The Current Status. Front. Immunol. 2020, 11, 552925. [Google Scholar] [CrossRef]
- Patberg, E.T.; Adams, T.; Rekawek, P.; Vahanian, S.A.; Akerman, M.; Hernandez, A.; Rapkiewicz, A.V.; Ragolia, L.; Sicuranza, G.; Chavez, M.R.; et al. Coronavirus disease 2019 infection and placental histopathology in women delivering at term. Am. J. Obstet. Gynecol. 2021, 224, 382.e1–382.e18. [Google Scholar] [CrossRef] [PubMed]
- Tallarek, A.C.; Urbschat, C.; Fonseca Brito, L.; Stanelle-Bertram, S.; Krasemann, S.; Frascaroli, G.; Thiele, K.; Wieczorek, A.; Felber, N.; Lütgehetmann, M.; et al. Inefficient Placental Virus Replication and Absence of Neonatal Cell-Specific Immunity Upon SARS-CoV-2 Infection During Pregnancy. Front. Immunol. 2021, 12, 698578. [Google Scholar] [CrossRef] [PubMed]
- Favre, G.; Mazzetti, S.; Gengler, C.; Bertelli, C.; Schneider, J.; Laubscher, B.; Capoccia, R.; Pakniyat, F.; Ben Jazia, I.; Eggel-Hort, B.; et al. Decreased Fetal Movements: A Sign of Placental SARS-CoV-2 Infection with Perinatal Brain Injury. Viruses 2021, 13, 2517. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, D.A. An analysis of 38 pregnant women with COVID-19, their newborn infants, and maternal-fetal transmission of SARS-CoV-2: Maternal coronavirus infections and pregnancy outcomes. Arch Pathol. Lab. Med. 2020, 144, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Zhang, J.; Xiong, C.; Li, X. The SARS-CoV-2 receptor ACE2 expression of maternal-fetal interface and fetal organs by single-cell transcriptome study. PLoS ONE 2020, 15, e0230295. [Google Scholar] [CrossRef] [PubMed]
- Pique-Regi, R.; Romero, R.; Tarca, A.L.; Luca, F.; Xu, Y.; Alazizi, A.; Leng, Y.; Hsu, C.D.; Gomez-Lopez, N. Does the human placenta express the canonical cell entry mediators for SARS-CoV-2? Elife 2020, 9, e58716. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.F.; Wong, S.F.; Lam, A.; Mak, Y.F.; Yao, H.; Lee, K.C.; Chow, K.M.; Yu, W.C.; Ho, L.C. The placentas of patients with severe acute respiratory syndrome: A pathophysiological evaluation. Pathology 2006, 38, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Hosier, H.; Farhadian, S.F.; Morotti, R.A.; Deshmukh, U.; Lu-Culligan, A.; Campbell, K.H.; Yasumoto, Y.; Vogels, C.B.; Casanovas-Massana, A.; Vijayakumar, P.; et al. SARS-CoV-2 infection of the placenta. J. Clin. Investig. 2020, 130, 4947–4953. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Sang, L.; Du, S.; Li, T.; Chang, Y.; Yang, X.A. Asymptomatic COVID-19 infection in late pregnancy indicated no vertical transmission. J. Med. Virol. 2020, 92, 1660–1664. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Flores, V.; Romero, R.; Xu, Y.; Theis, K.R.; Arenas-Hernandez, M.; Miller, D.; Peyvandipour, A.; Bhatti, G.; Galaz, J.; Gershater, M.; et al. Maternal-fetal immune responses in pregnant women infected with SARS-CoV-2. Nat. Commun. 2022, 13, 320. [Google Scholar] [CrossRef]
- Magatti, M.; Stefani, F.R.; Papait, A.; Cargnoni, A.; Masserdotti, A.; Silini, A.R.; Parolini, O. Perinatal Mesenchymal Stromal Cells and Their Possible Contribution to Fetal-Maternal Tolerance. Cells 2019, 8, 1401. [Google Scholar] [CrossRef]
- Bartha, A.I.; Foster-Barber, A.; Miller, S.P.; Vigneron, D.B.; Glidden, D.V.; Barkovich, A.J.; Ferriero, D.M. Neonatal encephalopathy: Association of cytokines with MR spectroscopy and outcome. Pediatr. Res. 2004, 56, 960–966. [Google Scholar] [CrossRef]
- Perico, L.; Benigni, A.; Casiraghi, F.; Ng, L.F.P.; Renia, L.; Remuzzi, G. Immunity, endothelial injury and complement-induced coagulopathy in COVID-19. Nat. Rev. Nephrol. 2021, 17, 46–64. [Google Scholar] [CrossRef]
- Bell, M.J.; Hallenbeck, J.M. Effects of intrauterine inflammation on developing rat brain. J. Neurosci. Res. 2002, 70, 570–579. [Google Scholar] [CrossRef]
- Meyer, U.; Feldon, J.; Yee, B.K. A review of the fetal brain cytokine imbalance hypothesis of schizophrenia. Schizophr. Bull. 2009, 35, 959–972. [Google Scholar] [CrossRef]
- Ohyu, J.; Marumo, G.; Ozawa, H.; Takashima, S.; Nakajima, K.; Kohsaka, S.; Hamai, Y.; Machida, Y.; Kobayashi, K.; Ryo, E.; et al. Early axonal and glial pathology in fetal sheep brains with leukomalacia induced by repeated umbilical cord occlusion. Brain Dev. 1999, 21, 248–252. [Google Scholar] [CrossRef]
- Gomez, R.; Romero, R.; Ghezzi, F.; Yoon, B.H.; Mazor, M.; Berry, S.M. The fetal inflammatory response syndrome. Am. J. Obstet. Gynecol. 1998, 179, 194–202. [Google Scholar] [CrossRef]
- Ramiro-Cortijo, D.; de la Calle, M.; Böger, R.; Hannemann, J.; Lüneburg, N.; López-Giménez, M.R.; Rodríguez-Rodríguez, P.; Martín-Cabrejas, M.Á.; Benítez, V.; de Pablo, Á.L.L.; et al. Male fetal sex is associated with low maternal plasma anti-inflammatory cytokine profile in the first trimester of healthy pregnancies. Cytokine 2020, 136, 155290. [Google Scholar] [CrossRef]
- Burd, I.; Balakrishnan, B.; Kannan, S. Models of fetal brain injury, intrauterine inflammation, and preterm birth. Am. J. Reprod. Immunol. 2012, 67, 287–294. [Google Scholar] [CrossRef]
- Raony, Í.; de Figueiredo, C.S.; Pandolfo, P.; Giestal-de-Araujo, E.; Oliveira-Silva Bomfim, P.; Savino, W. Psycho-Neuroendocrine-Immune Interactions in COVID-19: Potential Impacts on Mental Health. Front. Immunol. 2020, 11, 1170. [Google Scholar] [CrossRef] [PubMed]
- Littauer, E.Q.; Esser, E.S.; Antao, O.Q.; Vassilieva, E.V.; Compans, R.W.; Skountzou, I. H1N1 influenza virus infection results in adverse pregnancy outcomes by disrupting tissue-specific hormonal regulation. PLoS Pathog. 2017, 13, e1006757. [Google Scholar] [CrossRef]
- Seckl, J.R.; Holmes, M.C. Mechanisms of disease: Glucocorticoids, their placental metabolism and fetal ‘programming’ of adult pathophysiology. Nat. Clin. Pract. Endocrinol. Metab. 2007, 3, 479–488. [Google Scholar] [CrossRef]
- Atladóttir, H.O.; Thorsen, P.; Østergaard, L.; Schendel, D.E.; Lemcke, S.; Abdallah, M.; Parner, E.T. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J. Autism Dev. Disord. 2010, 40, 1423–1430. [Google Scholar] [CrossRef]
- Al-Haddad, B.J.S.; Oler, E.; Armistead, B.; Elsayed, N.A.; Weinberger, D.R.; Bernier, R.; Burd, I.; Kapur, R.; Jacobsson, B.; Wang, C.; et al. The fetal origins of mental illness. Am. J. Obstet. Gynecol. 2019, 221, 549–562. [Google Scholar] [CrossRef]
- Liao, X.; Liu, Y.; Fu, X.; Li, Y. Postmortem Studies of Neuroinflammation in Autism Spectrum Disorder: A Systematic Review. Mol. Neurobiol. 2020, 57, 3424–3438. [Google Scholar] [CrossRef] [PubMed]
- Meyer, U.; Feldon, J.; Schedlowski, M.; Yee, B.K. Immunological stress at the maternal-foetal interface: A link between neurodevelopment and adult psychopathology. Brain Behav. Immun. 2006, 20, 378–388. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Kalafat, E.; Blakeway, H.; Townsend, R.; O’Brien, P.; Morris, E.; Draycott, T.; Thangaratinam, S.; Le Doare, K.; Ladhani, S.; et al. Systematic review and meta-analysis of the effectiveness and perinatal outcomes of COVID-19 vaccination in pregnancy. Nat. Commun. 2022, 13, 2414. [Google Scholar] [CrossRef] [PubMed]
- Honein, M.A.; Christie, A.; Rose, D.A.; Brooks, J.T.; Meaney-Delman, D.; Cohn, A.; Sauber-Schatz, E.K.; Walker, A.; McDonald, L.C.; Liburd, L.C.; et al. Summary of Guidance for Public Health Strategies to Address High Levels of Community Transmission of SARS-CoV-2 and Related Deaths, December 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 1860–1867. [Google Scholar] [CrossRef] [PubMed]
- Davenport, M.H.; Marchand, A.A.; Mottola, M.F.; Poitras, V.J.; Gray, C.E.; Jaramillo Garcia, A.; Barrowman, N.; Sobierajski, F.; James, M.; Meah, V.L.; et al. Exercise for the prevention and treatment of low back, pelvic girdle and lumbopelvic pain during pregnancy: A systematic review and meta-analysis. Br. J. Sports Med. 2019, 53, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Bulgakova, V.A.; Poromov, A.A.; Grekova, A.I.; Pshenichnaya, N.Y.; Selkova, E.P.; Lvov, N.I.; Leneva, I.A.; Shestakova, I.V.; Maleev, V.V. Pharmacoepidemiological study of the course of influenza and other acute respiratory viral infections in risk groups. Ter. Arkhiv 2017, 89, 62–71. (In Russian) [Google Scholar] [CrossRef]
- Fedele, D.; De Francesco, A.; Riso, S.; Collo, A. Obesity, malnutrition, and trace element deficiency in the coronavirus disease (COVID-19) pandemic: An overview. Nutrition 2021, 81, 111016. [Google Scholar] [CrossRef]
- Mousa, A.; Naqash, A.; Lim, S. Macronutrient and Micronutrient Intake during Pregnancy: An Overview of Recent Evidence. Nutrients 2019, 11, 443. [Google Scholar] [CrossRef]
- Venter, C.; Eyerich, S.; Sarin, T.; Klatt, K.C. Nutrition and the Immune System: A Complicated Tango. Nutrients 2020, 12, 818. [Google Scholar] [CrossRef] [PubMed]
- Maggini, S.; Wintergerst, E.S.; Beveridge, S.; Hornig, D.H. Selected vitamins and trace elements support immune function by strengthening epithelial barriers and cellular and humoral immune responses. Br. J. Nutr. 2007, 98 (Suppl. 1), S29–S35. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yin, Y.L.; Li, D.; Kim, S.W.; Wu, G. Amino acids and immune function. Br. J. Nutr. 2007, 98, 237–252. [Google Scholar] [CrossRef]
- Akerström, S.; Mousavi-Jazi, M.; Klingström, J.; Leijon, M.; Lundkvist, A.; Mirazimi, A. Nitric oxide inhibits the replication cycle of severe acute respiratory syndrome coronavirus. J. Virol. 2005, 79, 1966–1969. [Google Scholar] [CrossRef]
- Paakkari, I.; Lindsberg, P. Nitric oxide in the central nervous system. Ann. Med. 1995, 27, 369–377. [Google Scholar] [CrossRef]
- Souci, S.W.; Fachmann, W.; Kraut, H. Food Composition and Nutrition Tables, 5th ed.; Medpharm Scientific Publishers: Stuttgart, Germany, 1994. [Google Scholar]
- Visek, W.J. Arginine needs, physiological state and usual diets. A reevaluation. J. Nutr. 1986, 116, 36–46. [Google Scholar] [CrossRef]
- Alexander, J.W.; Supp, D.M. Role of Arginine and Omega-3 Fatty Acids in Wound Healing and Infection. Adv. Wound Care 2014, 3, 682–690. [Google Scholar] [CrossRef]
- Li, X.; Bi, X.; Wang, S.; Zhang, Z.; Li, F.; Zhao, A.Z. Therapeutic Potential of ω-3 Polyunsaturated Fatty Acids in Human Autoimmune Diseases. Front. Immunol. 2019, 10, 2241. [Google Scholar] [CrossRef]
- Mildenberger, J.; Johansson, I.; Sergin, I.; Kjøbli, E.; Damås, J.K.; Razani, B.; Flo, T.H.; Bjørkøy, G. N-3 PUFAs induce inflammatory tolerance by formation of KEAP1-containing SQSTM1/p62-bodies and activation of NFE2L2. Autophagy 2017, 13, 1664–1678. [Google Scholar] [CrossRef]
- Miles, E.A.; Calder, P.C. Influence of marine n-3 polyunsaturated fatty acids on immune function and a systematic review of their effects on clinical outcomes in rheumatoid arthritis. Br. J. Nutr. 2012, 107 (Suppl. 2), S171–S184. [Google Scholar] [CrossRef]
- Chang, Y.F.; Hou, Y.C.; Pai, M.H.; Yeh, S.L.; Liu, J.J. Effects of ω-3 Polyunsaturated Fatty Acids on the Homeostasis of CD4+ T Cells and Lung Injury in Mice With Polymicrobial Sepsis. JPEN J. Parenter. Enteral. Nutr. 2017, 41, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Messina, G.; Polito, R.; Monda, V.; Cipolloni, L.; Di Nunno, N.; Di Mizio, G.; Murabito, P.; Carotenuto, M.; Messina, A.; Pisanelli, D.; et al. Functional Role of Dietary Intervention to Improve the Outcome of COVID-19: A Hypothesis of Work. Int. J. Mol. Sci. 2020, 21, 3104. [Google Scholar] [CrossRef]
- Madore, C.; Leyrolle, Q.; Lacabanne, C.; Benmamar-Badel, A.; Joffre, C.; Nadjar, A.; Layé, S. Neuroinflammation in Autism: Plausible Role of Maternal Inflammation, Dietary Omega 3, and Microbiota. Neural. Plast. 2016, 2016, 3597209. [Google Scholar] [CrossRef]
- Ye, Q.; Cai, S.; Wang, S.; Zeng, X.; Ye, C.; Chen, M.; Zeng, X.; Qiao, S. Maternal short and medium chain fatty acids supply during early pregnancy improves embryo survival through enhancing progesterone synthesis in rats. J. Nutr. Biochem. 2019, 69, 98–107. [Google Scholar] [CrossRef]
- Chesworth, R.; Gamage, R.; Ullah, F.; Sonego, S.; Millington, C.; Fernandez, A.; Liang, H.; Karl, T.; Münch, G.; Niedermayer, G.; et al. Spatial Memory and Microglia Activation in a Mouse Model of Chronic Neuroinflammation and the Anti-inflammatory Effects of Apigenin. Front. Neurosci. 2021, 15, 699329. [Google Scholar] [CrossRef]
- Projan, S.J.; Brown-Skrobot, S.; Schlievert, P.M.; Vandenesch, F.; Novick, R.P. Glycerol monolaurate inhibits the production of beta-lactamase, toxic shock toxin-1, and other staphylococcal exoproteins by interfering with signal transduction. J. Bacteriol. 1994, 176, 4204–4209. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, Y. Potential interventions for novel coronavirus in China: A systematic review. J. Med. Virol. 2020, 92, 479–490. [Google Scholar] [CrossRef]
- Gombart, A.F.; Pierre, A.; Maggini, S. A Review of Micronutrients and the Immune System-Working in Harmony to Reduce the Risk of Infection. Nutrients 2020, 12, 236. [Google Scholar] [CrossRef]
- Bastos Maia, S.; Rolland Souza, A.S.; Costa Caminha, M.F.; Lins da Silva, S.; Callou Cruz, R.S.B.L.; Carvalho Dos Santos, C.; Batista Filho, M. Vitamin A and Pregnancy: A Narrative Review. Nutrients 2019, 11, 681. [Google Scholar] [CrossRef]
- Ginde, A.A.; Sullivan, A.F.; Mansbach, J.M.; Camargo, C.A., Jr. Vitamin D insufficiency in pregnant and nonpregnant women of childbearing age in the United States. Am. J. Obstet. Gynecol. 2010, 202, 436.e1–436.e8. [Google Scholar] [CrossRef]
- Holick, M.F. The vitamin D deficiency pandemic: Approaches for diagnosis, treatment and prevention. Rev. Endocr. Metab. Disord. 2017, 18, 153–165. [Google Scholar] [CrossRef] [PubMed]
- BourBour, F.; Mirzaei Dahka, S.; Gholamalizadeh, M.; Akbari, M.E.; Shadnoush, M.; Haghighi, M.; Taghvaye-Masoumi, H.; Ashoori, N.; Doaei, S. Nutrients in prevention, treatment, and management of viral infections; special focus on Coronavirus. Arch Physiol. Biochem. 2020, 9, 1–10. [Google Scholar] [CrossRef]
- Hastie, C.E.; Mackay, D.F.; Ho, F.; Celis-Morales, C.A.; Katikireddi, S.V.; Niedzwiedz, C.L.; Jani, B.D.; Welsh, P.; Mair, F.S.; Gray, S.R.; et al. Vitamin D concentrations and COVID-19 infection in UK Biobank. Diabetes Metab. Syndr. 2020, 14, 561–565. [Google Scholar] [CrossRef]
- Bokharee, N.; Khan, Y.H.; Wasim, T.; Mallhi, T.H.; Alotaibi, N.H.; Iqbal, M.S.; Rehman, K.; Alzarea, A.I.; Khokhar, A. Daily versus stat vitamin D supplementation during pregnancy; A prospective cohort study. PLoS ONE 2020, 15, e0231590. [Google Scholar] [CrossRef]
- Calder, P.C.; Carr, A.C.; Gombart, A.F.; Eggersdorfer, M. Optimal Nutritional Status for a Well-Functioning Immune System Is an Important Factor to Protect against Viral Infections. Nutrients 2020, 12, 1181. [Google Scholar] [CrossRef]
- Galabov, A.S.; Mileva, M.; Simeonova, L.; Gegova, G. Combination activity of neuraminidase inhibitor oseltamivir and α-tocopherol in influenza virus A (H3N2) infection in mice. Antivir. Chem. Chemother. 2015, 24, 83–91. [Google Scholar] [CrossRef]
- Spinas, E.; Saggini, A.; Kritas, S.K.; Cerulli, G.; Caraffa, A.; Antinolfi, P.; Pantalone, A.; Frydas, A.; Tei, M.; Speziali, A.; et al. Crosstalk between Vitamin B and immunity. J. Biol. Regul. Homeost. Agents 2015, 29, 283–288. [Google Scholar]
- Kloss, O.; Eskin, N.A.M.; Suh, M. Thiamin deficiency on fetal brain development with and without prenatal alcohol exposure. Biochem. Cell Biol. 2018, 96, 169–177. [Google Scholar] [CrossRef]
- Stolakis, V.; Liapi, C.; Al-Humadi, H.; Kalafatakis, K.; Gkanti, V.; Bimpis, A.; Skandali, N.; Tsela, S.; Theocharis, S.; Zarros, A.; et al. Effects of gestational thiamine-deprivation and/or exposure to ethanol on crucial offspring rat brain enzyme activities. J. Matern. Fetal. Neonatal. Med. 2021, 34, 2458–2466. [Google Scholar] [CrossRef]
- Fernández-Villa, D.; Aguilar, M.R.; Rojo, L. Folic Acid Antagonists: Antimicrobial and Immunomodulating Mechanisms and Applications. Int. J. Mol. Sci. 2019, 20, 4996. [Google Scholar] [CrossRef]
- Burdge, G.C.; Lillycrop, K.A. Folic acid supplementation in pregnancy: Are there devils in the detail? Br. J. Nutr. 2012, 108, 1924–1930. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qin, L.; Gao, R.; Jin, X.; Cheng, K.; Zhang, S.; Hu, X.; Xu, W.; Wang, H. Neurodevelopmental effects of maternal folic acid supplementation: A systematic review and meta-analysis. Crit. Rev. Food Sci. Nutr. 2021, 21, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Mate, A.; Reyes-Goya, C.; Santana-Garrido, Á.; Sobrevia, L.; Vázquez, C.M. Impact of maternal nutrition in viral infections during pregnancy. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166231. [Google Scholar] [CrossRef] [PubMed]
- American College of Obstetricians and Gynecologists Committee on Practice Bulletins. Neural tube defects. Number 187. Obstet. Gynecol. 2017, 130, e279–e290. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.D.; Audibert, F.; Brock, J.A.; Carroll, J.; Cartier, L.; Gagnon, A.; Johnson, J.A.; Langlois, S.; Murphy-Kaulbeck, L.; Okun, N.; et al. Pre-conception Folic Acid and Multivitamin Supplementation for the Primary and Secondary Prevention of Neural Tube Defects and Other Folic Acid-Sensitive Congenital Anomalies. J. Obstet. Gynaecol. Can. 2015, 37, 534–552. [Google Scholar] [CrossRef]
- Batista, K.S.; Cintra, V.M.; Lucena, P.A.F.; Manhães-de-Castro, R.; Toscano, A.E.; Costa, L.P.; Queiroz, M.E.B.S.; de Andrade, S.M.; Guzman-Quevedo, O.; Aquino, J.S. The role of vitamin B12 in viral infections: A comprehensive review of its relationship with the muscle-gut-brain axis and implications for SARS-CoV-2 infection. Nutr. Rev. 2022, 80, 561–578. [Google Scholar] [CrossRef]
- Manzanares, W.; Hardy, G. Vitamin B12: The forgotten micronutrient for critical care. Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 662–668. [Google Scholar] [CrossRef]
- Yalcin Bahat, P.; Aldikactioglu Talmac, M.; Bestel, A.; Topbas Selcuki, N.F.; Aydın, Z.; Polat, İ. Micronutrients in COVID-19 Positive Pregnancies. Cureus 2020, 12, e10609. [Google Scholar] [CrossRef]
- Carr, A.C.; Maggini, S. Vitamin C and Immune Function. Nutrients 2017, 9, 1211. [Google Scholar] [CrossRef]
- Colacci, M.; Gold, W.L.; Shah, R. Modern-day scurvy. CMAJ 2020, 192, E96. [Google Scholar] [CrossRef]
- Carr, A.C.; Spencer, E.; Dixon, L.; Chambers, S.T. Patients with Community Acquired Pneumonia Exhibit Depleted Vitamin C Status and Elevated Oxidative Stress. Nutrients 2020, 12, 1318. [Google Scholar] [CrossRef] [PubMed]
- Casanueva, E.; Ripoll, C.; Tolentino, M.; Morales, R.M.; Pfeffer, F.; Vilchis, P.; Vadillo-Ortega, F. Vitamin C supplementation to prevent premature rupture of the chorioamniotic membranes: A randomized trial. Am. J. Clin. Nutr. 2005, 81, 859–863. [Google Scholar] [CrossRef] [PubMed]
- Ao, G.; Li, J.; Yuan, Y.; Wang, Y.; Nasr, B.; Bao, M.; Gao, M.; Qi, X. Intravenous vitamin C use and risk of severity and mortality in COVID-19: A systematic review and meta-analysis. Nutr. Clin. Pract. 2022, 37, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Rawat, D.; Roy, A.; Maitra, S.; Gulati, A.; Khanna, P.; Baidya, D.K. Vitamin C and COVID-19 treatment: A systematic review and meta-analysis of randomized controlled trials. Diabetes Metab. Syndr. 2021, 15, 102324. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Wang, L.; Tan, J.; Liu, H.; Ni, Y. High-dose vitamin C intravenous infusion in the treatment of patients with COVID-19: A protocol for systematic review and meta-analysis. Medicine 2021, 100, e25876. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.G.; Choo, Y.J.; Chang, M.C. The effectiveness of high-dose intravenous vitamin C for patients with coronavirus disease 2019: A systematic review and meta-analysis. Complement. Ther. Med. 2022, 64, 102797. [Google Scholar] [CrossRef]
- Cheng, R.Z. Can early and high intravenous dose of vitamin C prevent and treat coronavirus disease 2019 (COVID-19)? Med. Drug Discov. 2020, 5, 100028. [Google Scholar] [CrossRef]
- Liu, W.; Zhang, S.; Nekhai, S.; Liu, S. Depriving Iron Supply to the Virus Represents a Promising Adjuvant Therapeutic Against Viral Survival. Curr. Clin. Microbiol. Rep. 2020, 7, 13–19. [Google Scholar] [CrossRef]
- Augustine, L.F.; Mullapudi, V.; Subramanian, S.; Kulkarni, B. Infection-iron interaction during COVID-19 pandemic: Time to re-design iron supplementation programs. Med. Hypotheses 2020, 143, 110173. [Google Scholar] [CrossRef]
- Saxena, D.; Spino, M.; Tricta, F.; Connelly, J.; Cracchiolo, B.M.; Hanauske, A.R.; D’Alliessi Gandolfi, D.; Mathews, M.B.; Karn, J.; Holland, B.; et al. Drug-Based Lead Discovery: The Novel Ablative Antiretroviral Profile of Deferiprone in HIV-1-Infected Cells and in HIV-Infected Treatment-Naive Subjects of a Double-Blind, Placebo-Controlled, Randomized Exploratory Trial. PLoS ONE 2016, 11, e0154842. [Google Scholar] [CrossRef] [PubMed]
- Mocchegiani, E.; Romeo, J.; Malavolta, M.; Costarelli, L.; Giacconi, R.; Diaz, L.E.; Marcos, A. Zinc: Dietary intake and impact of supplementation on immune function in elderly. Age 2013, 35, 839–860. [Google Scholar] [CrossRef] [PubMed]
- Gammoh, N.Z.; Rink, L. Zinc in Infection and Inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn(2+) inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef] [PubMed]
- Anuk, A.T.; Polat, N.; Akdas, S.; Erol, S.A.; Tanacan, A.; Biriken, D.; Keskin, H.L.; Moraloglu Tekin, O.; Yazihan, N.; Sahin, D. The Relation between Trace Element Status (Zinc, Copper, Magnesium) and Clinical Outcomes in COVID-19 Infection during Pregnancy. Biol. Trace Elem. Res. 2021, 199, 3608–3617. [Google Scholar] [CrossRef]
- Jothimani, D.; Kailasam, E.; Danielraj, S.; Nallathambi, B.; Ramachandran, H.; Sekar, P.; Manoharan, S.; Ramani, V.; Narasimhan, G.; Kaliamoorthy, I.; et al. COVID-19: Poor outcomes in patients with zinc deficiency. Int. J. Infect. Dis. 2020, 100, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Read, S.A.; Obeid, S.; Ahlenstiel, C.; Ahlenstiel, G. The Role of Zinc in Antiviral Immunity. Adv. Nutr. 2019, 10, 696–710. [Google Scholar] [CrossRef] [PubMed]
- Hulisz, D. Efficacy of zinc against common cold viruses: An overview. J. Am. Pharm. Assoc. 2004, 44, 594–603. [Google Scholar] [CrossRef] [PubMed]
- Kurugöl, Z.; Akilli, M.; Bayram, N.; Koturoglu, G. The prophylactic and therapeutic effectiveness of zinc sulphate on common cold in children. Acta Paediatr. 2006, 95, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Hadrup, N.; Ravn-Haren, G. Acute human toxicity and mortality after selenium ingestion: A review. J. Trace Elem. Med. Biol. 2020, 58, 126435. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Al-Quraishy, S.; Dkhil, M.A.; Wunderlich, F.; Sies, H. Dietary selenium in adjuvant therapy of viral and bacterial infections. Adv. Nutr. 2015, 6, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Erol, S.A.; Polat, N.; Akdas, S.; Aribal Ayral, P.; Anuk, A.T.; Ozden Tokalioglu, E.; Goncu Ayhan, Ş.; Kesikli, B.; Ceylan, M.N.; Tanacan, A.; et al. Maternal selenium status plays a crucial role on clinical outcomes of pregnant women with COVID-19 infection. J. Med. Virol. 2021, 93, 5438–5445. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Taylor, E.W.; Bennett, K.; Saad, R.; Rayman, M.P. Association between regional selenium status and reported outcome of COVID-19 cases in China. Am. J. Clin. Nutr. 2020, 111, 1297–1299. [Google Scholar] [CrossRef] [PubMed]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [PubMed]
- Raha, S.; Mallick, R.; Basak, S.; Duttaroy, A.K. Is copper beneficial for COVID-19 patients? Med. Hypotheses 2020, 142, 109814. [Google Scholar] [CrossRef] [PubMed]
- Warnes, S.L.; Little, Z.R.; Keevil, C.W. Human Coronavirus 229E Remains Infectious on Common Touch Surface Materials. mBio 2015, 6, e01697-15. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.F.; Ding, H.; Jiao, R.Q.; Wu, X.X.; Kong, L.D. Possibility of magnesium supplementation for supportive treatment in patients with COVID-19. Eur. J. Pharmacol. 2020, 886, 173546. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J.; O’Keefe, J.H. Magnesium and Vitamin D Deficiency as a Potential Cause of Immune Dysfunction, Cytokine Storm and Disseminated Intravascular Coagulation in covid-19 patients. Mo. Med. 2021, 118, 68–73. [Google Scholar]
- Lal, C.S.; Kumar, S.; Ranjan, A.; Rabidas, V.N.; Verma, N.; Pandey, K.; Verma, R.B.; Das, S.; Singh, D.; Das, P. Comparative analysis of serum zinc, copper, magnesium, calcium and iron level in acute and chronic patients of visceral leishmaniasis. J. Trace Elem. Med. Biol. 2013, 27, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Nasser, R.; Naffaa, M.E.; Mashiach, T.; Azzam, Z.S.; Braun, E. The association between serum magnesium levels and community-acquired pneumonia 30-day mortality. BMC Infect. Dis. 2018, 18, 698. [Google Scholar] [CrossRef]
- D’Souza, R.; Ashraf, R.; Rowe, H.; Zipursky, J.; Clarfield, L.; Maxwell, C.; Arzola, C.; Lapinsky, S.; Paquette, K.; Murthy, S.; et al. Pregnancy and COVID-19: Pharmacologic considerations. Ultrasound Obstet. Gynecol. 2021, 57, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Leermakers, E.T.; Moreira, E.M.; Kiefte-de Jong, J.C.; Darweesh, S.K.; Visser, T.; Voortman, T.; Bautista, P.K.; Chowdhury, R.; Gorman, D.; Bramer, W.M.; et al. Effects of choline on health across the life course: A systematic review. Nutr. Rev. 2015, 73, 500–522. [Google Scholar] [CrossRef] [PubMed]
- Shim, E.; Park, E. Choline intake and its dietary reference values in Korea and other countries: A review. Nutr. Res. Pract. 2022, 16 (Suppl. 1), S126–S133. [Google Scholar] [CrossRef] [PubMed]
- Freedman, R.; Hunter, S.K.; Law, A.J.; Wagner, B.D.; D’Alessandro, A.; Christians, U.; Noonan, K.; Wyrwa, A.; Hoffman, M.C. Higher Gestational Choline Levels in Maternal Infection Are Protective for Infant Brain Development. J. Pediatr. 2019, 208, 198–206.e2, Erratum in: J. Pediatr. 2019, 213, 254. [Google Scholar] [CrossRef]
- Vasistha, N.A.; Pardo-Navarro, M.; Gasthaus, J.; Weijers, D.; Müller, M.K.; García-González, D.; Malwade, S.; Korshunova, I.; Pfisterer, U.; von Engelhardt, J.; et al. Maternal inflammation has a profound effect on cortical interneuron development in a stage and subtype-specific manner. Mol. Psychiatry 2020, 25, 2313–2329. [Google Scholar] [CrossRef] [PubMed]
- Rossi, A.; Pollice, R.; Daneluzzo, E.; Marinangeli, M.G.; Stratta, P. Behavioral neurodevelopment abnormalities and schizophrenic disorder: A retrospective evaluation with the Childhood Behavior Checklist (CBCL). Schizophr. Res. 2000, 44, 121–128. [Google Scholar] [CrossRef]
- Zhang, M.; Han, X.; Bao, J.; Yang, J.; Shi, S.Q.; Garfield, R.E.; Liu, H. Choline Supplementation During Pregnancy Protects Against Gestational Lipopolysaccharide-Induced Inflammatory Responses. Reprod. Sci. 2018, 25, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Ross, R.G.; Hunter, S.K.; Hoffman, M.C.; McCarthy, L.; Chambers, B.M.; Law, A.J.; Leonard, S.; Zerbe, G.O.; Freedman, R. Perinatal Phosphatidylcholine Supplementation and Early Childhood Behavior Problems: Evidence for CHRNA7 Moderation. Am. J. Psychiatry 2016, 173, 509–516, Erratum in: Am. J. Psychiatry 2016, 173, 735. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, S.W.; Carter, R.C.; Molteno, C.D.; Stanton, M.E.; Herbert, J.S.; Lindinger, N.M.; Lewis, C.E.; Dodge, N.C.; Hoyme, H.E.; Zeisel, S.H.; et al. Efficacy of Maternal Choline Supplementation during Pregnancy in Mitigating Adverse Effects of Prenatal Alcohol Exposure on Growth and Cognitive Function: A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Alcohol. Clin. Exp. Res. 2018, 42, 1327–1341. [Google Scholar] [CrossRef] [PubMed]
- Caudill, M.A.; Strupp, B.J.; Muscalu, L.; Nevins, J.E.H.; Canfield, R.L. Maternal choline supplementation during the third trimester of pregnancy improves infant information processing speed: A randomized, double-blind, controlled feeding study. FASEB J. 2018, 32, 2172–2180. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.C.; Freedman, R.; Law, A.J.; Clark, A.M.; Hunter, S.K. Maternal nutrients and effects of gestational COVID-19 infection on fetal brain development. Clin. Nutr. ESPEN 2021, 43, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Coronavirus Disease 2019 (COVID-19) and Pregnancy. 2020. Available online: https://www.cdc.gov/coronavirus/2019-ncov/cases-updates/special-populations/pregnancy-data-on-covid-19/what-cdc-is-doing.html (accessed on 7 July 2022).
- Li, Y.; Freedman, R. Prospects for improving future mental health of children through prenatal maternal micronutrient supplementation in China. Pediatr. Investig. 2020, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Hung, H.C.; Ke, Y.Y.; Huang, S.Y.; Huang, P.N.; Kung, Y.A.; Chang, T.Y.; Yen, K.J.; Peng, T.T.; Chang, S.E.; Huang, C.T.; et al. Discovery of M Protease Inhibitors Encoded by SARS-CoV-2. Antimicrob. Agents Chemother. 2020, 64, e00872-20. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.F.; Zhang, L.H.; Bai, F.; Wang, N.P.; Garner, R.E.; McKallip, R.J.; Zhao, Z.Q. Attenuation of myocardial fibrosis with curcumin is mediated by modulating expression of angiotensin II AT1/AT2 receptors and ACE2 in rats. Drug Des. Devel. Ther. 2015, 9, 6043–6054. [Google Scholar] [CrossRef]
- Tőzsér, J.; Benkő, S. Natural Compounds as Regulators of NLRP3 Inflammasome-Mediated IL-1β Production. Mediat. Inflamm. 2016, 2016, 5460302. [Google Scholar] [CrossRef]
- Ullah, F.; Liang, H.; Niedermayer, G.; Münch, G.; Gyengesi, E. Evaluation of Phytosomal Curcumin as an Anti-inflammatory Agent for Chronic Glial Activation in the GFAP-IL6 Mouse Model. Front. Neurosci. 2020, 14, 170. [Google Scholar] [CrossRef]
- Pastor, N.; Collado, M.C.; Manzoni, P. Phytonutrient and Nutraceutical Action against COVID-19: Current Review of Characteristics and Benefits. Nutrients 2021, 13, 464. [Google Scholar] [CrossRef]
- Ling, L.J.; Lu, Y.; Zhang, Y.Y.; Zhu, H.Y.; Tu, P.; Li, H.; Chen, D.F. Flavonoids from Houttuynia cordata attenuate H1N1-induced acute lung injury in mice via inhibition of influenza virus and Toll-like receptor signalling. Phytomedicine 2020, 67, 153150. [Google Scholar] [CrossRef]
- Mlcek, J.; Jurikova, T.; Skrovankova, S.; Sochor, J. Quercetin and Its Anti-Allergic Immune Response. Molecules 2016, 21, 623. [Google Scholar] [CrossRef]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef] [PubMed]
- Omotuyi, I.O.; Nash, O.; Ajiboye, B.O.; Olumekun, V.O.; Oyinloye, B.E.; Osuntokun, O.T.; Olonisakin, A.; Ajayi, A.O.; Olusanya, O.; Akomolafe, F.S.; et al. Aframomum melegueta secondary metabolites exhibit polypharmacology against SARS-CoV-2 drug targets: In vitro validation of furin inhibition. Phytother. Res. 2021, 35, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yi, D.; Ji, C.; Wu, T.; Wang, M.; Guo, S.; Wang, L.; Zhao, D.; Hou, Y. Monolaurin Confers a Protective Effect Against Porcine Epidemic Diarrhea Virus Infection in Piglets by Regulating the Interferon Pathway. Front. Immunol. 2022, 12, 797476. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Nutrients | Sources | RDA in Pregnancy | Action and Properties | References |
|---|---|---|---|---|
| Amino Acids | 71 g/day |
| [53] | |
| Soy protein, peanuts, walnuts, and fish. | 10 g/day |
| [54,55,56,57,58] |
| n-3 PUFA | Fish, fish oil, algae oil, walnuts, edible seeds, and flaxseeds | 200–300 mg/day |
| [59,62,63] |
| Fish, fish oil, and algae oil |
| [61] | |
| Fish, fish oil, and algae oil |
| [60] | |
| Fatty Acids (S/MCFA) | ||||
| coconut oil | 1–5 mg/day |
| [66,67] |
| Vitamin A (all-trans-retinol) | milk, fish, eggs, carrots, leafy greens, broccoli, cantaloupe, and squash. | 770 mcg retinol equivalents (avoid overdoses) |
| [68,69,70] |
| Vitamin E (Tocopherol) | olive and sunflower oils (α-tocopherol), soybean and corn oil (γ-tocopherol) | 15 mg/day (avoid overdoses) |
| [49,76,77] |
| Vitamin B1 (thiamine) | whole grains, legumes, meats, and fish | 1.4 mg/day |
| [78,79,80] |
| Vitamin B6 (pyridoxine) | fruit, vegetables, and grain | 1.7 mg/day |
| [69] |
| Vitamin B9 (folic acid) | beef liver, leafy vegetables, peas, beans, avocados, eggs, and milk | -low-risk pregnancies: 0.4 mg/day for 12 weeks -high-risk pregnancies: 4 mg/day in high-risk pregnancy |
| [81,82,83,84,85,86,87] |
| Vitamin B12 (cobalamin) | meat, clams, liver, fish, poultry, eggs, and dairy products | 2.6 µg/day |
| [69,88,89,90] |
| Vitamin C (ascorbic acid) | vegetables and fruits: citrus fruits, strawberries, potatoes, and tomatoes | 1.5 g/kg body weight |
| [73,91,92,93,94,95,96,97,98,99] |
| Iron | red meat, oysters, lentils, beans, poultry, fish, leaf vegetables, watercress, tofu, chickpeas, black-eyed peas, and blackstrap molasses. | 27 mg/day |
| [69,100,101,102] |
| Zinc | meat, fish, shellfish, fowl, eggs, dairy, wheat (germ and bran), sesame, poppy, alfalfa, celery, mustard, beans, nuts, almonds, whole grains, pumpkin seeds, sunflower seeds, and blackcurrant | 11 mg/day |
| [49,69,73,105,106,107,108,109,110] |
| Selenium | meat, nuts (Brazil nuts), cereals, and mushrooms | 60 µg/day |
| [111,112,113,114,115,116] |
| Copper | oysters, beef and lamb liver, Brazil nuts, blackstrap molasses, cocoa, black pepper, lobster, nuts and sunflower seeds, green olives, avocados, and wheat bran | 1000 μg/day |
| [117,118] |
| Magnesium | Spices, nuts, cereals, cocoa, green leafy vegetables such as spinach, coffee, and tea. | 360 mg/day |
| [119,120,121,122,123] |
| Choline | organ meats, egg yolks, grains, vegetables, fruit, and dairy products | 450–550 mg/day |
| [124,125,126,127,128,129,130,131,132,133,134,135] |
| Polyphenols | green tea, broccoli, and apples |
| ||
| Curcuma longa species plants | 3 mg/kg body weight/day |
| [136,137,138,139,140] |
| Flavonoids | red wine, oranges, red fruits and vegetables |
| [141] | |
| green tea | 120 mg/day |
| [63] |
| spinach, cabbage, kale and broccoli | 5–8 mg/day |
| [140] |
| onion, grapes, shallots, tea, Ginko bilboa and tomatoes | 500–1000 mg/day |
| [142,143,144] |
| parsley, celery, onions and oranges | 50 mg/day |
| [145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Germano, C.; Messina, A.; Tavella, E.; Vitale, R.; Avellis, V.; Barboni, M.; Attini, R.; Revelli, A.; Zola, P.; Manzoni, P.; et al. Fetal Brain Damage during Maternal COVID-19: Emerging Hypothesis, Mechanism, and Possible Mitigation through Maternal-Targeted Nutritional Supplementation. Nutrients 2022, 14, 3303. https://doi.org/10.3390/nu14163303
Germano C, Messina A, Tavella E, Vitale R, Avellis V, Barboni M, Attini R, Revelli A, Zola P, Manzoni P, et al. Fetal Brain Damage during Maternal COVID-19: Emerging Hypothesis, Mechanism, and Possible Mitigation through Maternal-Targeted Nutritional Supplementation. Nutrients. 2022; 14(16):3303. https://doi.org/10.3390/nu14163303
Chicago/Turabian StyleGermano, Chiara, Alessandro Messina, Elena Tavella, Raffaele Vitale, Vincenzo Avellis, Martina Barboni, Rossella Attini, Alberto Revelli, Paolo Zola, Paolo Manzoni, and et al. 2022. "Fetal Brain Damage during Maternal COVID-19: Emerging Hypothesis, Mechanism, and Possible Mitigation through Maternal-Targeted Nutritional Supplementation" Nutrients 14, no. 16: 3303. https://doi.org/10.3390/nu14163303
APA StyleGermano, C., Messina, A., Tavella, E., Vitale, R., Avellis, V., Barboni, M., Attini, R., Revelli, A., Zola, P., Manzoni, P., & Masturzo, B. (2022). Fetal Brain Damage during Maternal COVID-19: Emerging Hypothesis, Mechanism, and Possible Mitigation through Maternal-Targeted Nutritional Supplementation. Nutrients, 14(16), 3303. https://doi.org/10.3390/nu14163303