
Relevance of ω-6 GLA Added to ω-3 PUFAs Supplements for ADHD: A Narrative Review




{kind=link}
Abstract
:1. Introduction
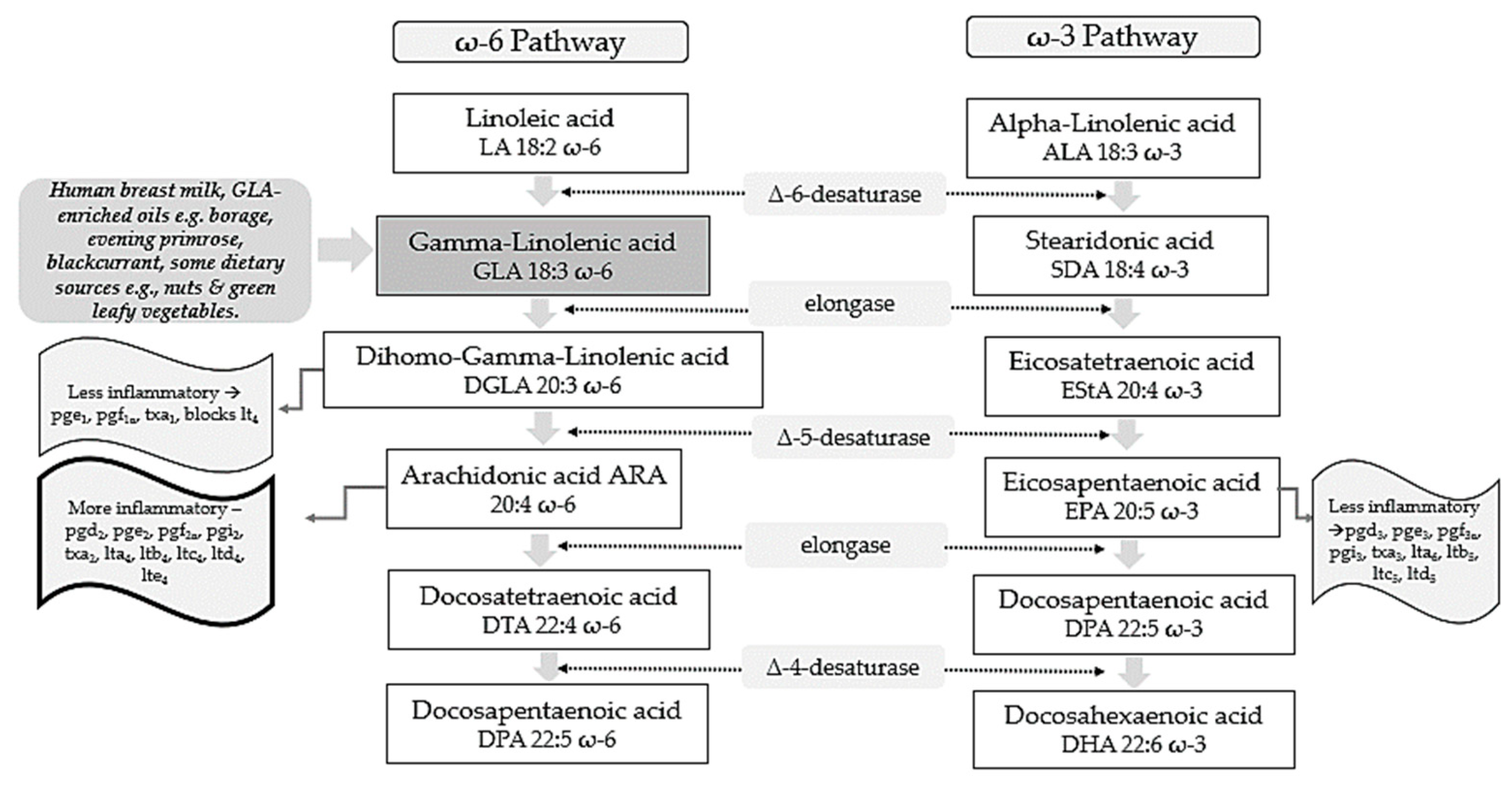
2. GLA as an Antioxidant and Anti-Inflammatory Molecule
3. GLA Synergy with EPA
4. GLA and ADHD Focus
4.1. Pathogenesis and Role of Neuroinflammation
4.2. Combined ω-6 and ω-3 PUFA Supplementation
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tesei, A.; Crippa, A.; Ceccarelli, S.B.; Mauri, M.; Molteni, M.; Agostoni, C.; Nobile, M. The potential relevance of docosahexaenoic acid and eicosapentaenoic acid to the etiopathogenesis of childhood neuropsychiatric disorders. Eur. Child Adolesc. Psychiatry 2017, 26, 1011–1030. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, M.G. The essentials of essential fatty acids. J. Diet Suppl. 2009, 6, 143–161. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Simopoulos, A.P. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 2002, 56, 365–379. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Importance of the ratio of omega-6/omega-3 essential fatty acids: Evolutionary aspects. World Rev. Nutr. Diet 2003, 92, 1–22. [Google Scholar] [CrossRef]
- Holub, B. Clinical nutrition: 4. Omega-3 fatty acids in cardiovascular care. JAMC 2002, 166, 608–615. [Google Scholar]
- Ander, B.; Dupasquier, C.; Prociuk, M.; Pierce, G. Polyunsaturated fatty acids and their effects on cardiovascular disease. Exp. Clin. Cardiol. 2003, 8, 164–172. [Google Scholar]
- Morris-Rosendahl, D.J.; Crocq, M.A. Neurodevelopmental disorders-the history and future of a diagnostic concept. Dialogues Clin. NeuroSci. 2020, 22, 65–72. [Google Scholar] [CrossRef]
- DSM. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Washington, DC, USA, 2013. [Google Scholar]
- Ahmed, R.; Borst, J.M.; Yong, C.W.; Aslani, P. Do parents of children with attention-deficit/hyperactivity disorder (ADHD) receive adequate information about the disorder and its treatments? A qualitative investigation. Patient Prefer. Adherence 2014, 8, 661–670. [Google Scholar] [CrossRef] [Green Version]
- Thomas, R.; Sanders, S.; Doust, J.; Beller, E.; Glasziou, P. Prevalence of attention-deficit/hyperactivity disorder: A systematic review and meta-analysis. Pediatrics 2015, 135, e994–e1001. [Google Scholar] [CrossRef] [Green Version]
- Sayal, K.; Prasad, V.; Daley, D.; Ford, T.; Coghill, D. ADHD in children and young people: Prevalence, care pathways, and service provision. Lancet Psychiatry 2018, 5, 175–186. [Google Scholar] [CrossRef]
- Mohammadi, M.R.; Zarafshan, H.; Khaleghi, A.; Ahmadi, N.; Hooshyari, Z.; Mostafavi, S.A.; Ahmadi, A.; Alavi, S.S.; Shakiba, A.; Salmanian, M. Prevalence of ADHD and Its Comorbidities in a Population-Based Sample. J. Atten. Disord. 2021, 25, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, K.W. Special issues in the diagnosis and treatment of ADHD in adolescents. Postgrad. Med. 2008, 120, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Shaw, M.; Hodgkins, P.; Caci, H.; Young, S.; Kahle, J.; Woods, A.G.; Arnold, L.E. A systematic review and analysis of long-term outcomes in attention deficit hyperactivity disorder: Effects of treatment and non-treatment. BMC Med. 2012, 10, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, J.P.; Su, K.P.; Mondelli, V.; Pariante, C.M. Omega-3 Polyunsaturated Fatty Acids in Youths with Attention Deficit Hyperactivity Disorder: A Systematic Review and Meta-Analysis of Clinical Trials and Biological Studies. Neuropsychopharmacology 2018, 43, 534–545. [Google Scholar] [CrossRef] [Green Version]
- Bonvicini, C.; Faraone, S.V.; Scassellati, C. Attention-deficit hyperactivity disorder in adults: A systematic review and meta-analysis of genetic, pharmacogenetic and biochemical studies. Mol. Psychiatry 2016, 21, 872–884. [Google Scholar] [CrossRef]
- Hansen, T.M.; Lerche, A.; Kassis, V.; Lorenzen, I.; Sondergaard, J. Treatment of rheumatoid arthritis with prostaglandin E1 precursors cis-linoleic acid and gamma-linolenic acid. Scand J. Rheumatol. 1983, 12, 85–88. [Google Scholar] [CrossRef]
- Horrobin, D.F. The role of essential fatty acids and prostaglandins in the premenstrual syndrome. J. Reprod. Med. 1983, 28, 465–468. [Google Scholar]
- Kunkel, S.L.; Ogawa, H.; Ward, P.A.; Zurier, R.B. Suppression of chronic inflammation by evening primrose oil. Prog. Lipid Res. 1981, 20, 885–888. [Google Scholar] [CrossRef]
- Innes, J.K.; Calder, P.C. Omega-6 fatty acids and inflammation. Prostaglandins Leukot. Essent. Fat. Acids 2018, 132, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.R.; Burger, B.; Kuhl, C.M.C.; Candreva, T.; Dos Anjos, M.B.P.; Rodrigues, H.G. Wound Healing and Omega-6 Fatty Acids: From Inflammation to Repair. Mediat. Inflamm. 2018, 2018, 2503950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, S.N.; Darsareh, F. Impact of evening primrose oil consumption on psychological symptoms of postmenopausal women: A randomized double-blinded placebo-controlled clinical trial. Menopause 2020, 27, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Majdinasab, N.; Namjoyan, F.; Taghizadeh, M.; Saki, H. The effect of evening primrose oil on fatigue and quality of life in patients with multiple sclerosis. Neuropsychiatr. Dis. Treat. 2018, 14, 1505–1512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeant, S.; Rahbar, E.; Chilton, F.H. Gamma-linolenic acid, Dihommo-gamma linolenic, Eicosanoids and Inflammatory Processes. Eur. J. Pharmacol. 2016, 785, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, R.; Huang, Y.S. Gamma linolenic acid: An antiinflammatory omega-6 fatty acid. Curr. Pharm. Biotechnol. 2006, 7, 531–534. [Google Scholar] [CrossRef] [Green Version]
- Dobryniewski, J.; Szajda, S.D.; Waszkiewicz, N.; Zwierz, K. Biology of essential fatty acids (EFA). Przegl. Lek. 2007, 64, 91–99. [Google Scholar]
- Williams, C.M.; Burdge, G. Long-chain n-3 PUFA: Plant v. marine sources. Proc. Nutr. Soc. 2006, 65, 42–50. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.A.; Ali, A.; Khan, S.A.; Zahran, S.A.; Damanhouri, G.; Azhar, E.; Qadri, I. Unraveling the complex relationship triad between lipids, obesity, and inflammation. Mediat. Inflamm. 2014, 2014, 502749. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.Y.; Chapkin, R.S. Importance of dietary gamma-linolenic acid in human health and nutrition. J. Nutr. 1998, 128, 1411–1414. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.M.; Swan, D.D.; Surette, M.E.; Stegner, J.; Chilton, T.; Fonteh, A.N.; Chilton, F.H. Dietary supplementation with gamma-linolenic acid alters fatty acid content and eicosanoid production in healthy humans. J. Nutr. 1997, 127, 1435–1444. [Google Scholar] [CrossRef]
- Chilton, L.; Surette, M.E.; Swan, D.D.; Fonteh, A.N.; Johnson, M.M.; Chilton, F.H. Metabolism of gammalinolenic acid in human neutrophils. J. Immunol. 1996, 156, 2941–2947. [Google Scholar]
- Du, Y.; Taylor, C.G.; Aukema, H.M.; Zahradka, P. Role of oxylipins generated from dietary PUFAs in the modulation of endothelial cell function. Prostaglandins Leukot. Essent. Fat. Acids 2020, 160, 102160. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.Y.; Ramos, K.S.; Chapkin, R.S. Dietary gamma-linolenic acid modulates macrophage-vascular smooth muscle cell interactions. Evidence for a macrophage-derived soluble factor that downregulates DNA synthesis in smooth muscle cells. Arter. Thromb. Vasc. Biol. 1995, 15, 1397–1403. [Google Scholar] [CrossRef]
- Heitmann, J.; Iversen, L.; Kragballe, K.; Ziboh, V.A. Incorporation of 15-hydroxyeicosatrienoic acid in specific phospholipids of cultured human keratinocytes and psoriatic plaques. Exp. Dermatol. 1995, 4, 74–78. [Google Scholar] [CrossRef]
- Gonzalez-Soto, M.; Mutch, D.M. Diet Regulation of Long-Chain PUFA Synthesis: Role of Macronutrients, Micronutrients, and Polyphenols on Delta-5/Delta-6 Desaturases and Elongases 2/5. Adv. Nutr. 2021, 12, 980–994. [Google Scholar] [CrossRef]
- Moghadas, M.; Essa, M.M.; Ba-Omar, T.; Al-Shehi, A.; Qoronfleh, M.W.; Eltayeb, E.A.; Guillemin, G.J.; Manivasagam, T.; Justin-Thenmozhi, A.; Al-Bulushi, B.S.; et al. Antioxidant therapies in attention deficit hyperactivity disorder. Front. BioSci. 2019, 24, 313–333. [Google Scholar] [CrossRef]
- Boyce, J.A. Eicosanoids in asthma, allergic inflammation, and host defense. Curr. Mol. Med. 2008, 8, 335–349. [Google Scholar] [CrossRef]
- Barham, J.B.; Edens, M.B.; Fonteh, A.N.; Johnson, M.M.; Easter, L.; Chilton, F.H. Addition of eicosapentaenoic acid to gamma-linolenic acid-supplemented diets prevents serum arachidonic acid accumulation in humans. J. Nutr. 2000, 130, 1925–1931. [Google Scholar] [CrossRef] [Green Version]
- Surette, M.E.; Stull, D.; Lindemann, J. The impact of a medical food containing gammalinolenic and eicosapentaenoic acids on asthma management and the quality of life of adult asthma patients. Curr. Med. Res. Opin. 2008, 24, 559–567. [Google Scholar] [CrossRef]
- Surette, M.E.; Koumenis, I.L.; Edens, M.B.; Tramposch, K.M.; Chilton, F.H. Inhibition of leukotriene synthesis, pharmacokinetics, and tolerability of a novel dietary fatty acid formulation in healthy adult subjects. Clin. Ther. 2003, 25, 948–971. [Google Scholar] [CrossRef]
- Weaver, K.L.; Ivester, P.; Seeds, M.; Case, L.D.; Arm, J.P.; Chilton, F.H. Effect of dietary fatty acids on inflammatory gene expression in healthy humans. J. Biol. Chem. 2009, 284, 15400–15407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.C.; Ivester, P.; Hester, A.G.; Sergeant, S.; Case, L.D.; Morgan, T.; Kouba, E.O.; Chilton, F.H. The impact of polyunsaturated fatty acid-based dietary supplements on disease biomarkers in a metabolic syndrome/diabetes population. Lipids Health Dis. 2014, 13, 196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pontes-Arruda, A.; Aragao, A.M.; Albuquerque, J.D. Effects of enteral feeding with eicosapentaenoic acid, gamma-linolenic acid, and antioxidants in mechanically ventilated patients with severe sepsis and septic shock. Crit. Care Med. 2006, 34, 2325–2333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Bo, L.; Liu, W.; Lu, X.; Jin, F. Enteral Immunomodulatory Diet (Omega-3 Fatty Acid, gamma-Linolenic Acid and Antioxidant Supplementation) for Acute Lung Injury and Acute Respiratory Distress Syndrome: An Updated Systematic Review and Meta-Analysis. Nutrients 2015, 7, 5572–5585. [Google Scholar] [CrossRef]
- Dawczynski, C.; Hackermeier, U.; Viehweger, M.; Stange, R.; Springer, M.; Jahreis, G. Incorporation of n-3 PUFA and gamma-linolenic acid in blood lipids and red blood cell lipids together with their influence on disease activity in patients with chronic inflammatory arthritis--a randomized controlled human intervention trial. Lipids Health Dis. 2011, 10, 130. [Google Scholar] [CrossRef] [Green Version]
- Buss, C. Maternal oxidative stress during pregnancy and offspring neurodevelopment. Brain Behav. Immun. 2021, 93, 6–7. [Google Scholar] [CrossRef]
- Han, V.X.; Patel, S.; Jones, H.F.; Dale, R.C. Maternal immune activation and neuroinflammation in human neurodevelopmental disorders. Nat. Rev. Neurol. 2021, 17, 564–579. [Google Scholar] [CrossRef]
- Laye, S.; Nadjar, A.; Joffre, C.; Bazinet, R.P. Anti-Inflammatory Effects of Omega-3 Fatty Acids in the Brain: Physiological Mechanisms and Relevance to Pharmacology. Pharmacol. Rev. 2018, 70, 12–38. [Google Scholar] [CrossRef]
- Caramia, G. The essential fatty acids omega-6 and omega-3: From their discovery to their use in therapy. Minerva. Pediatr. 2008, 60, 219–233. [Google Scholar]
- Brikell, I.; Kuja-Halkola, R.; Larsson, H. Heritability of attention-deficit hyperactivity disorder in adults. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2015, 168, 406–413. [Google Scholar] [CrossRef]
- Werenberg Dreier, J.; Nybo Andersen, A.M.; Hvolby, A.; Garne, E.; Kragh Andersen, P.; Berg-Beckhoff, G. Fever and infections in pregnancy and risk of attention deficit/hyperactivity disorder in the offspring. J. Child Psychol. Psychiatry 2016, 57, 540–548. [Google Scholar] [CrossRef] [PubMed]
- Silva, D.; Colvin, L.; Hagemann, E.; Bower, C. Environmental risk factors by gender associated with attention-deficit/hyperactivity disorder. Pediatrics 2014, 133, e14–e22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez, C.E.; Barry, C.; Sabhlok, A.; Russell, K.; Majors, A.; Kollins, S.H.; Fuemmeler, B.F. Maternal pre-pregnancy obesity and child neurodevelopmental outcomes: A meta-analysis. Obes. Rev. 2018, 19, 464–484. [Google Scholar] [CrossRef] [PubMed]
- Thapar, A.; Cooper, M.; Eyre, O.; Langley, K. What have we learnt about the causes of ADHD? J. Child Psychol. Psychiatry 2013, 54, 3–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vrijheid, M.; Casas, M.; Gascon, M.; Valvi, D.; Nieuwenhuijsen, M. Environmental pollutants and child health-A review of recent concerns. Int. J. Hyg. Environ. Health 2016, 219, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, F.X.; Lee, P.P.; Sharp, W.; Jeffries, N.O.; Greenstein, D.K.; Clasen, L.S.; Blumenthal, J.D.; James, R.S.; Ebens, C.L.; Walter, J.M.; et al. Developmental trajectories of brain volume abnormalities in children and adolescents with attention-deficit/hyperactivity disorder. JAMA 2002, 288, 1740–1748. [Google Scholar] [CrossRef]
- Reus, G.Z.; Fries, G.R.; Stertz, L.; Badawy, M.; Passos, I.C.; Barichello, T.; Kapczinski, F.; Quevedo, J. The role of inflammation and microglial activation in the pathophysiology of psychiatric disorders. Neuroscience 2015, 300, 141–154. [Google Scholar] [CrossRef]
- Hassan, W.; Noreen, H.; Castro-Gomes, V.; Mohammadzai, I.; da Rocha, J.B.; Landeira-Fernandez, J. Association of Oxidative Stress with Psychiatric Disorders. Curr. Pharm. Des. 2016, 22, 2960–2974. [Google Scholar] [CrossRef]
- Belmadani, A.; Tran, P.B.; Ren, D.; Miller, R.J. Chemokines regulate the migration of neural progenitors to sites of neuroinflammation. J. NeuroSci. 2006, 26, 3182–3191. [Google Scholar] [CrossRef]
- Sen, S.; Duman, R.; Sanacora, G. Serum brain-derived neurotrophic factor, depression, and antidepressant medications: Meta-analyses and implications. Biol. Psychiatry 2008, 64, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Kronfol, Z.; Remick, D.G. Cytokines and the brain: Implications for clinical psychiatry. Am. J. Psychiatry 2000, 157, 683–694. [Google Scholar] [CrossRef] [PubMed]
- Zayats, T.; Athanasiu, L.; Sonderby, I.; Djurovic, S.; Westlye, L.T.; Tamnes, C.K.; Fladby, T.; Aase, H.; Zeiner, P.; Reichborn-Kjennerud, T.; et al. Genome-wide analysis of attention deficit hyperactivity disorder in Norway. PLoS ONE 2015, 10, e0122501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.T.; Chen, Y.C.; Gau, S.S.; Yeh, T.H.; Fan, H.Y.; Hwang, Y.Y.; Lee, Y.L. Associations between allergic diseases and attention deficit hyperactivity/oppositional defiant disorders in children. Pediatr. Res. 2016, 80, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Instanes, J.T.; Halmoy, A.; Engeland, A.; Haavik, J.; Furu, K.; Klungsoyr, K. Attention-Deficit/Hyperactivity Disorder in Offspring of Mothers With Inflammatory and Immune System Diseases. Biol. Psychiatry 2017, 81, 452–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toto, M.; Margari, F.; Simone, M.; Craig, F.; Petruzzelli, M.G.; Tafuri, S.; Margari, L. Antibasal Ganglia Antibodies and Antistreptolysin O in Noncomorbid ADHD. J. Atten. Disord. 2015, 19, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Giana, G.; Romano, E.; Porfirio, M.C.; D’Ambrosio, R.; Giovinazzo, S.; Troianiello, M.; Barlocci, E.; Travaglini, D.; Granstrem, O.; Pascale, E.; et al. Detection of auto-antibodies to DAT in the serum: Interactions with DAT genotype and psycho-stimulant therapy for ADHD. J. Neuroimmunol. 2015, 278, 212–222. [Google Scholar] [CrossRef]
- Mittleman, B.B.; Castellanos, F.X.; Jacobsen, L.K.; Rapoport, J.L.; Swedo, S.E.; Shearer, G.M. Cerebrospinal fluid cytokines in pediatric neuropsychiatric disease. J. Immunol. 1997, 159, 2994–2999. [Google Scholar]
- Wohleb, E.S.; McKim, D.B.; Sheridan, J.F.; Godbout, J.P. Monocyte trafficking to the brain with stress and inflammation: A novel axis of immune-to-brain communication that influences mood and behavior. Front. NeuroSci. 2014, 8, 447. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.J.; Fowler, J.S.; Ding, Y.S. Imaging the effects of methylphenidate on brain dopamine: New model on its therapeutic actions for attention-deficit/hyperactivity disorder. Biol. Psychiatry 2005, 57, 1410–1415. [Google Scholar] [CrossRef]
- Arnold, L.E.; Hurt, E.; Lofthouse, N. Attention-deficit/hyperactivity disorder: Dietary and nutritional treatments. Child Adolesc. Psychiatr. Clin. N. Am. 2013, 22, 381–402. [Google Scholar] [CrossRef]
- Dunn, G.A.; Nigg, J.T.; Sullivan, E.L. Neuroinflammation as a risk factor for attention deficit hyperactivity disorder. Pharmacol. Biochem. Behav. 2019, 182, 22–34. [Google Scholar] [CrossRef]
- Spivak, B.; Vered, Y.; Yoran-Hegesh, R.; Averbuch, E.; Mester, R.; Graf, E.; Weizman, A. Circulatory levels of catecholamines, serotonin and lipids in attention deficit hyperactivity disorder. Acta Psychiatr. Scand. 1999, 99, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Quist, J.F.; Kennedy, J.L. Genetics of childhood disorders: XXIII. ADHD, Part 7: The serotonin system. J. Am. Acad. Child Adolesc. Psychiatry 2001, 40, 253–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, P.; Caballero, J.; Omidian, H. Use of serotonin norepinephrine reuptake inhibitors in the treatment of attention-deficit hyperactivity disorder in pediatrics. Ann. Pharmacother. 2014, 48, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Schuchardt, J.P.; Huss, M.; Stauss-Grabo, M.; Hahn, A. Significance of long-chain polyunsaturated fatty acids (PUFAs) for the development and behaviour of children. Eur. J. Pediatr. 2010, 169, 149–164. [Google Scholar] [CrossRef]
- Brikell, I.; Burton, C.; Mota, N.R.; Martin, J. Insights into attention-deficit/hyperactivity disorder from recent genetic studies. Psychol. Med. 2021, 51, 2274–2286. [Google Scholar] [CrossRef]
- Carlsson, T.; Molander, F.; Taylor, M.J.; Jonsson, U.; Bolte, S. Early environmental risk factors for neurodevelopmental disorders—a systematic review of twin and sibling studies. Dev. Psychopathol. 2021, 33, 1448–1495. [Google Scholar] [CrossRef]
- Lange, K.W. Dietary factors in the etiology and therapy of attention deficit/hyperactivity disorder. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 464–469. [Google Scholar] [CrossRef]
- Konikowska, K.; Regulska-Ilow, B.; Rozanska, D. The influence of components of diet on the symptoms of ADHD in children. Rocz. Panstw. Zakl. Hig. 2012, 63, 127–134. [Google Scholar]
- Konigs, A.; Kiliaan, A.J. Critical appraisal of omega-3 fatty acids in attention-deficit/hyperactivity disorder treatment. Neuropsychiatr. Dis. Treat. 2016, 12, 1869–1882. [Google Scholar] [CrossRef] [Green Version]
- Chang, C.Y.; Ke, D.S.; Chen, J.Y. Essential fatty acids and human brain. Acta Neurol. Taiwan 2009, 18, 231–241. [Google Scholar] [PubMed]
- Carver, J.D.; Benford, V.J.; Han, B.; Cantor, A.B. The relationship between age and the fatty acid composition of cerebral cortex and erythrocytes in human subjects. Brain Res. Bull. 2001, 56, 79–85. [Google Scholar] [CrossRef]
- Derbyshire, E. Do Omega-3/6 Fatty Acids Have a Therapeutic Role in Children and Young People with ADHD? J. Lipids 2017, 2017, 6285218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillies, D.; Sinn, J.; Lad, S.S.; Leach, M.J.; Ross, M.J. Polyunsaturated fatty acids (PUFA) for attention deficit hyperactivity disorder (ADHD) in children and adolescents. Cochrane Database Syst. Rev. 2012, 7, CD007986. [Google Scholar] [CrossRef] [PubMed]
- Puri, B.K.; Martins, J.G. Which polyunsaturated fatty acids are active in children with attention-deficit hyperactivity disorder receiving PUFA supplementation? A fatty acid validated meta-regression analysis of randomized controlled trials. Prostaglandins Leukot. Essent. Fat. Acids 2014, 90, 179–189. [Google Scholar] [CrossRef]
- Barragan, E.; Breuer, D.; Dopfner, M. Efficacy and Safety of Omega-3/6 Fatty Acids, Methylphenidate, and a Combined Treatment in Children With ADHD. J. Atten. Disord. 2017, 21, 433–441. [Google Scholar] [CrossRef]
- Matsudaira, T.; Gow, R.V.; Kelly, J.; Murphy, C.; Potts, L.; Sumich, A.; Ghebremeskel, K.; Crawford, M.A.; Taylor, E. Biochemical and Psychological Effects of Omega-3/6 Supplements in Male Adolescents with Attention-Deficit/Hyperactivity Disorder: A Randomized, Placebo-Controlled, Clinical Trial. J. Child Adolesc. Psychopharmacol. 2015, 25, 775–782. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.; Mansson, J.E.; Ostlund, S.; Fransson, G.; Areskoug, B.; Hjalmarsson, K.; Landgren, M.; Kadesjo, B.; Gillberg, C. Fatty acids in ADHD: Plasma profiles in a placebo-controlled study of Omega 3/6 fatty acids in children and adolescents. Atten. Defic. Hyperact. Disord. 2012, 4, 199–204. [Google Scholar] [CrossRef]
- Johnson, M.; Ostlund, S.; Fransson, G.; Kadesjo, B.; Gillberg, C. Omega-3/omega-6 fatty acids for attention deficit hyperactivity disorder: A randomized placebo-controlled trial in children and adolescents. J. Atten. Disord. 2009, 12, 394–401. [Google Scholar] [CrossRef] [Green Version]
- Dopfner, M.; Dose, C.; Breuer, D.; Heintz, S.; Schiffhauer, S.; Banaschewski, T. Efficacy of Omega-3/Omega-6 Fatty Acids in Preschool Children at Risk of ADHD: A Randomized Placebo-Controlled Trial. J. Atten. Disord. 2021, 25, 1096–1106. [Google Scholar] [CrossRef]
- Chang, J.P.; Su, K.P.; Mondelli, V.; Satyanarayanan, S.K.; Yang, H.T.; Chiang, Y.J.; Chen, H.T.; Pariante, C.M. High-dose eicosapentaenoic acid (EPA) improves attention and vigilance in children and adolescents with attention deficit hyperactivity disorder (ADHD) and low endogenous EPA levels. Transl. Psychiatry 2019, 9, 303. [Google Scholar] [CrossRef] [PubMed]
- Banaschewski, T.; Belsham, B.; Bloch, M.H.; Ferrin, M.; Johnson, M.; Kustow, J.; Robinson, S.; Zuddas, A. Supplementation with polyunsaturated fatty acids (PUFAs) in the management of attention deficit hyperactivity disorder (ADHD). Nutr. Health 2018, 24, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Derbyshire, E. Oily Fish and Omega-3 s Across the Life Stages: A Focus on Intakes and Future Directions. Front Nutr. 2019, 6, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchioni, D.M.; de Oliveira, M.F.; Carioca, A.A.F.; Miranda, A.A.M.; Carvalho, A.M.; Oki, E.; Norde, M.M.; Rogero, M.M.; Damasceno, N.R.T.; Fisberg, R.M. Plasma fatty acids: Biomarkers of dietary intake? Nutrition 2019, 59, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. “Cell Membrane Theory of Senescence” and the Role of Bioactive Lipids in Aging, and Aging Associated Diseases and Their Therapeutic Implications. Biomolecules 2021, 11, 241. [Google Scholar] [CrossRef] [PubMed]
- LaChance, L.; McKenzie, K.; Taylor, V.H.; Vigod, S.N. Omega-6 to Omega-3 Fatty Acid Ratio in Patients with ADHD: A Meta-Analysis. J. Can. Acad. Child Adolesc. Psychiatry 2016, 25, 87–96. [Google Scholar]
- Hester, A.G.; Murphy, R.C.; Uhlson, C.J.; Ivester, P.; Lee, T.C.; Sergeant, S.; Miller, L.R.; Howard, T.D.; Mathias, R.A.; Chilton, F.H. Relationship between a common variant in the fatty acid desaturase (FADS) cluster and eicosanoid generation in humans. J. Biol. Chem. 2014, 289, 22482–22489. [Google Scholar] [CrossRef] [Green Version]
- Keim, S.A.; Boone, K.M.; Klebanoff, M.A.; Turner, A.N.; Rausch, J.; Nelin, M.A.; Rogers, L.K.; Yeates, K.O.; Nelin, L.; Sheppard, K.W. Effect of Docosahexaenoic Acid Supplementation vs Placebo on Developmental Outcomes of Toddlers Born Preterm: A Randomized Clinical Trial. JAMA Pediatr. 2018, 172, 1126–1134. [Google Scholar] [CrossRef] [Green Version]
- Voigt, R.G.; Llorente, A.M.; Jensen, C.L.; Fraley, J.K.; Berretta, M.C.; Heird, W.C. A randomized, double-blind, placebo-controlled trial of docosahexaenoic acid supplementation in children with attention-deficit/hyperactivity disorder. J. Pediatr. 2001, 139, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Saini, R.K.; Prasad, P.; Sreedhar, R.V.; Akhilender Naidu, K.; Shang, X.; Keum, Y.S. Omega-3 Polyunsaturated Fatty Acids (PUFAs): Emerging Plant and Microbial Sources, Oxidative Stability, Bioavailability, and Health Benefits-A Review. Antioxidants 2021, 10, 1627. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Helft, J.; Caccialanza, R.; Derbyshire, E.; Maes, M. Relevance of ω-6 GLA Added to ω-3 PUFAs Supplements for ADHD: A Narrative Review. Nutrients 2022, 14, 3273. https://doi.org/10.3390/nu14163273
D’Helft J, Caccialanza R, Derbyshire E, Maes M. Relevance of ω-6 GLA Added to ω-3 PUFAs Supplements for ADHD: A Narrative Review. Nutrients. 2022; 14(16):3273. https://doi.org/10.3390/nu14163273
Chicago/Turabian StyleD’Helft, Jelle, Riccardo Caccialanza, Emma Derbyshire, and Michael Maes. 2022. "Relevance of ω-6 GLA Added to ω-3 PUFAs Supplements for ADHD: A Narrative Review" Nutrients 14, no. 16: 3273. https://doi.org/10.3390/nu14163273
APA StyleD’Helft, J., Caccialanza, R., Derbyshire, E., & Maes, M. (2022). Relevance of ω-6 GLA Added to ω-3 PUFAs Supplements for ADHD: A Narrative Review. Nutrients, 14(16), 3273. https://doi.org/10.3390/nu14163273